Introduction
Cellular junctions comprise a range of cell adhesion
molecules (CAM) and are essential for maintaining tissue
architecture (1). The four major CAM
families are integrins, cadherins, selectins and the immunoglobulin
CAM superfamily (2). Integrins are
composed of two or more noncovalently-associated membrane-spanning
subunits α and β (3). The specific
combination of α and β subunits confers specificity for various
extracellular ligands and their respective intracellular signaling
events, and each combination of α and β represents a significant
receptor family within the context of interaction with the
extracellular matrix (3). Cadherins
are calcium-dependent glycoproteins, which possess an extracellular
CAM domain with three to five internal repeats, a single-span
transmembrane domain and an intracellular domain (2). The extracellular domain of selectins
consists of a calcium-dependent lectin domain and an epidermal
growth factor (EGF)-like domain (2).
Selectins also contain a hydrophobic transmembrane domain and a
short cytoplasmic tail (2). The
Ig-CAMs are calcium-independent, with an extracellular domain
comprising a ligand-binding region of four to six Ig-like repeats,
one to five fibronectin-like repeats, a transmembrane domain and an
intracellular component (1).
Although these families are predominant, a number of CAMs do not
share any structural similarities with them, such as the epithelial
cell adhesion molecule (EpCAM) (4).
EpCAM is one of the first identified human
tumor-associated biomarkers (5) and
is now considered to be a marker of tumor-initiating cells
(6). EpCAM is a transmembrane,
calcium-independent, homophilic, intercellular adhesion
glycoprotein (314 amino acids; 40 kDa) with three distinct domains:
An extracellular domain (EpEX, 265 amino acids), a transmembrane
domain and an intracellular domain (EpICD, 26 amino acids)
(7). The cleaved EpICD enters the
nucleus, leading to the activation of the β-catenin/c-Myc signaling
pathway to promote cancer cell proliferation (8). EpCAM functions include cell signaling,
differentiation and migration in addition to adhesion and
proliferation (4). EpCAM has been
implicated in carcinogenesis and is expressed robustly in various
types of human epithelial cancers, such as lung, breast, ovarian,
cervical and colorectal cancer (CRC), suggesting that it may be a
promising target for cancer diagnosis and therapy (9–11).
According to GLOBOCAN 2018 data, CRC is the third
most commonly occurring cancer and second leading cause of
cancer-associated death in the world, with ~881,000 deaths
estimated for 2018 (12). Although
surgical removal of cancer followed by adjuvant therapy is one of
the most effective treatments, recurrences are inevitable (13–15).
Antibody-based treatments are also currently used in patients with
advanced CRC; however, the prognosis and clinical outcomes of
patients with CRC remain poor (16).
Therefore, new strategies are required to improve the effectiveness
of CRC treatment.
The present study developed an anti-EpCAM monoclonal
antibody (mAb) using cell-based immunization and screening (CBIS)
methods (17) aiming to determine
whether these anti-EpCAM mAbs induced antibody-dependent cellular
cytotoxicity (ADCC), complement-dependent cytotoxicity (CDC) or
antitumor activity against CRC in a murine xenograft model.
Materials and methods
Antibodies
Purified mouse IgG (cat. no. I8765) and mouse
IgG2a (cat. no. M7769) were purchased from
Sigma-Aldrich; Merck KGaA. Anti-EpCAM mAbs were purified using
Protein G-Sepharose (Cytiva).
Animals
All animal experiments were performed in accordance
with institutional guidelines and regulations to minimize animal
suffering and distress in the laboratory. The Institutional
Committee for Experiments of the Institute of Microbial Chemistry
(Numazu, Japan) approved the animal studies for ADCC and antitumor
activity (approval no. 2019-066). Mice were monitored for health
and weight every 1–5 days. Experiments on mice were conducted in ≤3
weeks. Weight loss >25% or tumor size >3,000 mm3
were identified as humane endpoints for euthanasia. At humane and
experimental endpoints, mice were euthanized by cervical
dislocation, and death was verified by validating respiratory and
cardiac arrest.
Cell lines
P3X63Ag8U.1 (P3U1), CHO-K1 and Caco-2 cells were
obtained from the American Type Culture Collection. The Genome
Network Project clone IRAK021G03 (EpCAM) was provided by the RIKEN
BioResource Research Center through the National BioResource
Project of the MEXT and AMED agencies of Japan (18–21).
EpCAM DNA plus a C-terminal PA tag recognized by the anti-PA tag
mAb (NZ-1) was subcloned into a pCAG-Ble vector (FUJIFILM Wako Pure
Chemical Corporation). CHO/EpCAM was established by transfecting
pCAG/EpCAM-PA into CHO-K1 cells using the Neon Transfection System
(Thermo Fisher Scientific, Inc.). CHO-K1 cells (1.5×106)
were transfected with 10 µg of plasmid DNA using 100 µl Neon tip,
at room temperature. After 4 days, cells were incubated with 1
µg/ml anti-EpCAM mAb (clone 9C4; cat. no. 324202; BioLegend, Inc.)
for 30 min on ice and subsequently with Alexa Fluor 488-conjugated
anti-mouse IgG (1:1,000; cat. no. 4408; Cell Signaling Technology,
Inc.) for 30 min on ice. Positive cells for anti-EpCAM mAb were
sorted using an SH800 cell sorter (Sony Corporation), and stable
transfectants were cultivated in RPMI-1640 medium (Nacalai Tesque,
Inc.) containing 0.5 mg/ml zeocin (InvivoGen). Using TruGuide gRNA
tool, gRNA of EpCAM (NM_002354) was selected from GeneArt
predesigned gRNAs database (Thermo Fisher Scientific, Inc.). gRNA
sequence used was GATCCTGACTGCGATGAGAG(cgg), which targeted exon 3
of EpCAM (Assay ID, CRISPR701274). Double strand gRNA sequence was
subcloned into GeneArt CRISPR Nuclease Vector with OFP Reporter
(Thermo Fisher Scientific, Inc.). Caco-2/EpCAM-knockout (BINDS-16)
cells were generated by transfecting 10 μg of CRISPR/Cas9 plasmids
for EpCAM (Thermo Fisher Scientific, Inc.) into Caco-2 cells
(1.5×106) for 7 days using a Neon transfection system
with 100 µl Neon tip. Stable transfectants were established by cell
sorting as aforementioned. P3U1, CHO-K1 and CHO/EpCAM cells were
cultured in RPMI-1640 medium (Nacalai Tesque, Inc.). Caco-2 and
BINDS-16 were cultured in Dulbecco's modified Eagle's medium (DMEM;
Nacalai Tesque, Inc.). The medium was supplemented with 10%
heat-inactivated fetal bovine serum (FBS; Thermo Fisher Scientific
Inc.), 100 U/ml penicillin, 100 µg/ml streptomycin and 0.25 µg/ml
amphotericin B (Nacalai Tesque, Inc.), and the cells were incubated
at 37°C in a humidified atmosphere containing 5%
CO2.
Hybridoma production
The CBIS method (17)
was used in the present study to develop mAbs against EpCAM.
Briefly, one BALB/c mouse was intraperitoneally (i.p.) immunized
with CHO/EpCAM cells (1×108 cells/500 µl) with Imject
Alum adjuvant (Thermo Fisher Scientific Inc.). The procedure
included three additional immunizations, followed by a final
booster injection administered i.p. 2 days before spleen cell
harvesting. Spleen cells (1×108 cells) were then fused
with mouse plasma cell myeloma P3U1 cells (1×107 cells)
using PEG1500 (Roche Diagnostics). The hybridomas were cultured in
RPMI-1640 medium supplemented with hypoxanthine, aminopterin and
thymidine for selection (50X solution; Thermo Fisher Scientific
Inc.). Cell culture supernatants of hybridomas in each well of
96-well plates were mixed with 1×105 CHO/EpCAM cells and
were directly screened using flow cytometry.
Flow cytometry
Caco-2 cells (2.5×105 cells/ml) were
harvested after brief exposure to 0.25% trypsin in 1 mM
ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque, Inc.).
Following washing with 0.1% bovine serum albumin (BSA; Nacalai
Tesque, Inc.) in phosphate-buffered saline (PBS), Caco-2 cells were
treated with 1 µg/ml of anti-EpCAM mAbs for 30 min at 4°C, followed
by Alexa Fluor 488-conjugated anti-mouse IgG (1:1,000; cat. no.
4408; Cell Signaling Technology, Inc.). Fluorescence data were
obtained using an EC800 Cell Analyzer (Sony Corporation), and
analyzed using the originally installed EC800 software v1.3.6 (Sony
Corporation).
Western blot analysis
Cell pellets were lysed in PBS with 1% Triton X-100
and 50 µg/ml aprotinin (cat. no. 03346-84; Nacalai Tesque, Inc.).
Protein concentration was determined using the BCA assay. Cell
lysates of CHO-K1, CHO/EpCAM, Caco-2 and BINDS-16 cells were boiled
in sodium dodecyl sulfate sample buffer (Nacalai Tesque, Inc.). The
samples (10 µg/lane) were then electrophoresed on 5–20%
polyacrylamide gels (Nacalai Tesque, Inc.) and transferred to
polyvinylidene difluoride membranes (Merck KGaA). Following
blocking with 4% milk (Nacalai Tesque, Inc.) for 1 h, the membrane
was incubated with anti-EpCAM (1 µg/ml) or anti-β-actin (1 µg/ml;
clone AC-15; cat no. A5441; Sigma-Aldrich; Merck KGaA) antibodies
for 1 h, followed by incubation with HRP-conjugated anti-mouse IgG
(cat. no. P0260, Agilent Technologies, Inc.) or anti-rat IgG (cat.
no. A9542; Sigma-Aldrich; Merck KGaA) at a 1:2,000 dilution for 1 h
at room temperature. The membrane was developed using the
ImmunoStar LD Chemiluminescence Reagent (FUJIFILM Wako Pure
Chemical Corporation) and a Sayaca-Imager (DRC Co., Ltd.). All
western blotting procedures were performed at room temperature.
Immunohistochemical analyses
Histological sections (4 µm) of a colorectal
adenocarcinoma tissue array (cat. no. CO243b; US Biomax Inc.) were
autoclaved directly in citrate buffer (pH 6.0; Nichirei Bioscience,
Inc.) for 20 min. The sections were then incubated with 1 μg/ml
anti-EpCAM mAb for 1 h at room temperature and treated using an
Envision+ kit (Agilent Technologies, Inc.) for 30 min at room
temperature. The color was developed using 3,3′-diaminobenzidine
tetrahydrochloride (Agilent Technologies Inc.) for 2 min at room
temperature, and sections were then counterstained with hematoxylin
(FUJIFILM Wako Pure Chemical Corporation) for 2 min at room
temperature. Hematoxylin and eosin (H&E) staining (FUJIFILM
Wako Pure Chemical Corporation) was performed using consecutive
colorectal adenocarcinoma tissue sections for 2 min at room
temperature.
Determination of the binding
affinity
Caco-2 cells were suspended in 100 µl serially
diluted anti-EpCAM mAb (0.006–100 µg/ml), followed by the addition
of Alexa Fluor 488-conjugated anti-mouse IgG (1:200; cat. no. 4408;
Cell Signaling Technology, Inc.). Fluorescence data were obtained
using an EC800 Cell Analyzer, and analyzed using the originally
installed EC800 v1.3.6 software (Sony Corporation). The
dissociation constant (KD) was calculated by
fitting binding isotherms to built-in, one-site binding models in
GraphPad Prism 6 (GraphPad Software, Inc.).
ADCC
ADCC induction by EpCAM was assayed as follows: A
total of 6 female 5-week-old BALB/c nude mice were purchased from
Charles River Laboratories, Inc. Following euthanasia by cervical
dislocation, the spleens were removed aseptically, and single-cell
suspensions were obtained by forcing spleen tissues through a
sterile cell strainer (cat. no. 352360; Corning, Inc.) with a
syringe. Erythrocytes were lysed by 10-sec exposure to ice-cold
distilled water. The splenocytes were washed with DMEM and
resuspended in DMEM with 10% FBS; this yield was designated as
effector cells. Caco-2 cells were labeled with 10 µg/ml Calcein-AM
(Thermo Fisher Scientific, Inc.) and resuspended in DMEM with 10%
FBS. Caco-2 cells were transferred to 96-well plates at
2×104 cells/well and mixed with the effector cells at an
effector-to-target ratio of 50:1, along with 100 µg/ml anti-EpCAM
mAb or control mouse IgG2a. Following a 4-h incubation,
Calcein-AM release into the supernatant was measured in each well
using a Power Scan HT microplate reader (BioTek Instruments, Inc.)
with an excitation wavelength of 485 nm and an emission wavelength
of 538 nm. Cytolytic activity was determined as a percentage of
lysis and calculated using the following equation: Lysis (%) = (E -
S) / (M - S) × 100, where E is the fluorescence measured in the
co-cultures of target and effector cells, S is the spontaneous
fluorescence of the target cells, and M is the maximum fluorescence
measured after lysis of all cells with a buffer containing 0.5%
Triton X-100, 10 mM Tris-HCl (pH 7.4) and 10 mM EDTA.
CDC
CDC by EpCAM was assayed as follows: Caco-2 cells
were labeled with 10 µg/ml Calcein-AM and resuspended in DMEM with
10% FBS. Caco-2 cells were plated in 96-well plates at
2×104 cells/well, and 10% Low-Tox-M rabbit complement
(Cedarlane Laboratories) with 100 µg/ml anti-EpCAM mAb or control
mouse IgG2a was added to each well. Following a 4-h
incubation at 37°C, Calcein-AM release into the supernatant was
determined in each well. Fluorescence intensity was calculated as
described in the ADCC section.
Antitumor activity of anti-EpCAM mAb
in colorectal adenocarcinoma xenografts
A total of 32 5-week-old female BALB/c nude mice
were purchased from Charles River Laboratories, Inc. After a 2-week
acclimation period, the mice were used in experiments at 7 weeks of
age. Caco-2 cells (0.3 ml; 1.33×108 cells/ml in DMEM)
were mixed with 0.5 ml BD Matrigel Matrix Growth Factor Reduced (BD
Biosciences), and 100 µl of this suspension (5×106
cells) was injected subcutaneously into the left flank of each
animal. On day 1 post-inoculation, 100 µg anti-EpCAM mAb or control
mouse IgG in 100 µl PBS was injected i.p. Additional antibody
inoculations were performed on days 7 and 12. Tumor formation was
measured in mice in the treatment and control groups on days 7, 8,
12, 15 and 17 after Caco-2 cell injection. On day 17 after cell
implantation, all mice were euthanized by cervical dislocation, and
tumor diameters and volumes were measured and recorded.
Statistical analyses
Data are presented as the mean ± SEM. Statistical
analysis was conducted with one-way ANOVA and Tukey's multiple
comparisons tests for ADCC and CDC; one-way ANOVA and Sidak's
multiple comparisons tests for tumor volume and mouse weight; and
Welch's t-test for tumor weight. All calculations were performed
with GraphPad Prism 7 (GraphPad Software, Inc.). P<0.05 was
considered to indicate a statistically significant difference.
Results
Establishment and characterization of
the anti-EpCAM mAb
The anti-EpCAM mAb was established by immunizing one
mouse with CHO/EpCAM cells and fusing its spleen cells with P3U1
cells. Supernatants from hybridomas, which were positive for
CHO/EpCAM and negative for CHO-K1, were selected by flow cytometry.
Further screening by immunohistochemistry and western blotting was
performed for validation, resulting in the establishment of
EpMab-16 (IgG2a, κ).
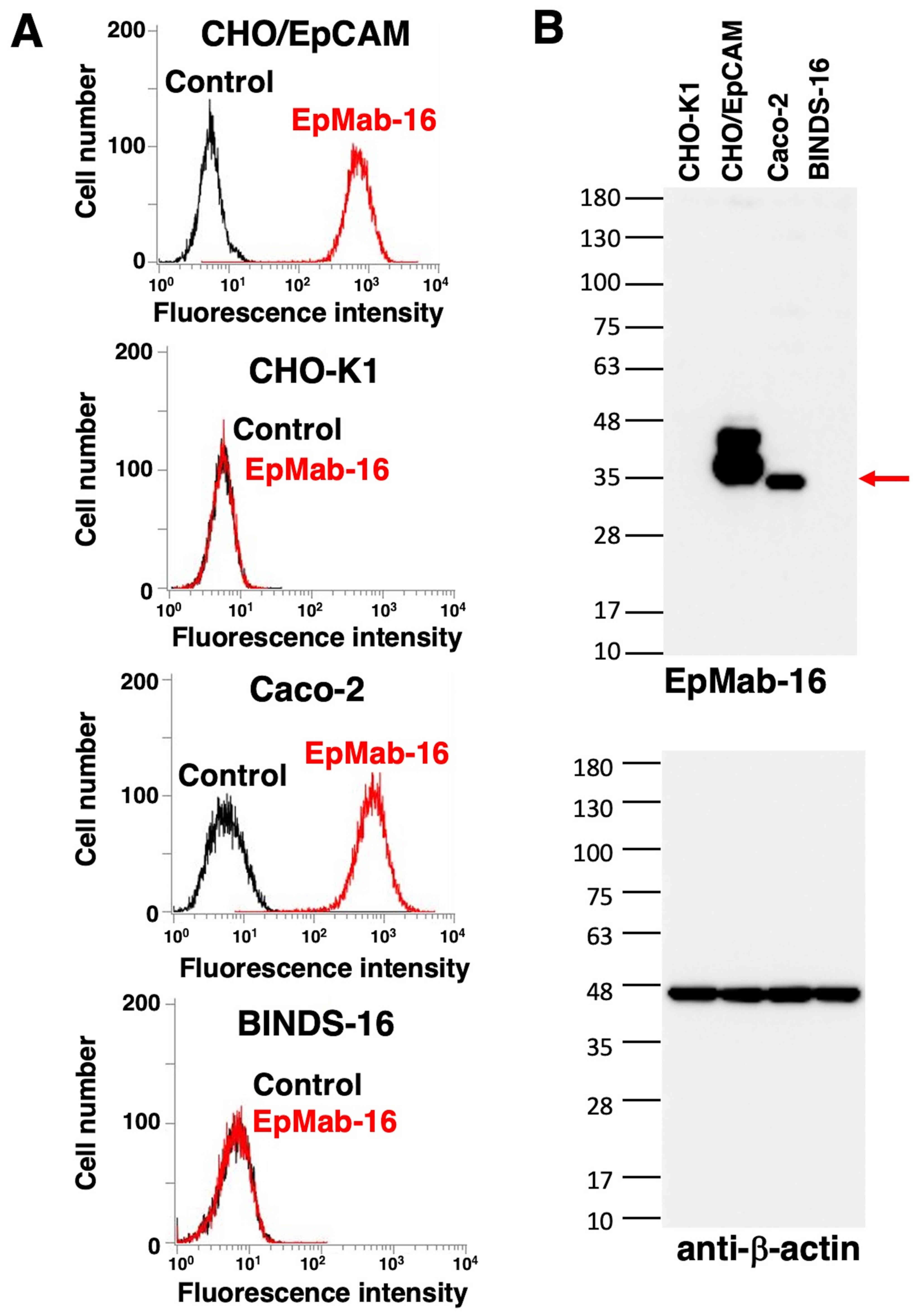
Flow cytometry was performed to assess the
sensitivity of EpMab-16 in CHO/EpCAM cells and the Caco-2
colorectal adenocarcinoma cell line. As presented in Fig. 1A, EpMab-16 bound to CHO/EpCAM cells,
but not CHO-K1 cells. EpMab-16 also bound to Caco-2 but not
BINDS-16 cells, indicating that EpMab-16 was specific for EpCAM in
the colorectal adenocarcinoma cell line.
Western blotting was performed to further assess the
sensitivity of EpMab-16. Lysates of CHO-K1, CHO/EpCAM, Caco-2 and
BINDS-16 cells were probed. As demonstrated in Fig. 1B, EpMab-16 detected the 35-kDa band
of EpCAM in lysates from CHO/EpCAM and Caco-2 cells, whereas this
band was not present in lysates from CHO-K1 and BINDS-16 cells,
indicating that EpMab-16 specifically detected both exogenous and
endogenous EpCAM. The molecular weight of EpCAM between CHO/EpCAM
and Caco-2 was different as a PA tag (12 amino acids) was added to
C-terminus of EpCAM in CHO/EpCAM cells.
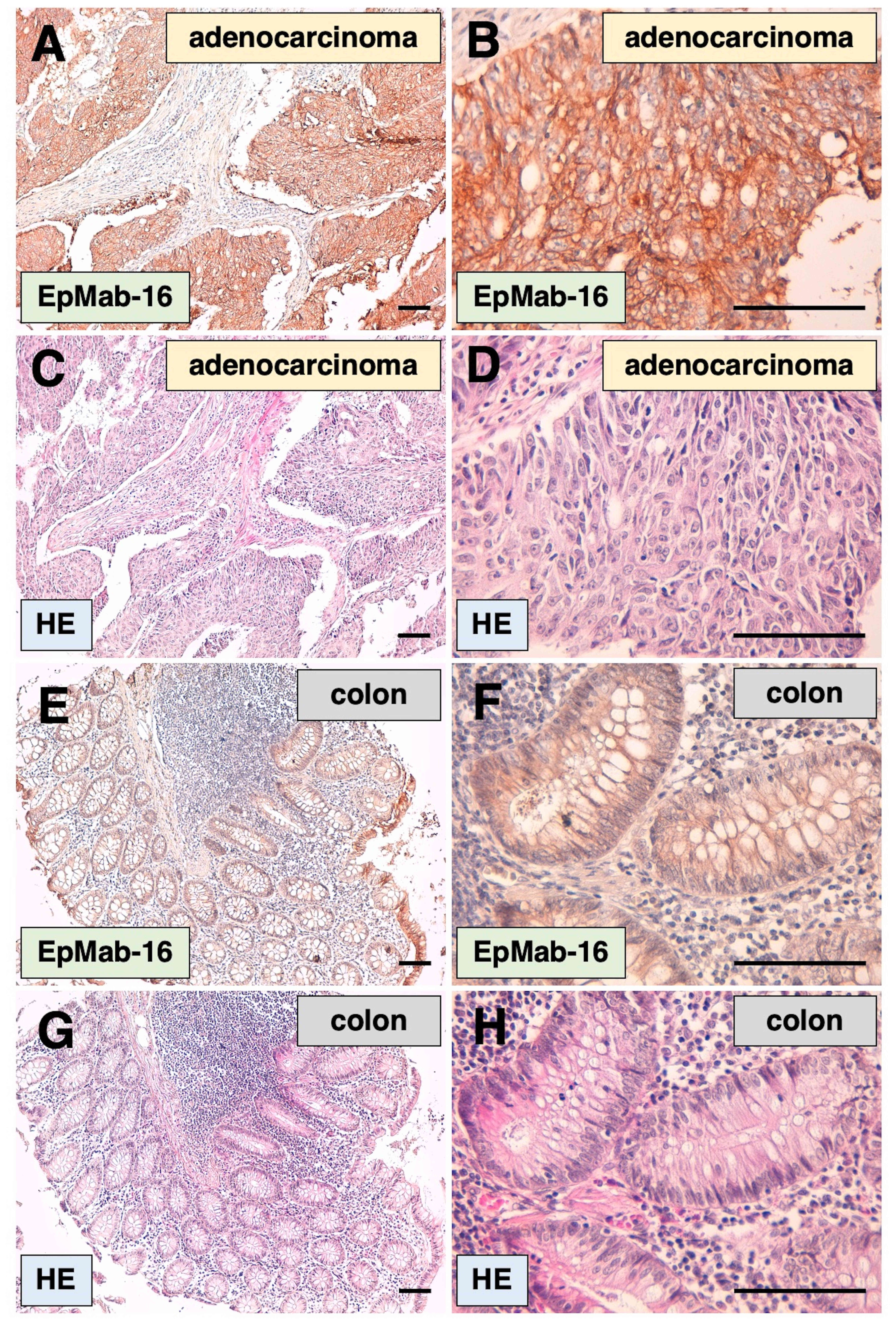
EpMab-16 detected membrane antigens in colorectal
adenocarcinoma tissues in the immunohistochemical analysis
(Fig. 2A and B). Among six cases of
colorectal adenocarcinoma, five (83%) were positively stained by
EpMab-16. No staining was observed without the primary mAb (data
not shown). H&E staining was performed to stain nucleus as blue
and cytosol as pink, using consecutive colorectal adenocarcinoma
tissue sections (Fig. 2C and D).
Furthermore, EpMab-16 weakly detected membrane antigens in normal
colon tissues (Fig. 2E and F). Among
six samples of normal colon tissues, three tissues (50%) were
stained by EpMab-16. No staining was observed without the primary
mAb (data not shown). H&E staining was performed to stain
nucleus as blue and cytosol as pink, using consecutive normal colon
tissue sections (Fig. 2G and H).
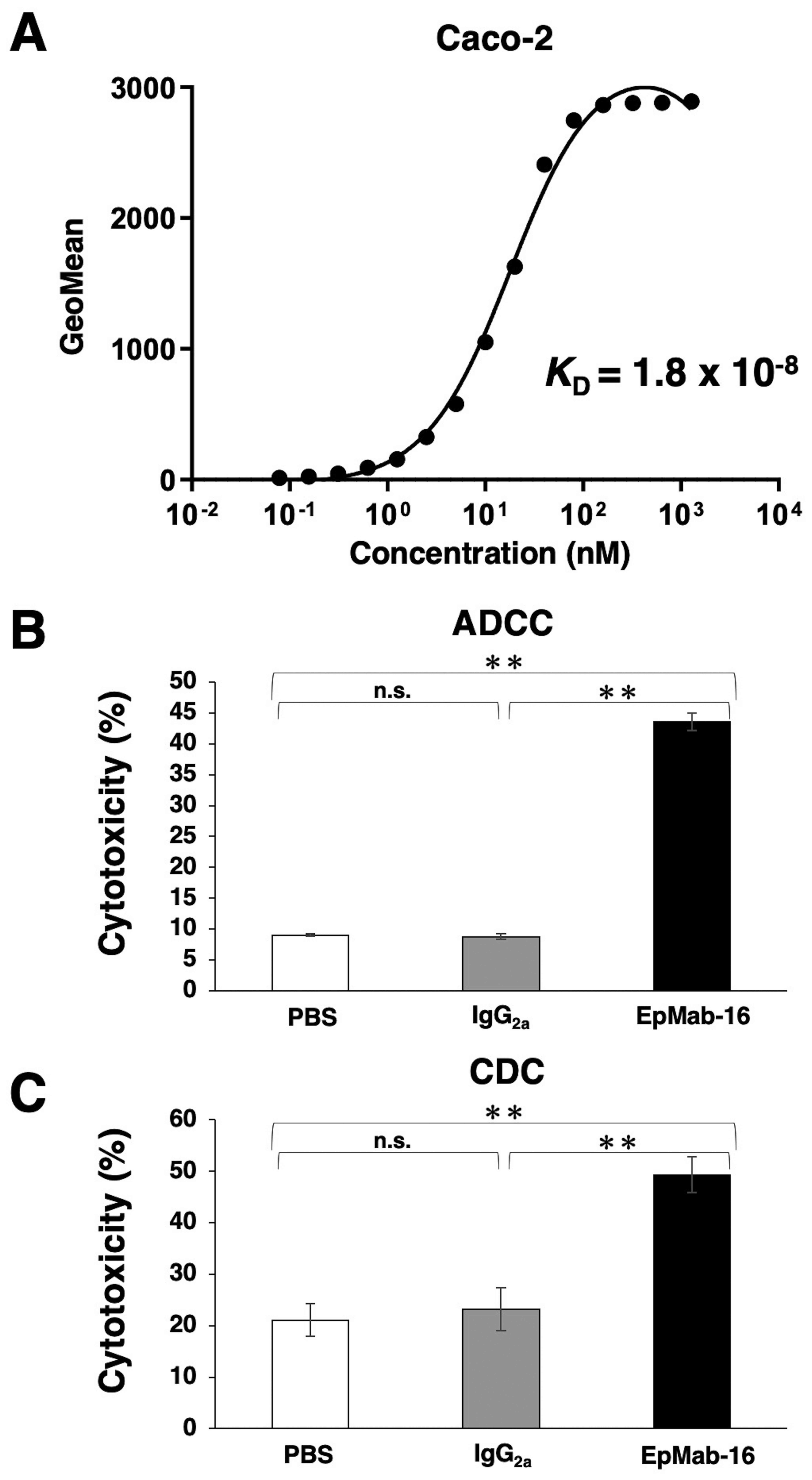
Kinetic analysis of the interactions of EpMab-16
with Caco-2 cells was subsequently analyzed using flow cytometry.
The KD for EpMab-16 in Caco-2 cells was
calculated to be 1.8×10−8 M (Fig. 3A), indicating a moderate binding
affinity of EpMab-16 to colorectal adenocarcinoma cells.
ADCC and CDC activities of EpMab-16 in
a colorectal adenocarcinoma cell line
The present study further investigated whether
EpMab-16 induced ADCC and CDC antitumor activity in an
EpCAM-expressing Caco-2 colorectal adenocarcinoma cell line. As
presented in Fig. 3B, EpMab-16
exhibited higher ADCC (44% cytotoxicity) in Caco-2 cells compared
with that of control mouse IgG2a (8.7% cytotoxicity;
P<0.01) or control PBS (9.0% cytotoxicity; P<0.01) treatment.
EpMab-16 was also associated with more robust CDC activity (49%
cytotoxicity) in Caco-2 cells compared with that of control mouse
IgG2a (23% cytotoxicity; P<0.01) or control PBS (21%
cytotoxicity; P<0.01) treatment (Fig.
3C). These results suggested that EpMab-16 induced strong ADCC
and CDC antitumor activity in vitro.
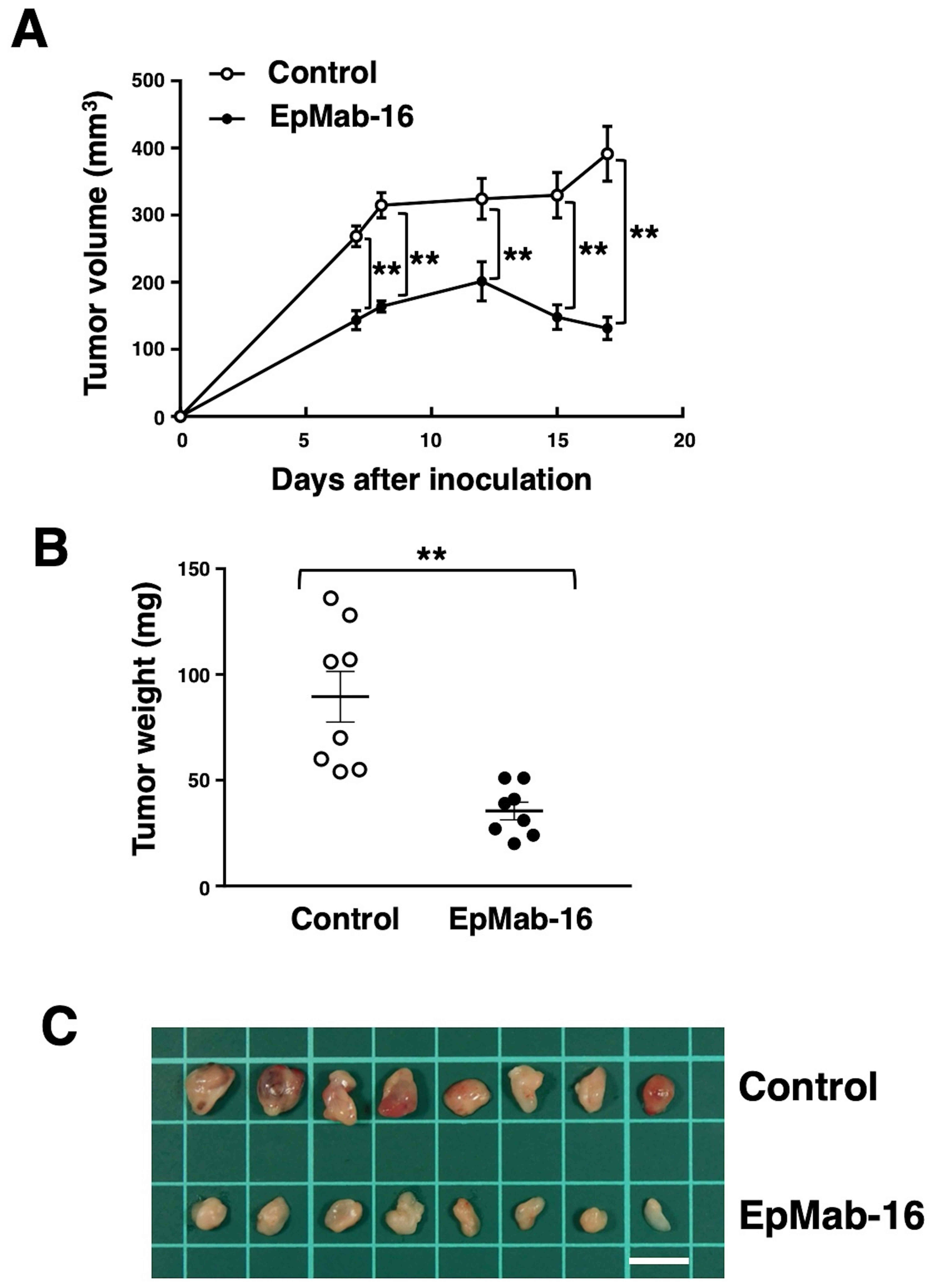
EpMab-16 antitumor activity in mouse
xenografts of Caco-2 colorectal adenocarcinoma cells
We further examined whether EMab-16 exerts antitumor
activity in vivo. On days 1, 7 and 12 after Caco-2 cell
injections into the mice, the Caco-2 ×enograft mouse models were
injected with EpMab-16 or control mouse IgG. During the animal
experiment, no apparent weight loss due to tumor burden or organ
failure was observed among the mice. EpMab-16-treated mice
exhibited significantly lower tumor growth on days 7, 8, 12, 15 and
17 (all P<0.01) compared with that in IgG-treated control mice
(Fig. 4A). Tumor volume reduction by
EpMab-16 treatment reached 66% relative to the control group on day
17. Tumors from EpMab-16-treated mice weighed significantly less
compared with those from IgG-treated control mice (60% reduction;
P<0.01; Fig. 4B). Resected tumors
on day 17 are depicted in Fig. 4C.
Total body weights did not significantly differ between the
treatment and control groups (Fig.
S1). These results indicated that EpMab-16 reduced the growth
of Caco-2 ×enografts, but did not altogether eliminate it.
Discussion
The present study aimed to determine whether a novel
anti-EpCAM mAb may be useful for treating colorectal
adenocarcinoma. First, a sensitive and specific anti-EpCAM mAb,
EpMab-16, was developed, which exhibited high reactivity for
colorectal adenocarcinoma by flow cytometry, western blotting and
immunohistochemical analyses. Notably, the results also suggested
that EpMab-16 exhibited diagnostic efficacy in FFPE tissues as
pathological diagnosis utilizes FFPE tissues. EpMab-16 was
demonstrated to possess strong ADCC and CDC activity against the
Caco-2 colorectal adenocarcinoma cell line in vitro, and
significantly reduced not only the volume but also the weight of
Caco-2 ×enografts in vivo. Although tumor reduction was
determined to be 66% by volume and 60% by weight on day 17, the
reduction was not sufficient to eliminate the tumor entirely.
Cancer stem cells (CSCs) exhibit specific
characteristics including decontrolled self-renewal,
tumor-initiating and tumor-promoting properties, and chemotherapy
resistance. Therefore, targeting CSCs is thought to be a promising
approach to treat cancer (22).
Since CD133 is thought to be one of CSC markers for CRC (23–25), we
previously developed an anti-CD133 mAb (CMab-43, mouse
IgG2a) (17) and
investigated whether CD133 may represent a therapeutic target in
colorectal adenocarcinoma using CMab-43 (26). Importantly, CMab-43 exerted antitumor
activity in Caco-2 ×enograft models at a dose of 100 µg/mouse/week
administered three times, suggesting that CMab-43 may be useful for
antibody therapy against CD133-positive CRC (26). In the present study, the effects of
an anti-EpCAM mAb, EpMab-16, were examined in vitro and
in vivo, as EpCAM is also known to be a CSC marker for CRC
(25). In the present study,
EpMab-16 also showed antitumor activity against CRC in a murine
xenograft model. CSC marker-expressing cells may be heterogenous;
therefore, the combination of mAbs or anticancer drugs to kill the
CSC marker-expressing cancer cells could be an ideal strategy to
treat CRC.
In another recent study, we established an anti-HER2
mAb (H2Mab-19, mouse IgG2b) (27). Our previous study demonstrated that
H2Mab-19 significantly reduced tumor development in
HER2-expressing breast cancer cells (BT-474), oral cancer cells
(HSC-2 and SAS) and CRC cells (Caco-2) xenograft models, suggesting
that treatment with H2Mab-19 may be a useful therapy for
patients with HER2-expressing cancers (27,28).
Although H2Mab-19 showed significant antitumor activity,
immunohistochemical analysis revealed that HER2 expression was
diminished in the remaining cancer cells after H2Mab-19
treatment (28). These results
suggest that H2Mab-19 treatment may not be fully
effective for patients with HER2-expressing cancers. In the current
study, an EpCAM-targeting mAb, EpMab-16, was developed. To enhance
the therapeutic effects in vivo, the combinational use of
different mAbs, such as H2Mab-19 and EpMab-16 may be
required to treat HER2 and/or EpCAM-expressing CRC cells.
The success of antibody-based therapeutics of cancer
depends on the target antigen and the antibody to be used. It has
been reported that EpCAM is upregulated in 94% of CRCs (29), whereas EpCAM expression is restricted
to only the basolateral membrane of epithelial cells in normal
tissues, and therapeutic agents have limited access to it (30), underscoring the importance of EpCAM
as a therapeutic target. Since EpMab-16 appears to react with not
only the basolateral membrane, but also the apical membrane of
epithelial cells according to the results of the present study, the
expression pattern of EpCAM should be further investigated by
comparing EpMab-16 with other anti-EpCAM mAbs.
A number of antibody-based therapeutic approaches
targeting EpCAM have been developed (10). Previously, Liao et al
(31) generated an anti-EpCAM mAb
EpAb2-6. Using a colon cancer xenograft model, they demonstrated
that EpAb2-6 induced CRC cell death by inhibiting EpCAM signaling
rather than by acting through the ADCC or CDC (31). On the other hand, EpMab-16 generated
in the present study possessed high ADCC and CDC activities against
CRC cells and exhibited antitumor activity in vivo. Another
previous study demonstrated that an anti-EpCAM toxin-conjugated
antibody Oportuzumab monatox (also termed VB4-845) was effective
against squamous cell carcinomas of the head and neck and
non-muscle invasive bladder cancer, and well-tolerated in clinical
trials (phase I and II) (32).
Considering that EpCAM is also expressed in tumor-initiating cells,
antibody-based therapies against EpCAM may kill not only
proliferating cancer cells, but also drug-resistant dormant CRC
cells (6). These results suggest
that the EpCAM-targeted immunotherapy may be a promising
therapeutic strategy for CRC. To further develop EpCAM-targeted
cancer therapy, further studies are required to elucidate the
precise role of EpCAM inhibition among various cancer cells.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Mr. Takuro Nakamura,
Ms. Miyuki Yanaka, Ms. Saori Handa, Ms. Saki Okamoto and Mr. Yu
Komatsu (Department of Antibody Drug Development, Tohoku University
Graduate School of Medicine, Sendai, Japan) for technical
assistance with the in vitro experiments, and Ms. Akiko
Harakawa [Institute of Microbial Chemistry (BIKAKEN), Microbial
Chemistry Research Foundation, Numazu, Japan] for technical
assistance with the animal experiments.
Funding
This research was supported in part by the Japan
Agency for Medical Research and Development (grant nos.
JP20am0401013 to YK, JP20am0101078 to YK and JP20ae0101028 to
YK).
Availability of data and materials
The datasets used and/or analyzed during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
HH, TA, YS, and MKK performed the in vitro
experiments. TO performed in vivo experiments. JT analyzed
the experimental data. MK and YK designed the study and wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Animal studies for ADCC and the antitumor activity
were approved by the Institutional Committee for Experiments of the
Institute of Microbial Chemistry (Numazu, Japan; approval no.
2019-066).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ADCC
|
antibody-dependent cellular
cytotoxicity
|
|
CBIS
|
cell-based immunization and
screening
|
|
CDC
|
complement-dependent cytotoxicity
|
|
EpCAM
|
epithelial cell adhesion molecule
|
|
mAb
|
monoclonal antibody
|
References
|
1
|
Okegawa T, Pong RC, Li Y and Hsieh JT: The
role of cell adhesion molecule in cancer progression and its
application in cancer therapy. Acta Biochim Pol. 51:445–457. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petruzzelli L, Takami M and Humes HD:
Structure and function of cell adhesion molecules. Am J Med.
106:467–476. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maheshwari G, Brown G, Lauffenburger DA,
Wells A and Griffith LG: Cell adhesion and motility depend on
nanoscale RGD clustering. J Cell Sci. 113:1677–1686.
2000.PubMed/NCBI
|
|
4
|
Trzpis M, McLaughlin PM, de Leij LM and
Harmsen MC: Epithelial cell adhesion molecule: More than a
carcinoma marker and adhesion molecule. Am J Pathol. 171:386–395.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Herlyn M, Steplewski Z, Herlyn D and
Koprowski H: Colorectal carcinoma-specific antigen: Detection by
means of monoclonal antibodies. Proc Natl Acad Sci USA.
76:1438–1442. 1979. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Imrich S, Hachmeister M and Gires O: EpCAM
and its potential role in tumor-initiating cells. Cell Adhes Migr.
6:30–38. 2012. View Article : Google Scholar
|
|
7
|
Litvinov SV, Velders MP, Bakker HA,
Fleuren GJ and Warnaar SO: Ep-CAM: A human epithelial antigen is a
homophilic cell-cell adhesion molecule. J Cell Biol. 125:437–446.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Maetzel D, Denzel S, Mack B, Canis M, Went
P, Benk M, Kieu C, Papior P, Baeuerle PA, Munz M, et al: Nuclear
signalling by tumour-associated antigen EpCAM. Nat Cell Biol.
11:162–171. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sen S and Carnelio S: Expression of
epithelial cell adhesion molecule (EpCAM) in oral squamous cell
carcinoma. Histopathology. 68:897–904. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Baeuerle PA and Gires O: EpCAM (CD326)
finding its role in cancer. Br J Cancer. 96:417–423. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Went P, Vasei M, Bubendorf L, Terracciano
L, Tornillo L, Riede U, Kononen J, Simon R, Sauter G and Baeuerle
PA: Frequent high-level expression of the immunotherapeutic target
Ep-CAM in colon, stomach, prostate and lung cancers. Br J Cancer.
94:128–135. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wolpin BM, Meyerhardt JA, Mamon HJ and
Mayer RJ: Adjuvant treatment of colorectal cancer. CA Cancer J
Clin. 57:168–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Longo WE and Johnson FE: The preoperative
assessment and postoperative surveillance of patients with colon
and rectal cancer. Surg Clin North Am. 82:1091–1108. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
O'Connell MJ, Colangelo LH, Beart RW,
Petrelli NJ, Allegra CJ, Sharif S, Pitot HC, Shields AF, Landry JC,
Ryan DP, et al: Capecitabine and oxaliplatin in the preoperative
multimodality treatment of rectal cancer: Surgical end points from
National Surgical Adjuvant Breast and Bowel Project trial R-04. J
Clin Oncol. 32:1927–1934. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Duffy MJ, Lamerz R, Haglund C, Nicolini A,
Kalousová M, Holubec L and Sturgeon C: Tumor markers in colorectal
cancer, gastric cancer and gastrointestinal stromal cancers:
European group on tumor markers 2014 guidelines update. Int J
Cancer. 134:2513–2522. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Itai S, Fujii Y, Nakamura T, Chang YW,
Yanaka M, Saidoh N, Handa S, Suzuki H, Harada H, Yamada S, et al:
Establishment of CMab-43, a sensitive and specific anti-CD133
monoclonal antibody, for immunohistochemistry. Monoclon Antib
Immunodiagn Immunother. 36:231–235. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ota T, Suzuki Y, Nishikawa T, Otsuki T,
Sugiyama T, Irie R, Wakamatsu A, Hayashi K, Sato H, Nagai K, et al:
Complete sequencing and characterization of 21,243 full-length
human cDNAs. Nat Genet. 36:40–45. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Otsuki T, Ota T, Nishikawa T, Hayashi K,
Suzuki Y, Yamamoto J, Wakamatsu A, Kimura K, Sakamoto K, Hatano N,
et al: Signal sequence and keyword trap in silico for selection of
full-length human cDNAs encoding secretion or membrane proteins
from oligo-capped cDNA libraries. DNA Res. 12:117–126. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kimura K, Wakamatsu A, Suzuki Y, Ota T,
Nishikawa T, Yamashita R, Yamamoto J, Sekine M, Tsuritani K,
Wakaguri H, et al: Diversification of transcriptional modulation:
Large-scale identification and characterization of putative
alternative promoters of human genes. Genome Res. 16:55–65. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Itoh M, Yasunishi A, Imamura K,
Kanamori-Katayama M, Suzuki H, Suzuki M, Carninci P, Kawai J and
Hayashizaki Y: Constructing ORFeome resources with removable
termination codons. Biotechniques. 41:44–50, 46, 48 passim. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dzobo K, Senthebane DA, Ganz C, Thomford
NE, Wonkam A and Dandara C: Advances in therapeutic targeting of
cancer stem cells within the tumor microenvironment: An updated
review. Cells. 9:18962020. View Article : Google Scholar
|
|
23
|
O'Brien CA, Pollett A, Gallinger S and
Dick JE: A human colon cancer cell capable of initiating tumour
growth in immunodeficient mice. Nature. 445:106–110. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ricci-Vitiani L, Lombardi DG, Pilozzi E,
Biffoni M, Todaro M, Peschle C and De Maria R: Identification and
expansion of human colon-cancer-initiating cells. Nature.
445:111–115. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lugli A, Iezzi G, Hostettler I, Muraro MG,
Mele V, Tornillo L, Carafa V, Spagnoli G, Terracciano L and Zlobec
I: Prognostic impact of the expression of putative cancer stem cell
markers CD133, CD166, CD44s, EpCAM, and ALDH1 in colorectal cancer.
Br J Cancer. 103:382–390. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kato Y, Ohishi T, Yamada S, Itai S,
Furusawa Y, Sano M, Nakamura T, Kawada M and Kaneko MK: Anti-CD133
monoclonal antibody CMab-43 exerts antitumor activity in a mouse
xenograft model of colon cancer. Monoclon Antib Immunodiagn
Immunother. 38:75–78. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takei J, Kaneko MK, Ohishi T, Kawada M,
Harada H and Kato Y: H2Mab-19, an anti-human epidermal
growth factor receptor 2 monoclonal antibody exerts antitumor
activity in mouse oral cancer xenografts. Exp Ther Med. 20:846–853.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kato Y, Ohishi T, Takei J, Nakamura T,
Sano M, Asano T, Sayama Y, Hosono H, Kawada M and Kaneko MK: An
anti-human epidermal growth factor receptor 2 monoclonal antibody
H2Mab-19 exerts antitumor activity in mouse colon cancer
xenografts. Monoclon Antib Immunodiagn Immunother. 39:123–128.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Spizzo G, Fong D, Wurm M, Ensinger C,
Obrist P, Hofer C, Mazzoleni G, Gastl G and Went P: EpCAM
expression in primary tumour tissues and metastases: An
immunohistochemical analysis. J Clin Pathol. 64:415–420. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Balzar M, Prins FA, Bakker HA, Fleuren GJ,
Warnaar SO and Litvinov SV: The structural analysis of adhesions
mediated by Ep-CAM. Exp Cell Res. 246:108–121. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liao MY, Lai JK, Kuo MY, Lu RM, Lin CW,
Cheng PC, Liang KH and Wu HC: An anti-EpCAM antibody EpAb2-6 for
the treatment of colon cancer. Oncotarget. 6:24947–24968. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Eyvazi S, Farajnia S, Dastmalchi S,
Kanipour F, Zarredar H and Bandehpour M: Antibody Based EpCAM
Targeted Therapy of Cancer, Review and Update. Curr Cancer Drug
Targets. 18:857–868. 2018. View Article : Google Scholar : PubMed/NCBI
|