Introduction
According to the grading system developed by the
World Health Organization (WHO), gliomas are classified into grades
I–IV (1). Grade I and II gliomas are
low-grade gliomas and the prognosis of the patients is relatively
good. Grade III and IV gliomas are high-grade gliomas (HGG). The
prognosis of the patients with these types of tumors is relatively
poor (2). Diffuse astrocytoma is
classified as a WHO grade II tumor, whereas anaplastic astrocytoma
is a WHO grade III tumor. Glioblastoma is a WHO grade IV tumor
(3). In the last 30 years, the
5-year survival rate of gliomas has not significantly improved
(4). Based on this evidence, the
identification of new treatments for patients with GBM is
imperative.
Tumor necrosis factor-α (TNF-α) is a polypeptide
cytokine produced by mononuclear macrophages and is considered an
important inflammatory factor in the tumor inflammatory
microenvironment (5). Tumor necrosis
factor receptor 1 (TNFR1) is one of the receptors of TNF-α that
mediates the majority of its biological functions, such as
antiviral activity (6), apoptosis
induction (7) and activation of the
NF-κB protein (8,9). The receptor of TNF-α is a type I
transmembrane glycoprotein distributed on the surface of the normal
cell membrane (10). A previous
study has reported that TNFR1 exerts a significant influence on the
malignant progression of tumors and that it is associated with the
degree of differentiation or the depth of invasion of the tumors
(11).
Pyruvate dehydrogenase (PDH) α1 (PDHA1) is a subunit
of PDH. PDHA1 has a molecular weight of 43.3 kDa and a genomic DNA
full length of 15.92 Kbp which includes 11 exons. It is located on
the X chromosome (Xp22.1–22.2) and is also known as PDC E1α
(12). In mitochondria, PDH exists
in the form of PDH complex (PDHc) and a specific phosphorylation
site of PDHc on PDHA1 serves an important role in regulating the
activity and energy metabolism of PDHc (13). PDH kinase (PDK)1 primarily acts on
PDHA1 in PDHc. The regulation of PDHA1 phosphorylation can increase
PDHc activity (14). The normal
expression of PDHA1 is a prerequisite for the normal development of
the tricarboxylic acid (TCA) cycle and the oxidative
phosphorylation in the mitochondria (15). The function of PDHA1 is to transform
aerobic glycolysis into oxidative phosphorylation. In the absence
of oxygen, pyruvate is reduced to lactate. In the presence of
oxygen, pyruvate is oxidized by PDH to acetyl-CoA, which enters the
mitochondria and is metabolized further by the TCA cycle. When the
phosphorylation of PDHA1 is inhibited, it can inactivate PDHc
(16). Furthermore, the Warburg
effect is enhanced, and the invasiveness of tumor cells is
increased (17). Therefore, the
ability of phosphorylated (p)-PDHA1 to mediate TNFα-induced cell
migration in glioma cells is worth determining.
Materials and methods
Patients and tissue samples
A total of 80 paraffin embedded samples were
obtained from the Affiliated Hospital of Nantong University between
January 2013 and January 2017. The present study was approved by
the Human Research Ethics Committee of Nantong University, China.
The patients were recruited based on: i) Postoperative pathological
diagnosis; and ii) Magnetic resonance imaging data. Moreover, all
glioma specimens were obtained from patients who had not previously
received radiotherapy or chemotherapy. The sections of the glioma
samples were analyzed by two professionals from the Department of
Pathology, Affiliated Hospital of Nantong University according to
the WHO (2007) tumor grading standards (18). The grading system adopted the
double-blind method. The follow-up data were collected by telephone
visit and concomitant household registration data visit (follow-up
time ≤60 months). The follow-up rate was 93.7% (75/80).
Cell lines
The cell lines used were purchased from The Cell
Bank of Type Culture Collection of the Chinese Academy of Sciences.
Specifically, U251, U87 (glioblastoma of unknown origin), A172, H4
and SHG44 glioma cell lines, and 293T cells were used in the
present study. U251, U87, A172, H4 and SHG44 glioma cell lines were
cultured in DMEM medium (Gibco; Thermo Fisher Scientific, Inc.).
293T cells were cultured in RPMI 1640 medium (Sigma-Aldrich; Merck
KGaA). In total, 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc.) was added into all culture media. Cell culture
was performed at 37°C in a 5% CO2 incubator.
Antibodies
The primary antibodies used in the present study
included the following: PDHA1 (1:500; cat. no. 377092 Santa Cruz
Biotechnology, Inc.), p-PDHA1 (1:1,000; no. ab115343; Abcam), TNFR1
(1:1,000; cat. no. ab68160; Abcam), MMP2 (1:500; cat. no. 13595;
Santa Cruz Biotechnology, Inc.), MMP14 (1:500; cat. no. 373908;
Santa Cruz Biotechnology, Inc.), E-cadherin (1:1,000; cat. no.
14472; Cell Signaling Technology, Inc.), GAPDH (1:5,000; cat. no.
ab8245; Abcam).
Drug treatment
DCA stock solutions of 1/10 mM (Sigma-Aldrich; Merck
KGaA) were diluted in PBS for in vitro experiments. Prior to
western blotting experiments, DCA was diluted into 5, 10 and 20 mM
concentrations to stimulate U251 cells for 24 h at 37°C in a 5%
CO2 incubator. Prior to migration and wound-healing
experiments, the concentration of 20 mM DCA was chosen to stimulate
U251 cell for 24 and 48 h at 37°C in a 5% CO2
incubator.
Cell protein extraction
Cells at 0, 4, 8, 12 and 24 h were washed three
times with PBS at room temperature for 5 min. RIPA cell protein
lysate (Thermo Fisher Scientific, Inc.; 400 µl) was added to the
cells on ice for 40 min. Samples were then centrifuged at 4°C for
15 min at 12,000 × g.
Cell proliferation assays
The U251 glioma cells were seeded in 96-well plates
at a density of 2×106 cells/well for 24 h and
subsequently the Cell Counting Kit (CCK)-8 reagent (Dojindo
Molecular Technologies, Inc.) was added to each well for 4 h,
taking care to avoid light. The absorbance was measured at 450 nm
by a microplate reader (Bio-Rad Laboratories, Inc.).
Western blot analysis
Proteins were extracted from the transfected cells
using RIPA buffer on ice. Protein extracts were mixed with 2X
loading buffer. The protein was quantified using BCA determination
method. Proteins (60 µg/lane) were separated by 10% SDS-PAGE. The
protein samples were transferred to polyvinylidene fluoride (PVDF)
membranes following sodium dodecyl sulfate-polyacrylamide gel
electrophoresis. The PVDF membrane was blocked with skimmed milk at
room temperature for 2 h, before the aforementioned primary
antibodies were added at 4°C overnight. Membranes were then
incubation with horseradish peroxidase conjugated goat anti-rabbit
secondary antibody (1:5,000; cat. no. ab7090; Abdam) at room
temperature for 2 h. Pierce™ Electro-chemiluminescence (Thermo
Fisher Scientific, Inc.) was used to detect protein expression. The
ImageJ software (v1.50; National Institutes of health) was used to
conduct densitometric analysis.
Immunofluorescence staining of
cytoskeleton
U251MG cells were cultured in a 6-well plate. After
4 h growth, the cells were collected. The cells were fixed with 4%
paraformaldehyde for 1 h at 37°C. Subsequently, they were treated
with 0.2% Triton X-100 and blocked with 1% bovine serum albumin
(Sigma-Aldrich-Merck KGaA) at room temperature for 30 min. Then
cells were immunolabeled with anti-vinculin (1:200; cat. no.
00162950; Sigma-Aldrich; Merck KGaA) at 37°C for 1 h followed by
FITC-anti-mouse (1:200; cat. no. F4516; Sigma-Aldrich; Merck KGaA)
at room temperature for 2 h. Subsequently,
phalloidin-tetramethylrhodamine (1:1,000, Abcam) was added to the
cells and placed in a 37°C incubator for 1 h. Finally, the nuclei
were stained with 4′,6-diamidino-2-phenylindole at room temperature
for 1 h. The cytoskeletal structure was detected by an Olympus
fluorescence microscope (magnification, ×1,000).
Wound healing assay
The U251 glioma cells were cultured in a 6-well
plate according to different experimental conditions. A line was
drawn in the fused cell layer (5×105 cells/well) with a
100 µl micropipette tip. The cells were washed three times with PBS
and grown in serum-free DMEM, before being incubated at 37°C for 48
h. Then, the cells were imaged at 0, 24 and 48 h using an inverted
Leica phase contrast microscope (magnification, ×200). The wound
closure rate was calculated using ImageJ (version 1.6; National
Institutes of Health) and relative wound closure was
determined.
Cell invasion assay
A total of 200 µl U251 cells (5×105
cells) was added in serum-free DMEM to the upper layer of each
chamber and 600 µl DMEM containing 10% fetal bovine serum was added
to the lower layer of each chamber. The cells were cultured at 37°C
in a 5% CO2 incubator for 24 h. At appropriate time
points, the cells in the upper chamber were removed with a cotton
swab and the migrated cells were fixed and stained with 4% crystal
violet at room temperature for 5 min. The migrated cells were
counted using an inverted Leica phase contrast microscope
(magnification, ×100).
Immunohistochemical analysis
The tissue slices were put into the oven for 8 h and
subsequently into gradient ethanol and xylene. The sections were
boiled in citrate solution to retrieve the antigens and washed with
PBS three times. The endogenous peroxidase activity was blocked
with 0.3% hydrogen peroxide at room temperature for 1 h. The
primary antibody (p-PDHA1; 1:1,000; cat. no. ab115343; Abcam),
E-cadherin (1:1,000; cat. no. 14472; Cell Signaling Technology,
Inc.) was added to the sections at 4°C for overnight incubation.
The following day, the sections were washed with PBS and the
secondary antibody was added at room temperature for 2 h. The
slices were stained with 3,3-diaminobenzidine and hematoxylin at
room temperature for 5 min. The sections were differentiated with
hydrochloric acid alcohol and dehydrated with gradient alcohol.
Finally, the slices were sealed with neutral balsam. The sections
were examined by two professional pathologists who did not have
previous relevant information regarding the samples. The staining
intensity of the sections was classified as follows: 0 (no
staining), 1 (weak staining), 2 (moderate staining) and 3 (strong
staining). The proportion of cell staining corresponded to the
following scores: 0, <1%; 1, 1–20%; 2, 21–50%; 3, 51–75%; and 4,
>75%. PBS was used as the negative control and the sections that
were stained positive were used as the positive control. Following
the non-specific staining of the tumor margin, the necrotic area
and adjacent brain tissue area were excluded. The scores of the
staining intensity and positive cell proportion were added.
Cell transfection
The small interfering (si)RNA sequences used for
PDHA1 knockdown were synthesized by the Shanghai GeneChem Co., Ltd.
The sequences were the following: siPDHA1-1,
5′-CAATCAGTGGATCAAGTT-3′, siPDHA1-2, 5′-AATGGAGTTGAAAGCAGAT-3′,
siPDHA1-3, 5′-TGGTAGCATCCCGTAATTT-3′, siPDHA1-4,
5′-AGAAATTCTCGCAGAGCTT-3′. The negative control (NC) siRNA sequence
(siNC) was the following: 5′-TTCTCCGAACGTGTCACGT-3′. Subsequent
experiments were conducted at 48 h post-transfection. The U251
glioma cells were grown at a density of 80% and transfected with
100 nM siRNA targeting specific genes using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific), according to the manufacturer's instructions.
Statistical analysis
The data were analyzed by the SPSS 19.0 software
(IBM Corp.). The association between PDHA1 and Ki-67 was evaluated
by the Spearman's correlation coefficient. The χ2 test
was used to compare the clinicopathological parameters and the
levels of the p-PDHA1 protein in 80 patients. The Kaplan-Meier
(K-M) by SPSS (v.22.0; IBM Corp.) survival curve was used to
analyze the survival rate of the patients. The log rank test was
used to compare the survival curve data. The differences between
two groups were analyzed using the Student's paired t-test. The
differences between multiple groups were analyzed by one-way ANOVA
followed by Tukey's post hoc test. Cox proportional analysis was
used to analyze the independent effect of p-PDHA1 on the prognosis
of patients. All experiments were repeated at least three times.
P<0.05 was considered to indicate a statistically significant
difference.
Results
p-PDHA1 levels in different grades of
human glioma tissues
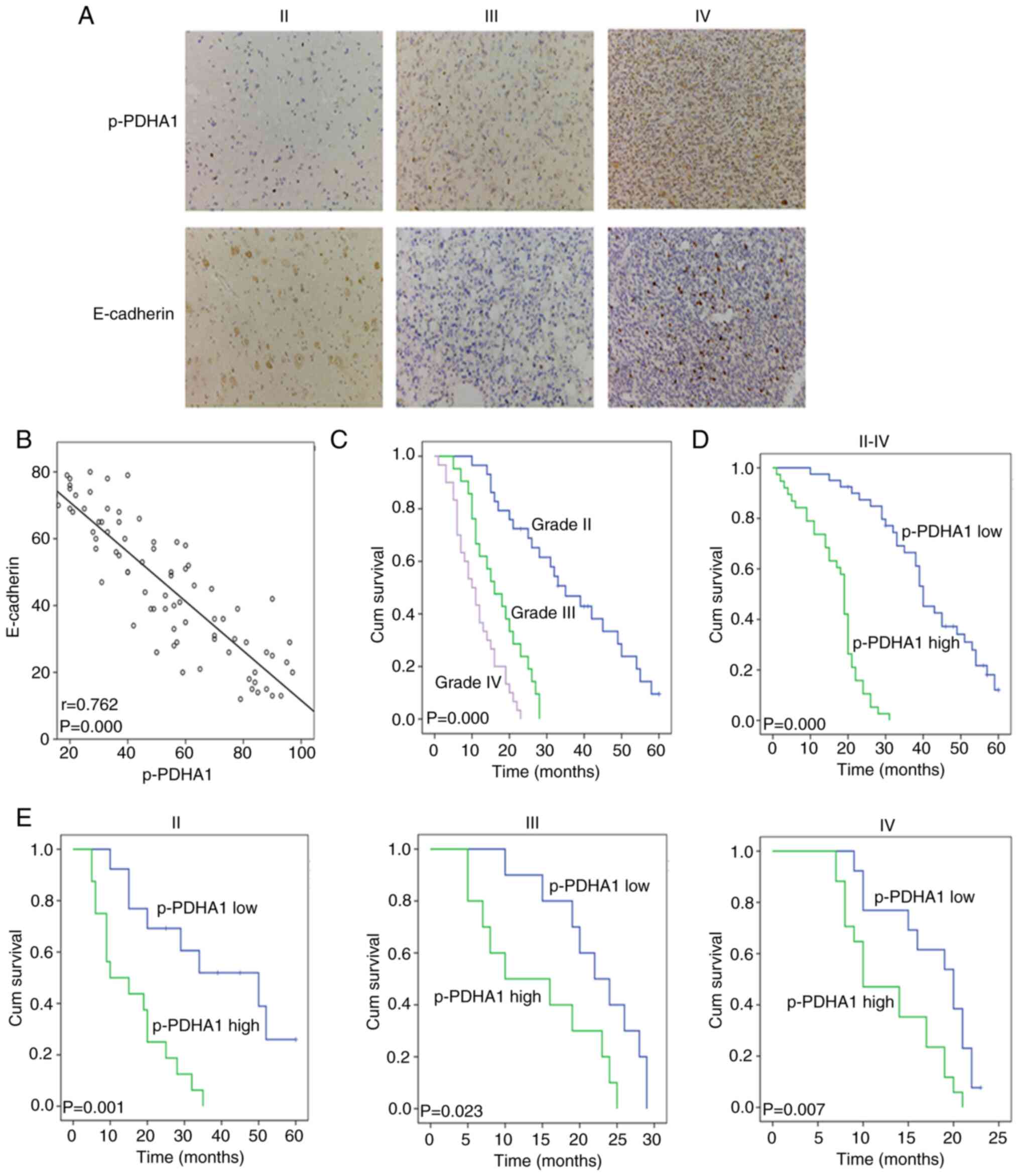
The protein levels of p-PDHA1 and E-cadherin in
tissues were detected by immunohistochemistry (Fig. 1A). The staining results of 80
patients with glioma were assessed using the Spearman rank test.
The data demonstrated that the protein levels of p-PDHA1 correlated
negatively with E-cadherin (Fig. 1B;
P<0.001). The clinical data of 80 patients with glioma were
analyzed, including 29 patients with grade II glioma, 20 patients
with grade III glioma and 31 patients with grade IV glioma. The
χ2 test indicated that p-PDHA1 level significantly
differed between the different grades of glioma (P=0.019; Table I). The K-M survival curve and
log-rank test were used to analyze the survival between different
WHO grades and p-PDHA1 level in patients with glioma (Fig. 1C; P=0.000). Subsequently, the
association of the survival time between high and low p-PDHA1 level
was analyzed using the K-M curve and the log rank test (Fig. 1D; P=0.000). Patients were then
divided into high and low p-PDHA1 expression groups. The high
expression group was associated with the different grades of glioma
(Table I). It was revealed that the
survival time of patients with high p-PDHA1 level in glioma was
lower than that noted in low p-PDHA1 group. The K-M survival curves
of high and low p-PDHA1 level in the different glioma grades
indicated that low p-PDHA1 level was associated with longer
survival time of patients with glioma in tissues of all grades
(Fig. 1E; P<0.05). Multivariate
survival analysis demonstrated that p-PDHA1 level was an
independent prognostic factor for glioma (P=0.003; Table II).
 | Table I.Protein level of p-PDHA1 and
clinicopathological characteristics in 80 cases of glioma. |
Table I.
Protein level of p-PDHA1 and
clinicopathological characteristics in 80 cases of glioma.
|
|
| p-PDHA1
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Variables | Total, n | Low | High |
χ2-value | P-value |
|---|
| Age, years |
|
<40 | 38 | 23 | 15 | 0.317 | 0.573 |
|
≥40 | 42 | 29 | 13 |
|
|
| Sex |
|
Female | 43 | 18 | 25 | 0.747 | 0.387 |
|
Male | 37 | 20 | 17 |
|
|
| Tumor location |
|
Frontal | 20 | 7 | 13 | 1.312 | 0.726 |
|
Parietal | 17 | 9 | 8 |
|
|
|
Occipital | 32 | 14 | 18 |
|
|
|
Temporal | 11 | 5 | 6 |
|
|
| Tumor size, cm |
|
<4 | 35 | 20 | 15 | 0.722 | 0.395 |
| ≥4 | 45 | 31 | 14 |
|
|
| WHO grade |
| II | 29 | 10 | 19 | 7.885 | 0.019 |
|
III | 20 | 6 | 14 |
|
|
| IV | 31 | 20 | 11 |
|
|
| Extent of
resection |
|
Biopsy | 14 | 8 | 6 | 1.564 | 0.458 |
| Total
resection | 36 | 15 | 21 |
|
|
|
Subtotal resection | 30 | 17 | 13 |
|
|
| Table II.Contribution of various potential
prognostic factors to survival by Cox regression analysis on 80
glioma specimens. |
Table II.
Contribution of various potential
prognostic factors to survival by Cox regression analysis on 80
glioma specimens.
|
Characteristics | Hazard ratio | 95% CI | P-value |
|---|
| Age |
1.089 | 0.636–1.865 | 0.757 |
| Sex |
1.290 | 0.760–2.189 | 0.345 |
| Tumor location |
0.822 | 0.639–1.057 | 0.127 |
| Tumor size |
1.340 | 0.800–2.244 | 0.266 |
| WHO Grade | 13.661 | 7.046–26.488 | 0.000 |
| Extent of
resection |
0.944 | 0.688–1.295 | 0.721 |
| p-PDHA1 |
2.432 | 1.344–4.400 | 0.003 |
Protein level of p-PDHA1 in glioma
cells
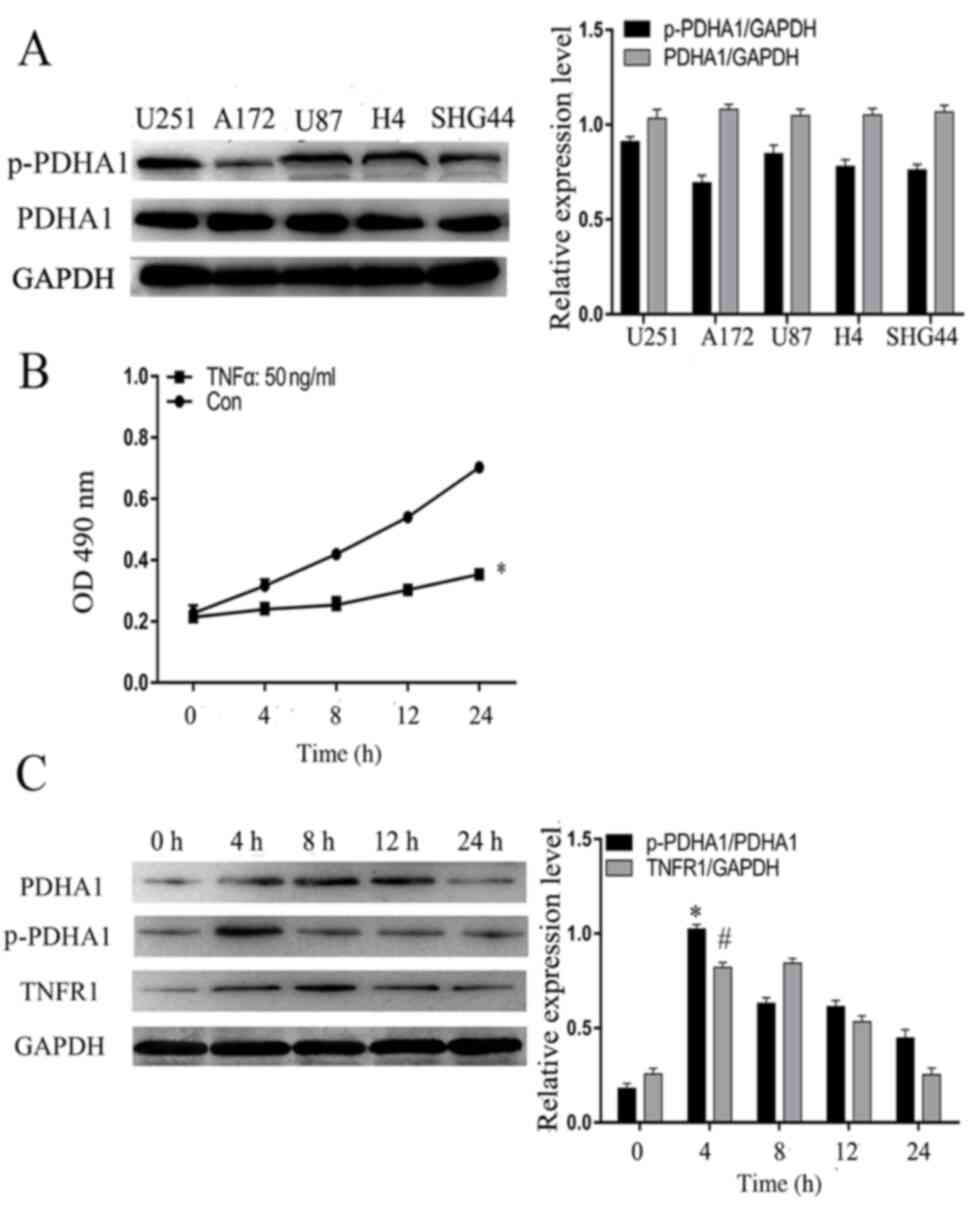
Initially, five glioma cell lines were selected as
follows: H4, A172, SHG44, U251 and U87. Western blot analysis
indicated that the levels of p-PDHA1 were increased in U251 glioma
cells compared with total protein (Fig.
2A). Subsequently, TNF-α (50 ng/ml) was used to stimulate U251
glioma cells and the absorbance was detected at 0, 4, 8, 12 and 24
h at 490 nm. CCK-8 analysis indicated that following the
stimulation of TNF-α, compared with 0 h, the proliferation of
glioma cells was increased (Fig.
2B). Furthermore, U251 glioma cells were treated with TNF-α (50
ng/ml) and the cell protein was collected at 0, 4, 8, 12 and 24 h,
in order to assess the highest protein level time point of p-PDHA1.
Western blot analysis indicated that p-PDHA1 and TNFR1 levels were
increased at 4 h in U251 glioma cells compared with those noted at
0 h (Fig. 2C).
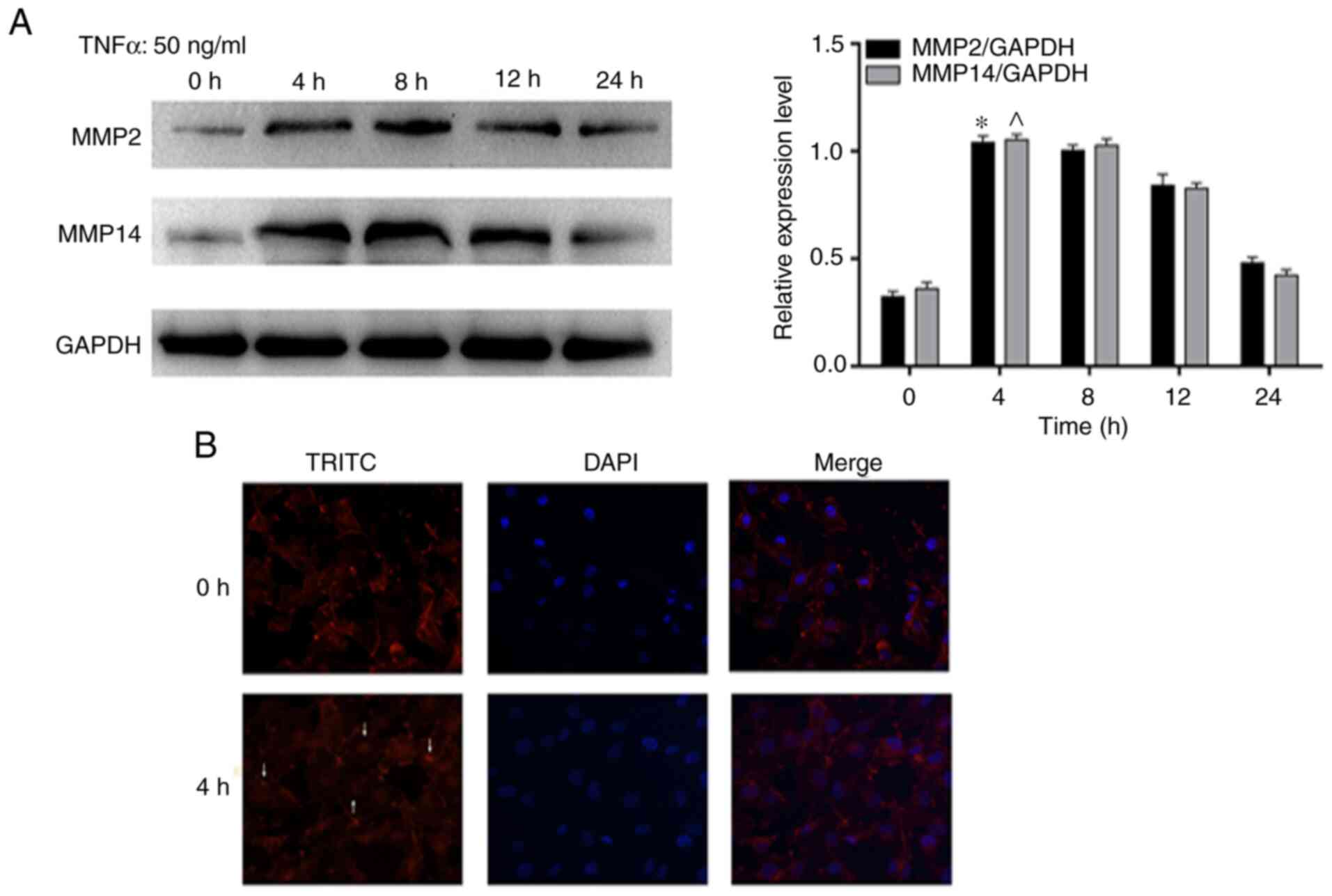
p-PDHA1 promotes glioma cell migration
and invasion following TNF-a stimulation
In order to further investigate whether p-PDHA1
exhibited migratory effects on glioma cells, the effects of TNF-α
stimulation on specific migratory markers were assessed in U251
glioma cells. The results indicated that the protein levels of MMP2
and MMP14 in glioma cells were significantly increased at 4 and 8 h
compared with 0 h. (Fig. 3A). In
addition, these results were further verified by immunofluorescence
staining of the cytoskeleton. Immunofluorescence microscopy
demonstrated that F-actin formed strong and dense stress fibers in
U251 glioma cells treated with TNF-α for 4 h (Fig. 3B).
Knockdown of PDHA1 is associated with
the migration and invasion capacities of glioma cells
Following stimulation with TNF-α, the expression
levels of the migratory markers were increased suggesting that they
were also associated with p-PDHA1 levels. U251 cells were
transfected with four PDHA1-specific siRNAs and the data
demonstrated that siPDHA1-4 exhibited the most significant
knockdown efficiency in U251 glioma cells (Fig. 4A). Therefore, siPDHA1-4-transfected
U251 cells were selected and grouped according to the experimental
design. The stimulation time of TNF-α (50 ng/ml) was 4 h. The
association between p-PDHA1 and glioma cell was studied using cell
migration and wound-healing assays. The results indicated that
knockdown of p-PDHA1 significantly inhibited the migratory and
migration efficiency of glioma cells at 24 and 48 h compared with 0
h. TNF-α stimulation did not enhance cell migration in
PDHA1-siRNA-transfected cells compared with the TNF-α stimulation
group (Fig. 4B and C). Subsequently,
western blot analysis was performed, and the data indicated that
TNFR1 levels were upregulated following TNF-α stimulation.
Knockdown of p-PDHA1 also reduced the expression levels of MMP2 and
MMP14. TNF-α stimulation increased the expression levels of MMP2,
MMP14 and p-PDHA1/PDHA1 (P<0.05). Whereas it did not increase
the expression levels of MMP2, MMP14 and p-PDHA1/PDHA1 (P<0.05)
in PDHA1-siRNA transfected cells (Fig.
4D).
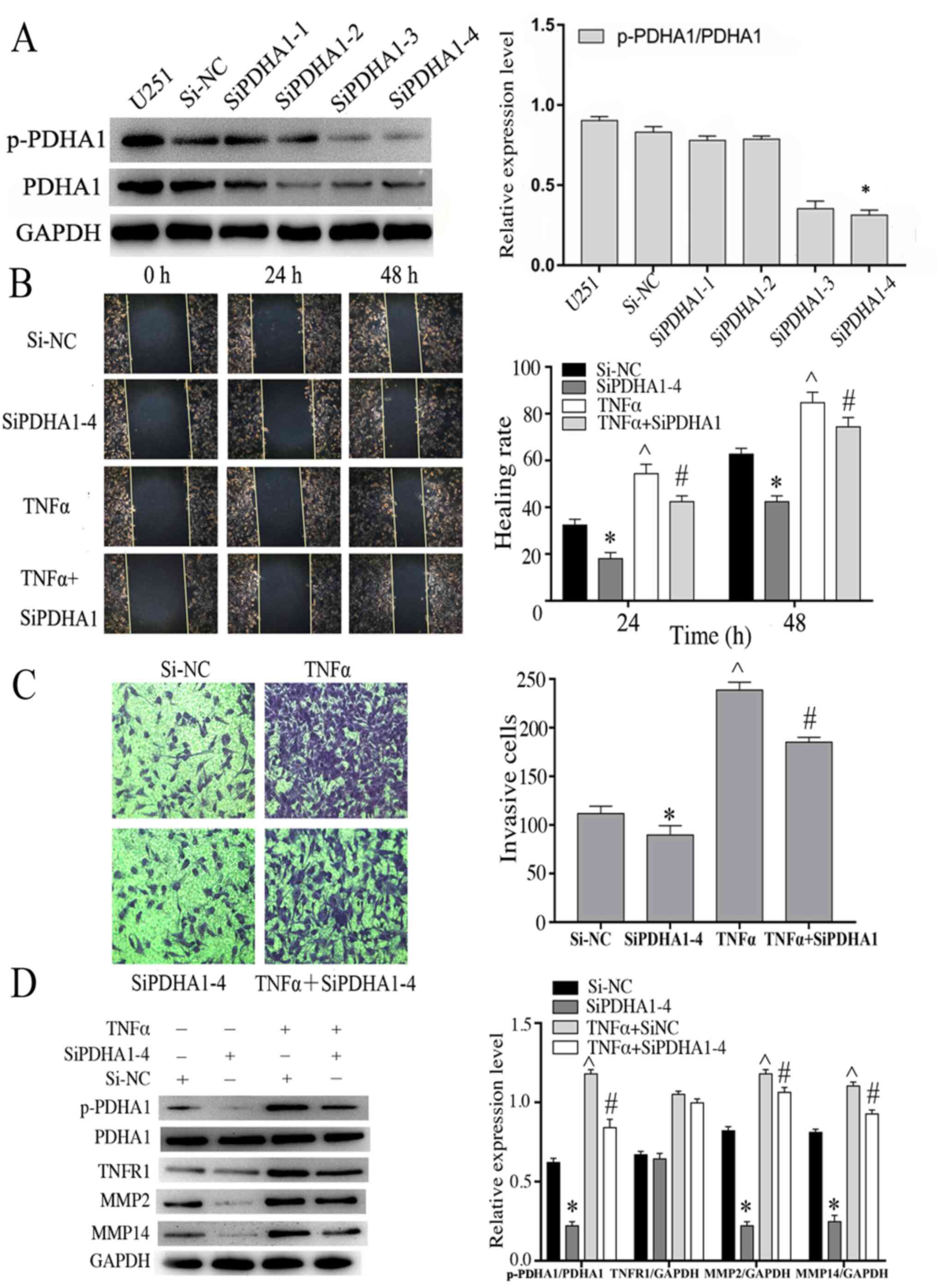 | Figure 4.Knockdown of PDHA1 is associated with
the migration and invasion capacities of glioma cells. (A) Western
blot analysis of the interference efficiency of U251 cells
transfected with siPDHA1. The bar chart indicates the ratio of
p-PDHA1 to PDHA1 expression. *P<0.05 vs. siNC. (B) Wound healing
assays of U251MG cells treated with si-NC, si-PDHA1-4, TNF-α and
siPDHA1 + TNF-α. The migration of glioma cells to the wound surface
at 0, 24 and 48 h was observed using the inverted Leica phase
contrast microscope (magnification, ×200). The bar chart indicates
the quantitative results of the wound healing assays *P<0.05,
^P<0.05 vs. siNC; #P<0.05 vs. TNF-α.
(C) Transwell invasion assays of U251 glioma cells in the si-NC,
siPDHA1-4, TNF-α and siPDHA1 + TNFα groups. The bar chart indicated
the quantitative results of the Transwell invasion assays
(magnification, ×100). *P<0.05, ^P<0.05 vs. siNC;
#P<0.05 vs. TNF-α. (D) Western blot analysis of
siPDHA1-transfected and/or TNF-α-stimulated U251 glioma cells. The
bar chart indicates the ratio of p-PDHA1 to PDHA1 expression, as
well as the expression levels of TNFR1, MMP2 and MMP14 normalized
to those of GAPDH. *P<0.05, ^P<0.05 vs. siNC;
#P<0.05 vs. TNF-α + siNC. p-PDHA1, phosphorylated
pyruvate dehydrogenase α1; PDHA1, pyruvate dehydrogenase α1; TNF-α,
tumor necrosis factor-α; TNFR1, tumor necrosis factor receptor
1. |
Effects of DCA on the migration and
invasion capacities of glioma cells stimulated by TNF-α
It has been reported that DCA is the inhibitor with
preliminary clinical effects on glioblastoma (19). PDK is an important kinase that
regulates the activity of the PDHc. Therefore, the protein levels
of p-PDHA1 were examined in U251 glioma cells at various
concentrations of DCA. The results indicated that p-PDHA1/PDHA1
level (P<0.05) was significantly lower following treatment of
the cells with 20 mM DCA compared with that noted in the control
cells (Fig. 5A). U251 glioma cells
were incubated with 20 mM DCA for 24 h and grouped according to the
experimental design. The stimulation time of TNF-α (50 ng/ml) was 4
h. Based on the analysis of the wound healing and transwell
invasion assays, DCA-treatment of U251 cells resulted in a
significant inhibition in their migratory and invasive efficacy.
TNF-α stimulation did not enhance cell migration in the DCA-treated
cells compared with the cells treated with TNF-α alone (Fig. 5B and C). Western blot analysis
indicated that DCA reduced the expression levels of MMP2 and MMP14.
TNF-α stimulation increased the expression levels of MMP2, MMP14
and p-PDHA1/PDHA1 (P<0.05). Whereas it did not increase the
expression levels of MMP2, MMP14 and p-PDHA1/PDHA1 (P<0.05) in
the DCA-treated glioma cells (Fig.
5D).
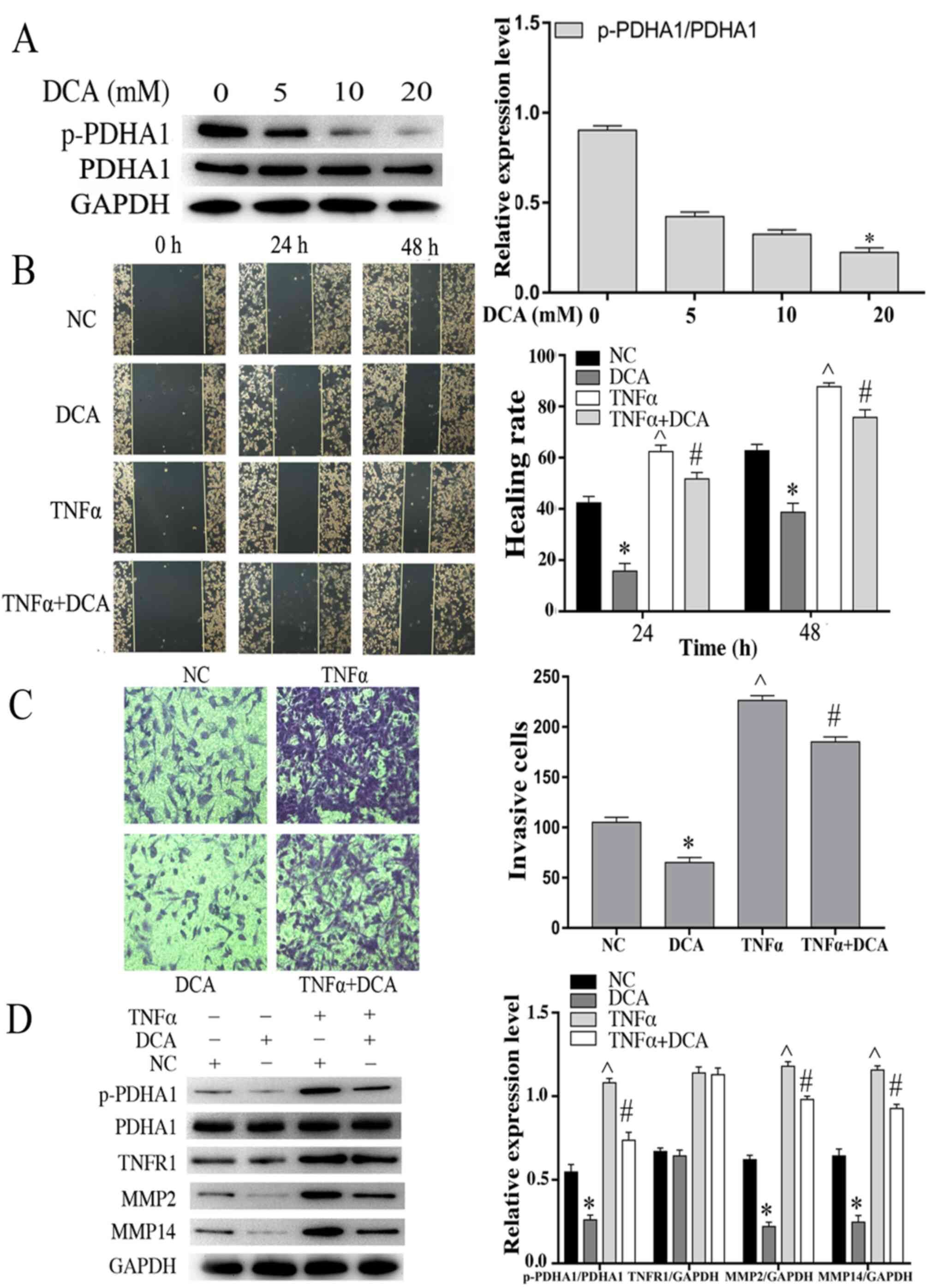 | Figure 5.Effects of DCA on the migration and
invasion of glioma cells stimulated by TNF-α. (A) Western blot
analysis of U251 glioma cells incubated with different
concentrations of DCA for 24 h. The bar chart indicates the ratio
of p-PDHA1 to PDHA1 expression. *P<0.05 vs. 0 mM. (B) Wound
healing assays of NC, DCA, TNF-α and DCA+TNF-α U251 cells. The bar
chart is representative of the quantitative results of the wound
healing assays. *P<0.05, ^P<0.05 vs. NC;
#P<0.05 vs. DCA. (C) Transwell invasion assays of NC,
DCA, TNF-α and DCA + TNFα U251 cell groups. The bar chart is
indicative of the quantitative results of the Transwell invasion
assay. *P<0.05, ^P<0.05 vs. NC;
#P<0.05 vs. DCA. (D) Western blot analysis of U251MG
cells treated with DCA and/or TNF-α. The bar chart indicated the
ratio of p-PDHA1 to PDHA1 expression and the expression levels of
TNFR1, MMP2 and MMP14 normalized to those of GAPDH. *P<0.05,
^P<0.05 vs. NC; #P<0.05 vs. DCA. DCA,
dichloroacetic acid; TNF-α, tumor necrosis factor-α; p-PDHA1,
phosphorylated pyruvate dehydrogenase α1; NC, negative control. |
Discussion
Invasion and migration are the main biological
characteristics of tumor malignancy. A previous study has reported
that the three main characteristics of tumor cell migration are the
following: Tumor cells adhere to ECM and degrade it using lyase,
cathepsin and autocrine or paracrine cytokines, such as 5-HT,
histamine and bradykinin (20). All
these processes alter cell morphology and enable the migration
movement using pseudopodia (20).
This partially explains the poor survival rate of patients with
glioma. Migration is the key to invasion; therefore, the control of
the migration of glioma cells is particularly important for the
effective treatment of this disease (21).
Several inflammatory cytokines have been
demonstrated to be involved in the tumor microenvironment, such as
interleukin (IL)-1, IL-6, IL-1β, TNF-α and TGF-β, which not only
recruit inflammatory cells to the tumor site, but also amplify the
inflammatory effect. They also promote the proliferation and
metastasis of tumor cells, and accelerate the formation of tumor
blood vessels and lymphatic vessels (22–24).
Tumor necrosis factor can induce a variety of processes, such as
apoptosis, necrosis and cell migration (25–27).
There are two types of TNFR receptors: TNFR1 and TNFR2. The
majority of the biological effects of TNF-α are produced by TNFR1
(28). In the present study, the
role of p-PDHA1 was investigated in the regulation of genes
involved in cell migration and TNF-α-induced gene expression in
glioma cells.
Normal expression of PDHA1 is a prerequisite for the
carboxylic acid cycle and oxidative phosphorylation occurring in
the mitochondria. PDHA1 is an important catalytic component of
PDHc. Phosphorylation of PDHA1 occurs at three specific serine
residues and the single phosphorylation of each site can cause PDHc
inactivation. Phosphorylation of site 1 is the most rapid reaction;
covalent modification occurs at specific serine residues in PDHA1
(29). Tumor progression promotes
the development of hypoxia and induces the expression of hypoxia
inducible factors (HIFs). HIFs induce high expression of PDK, which
phosphorylates PDHA1 and inactivates PDHc (30). Although aerobic glycolysis has been
widely accepted as a metabolic feature of tumors, its mechanism in
tumor progression remains unclear. In the present study, the data
demonstrated that the protein levels of TNFR1 and p-PDHA1 were
altered in glioma cells following the stimulation with TNF-α. The
present study examined whether p-PDHA1 was associated with the
inflammatory microenvironment, and its biological interaction with
the processes of invasion and migration of glioma cells was further
explored. Western blotting was used to analyze the activity of
glioma cell migration following stimulation by TNF-α. The potential
of glioma cell migration was evaluated by immunofluorescence
staining of the cytoskeleton. It has been reported that PDK and PDH
phosphatase regulate the activity of PDHA1. Phosphorylation of
PDHA1 can cause PDHc inactivation. It can enhance the Warburg
effect and the invasiveness of tumor cells (17). Therefore, in the present study, the
involvement of PDHA1 phosphorylation in glioma cell migration and
invasion was further assessed.
Various proteases, including MMPs, are involved in
the destruction of normal brain tissue (31). The wound healing and Transwell
invasion assays demonstrated that knockdown of the expression of
PDHA1 and reduced levels of p-PDHA1 decreased the migration and
invasion of glioma cells. TNF-α stimulation promoted glioma cell
migration and invasion, whereas this effect was not noted in PDHA1
siRNA-transfected cells. DCA is an inhibitor of PDK, which is an
important kinase that regulates the activity of PDHc (19). The present study indicated that DCA
treatment significantly inhibited the migratory and invasive
efficiency of glioma cells. TNF-α stimulation promoted glioma cell
migration and invasion, whereas TNF-α stimulation did not enhance
cell migration in the DCA-treated cells.
In conclusion, the present study demonstrated that
p-PDHA1 served an important role in cell migration and invasion of
glioma cells, suggesting that its level was associated with the
development of glioma. Furthermore, the data indicated that TNF-α
induced the level of p-PDHA1 in glioma cells. Therefore, p-PDHA1
may represent a key messenger for the regulation of cellular
movement and gene expression. A more comprehensive understanding of
this pathway may be helpful in establishing new therapeutic
strategies for glioma.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant nos. 81572491,
81602201 and 81502169).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
CZ and DW designed the study. ZY and YW performed
the experiment and wrote the manuscript. LZ analyzed the clinical
data. All authors read, revised and approved the final version of
the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Nantong University, Jiangsu, China (grant no.
20190313). The present manuscript is one of the results of the
project. All patients or their family members provided informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Reference
|
1
|
Rasmussen BK, Hansen S, Laursen RJ,
Kosteljanetz M, Schultz H, Norgard BM, Guldberg R and Gradel KO:
Epidemiology of glioma: Clinical characteristics, symptoms, and
predictors of glioma patients grade I–IV in the the Danish
Neuro-oncology registry. J Neurooncol. 135:571–579. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hakyemez B, Erdoğan C, Ercan I, Ergin N,
Uysal S and Atahan S: High-grade and low-grade gliomas:
Differentiation by using perfusion MR imaging. Clin Radiol.
60:493–502. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brat DJ, Aldape K, Colman H, Holland EC,
Louis DN, Jenkins RB, Kleinschmidt-DeMasters BK, Perry A,
Reifenberger G, Stupp R, et al: cIMPACT-NOW update 3: Recommended
diagnostic criteria for ‘Diffuse astrocytic glioma, IDH-wildtype,
with molecular features of glioblastoma, WHO grade IV’. Acta
Neuropathol. 136:805–810. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Anjum K, Shagufta BI, Abbas SQ, Patel S,
Khan I, Shah SAA, Akhter N and Hassan SSU: Current status and
future therapeutic perspectives of glioblastoma multiforme (GBM)
therapy: A review. Biomed Pharmacother. 92:681–689. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hwang JS, Jung EH, Kwon MY and Han IO:
Glioma-secreted soluble factors stimulate microglial activation:
The role of interleukin-1β and tumor necrosis factor-α. J
Neuroimmunol. 298:165–171. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Camelo S, Lafage M, Galelli A and Lafon M:
Selective role for the p55 Kd TNF-alpha receptor in immune
unresponsiveness induced by an acute viral encephalitis. J
Neuroimmunol. 113:95–108. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jiang Y, Yu M, Hu X, Han L, Yang K, Ba H,
Zhang Z, Yin B, Yang XP, Li Z and Wang J: STAT1 mediates
transmembrane TNF-alpha-induced formation of death-inducing
signaling complex and apoptotic signaling via TNFR1. Cell Death
Differ. 24:660–671. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ting AT and Bertrand MJM: More to Life
than NF-κB in TNFR1 signaling. Trends Immunol. 37:535–545. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang X, Yin N, Guo A, Zhang Q, Zhang Y,
Xu Y, Liu H, Tang B and Lai L: NF-κB signaling and cell-fate
decision induced by a fast-dissociating tumor necrosis factor
mutant. Biochem Biophys Res Commun. 489:287–292. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang S, Xie C, Chen Y, Wang J, Chen X, Lu
Z, June RR and Zheng S: Differential roles of TNFα-TNFR1 and
TNFα-TNFR2 in the differentiation and function of
CD4+Foxp3+induced Treg cells in vitro and in
vivo periphery in autoimmune diseases. Cell Death Dis. 10:272019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Martínez-Reza I, Díaz L and García-Becerra
R: Preclinical and clinical aspects of TNF-α and its receptors
TNFR1 and TNFR2 in breast cancer. J Biomed Sci. 24:902017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Z, Yu M, Fei B, Fang X, Ma T and Wang
D: miR-21-5p targets PDHA1 to regulate glycolysis and cancer
progression in gastric cancer. Oncol Rep. 40:2955–2963.
2018.PubMed/NCBI
|
|
13
|
Ferriero R, Nusco E, De Cegli R, Carissimo
A, Manco G and Brunetti-Pierri N: Pyruvate dehydrogenase complex
and lactate dehydrogenase are targets for therapy of acute liver
failure. J Hepatol. 69:325–335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stacpoole PW: Therapeutic targeting of the
pyruvate dehydrogenase complex/pyruvate dehydrogenase kinase
(PDC/PDK) axis in cancer. J Natl Cancer Inst. 109:2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu L, Li Y, Zhou L, Dorfman RG, Liu L, Cai
R, Jiang C, Tang D, Wang Y, Zou X, et al: SIRT3 elicited an
anti-Warburg effect through HIF1α/PDK1/PDHA1 to inhibit
cholangiocarcinoma tumorigenesis. Cancer Med. 8:2380–2391. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu F, Zhang W, You X, Liu Y, Li Y, Wang
Z, Wang Y, Zhang X and Ye L: The oncoprotein HBXIP promotes glucose
metabolism reprogramming via downregulating SCO2 and PDHA1 in
breast cancer. Oncotarget. 29:27199–27213. 2015. View Article : Google Scholar
|
|
17
|
Liu L, Cao J, Zhao J, Li X, Suo Z and Li
H: PDHA1 Gene Knockout in Human esophageal squamous cancer
cells resulted in greater warburg effect and aggressive features in
vitro and in vivo. Onco Targets Ther. 12:9899–9913. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Glotsos D, Spyridonos P, Petalas P,
Cavouras D, Ravazoula P, Dadioti PA, Lekka I and Nikiforidis G:
Computer-based malignancy grading of astrocytomas employing a
support vector machine classifier, the WHO grading system and the
regular hematoxylin-eosin diagnostic staining procedure. Anal Quant
Cytol Histol. 2:77–83. 2004.
|
|
19
|
Velpula KK, Guda MR, Sahu K, Tuszynski J,
Asuthkar S, Bach SE, Lathia JD and Tsung AJ: Metabolic targeting of
EGFRvIII/PDK1 axis in temozolomide resistant glioblastoma.
Oncotarget. 8:35639–35655. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Oudin MJ and Weaver VM: Physical and
chemical gradients in the tumor microenvironment regulate tumor
cell invasion, migration, and metastasis. Cold Spring Harb Symp
Quant Biol. 81:189–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Andreozzi M, Quintavalle C, Benz D,
Quagliata L, Matter M, Calabrese D, Tosti N, Ruiz C, Trapani F,
Tornillo L, et al: HMGA1 expression in human hepatocellular
carcinoma correlates with poor prognosis and promotes tumor growth
and migration in in vitro models. Neoplasia. 18:724–731. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yao X, Huang J, Zhong H, Shen N, Faggioni
R, Fung M and Yao Y: Targeting interleukin-6 in inflammatory
autoimmune diseases and cancers. Pharmacol Ther. 141:125–139. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Castano Z, San Juan BP, Spiegel A, Pant A,
DeCristo MJ, Laszewski T, Ubellacker JM, Janssen SR, Dongre A,
Reinhardt F, et al: IL-1β inflammatory response driven by primary
breast cancer prevents metastasis-initiating cell colonization. Nat
Cell Biol. 20:1084–1097. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Batlle E and Massague J: Transforming
growth factor-β signaling in immunity and cancer. Immunity.
50:924–940. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang BP, Lin CS, Wang CJ and Kao SH:
Upregulation of heat shock protein 70 and the differential protein
expression induced by tumor necrosis factor-alpha enhances
migration and inhibits apoptosis of hepatocellular carcinoma cell
HepG2. Int J Med Sci. 14:284–293. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shiozaki A, Shimizu H, Ichikawa D, Konishi
H, Komatsu S, Kubota T, Fujiwara H, Okamoto K, Iitaka D, Nakashima
S, et al: Claudin 1 mediates tumor necrosis factor alpha-induced
cell migration in human gastric cancer cells. World J
Gastroenterol. 20:17863–17876. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ooppachai C, Limtrakul Dejkriengkraikul P
and Yodkeeree S: Dicentrine potentiates TNF-α-induced apoptosis and
suppresses invasion of A549 lung adenocarcinoma cells via
modulation of NF-κB and AP-1 activation. Molecules. 24:41002019.
View Article : Google Scholar
|
|
28
|
Van Horssen R, Ten Hagen TL and Eggermont
AM: TNF-alpha in cancer treatment: Molecular insights, antitumor
effects, and clinical utility. Oncologist. 11:397–408. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Holness MJ and Sugden MC: Regulation of
pyruvate dehydrogenase complex activity by reversible
phosphorylation. Biochem Soc Trans. 31:1143–1151. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Izquierdo-Garcia JL, Viswanath P, Eriksson
P, Cai L, Radoul M, Chaumeil MM, Blough M, Luchman HA, Weiss S,
Cairncross JG, et al: IDH1 mutation induces reprogramming of
pyruvate metabolism. Cancer Res. 75:2999–3009. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou W, Yu X, Sun S, Zhang X, Yang W,
Zhang J, Zhang X and Jiang Z: Increased expression of MMP-2 and
MMP-9 indicates poor prognosis in glioma recurrence. Biomed
Pharmacother. 118:1093692019. View Article : Google Scholar : PubMed/NCBI
|