Introduction
Hepatocellular cancer (HCC) is one of the most
common invasive malignancies, ranking 4th among cancer-related
causes of death in the world in 2018 (1). Despite advances in diagnostic and
treatment strategies, the prognosis for HCC remains poor (2). The high proliferation and invasion
activities of HCC cells are the main reasons for the poor prognosis
of patients with HCC (3). The
hypoxic characteristic of solid tumor tissue is caused by the rapid
growth of the tumor and relatively low density of microvessels
(4). It has been reported that
hypoxia is involved in the angiogenesis, metastasis, proliferation,
and chemotherapy resistance in patients with cancer (5–7). For
instance, hypoxia-induced miR-182 accelerates the angiogenesis of
HCC cells by downregulating Ras GTPase-activating protein 1
(5). Moreover, hypoxia-induced
circular RNA (circRNA) circELP3 facilitates cisplatin resistance
and tumor growth in bladder cancer (7). Though hypoxia exerts vital roles in the
progression of HCC, its potential molecular mechanisms remain
unclear.
CircRNAs are a novel type of RNA with a closed
circular structure (8). They
regulate gene expression by modulating protein, miRNA and
transcription functions (9).
Moreover, circRNAs were revealed to be associated with the
carcinogenesis of a range of malignant tumors (10). For example, circRNA circNT5E promotes
the tumorigenesis of glioblastoma through sponging miR-422a
(11). circRNA circPRKCI accelerates
lung adenocarcinoma growth by elevating E2F7 expression by sponging
miR-589 and miR-545 (12).
Furthermore, circRNAs were involved in hypoxia-mediated cancer
progression (13–15). Circular RNA hsa_circ_0008450
(circ_0008450), located at chromosome 16, is formed by the reverse
splicing of the CMTM3 [CKLF-like MARVEL transmembrane domain
containing (CMTM) 3] gene (from exon 3 to exon 5). Furthermore, it
has been reported that circ_0008450 was associated with the
development of nasopharyngeal cancer (16). Additionally, circ_0008450 may be
acting as a potential prognostic marker for HCC (17). However, it remains unclear whether
circ_0008450 is involved in the malignant behavior of HCC under
hypoxic conditions.
MicroRNAs (miRNAs or miRs) are a type of key
regulator of gene expression and mainly regulate gene expression at
the post-transcriptional level (18). They are involved in cell
differentiation and metabolism, organismal development, oncogenesis
and viral infection (19). miR-431
has been reported to act as a suppressor in multiple cancers,
including pancreatic cancer (20),
colorectal cancer (21) and lung
cancer (22). Additionally, miR-431
participated in the advancement of HCC (23). However, whether miR-431 mediated the
development of HCC under hypoxic conditions has not been
reported.
A-kinase anchor protein 1 (AKAP1) is a member of the
A-kinase Ankyrin family and serves important roles in intracellular
cAMP signal transmission to the outer mitochondrial member
(24). Moreover, AKAP1 regulated
oxidative metabolism, organelle biogenesis and cell survival
(25). Additionally, AKAP1 regulated
endothelial cells behavior and vascular function (26). High AKAP1 expression predicted poor
prognosis of HCC (27). However, the
molecular mechanism of AKAP1 involved in the malignant behavior of
HCC under hypoxic conductions needs to be further investigated.
The current study aimed to investigate the impacts
of circ_0008450 on viability, apoptosis and glycolysis of HCC cells
under hypoxic conductions.
Materials and methods
Study subjects
The current study was authorized by the Ethics
Committee of Lanzhou University Second Hospital, Lanzhou, China. A
total of 30 paired HCC tissues and corresponding adjacent normal
tissues (>2.0 cm from the tumor margin) were obtained from
patients with HCC who underwent resections at Lanzhou University
Second Hospital. The clinicopathological features of HCC patients
were presented in Table I. Moreover,
serum samples were drawn from 30 patients with HCC and 30 healthy
volunteers from the same hospital. All subjects were recruited
between April 2017 and June 2019. The inclusion criteria for
patients with HCC: i) Diagnosed with HCC and no other tumors and
ii) without any treatment before surgery. The inclusion criteria
for healthy volunteers were healthy individuals who were
age-matched with the patients with HCC and had never been diagnosed
with cancer. Written informed consent was obtained from patients
with HCC and healthy volunteers.
 | Table I.Association between circ_0008450
expression and clinical clinicopathological parameters of patients
with hepatocellular cancer. |
Table I.
Association between circ_0008450
expression and clinical clinicopathological parameters of patients
with hepatocellular cancer.
|
|
| circ_0008450
expression |
|
|---|
|
|
|
|
|
| Parameter | Patients, n | Low n=14 | High n=16 | P-value |
|---|
| Age, years |
|
|
| 0.07 |
|
≤60 | 14 | 9 | 5 |
|
|
>60 | 16 | 5 | 11 |
|
| Sex |
|
|
| 0.232 |
|
Female | 18 | 10 | 8 |
|
|
Male | 12 | 4 | 8 |
|
| Tumor size, cm |
|
|
| 0.003a |
| ≤5 | 17 | 12 | 5 |
|
|
>5 | 13 | 2 | 11 |
|
| TNM stage |
|
|
| 0.003a |
|
I–II | 15 | 11 | 4 |
|
|
III | 15 | 3 | 12 |
|
| AFP, µg/l |
|
|
| 0.696 |
|
≤400 | 16 | 8 | 8 |
|
|
>400 | 14 | 6 | 8 |
|
| Lymphatic
metastasis |
|
|
| 0.0001a |
|
Negative | 12 | 11 | 1 |
|
|
Positive | 18 | 3 | 15 |
|
| Vascular
invasion |
|
|
| 0.961 |
|
Absent | 13 | 6 | 7 |
|
|
Present | 17 | 8 | 9 |
|
| Distant
metastasis |
|
|
| 0.0003a |
| M0 | 15 | 12 | 3 |
|
| M1 | 15 | 2 | 13 |
|
Cell culture and treatment
HCC cell lines SNU-387 and Huh7, and human normal
liver epithelial cells THLE-2, were purchased from BeNa Culture
Collection. All cells were cultivated in RPMI-1640 medium
(Sigma-Aldrich; Merck KGaA) supplemented with 10% FBS
(Sigma-Aldrich; Merck KGaA) and 1% penicillin/streptomycin stock
solution (Sigma-Aldrich; Merck KGaA) in a humidified atmosphere
with 5% CO2 at 37°C. For the hypoxia treatment, SNU-387
and Huh7 cells were incubated in a humidified atmosphere with 94%
N2, 5% CO2 and 1% O2 for 48 h.
Cell transfection
Small interference RNA targeting circ_0008450
(si-circ_0008450, 5′-ACAAGACAGAAGGGTCTCTCA-3′) and negative
controls (si-NC, 5′-GCGCGATAGCGCGAATATA-3′) were purchased from
Guangzhou RiboBio Co., Ltd. miR-431 mimics
(3′-ACGUACUGCCGGACGUUCUGU-5′), inhibitors
(3′-ACAGAACGTCCGGCAGTACGT-5′) and their negative controls (miRNA NC
and inhibitor NC) were obtained from Shanghai GenePharma, Co., Ltd.
The full-length sequences of circ_0008450 and AKAP1 were cloned
into the pcDNA3.1 vector (vector; Invitrogen; Thermo Fisher
Scientific, Inc.) or pCD5-ciR vector (pc-NC; Greenseed Biotech,
Co.) to obtain the overexpression vectors for circ_0008450 and
AKAP1, respectively. Oligonucleotides or vectors were transiently
transfected into SNU-387 and Huh7 cells using the Lipofectamine
3000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
transfection concentration of oligonucleotides was as follows:
si-NC (40 nM), si-circ_0008450 (40 nM), miR-431 Mimics (50 nM),
miRNA NC (50 nM), miR-431 inhibitors (50 nM), inhibitor NC (50 nM).
After transfection, cells were treated under hypoxia conditions for
48 h.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from tissues, serum and cells was
extracted using the TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). cDNA for circ_0008450, CMTM3, AKAP1 and
miR-431 was generated using a Moloney Murine Leukemia Virus (M-MLV)
First Strand kit (Thermo Fisher Scientific, Inc.) or a miRNA
Reverse Transcription kit (Takara Biotechnology Co., Ltd.)
according to the manufacturer's protocols. RT-qPCR was performed
using SYBR Premix Ex Taq Takara Biotechnology Co., Ltd., with the
following thermocycling conditions: 95°C for 30 sec, followed by 40
cycles of 95°C for 5 sec and 60°C for 20 sec. The expression of
circ_0008450, CMTM3, AKAP1, HIF-1α and miR-431 was quantified using
the 2−ΔΔCq method (28).
Primers used were as follows: circ_0008450 forward (divergent),
5′-TGATGCATCCCATCCACCCT-3′ and reverse (divergent),
5′-AGCCCTGCCACTTGTCATTC-3′; CMTM3 forward (convergent)
5′-TTTTATCTGCTATGTGGCGTCC-3′ and reverse (convergent),
5′-TGTCTTGTGGGCTGTGGTCTC-3′; AKAP1 forward,
5′-GCTTACGGCTTGTACCTGAAG-3′ and reverse,
5′-ATGGTGCTCTTGGAAATACGC-3′; miR-431 forward,
5′-CAGGCCGTCATGCAAA-3′ and reverse, 5′-CGCTTCAGAATTTGCGTGTCAT-3′;
U6 small nuclear RNA (snRNA) forward, 5′-GCTCGCTTCGGCAGCACA-3′ and
reverse, 5′-GAGGTATTCGCACCAGAGGA-3′; HIF-1α forward,
5′-TATGAGCCAGAAGAACTTTTAGGC-3′ and reverse,
5′-CACCTCTTTTGGCAAGCATCCTG-3′; GAPDH forward (convergent),
5′-GACTCCACTCACGGCAAATTCA-3′ and reverse (convergent),
5′-TCGCTCCTGGAAGATGGTGAT-3′; and GAPDH forward (divergent),
5′-GAAGGTGAAGGTCGAGTC-3′ and reverse (divergent),
5′-GAAGATGGTGATGGGATTTC-3′. Oligo(dt)18 primers (cat.
no. SO132) and random primers (cat. no. 48190011) were bought from
(Invitrogen; Thermo Fisher Scientific, Inc.). GAPDH or U6 snRNA
served as an internal control. To verify the splicing from
head-to-tail of circRNA, we designed divergent primers and
convergent primers to amplify circRNA and linear RNA, and GAPDH was
used as negative control.
Actinomycin D treatment and RNase R
digestion
Huh7 cells were treated with 50 ng/ml Actinomycin D
(Sigma-Aldrich; Merck KGaA) at different times (0, 4, 8, 12 or 24
h) to block new RNA synthesis. Additionally, total Huh7 RNA was
degraded with 3 U/µg RNase R (Epicentre; Illumina, Inc.) at 37°C
for 15 min.
Cell viability assay
Viability of transfected SNU-387 and Huh7 cells was
assessed by Cell Counting kit-8 (CCK-8) assays. The cells were
cultured under hypoxia conditions for 48 h after transfection.
Cells were then incubated with 10 µl CCK-8 reagent (Dojindo
Molecular Technologies, Inc.) for 2 h. Color reactions were
determined using a Microplate Absorbance Reader (Bio-Rad
Laboratories, Inc.).
Flow cytometry assay
Apoptosis rate of transfected and hypoxia-treated
SNU-387 and Huh7 cells was evaluated using an Annexin
V-FITC/propidium iodide (PI) apoptosis detection kit
(Sigma-Aldrich; Merck KGaA) according to the manufacturer's
protocols. Briefly, the cells were collected and resuspended in
binding buffer and labeled with 5 µl Annexin V-FITC and 10 µl PI in
the dark at room temperature for 15 min. The apoptotic rate (early
+ late apoptotic cells) was analyzed using a FACScan flow cytometer
(Becton-Dickinson and Company) with Cell Quest software (version
4.0.2, Becton-Dickinson and Company).
Measurement of glucose consumption and
lactate production
Cell supernatants of transfected and hypoxia-treated
SNU-387 and Huh7 cells were collected and the glucose consumption
and lactate production of the cells were assessed using a Glucose
Assay kit or a Lactic Acid Assay kit (both from Sigma-Aldrich;
Merck KGaA) in accordance with the manufacturer's procedures.
Dual-luciferase reporter assay
Binding sites of miR-431 in circ_0008450 or AKAP1
were predicted using CircRNA Interactome (https://circinteractome.irp.nia.nih.gov/) or Starbase
(http://starbase.sysu.edu.cn/) databases.
Following this, fragments of circ_0008450 (position, 68–73; CAAGAC)
or the 3′-untranslated region (UTR) of AKAP1 (position, 431–438;
GCAAGAC; harbored miR-431 binding sites) was amplified and inset
into the pGL3 vector (Promega Corporation) for the construction of
wild-type luciferase reporter vectors. Mutant luciferase reporter
vectors for circ_0008450 (GUUCUG) or AKAP1 (CGUUCUG; within miR-431
binding sites) were established using the same method. Luciferase
reporter vectors and miRNA NC, miR-431 mimics, inhibitor NC or
miR-431 inhibitors were cotransfected into 293T cells using
Lipofectamine 3000 reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). After 48 h, the luciferase intensities of the luciferase
reporter vectors in 293T cells were determined using a luciferase
reporter assay kit (Promega Corporation) based on the
manufacturer's procedures. Relative activity was normalized to
Renilla luciferase activity.
Western blotting
HCC cells were lysed using lysis buffer (Beyotime
Institute of Biotechnology) to extract total protein and western
blotting was conducted as previously described (29). A BCA™ Protein Assay kit (Pierce;
Thermo Fisher Scientific, Inc.) was utilized to quantify the
concentration of total protein. Briefly, 10% SDS-PAGE was used to
separate the total protein (30 µg/lane). Membranes with transferred
proteins were blocked with TBST supplemented with 5% BSA
(Sigma-Aldrich; Merck KGaA). Primary antibodies used were as
follows: rabbit anti-AKAP1 (cat. no. ab156004; 1:1,000; Abcam),
rabbit anti-cleaved caspase-3 (caspase-3) (cat. no. ab2302; 1:300;
Abcam), and rabbit-anti-GAPDH (1:10,000; cat. no. ab181602).
Secondary antibodies used were goat anti-rabbit antibodies (cat.
no. ab97051; 1:10,000; Abcam). Protein bands were visualized using
ImmunoStar LD (Fujifilm Wako Pure Chemical Corporation) and GAPDH
was used as the loading control. Densitometry analysis was
performed using ImageJ software (version 1.6; National Institutes
of Health).
Xenograft assay
Animal experiments were approved by the Animal
Ethics Committee of Lanzhou University Second Hospital. A total of
8 female BALB/c nude mice (weight, 17–22 g; age, 4–6 weeks) were
obtained from the Shanghai Experimental Animal Center for the
xenograft assays. Briefly, PHBLV-U6-ZSGreen-puro lentivirus vectors
(Hanbio Biotechnology Co., Ltd.) with short hairpin
(sh)-circ_0008450 were infected into Huh7 cells for 1 week under
puromycin. Following this, Huh7 cells (5×106)
transfected with sh-NC or lentivirus-mediated sh-circ_0008450 were
resuspended in 50 µl PBS (Beijing Solarbio Science & Technology
Co., Ltd.) and subcutaneously injected into the dorsal side of nude
mice (4 mice/group). All mice were kept in a specific pathogen-free
room (50±5% humidity) with a 12 h light/12 h dark cycle at 25°C and
free access to water and food. Tumor volumes were measured using a
digital caliper every 4 days from day 7 post-injection and
calculated using the following equation: Volume = (length ×
width2)/2. Mice were anesthetized with 10 mg/kg xylazine
and euthanized by cervical decapitation on day 27 post-injection.
Tumor tissues were excised after death as assessed by pupil
dilation and cardiac arrest. Tumor tissues were weighed and then
digested with TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) for the evaluation of the expression levels of
circ_0008450, miR-431 and AKAP1.
Statistical analysis
Experiments were performed in triplicate. GraphPad
Prism software (version 5.0; GraphPad Software, Inc.) and SPSS
software (version 22.0; SPSS, Inc.) were used for statistical
analysis. Data are presented as mean ± standard deviation. Paired
or unpaired Student's t-tests were performed to analyze the
differences between two groups and ANOVA with Tukey's pos-hoc test
was used to determine differences between ≥3 groups. The
association between circ_0008450 expression and clinical
clinicopathological parameters of patients with HCC were determined
using the χ2 test. Receiver operating characteristic
curve (ROC) analysis was performed to analyze differences between
patients with HCC and healthy controls. P<0.05 was considered to
indicate a statistically significant difference.
Results
Circ_0008450 is upregulated in the
tissues and serum of patients with HCC patients and HCC cells
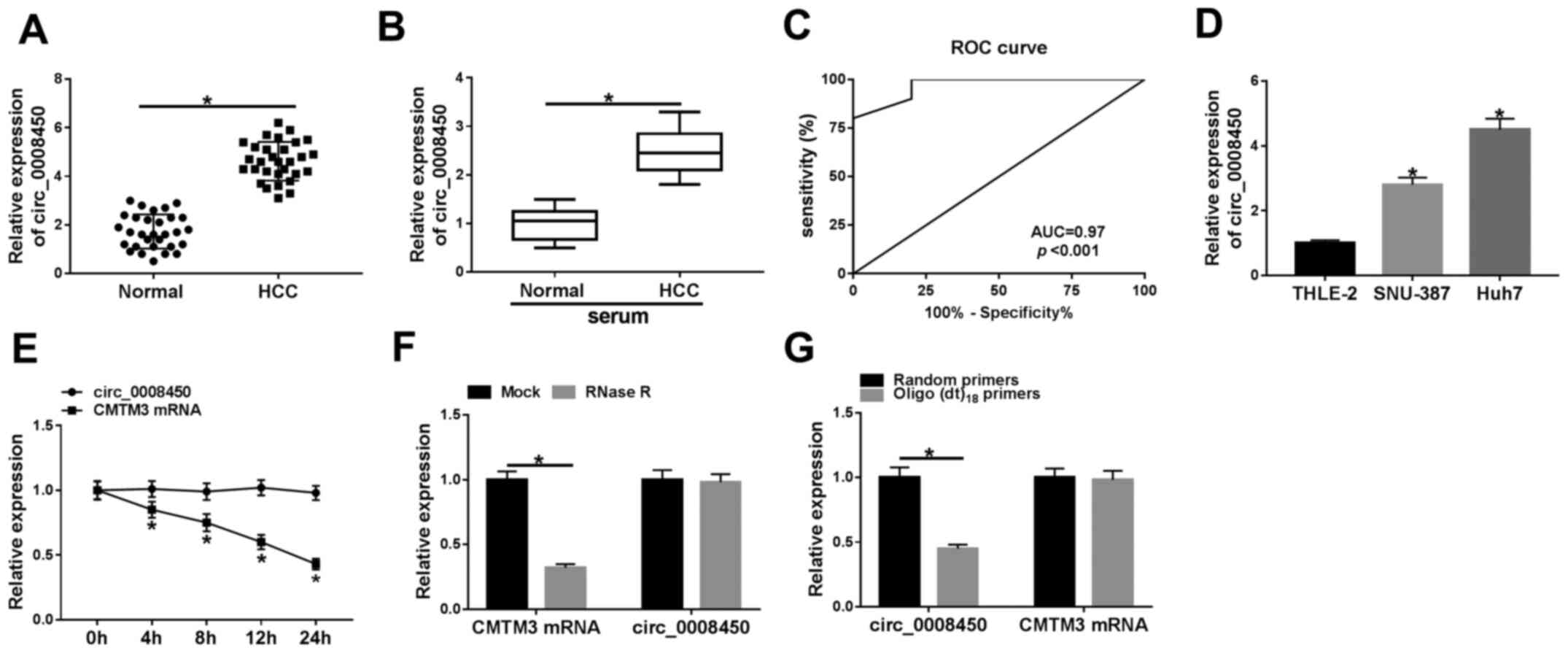
To verify the expression pattern of circ_0008450 in
HCC, the expression of circ_0008450 in 30 paired HCC tissues and
adjacent normal tissues were examined using RT-qPCR. The results
demonstrated that circ_0008450 expression was significantly
increased in HCC tissues compared with adjacent normal tissues
(Fig. 1A). Furthermore, the
expression of HIF-1α was increased in HCC tissues compared with
adjacent normal tissues (Fig. S1).
The expression of circ_0008450 was significantly increased in the
serum of patients with HCC patients compared with healthy controls
(Fig. 1B). Moreover, ROC analysis
demonstrated that circ_0008450 exhibited extremely high diagnostic
value to distinguish patients with HCC from healthy controls
(AUC=0.97; Fig. 1C). High
circ_0008450 expression was associated with tumor size (P=0.003),
TNM stage (P=0.003), lymphatic metastasis (P=0.0001) and distant
metastasis (P=0.0003); however, circ_0008450 expression was not
associated to age, sex, α-fetoprotein (AFP) or vascular invasion
(Table I). Furthermore, the
expression of circ_0008450 was elevated in HCC cells (SNU-387 and
Huh7) compared with THLE-2 cells (Fig.
1D). Circ_0008450 and linear GAPDH were amplified based on cDNA
and genomic DNA (gDNA) using divergent and convergent primers. The
results demonstrated that circ_0008450 was amplified by primers in
cDNA; however, circ_000845 was not amplified by gDNA (Fig. S2). Subsequently, the circularization
of circ_0008450 was further verified using Huh7 cells. Circ_0008450
was more stable and resistant to actinomycin D treatment compared
with linear CMTM3 (Fig. 1E). There
was no significant difference in circ_0008450 expression following
RNase R treatment; however, the linear CMTM3 gene was digested
(Fig. 1F). Furthermore, circ_0008450
expression was significantly decreased using oligo(dt)18
primers compared with random primers, while the expression of CMTM3
did not exhibit a significant change, indicating that circ_0008450
did not have a ploy-A tail structure (Fig. 1G). These data indicated that
circ_0008450 had high diagnostic value and its elevation may be
associated with the progression of HCC.
Circ_0008450 depletion represses
viability and glycolysis, and accelerates apoptosis of HCC cells
following hypoxia treatment
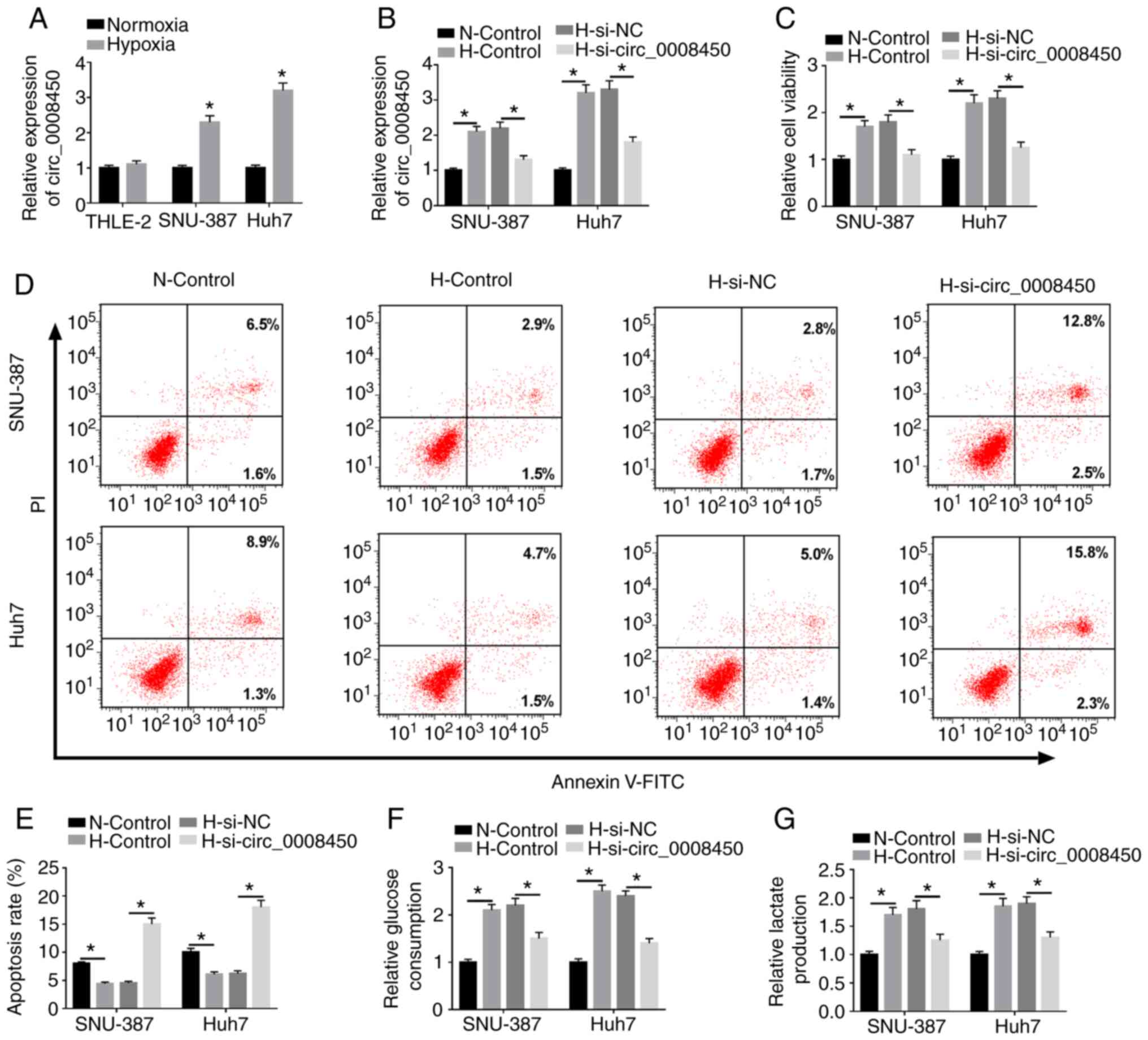
Due to the upregulation of circ_0008450 in HCC, the
role of circ_0008450 in HCC under hypoxia was investigated. RT-qPCR
results revealed that circ_0008450 was significantly upregulated in
SNU-387 and Huh7 cells treated with hypoxia compared with SNU-387
or Huh7 cells under normoxic conditions (Fig. 2A). Additionally, no significant
change in THLE-2 cells. Regardless of whether the circ_0008450 gene
was suppressed, the apoptosis and glycolysis of THLE-2 cells were
not significantly different under normoxic or hypoxic conditions
(Fig. S3). Hypoxia-induced
enhancement of circ_0008450 in SNU-387 and Huh7 cells was abolished
by the introduction of si-circ_0008450 (Fig. 2B). Additionally, the viability of
SNU-387 and Huh7 cells was increased under hypoxia and this effect
was reversed by circ_0008450 inhibition (Fig. 2C). The results of flow cytometry
assays demonstrated that the promotive effect of hypoxia on the
apoptosis of SNU-387 and Huh7 cells was abolished by circ_0008450
suppression (Fig. 2D and E).
Furthermore, circ_0008450 knockdown partly reversed the
downregulation of caspase-3 in SNU-387 and Huh7 cells following
hypoxia treatment (Fig. S4).
Additionally, the results of glycolysis assays demonstrated that
glucose consumption (Fig. 2F) and
lactate production (Fig. 2G) were
increased in SNU-387 and Huh7 cells following hypoxia treatment.
These effects were reversed by circ_0008450 knockdown.
Collectively, these findings revealed that circ_0008450 silencing
reverse cell viability, glycolysis and lactate production, and
accelerated cell apoptosis in hypoxia-treated HCC cells.
Circ_0008450 targets miR-431 in
hypoxia-treated HCC cells
To examine the regulatory mechanism of circ_0008450
in HCC, the underlying target for circ_0008450 was predicted. The
CircRNA interactome database revealed that circ_0008450 had binding
sites for miR-431 (Fig. 3A). The
data revealed that miR-431 mimics significantly repressed the
luciferase intensity of luciferase reporters containing
WT-circ_0008450 in 293T cells compared with miRNA NC, while the
luciferase intensity of luciferase reporters containing
MUT-circ_0008450 was not significantly altered (Fig. 3B). Additionally, the luciferase
activity of luciferase reporters containing WT-circ_0008450 in 293T
was significantly increased by miR-431 inhibitors (Fig. 3C) and circ_0008450 expression was
significantly elevated in SNU-387 and Huh7 cells following
circ_0008450 transfection compared to control vectors (Fig. S5). Furthermore, miR-431 expression
was significantly increased by circ_0008450 downregulation and was
significantly decreased by circ_0008450 enhancement in SNU-387 and
Huh7 cells (Fig. 3D). miR-431
expression was reduced in SNU-387 and Huh7 cells under hypoxia
treatment (Fig. 3E). Additionally,
miR-431 was significantly upregulated in SNU-387 and Huh7 cells
transfected with miR-431 mimic and was significantly downregulated
in cells transfected with miR-431 inhibitor under hypoxic
conditions (Fig. 3F and G). Overall,
these data indicated that circ_0008450 acted as a sponge for
miR-431 in HCC.
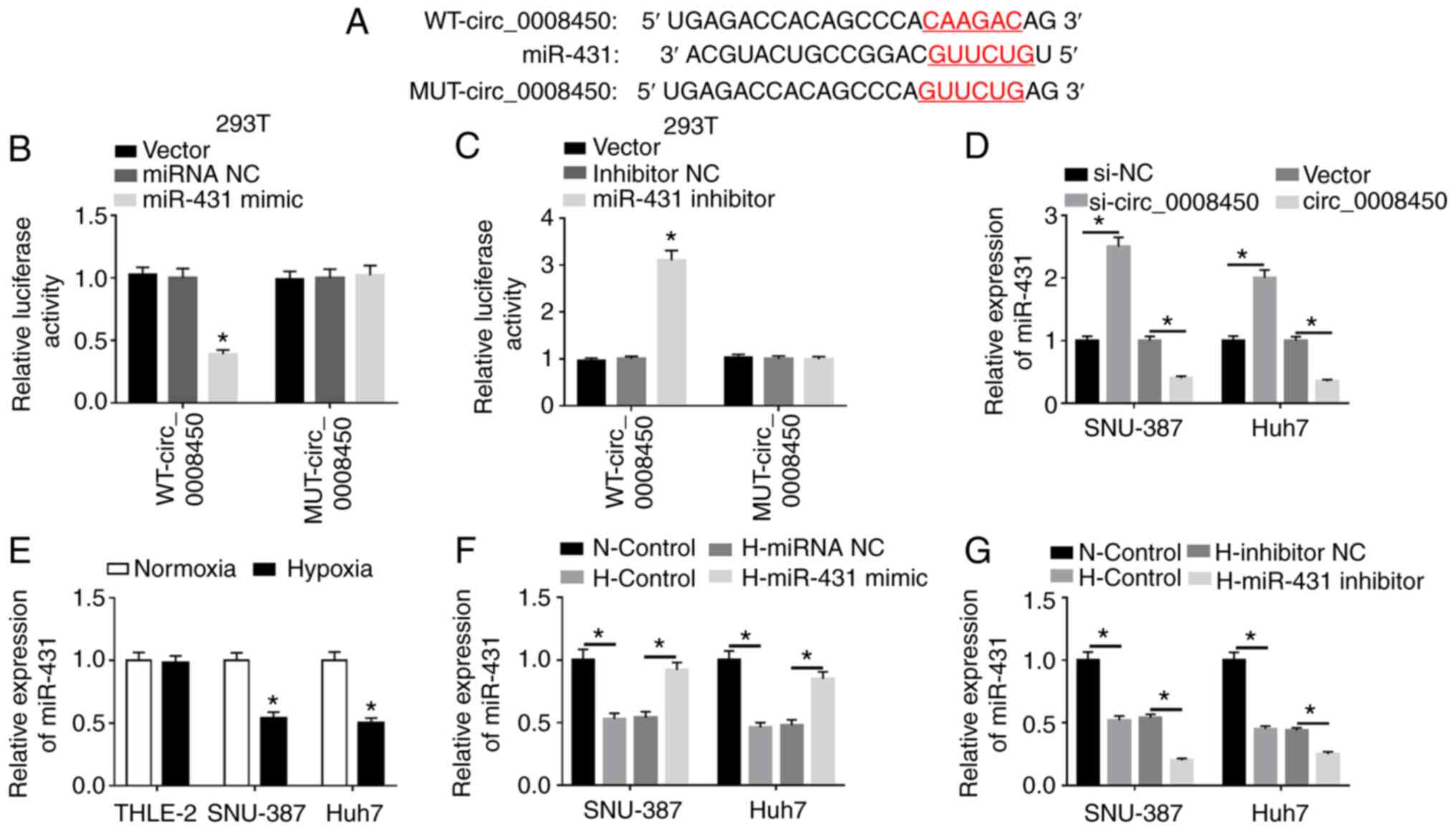 | Figure 3.miR-431 acted as a target for
circ_0008450 in hepatocellular cancer cells. (A) The binding sites
of circ_0008450 in miR-431 were predicted using the CircRNA
interactome database. Luciferase intensities of the WT and MUT
luciferase reporter vectors of circ_0008450 in 293T cells
transfected with (B) miRNA NC, miR-431 mimic, (C) inhibitor NC or
miR-431 inhibitor were determined by dual-luciferase reporter
assays. (D) Effect of circ_0008450 on the expression of miR-431 was
assessed using RT-qPCR. (E) The expression of miR-431 in THLE-2,
SNU-387 and Huh7 cells treated with hypoxia was detected by
RT-qPCR. The transfection efficiency of miR-431 (F) overexpression
and (G) silencing on SNU-387 and Huh7 cells treated with hypoxia
was evaluated by RT-qPCR. *P<0.05. miR, microRNA; circ,
circular; WT, wild-type; MUT, mutant; NC, negative control;
RT-qPCR, reverse transcription-quantitative PCR; N, normoxic; H,
hypoxic. |
miR-431 inhibitor reverses
circ_0008450 knockdown-mediated effects on viability, apoptosis and
glycolysis of HCC cells following hypoxia treatment
Based on the above findings, whether miR-431 was
associated with the viability, apoptosis and glycolysis of HCC
cells mediated by circ_0008450 following hypoxia was explored. The
upregulation of miR-431 in hypoxia-treated SNU-387 and Huh7 cells
caused by circ_0008450 silencing was partly recovered by miR-431
inhibition (Fig. 4A). CCK-8 assays
demonstrated that the repressive effect of circ_0008450 knockdown
on viability of SNU-387 and Huh7 cells under hypoxia treatment was
restored by miR-431 suppression (Fig.
4B). Flow cytometry assays revealed that decreased miR-431
expression reversed the increased apoptosis of hypoxia-disposed
SNU-387 and Huh7 cells induced by circ_0008450 silencing (Fig. 4C). Additionally, miR-431 knockdown
overturned the decrease of glucose consumption and lactate
production in SNU-387 and Huh7 cells caused by circ_0008450
depletion under hypoxia treatment (Fig.
4D and E). These results indicated that circ_0008450 mediated
the viability, apoptosis and glycolysis of hypoxia-exposed SNU-387
and Huh7 cells by sponging miR-431.
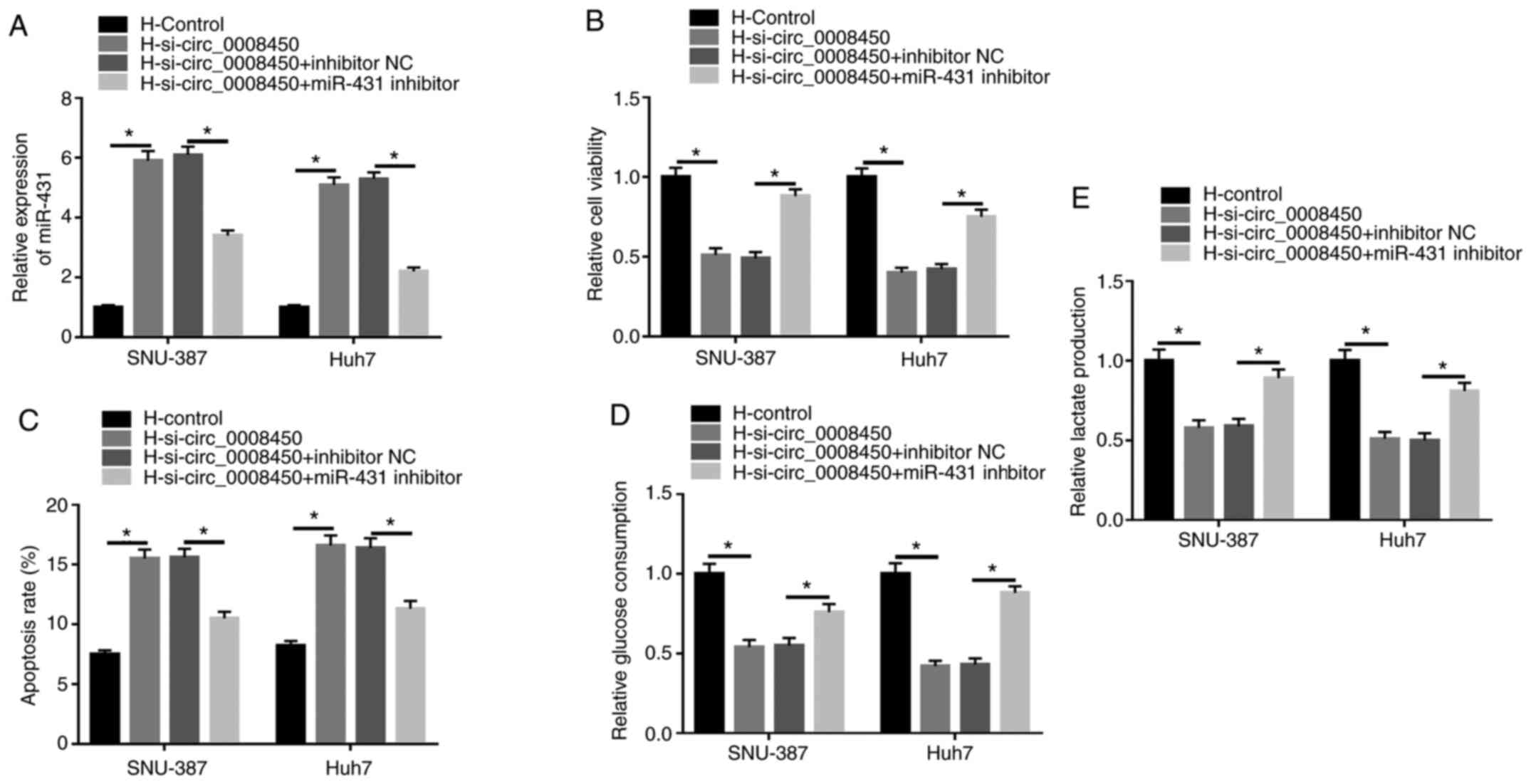 | Figure 4.Circ_0008450 regulated the viability,
apoptosis and glycolysis of hepatocellular cancer cells via miR-431
following hypoxia treatment. (A) Influence of miR-431 repression on
circ_0008450 downregulation-mediated miR-431 expression was
analyzed by reverse transcription-quantitative PCR. Effect of
miR-431 inhibition on circ_0008450 reduction-mediated (B)
viability, (C) apoptosis, (D and E) glycolysis in hypoxia-treated
SNU-387 and Huh7 cells were determined using Cell Counting kit-8,
flow cytometry and glycolysis assays. *P<0.05. circ, circular;
miR, microRNA; si-, small interfering; H, hypoxic; N, normoxic; NC,
negative control. |
AKAP1 is a target for miR-431
Subsequently, the target for miR-431 was
investigated using the Starbase database. The results demonstrated
that the 3′UTR of AKAP1 mRNA possessed possible binding sites for
miR-431 (Fig. 5A). Luciferase
intensity in 293T cells was significantly reduced following
cotransfection luciferase reporters containing WT-AKAP1 3′UTR and
miR-431 mimic, while the luciferase intensity of luciferase
reporters containing MUT-AKAP13′UTR were not significantly
different (Fig. 5B). Luciferase
intensity of luciferase reporters containing WT-AKAP1 3′UTR was
enhanced in miR-431-inhibited 293T cells (Fig. 5C). Moreover, miR-431 mimics
significantly decreased the levels of AKAP1 mRNA and protein in
SNU-387 and Huh7 cells, while the levels of AKAP1 mRNA and protein
were significantly elevated by miR-431 repression (Fig. 5D and E). Additionally, the levels of
AKAP1 mRNA were upregulated in SNU-387 and Huh7 cells transfected
with pc-AKAP1 compared with control pc-NC (Fig. S5). Furthermore, the mRNA and protein
levels of AKAP1 were signally increased in SNU-387 and Huh7 cells
following hypoxia (Fig. 5F and G).
In summary, AKAP1 served as a target for miR-431 in HCC cells.
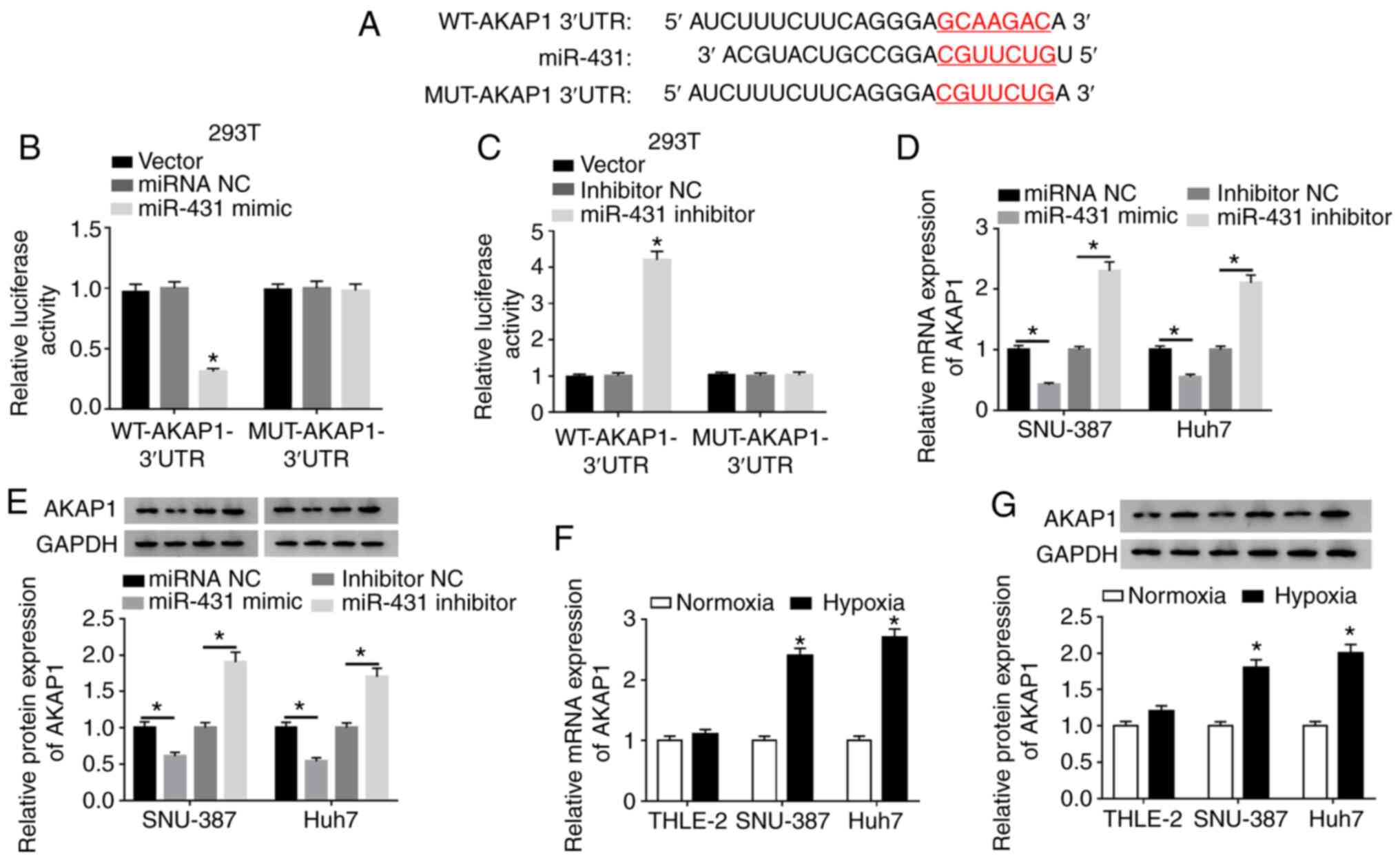 | Figure 5.AKAP1 was a target of miR-431 in
hepatocellular cancer cells. (A) The binding sites between miR-431
and AKAP1 were predicted using the Starbase database.
Dual-luciferase reporter assays were conducted for the evaluation
of the luciferase activity in 293T cells contransfected with WT or
MUT reporter vectors of AKAP1 and (B) miRNA NC, miR-431 mimic, (C)
inhibitor NC or miR-431 inhibitor. Effect of miR-431 on expression
of AKAP1 (D) mRNA and (E) protein of SNU-387 and Huh7 cells was
analyzed by RT-qPCR and western blot analysis, respectively. Under
hypoxic conditions, the (F) mRNA and (G) protein levels of AKAP1 in
SNU-387 and Huh7 cells were detected by RT-qPCR and western blot
analysis, respectively. *P<0.05. AKAP1, A-kinase anchor protein
1; miR, microRNA; WT, wild-type; MUT, mutant; NC, negative control;
RT-qPCR, reverse transcription-quantitative PCR; UTR, untranslated
region. |
AKAP1 overexpression abolishes miR-431
mimic-mediated influence on viability, apoptosis and glycolysis of
HCC following hypoxia treatment
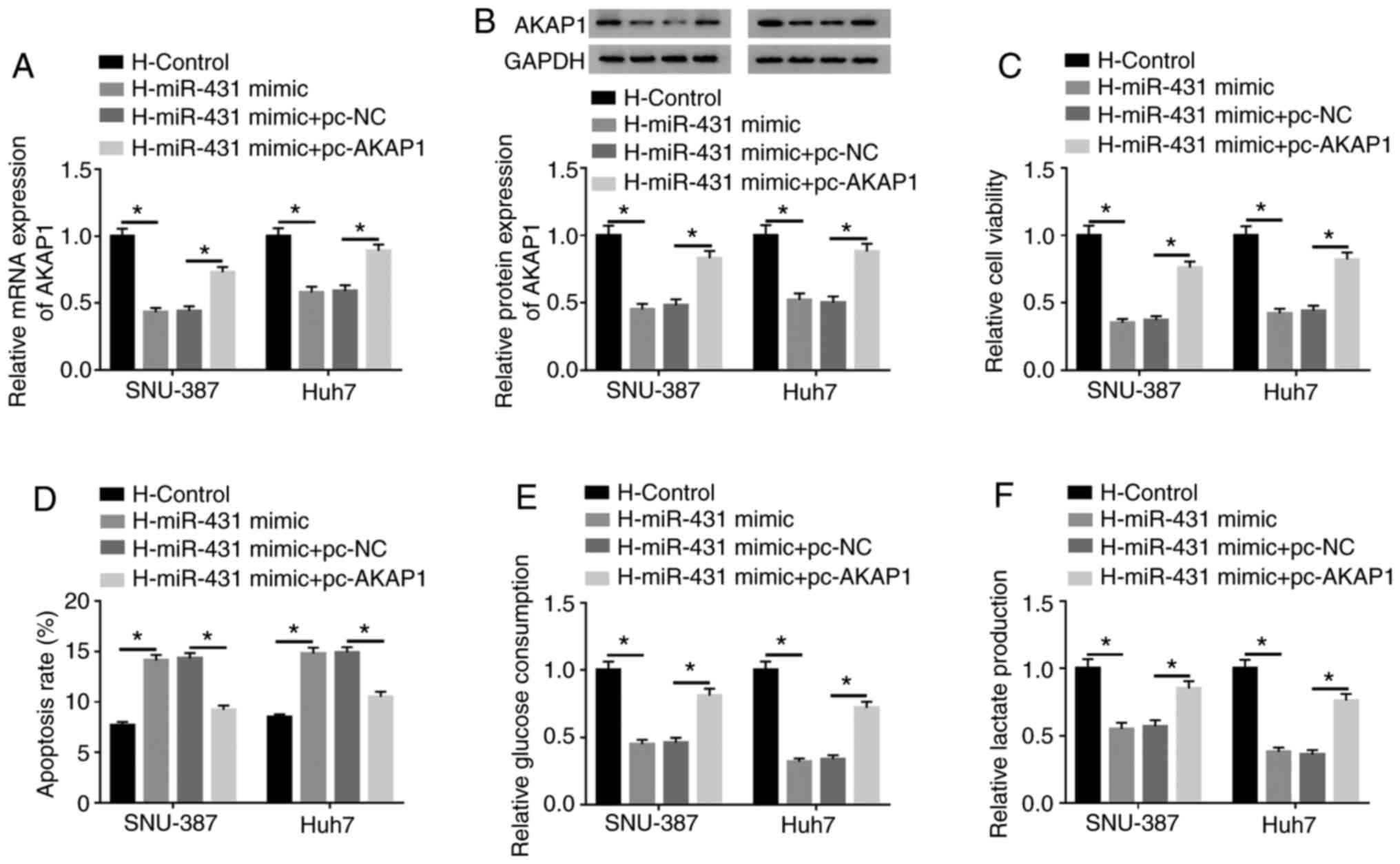
Considering miR-431 targeted AKAP1 in HCC cells,
whether miR-431 regulated the viability, apoptosis and glycolysis
of HCC via AKAP1 following hypoxia treatment was investigated. The
results presented that miR-431 overexpression reversed the
upregulation of AKAP1 mRNA (Fig. 6A)
and protein (Fig. 6B) in
hypoxia-treated SNU-387 and Huh7 cells. Furthermore, AKAP1
overexpression significantly abolished this effect. Increased
miR-431 expression inhibited cell viability in hypoxia-disposed
SNU-387 and Huh7 cells and this repression was overturned by AKAP1
overexpression (Fig. 6C). The
results of the flow cytometry assays revealed an elevated apoptosis
rate in hypoxia-treated SNU-387 and Huh7 cells caused by increased
miR-431 expression was reversed by the introduction of AKAP1
(Fig. 6D). Additionally, the
inhibitory impacts of miR-431 upregulation on glucose consumption
and lactate production in hypoxia-disposed SNU-387 and Huh7 cells
were restored by AKAP1 elevation (Fig.
6E and F). In summary, these results indicated that miR-431
mediated cell viability, apoptosis and glycolysis through AKAP1 in
hypoxia-disposed HCC cells.
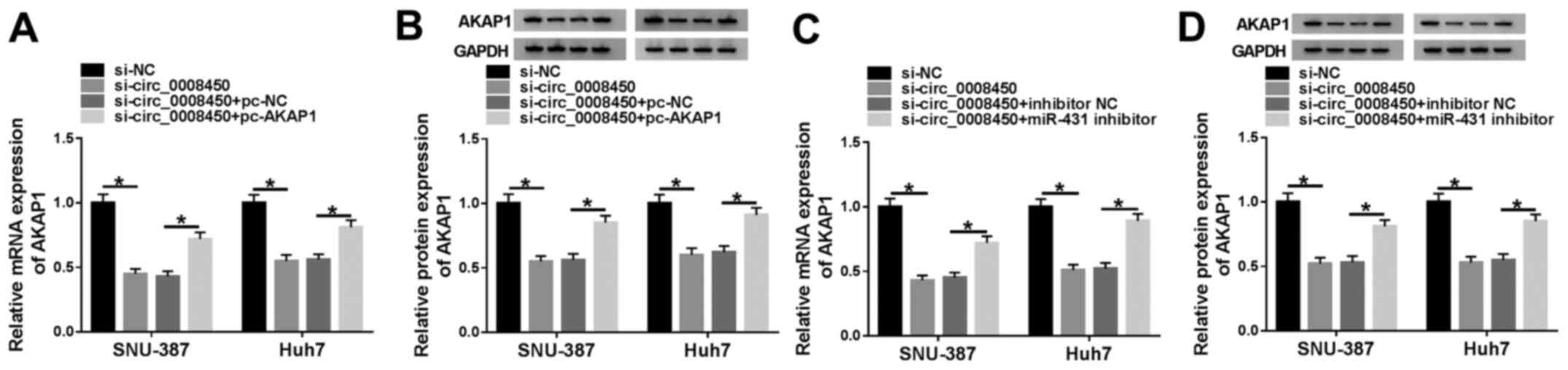
AKAP1 is modulated by circ_0008450 by
sponging miR-431
Considering miR-431 was a target of circ_0008450 and
miR-431 targeted AKAP1 in SNU-387 and Huh7 cells, whether
circ_0008450 regulated the expression of AKAP1 via miR-431 in
SNU-387 and Huh7 cells was explored. The results demonstrated that
the mRNA (Fig. 7A) and protein
(Fig. 7B) levels of AKAP1 were
decreased following the downregulation of circ_0008450 in SNU-387
and Huh7 cells, and that this effect was reversed by AKAP1
overexpression. Moreover, miR-431 inhibitor abolished the
inhibitory influence of circ_0008450 silencing on the levels of
AKAP1 mRNA (Fig. 7C) and protein
(Fig. 7D) of SNU-387 and Huh7 cells.
These data indicated that circ_0008450 regulated the expression of
AKAP1 via miR-431 in SNU-387 and Huh7 cells.
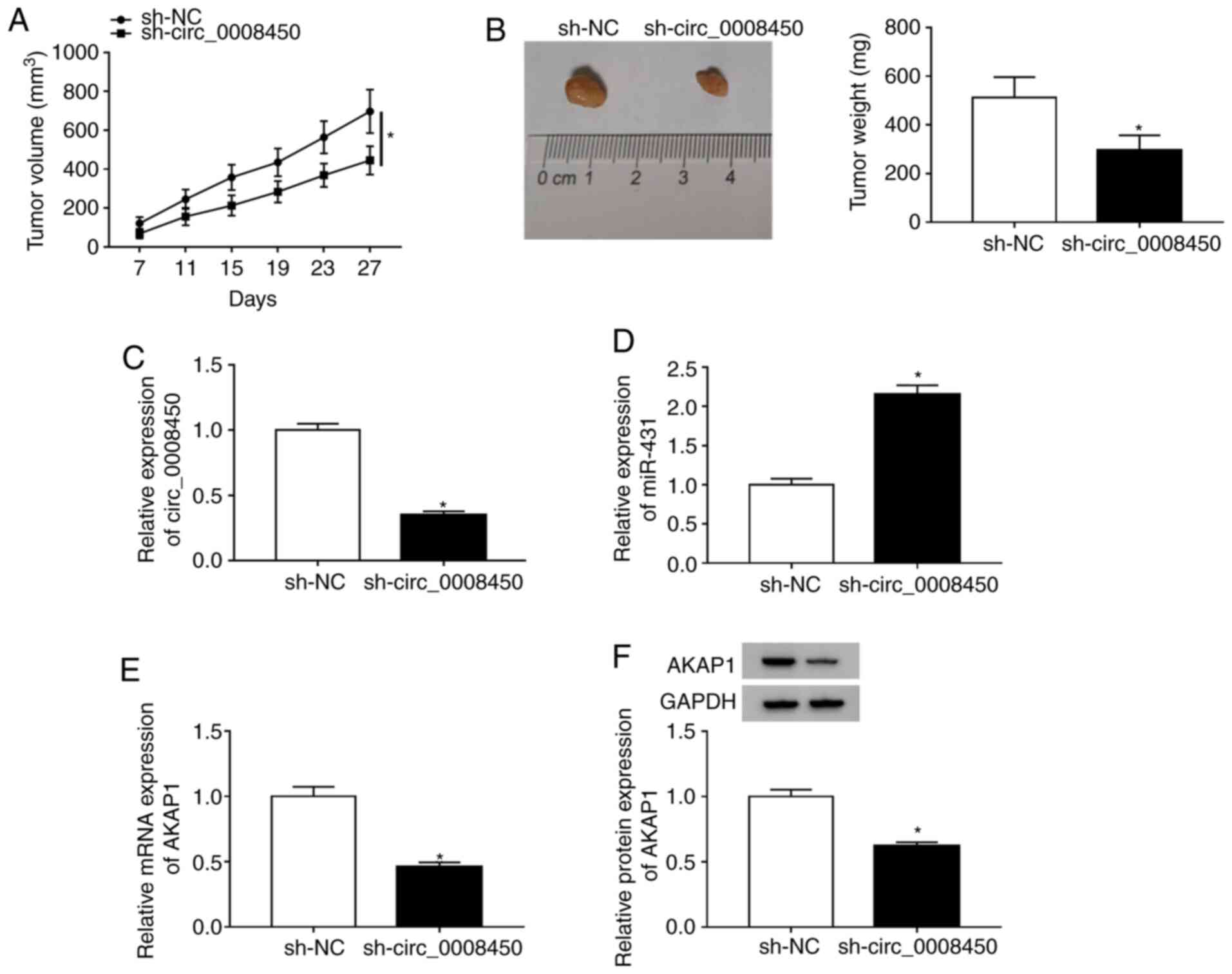
Circ_0008450 depletion reduced HCC
growth
To explore the effect of circ_0008450 on HCC tumor
growth in vivo, xenograft assay in nude mice was conducted.
The results presented that circ_0008450 expression was reduced in
Huh7 cells after transfected with sh-circ_0008450 (Fig. S6). Moreover, tumor volumes (Fig. 8A) and weights (Fig. 8B) were reduced in the sh-circ_0008450
group compared to the sh-NC group. Furthermore, circ_0008450
expression was significantly decreased (Fig. 8C) and miR-431expression significantly
increased (Fig. 8D) in the
sh-circ_0008450 group compared with the sh-NC group. Additionally,
AKAP1 mRNA (Fig. 8E) and protein
(Fig. 8F) levels were significantly
decreased in the sh-circ_0008450 group compared with the control
group. Additionally, the levels of caspase-3 were elevated in mice
tumor tissues of the sh-circ_0008450 group compared with the sh-NC
group (Fig. S7). Collectively,
these data demonstrated that circ_0008450 downregulation suppressed
tumor growth in vivo.
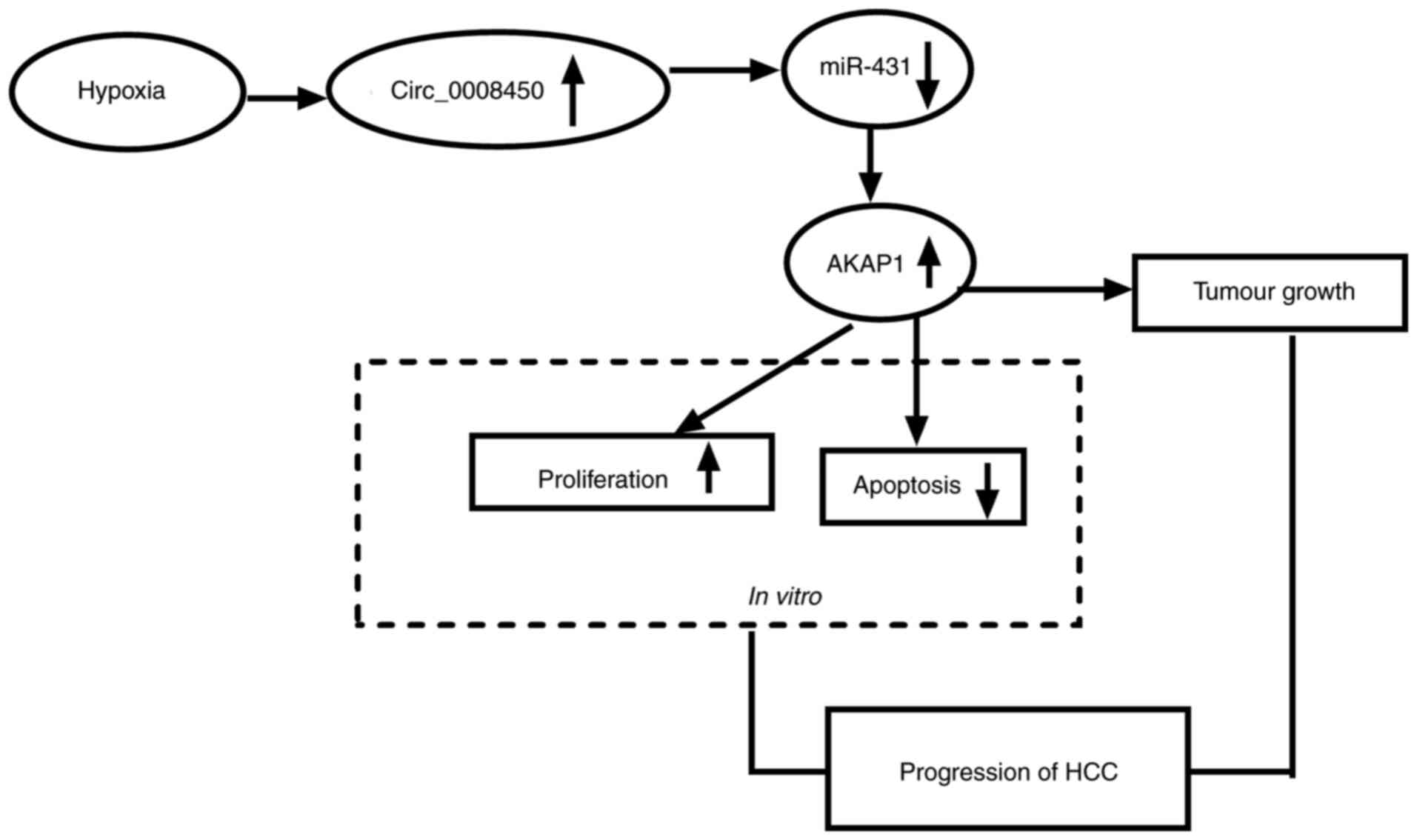
Hypoxia-induced circ_000845 regulates
HCC growth by the miR-431/AKAP1 pathway
Based on the above findings, we concluded that
hypoxia-induced circ_000845 elevated AKAP1 expression through
competitively binding to miR-431, thereby accelerating
proliferation and constraining apoptosis of HCC cells, leading to
the growth of HCC (Fig. 9).
Discussion
The rapid growth of HCC cells usually exceeds the
growth of functional blood vessels, which often causes insufficient
oxygen delivery in HCC tumors (30).
However, in order to provide sufficient energy and substances for
the anabolic metabolism of cancer cells, these cells alter their
glucose metabolism from oxidative to glycolysis to adapt to hypoxic
stress (31). In recent years,
circRNAs have been reported to be associated with the progression
of hypoxia-mediated tumors. For instance, hypoxia-induced
circDENND2A accelerated cell malignancy in glioma cells by sponging
miR-625-5p (15). Additionally,
circ_0000977 repression elevated the killing effect of natural
killer cells in pancreatic cancer cells under hypoxic conditions
(32). In the current study,
circ_0008450 was upregulated in HCC tissues and hypoxia-treated HCC
cells. High circ_0008450 expression was associated with tumor size,
TNM stage, lymphatic metastasis and distant metastasis in patients
with HCC. Moreover, circulating circ_0008450 had a high diagnostic
value. These results indicated that hsa_circ_0084927 may be a novel
biomarker for the diagnosis of HCC. Furthermore, circ_0008450
inhibition reduced tumor growth in vivo and reversed its
effects on viability and glycolysis, and promoted the apoptosis of
hypoxia-disposed HCC cells in vitro. Zhang et al
(17) revealed that circ_0008450
downregulation induced cell apoptosis and repressed cell invasion,
proliferation and migration in HCC cells. Moreover, Lin et
al (33) indicated that
circ_0008450 knockdown reversed invasion, proliferation and
migration of HCC cells. Furthermore, circ_0008450 was revealed to
be an oncogene in nasopharyngeal cancer (16). These data indicated that circ_0008450
acted as a potential diagnostic biomarker for HCC and
hypoxia-induced circ_0008450 served a cancerogenic role in HCC.
It has been reported that circ_0008450 regulated
tumor progression by acting as a sponge for miRNAs (16,33). For
instance, hsa_circ_0008450 knockdown impeded the advancement of
hepatocellular carcinoma by downregulating enhancer of zeste
homolog 2 by sponging miR-214-3p (33). Furthermore, miR-431 was revealed to
act as an anti-tumor gene in a variety of cancers. For example, a
previous report demonstrated that miR-431 inhibited metastasis and
proliferation of lung cancer cells by repressing DEAD box protein
expression 5 (22). Another study
revealed that silenced miR-431 expression accelerated the invasion
of papillary thyroid cancer and was associated with lymph node
metastasis (34). Previous studies
have demonstrated that miR-431 reversed cell invasion and invasion
in HCC cells (23,35). In the current study, miR-431
expression was reduced in HCC cells treated with hypoxia and acted
as a target for circ_0008450. Additionally, miR-431 suppression
reversed circ_0008450 silencing-mediated effects on the viability,
apoptosis and glycolysis of hypoxia-treated HCC cells. These
results indicated that hypoxia-induced circ_0008450 modulated the
malignant behavior of HCC cells through miR-431 under hypoxic
conditions.
As a scaffold protein, AKAP1 has been reported to
serve a vital role in controlling mitochondrial function (36). A previous report reported that AKAP1
supported the mTOR pathway and increased the growth of tumor cells
(25). Increased AKAP1 expression
was associated portal vein thrombosis, tumor size and
tumor-mode-metastasis stage of patients with HCC and AKAP1 may act
as a potential prognostic biomarker for the prediction of survival
in patients with HCC followed by radical resection (27). In the current study, AKAP1 was
regulated by circ_0008450 via miR-431. AKAP1 elevation reversed the
effects of miR-431 on the viability, apoptosis and glycolysis of
HCC cells following hypoxia treatment. Therefore, circ_0008450
contributed to HCC progression by upregulating AKAP1 and sponging
miR-431 under hypoxia conditions. Unfortunately, only molecular
biology research was conducted in the current study and clinical
analysis was not performed on the samples. This can be investigated
in future studies.
In conclusion, the current study demonstrated that
hypoxia-induced circ_0008450 mediated the malignant behavior of HCC
cells under hypoxia through the miR-431/AKAP1 axis and circ_0008450
may act as a possible therapeutic target for HCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request
Authors' contributions
QD participated in study design, methodology and
data interpretation, and drafted the manuscript. JH performed data
collection, analysis and statistical analysis. SG participated in
the methodology and data interpretation. SZ performed data
interpretation and methodology. YP interpreted data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
The current study was authorized by the Ethics
Committee of Lanzhou University Second Hospital, Lanzhou,
China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular cancer
|
|
AKAP1
|
A-kinase anchor protein 1
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Forner A, Llovet JM and Bruix J:
Hepatocellular carcinoma. Lancet. 379:1245–1255. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
El-Serag HB and Rudolph KL: Hepatocellular
carcinoma: Epidemiology and molecular carcinogenesis.
Gastroenterology. 132:2557–2576. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhu P, Ning Y, Yao L, Chen M and Xu C: The
proliferation, apoptosis, invasion of endothelial-like epithelial
ovarian cancer cells induced by hypoxia. J Exp Clin Cancer Res.
29:1242010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Du C, Weng X, Hu W, Lv Z, Xiao H, Ding C,
Gyabaah OA, Xie H, Zhou L, Wu J, et al: Hypoxia-inducible MiR-182
promotes angiogenesis by targeting RASA1 in hepatocellular
carcinoma. J Exp Clin Cancer Res. 34:672015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rankin EB and Giaccia AJ: Hypoxic control
of metastasis. Science. 352:175–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Su Y, Yang W, Jiang N, Shi J, Chen L,
Zhong G, Bi J, Dong W, Wang Q, Wang C, et al: Hypoxia-elevated
circELP3 contributes to bladder cancer progression and cisplatin
resistance. Int J Biol Sci. 15:441–452. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing. Mol
Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ebbesen KK, Hansen TB and Kjems J:
Insights into circular RNA biology. RNA Biol. 14:1035–1045. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao ZJ and Shen J: Circular RNA
participates in the carcinogenesis and the malignant behavior of
cancer. RNA Biol. 14:514–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang R, Zhang S, Chen X, Li N, Li J, Jia
R, Pan Y and Liang H: CircNT5E acts as a sponge of miR-422a to
promote glioblastoma tumorigenesis. Cancer Res. 78:4812–4825. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Qiu M, Xia W, Chen R, Wang S, Xu Y, Ma Z,
Xu W, Zhang E, Wang J, Fang T, et al: The circular RNA circPRKCI
promotes tumor growth in lung adenocarcinoma. Cancer Res.
78:2839–2851. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ren S, Liu J, Feng Y, Li Z, He L, Li L,
Cao X, Wang Z and Zhang Y: Knockdown of circDENND4C inhibits
glycolysis, migration and invasion by up-regulating miR-200b/c in
breast cancer under hypoxia. J Exp Clin Cancer Res. 38:3882019.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei Y, Zhang Y, Meng Q, Cui L and Xu C:
Hypoxia-induced circular RNA has_circRNA_403658 promotes bladder
cancer cell growth through activation of LDHA. Am J Transl Res.
11:6838–6849. 2019.PubMed/NCBI
|
|
15
|
Su H, Zou D, Sun Y and Dai Y:
Hypoxia-associated circDENND2A promotes glioma aggressiveness by
sponging miR-625-5p. Cell Mol Biol Lett. 24:242019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wei H, Liu D, Sun J, Mao Y, Zhao L, Zhu W,
Xu G and Gao Z: Circular RNA circ_0008450 upregulates CXCL9
expression by targeting miR-577 to regulate cell proliferation and
invasion in nasopharyngeal carcinoma. Exp Mol Pathol.
110:1042882019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang J, Chang Y, Xu L and Qin L: Elevated
expression of circular RNA circ_0008450 predicts dismal prognosis
in hepatocellular carcinoma and regulates cell proliferation,
apoptosis, and invasion via sponging miR-548p. J Cell Biochem.
120:9487–9494. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang J, Zhu H, Jin Y and Song Y: MiR-431
inhibits cell proliferation and induces cell apoptosis by targeting
CDK14 in pancreatic cancer. Eur Rev Med Pharmacol Sci.
22:4493–4499. 2018.PubMed/NCBI
|
|
21
|
Su WB and Liu ZY: MiR-431 inhibits
colorectal cancer cell invasion via repressing CUL4B. Eur Rev Med
Pharmacol Sci. 22:3047–3052. 2018.PubMed/NCBI
|
|
22
|
Xu CM, Chen LX, Gao F, Zhu MF, Dai Y, Xu Y
and Qian WX: MiR-431 suppresses proliferation and metastasis of
lung cancer via down-regulating DDX5. Eur Rev Med Pharmacol Sci.
23:699–707. 2019.PubMed/NCBI
|
|
23
|
Sun K, Zeng T, Huang D, Liu Z, Huang S,
Liu J and Qu Z: MicroRNA-431 inhibits migration and invasion of
hepatocellular carcinoma cells by targeting the ZEB1-mediated
epithelial-mensenchymal transition. FEBS Open Bio. 5:900–907. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Feliciello A, Gottesman ME and Avvedimento
EV: The biological functions of A-kinase anchor proteins. J Mol
Biol. 308:99–114. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rinaldi L, Sepe M, Delle Donne R, Conte K,
Arcella A, Borzacchiello D, Amente S, De Vita F, Porpora M, Garbi
C, et al: Mitochondrial AKAP1 supports mTOR pathway and tumor
growth. Cell Death Dis. 8:e28422017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Schiattarella GG, Cattaneo F, Carrizzo A,
Paolillo R, Boccella N, Ambrosio M, Damato A, Pironti G, Franzone
A, Russo G, et al: Akap1 regulates vascular function and
endothelial cells behavior. Hypertension. 71:507–517. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yu J, Zhang Y, Zhou D, Wu J and Luo R:
Higher expression of A-kinase anchoring-protein 1 predicts poor
prognosis in human hepatocellular carcinoma. Oncol Lett.
16:131–136. 2018.PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang ZJ, Shen QH, Chen HY, Yang Z, Shuai
MQ and Zheng SS: Galectin-1 gene silencing inhibits the activation
and proliferation but induces the apoptosis of hepatic stellate
cells from mice with liver fibrosis. Int J Mol Med. 43:103–116.
2019.PubMed/NCBI
|
|
30
|
Rankin EB, Nam JM and Giaccia AJ: Hypoxia:
Signaling the metastatic cascade. Trends Cancer. 2:295–304. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang X, Li Y, Ma Y, Yang L, Wang T, Meng
X, Zong Z, Sun X, Hua X and Li H: Yes-associated protein (YAP)
binds to HIF-1α and sustains HIF-1α protein stability to promote
hepatocellular carcinoma cell glycolysis under hypoxic stress. J
Exp Clin Cancer Res. 37:2162018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ou ZL and Luo Z: Hypoxia-induced shedding
of MICA and HIF1A-mediated immune escape of pancreatic cancer cells
from NK cells: Role of circ_0000977/miR-153 axis. RNA Biol.
16:1592–1603. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lin T, Dai Y, Guo X, Chen W, Zhao J, Cao L
and Wu Z: Silencing of hsa_circ_0008450 represses hepatocellular
carcinoma progression through regulation of microRNA-214-3p/EZH2
axis. Cancer Manag Res. 11:9133–9143. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu Y, Li L, Liu Z, Yuan Q and Lu X:
Downregulation of MiR-431 expression associated with lymph node
metastasis and promotes cell invasion in papillary thyroid
carcinoma. Cancer Biomark. 22:727–732. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li MF, Li YH, He YH, Wang Q, Zhang Y, Li
XF, Meng XM, Huang C and Li J: Emerging roles of hsa_circ_0005075
targeting miR-431 in the progress of HCC. Biomed Pharmacother.
99:848–858. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Carlucci A, Adornetto A, Scorziello A,
Viggiano D, Foca M, Cuomo O, Annunziato L, Gottesman M and
Feliciello A: Proteolysis of AKAP121 regulates mitochondrial
activity during cellular hypoxia and brain ischaemia. EMBO J.
27:1073–1084. 2008. View Article : Google Scholar : PubMed/NCBI
|