Introduction
Multiple myeloma (MM) is a malignancy of plasma
cells, and is the second most common type of hematopoietic
malignancy, accounting for 10–15% of total hematopoietic cancer
types (1). The prevalence of MM
increases with age and the median age at diagnosis is 69 years old
in the United States (1). Due to the
development of novel drugs and therapeutic methods, the overall
survival of patients with MM has significantly improved in recent
decades (2). However, MM remains an
incurable disease. One reason for this is the drug-resistance
developed in patients during the treatment (3,4).
Therefore, novel drugs and therapeutic methods are required to
improve the treatment strategies for patients with MM.
In cancer, epigenetic regulation contributes to
reversible modification of chromatin structure and gene
transcription (5). Oncogenes are
always upregulated or activated whereas tumor suppressor genes are
always downregulated or silenced in this process (5). Acetylation/deacetylation is one of the
mechanisms that is involved in this regulatory process. Lysine
residues of histone or non-histone can be acetylated by
histone/protein acetyltransferases, which is contrary to the
activity of histone deacetylases (HDAC) (6). Deacetylation of histone or non-histone
can lead to transcription silence of genes and deactivation of
protein (6,7). In cancer cells, inhibition of HDAC
leads to cell cycle arrest and induction of apoptosis. Targeted
inhibition of HDAC may therefore be considered as a potential
therapeutic method in MM treatment (8).
The silent mating type information regulation 2
homolog (SIRTUIN) family belongs to Class III HDACs and is a group
of proteins with nicotinamide adenine dinucleotide (NAD)-dependent
deacetylase activity (9). Their
substrates include histone and non-histone proteins (9). It has important roles in cancer cell
survival and apoptosis by deacetylating key cell signaling
molecules and apoptotic related proteins, including TATA binding
protein-associated factor 168, p53 and forkhead box O1 (FOXO1)
(10). Among the SIRTUIN family,
silent mating type information regulation 2 homolog 1 (SIRT1) is
the most-studied member in basic and translational oncology
research (11,12). Previous studies have reported that
SIRT1 contributes to the development of several solid tumors and
hematopoietic malignancies (13,14). In
addition, small compounds targeting SIRT1 could effectively inhibit
the proliferation of tumor cells. Therefore, the present study
aimed to investigate the antitumor effect of the SIRT1 inhibitor
cambinol on MM cells.
Materials and methods
Reagents and cell lines
MM cell lines RPMI8226 and U266 were ordered from
the American Type Culture Collection. A Cell Counting Kit (CCK)-8
assay was purchased from Beyotime Institute of Biotechnology.
Cambinol, Annexin V and PI were purchased from Sigma-Aldrich (Merck
KGaA). Anti-procaspase-3 (cat. no. 610322; 1:1,000), and
poly(ADP-ribose) polymerase 1 (PARP) (cat. no. 556362; 1:2,000)
antibodies were purchased from BD Biosciences. The anti-β-actin
antibody (cat. no. ZRB1312; 1:1,000) was purchased from
Sigma-Aldrich (Merck KGaA). Anti-p53 (cat. no. 48818; 1:1,000),
cleaved caspase 3 (cat. no. 9664; 1:1,000), acetylated p53 (Lys382;
Ac-p53) (cat. no.2525; 1:1,000), Bcl-2 (cat. no. 15071; 1:1,000),
cyclin D1 (cat. no.2922; 1:1,000) and p21 (cat. no. 2947; 1:1,000)
antibodies were purchased from Cell Signaling Technology, Inc. The
anti-mouse and rabbit horseradish peroxidase-conjugated secondary
antibodies (cat. no. 7076, 7074; 1:5,000) were purchased from Cell
Signaling Technology, Inc. Culture RPMI-1640 medium and FBS were
purchased from Cytiva. Fluorescence Activated Cell Sorter (FACS)
was obtained from BD Biosciences.
Cell viability assay
MM cells were routinely cultured in RPMI-1640 medium
supplemented with 10% FBS at 37°C in a humidified incubator with 5%
CO2. For the proliferation assay, cells were seeded in
96-well plate at a density of 2×104 cell/well. The cells
were incubated with various concentrations of cambinol (20, 40, 80,
160 and 320 µM) dissolved in DMSO for 24 and 48 h (at 37°C in 5%
CO2). Then, 20 µl CCK-8 solution was added to each well
and the cells were incubated at 37°C in 5% CO2 for 4 h
according to the manufacturer's protocol. The absorbance was
measured at 450 nm and values are presented as mean ± SD.
Cell apoptosis assay
Apoptosis measurement was performed according to the
protocol of Annexin V-FITC Apoptosis Detection Kit (Beyotime
Institute of Biotechnology). Cells were treated with 80 µM cambinol
for 48 h at 37°C in 5% CO2 and were collected via
centrifugation (1,000 × g for 5 min at 4°C). Then, cells were
washed once with 1X washing buffer. Cells were resuspended in 500
µl working staining solution and incubated in ice for 5 min,
subsequently 5 µl Annexin V was added. Then, 5 µl PI solution was
added to each tube and cells were incubated in ice for 15 min, the
cells were detected using FACS. Results are representative of three
independent experiments.
Cell cycle analysis
Cells were treated with 80 µM cambinol at 37°C for
48 h and were collected via centrifugation (1,000 × g for 5 min at
room temperature). The collected cells were resuspended in 70%
ethanol and incubated at −20°C overnight. On the following day,
cells were washed twice with PBS and resuspended in 0.5 ml PBS
containing 0.1% Triton X-100, 0.2 mg/ml DNase-free RNase and 2
µg/ml PI. Cells were incubated on ice in the dark and cell cycle
was assessed via FACS within 2 h.
SIRT1 activity assay
SIRT1 activity was analyzed using a fluorogenic
SIRT1 assay kit (cat. no. CS1040; Sigma-Aldrich; Merck KGaA).
According to the manufacturer's instructions, whole cell lysate
proteins were extracted with RIPA buffer (Beyotime Institute of
Biotechnology) and the fluorogenic SIRT1 assay kit was used to
perform the assay.
Western blotting
After drug treatment for 48 h, cells in a 60-mm
culture dish were washed twice with PBS, lysed in 150 µl ice-cold
high-efficiency RIPA buffer with the protease inhibitor PMSF (1 mM)
for 30 min, then collected to a 1.5 ml tube and centrifuged at 4°C
12,000 × g for 10 min. The protein concentration was quantified in
supernatant using a BCA assay kit. The supernatant was mixed with
5X loading buffer and heated at 100°C for 5 min and samples (40 µg
per lane) were loaded and separated via SDS-PAGE (10% separating
gel). The 0.2-µm PVDF membrane was used for protein transfer.
Membranes were blocked with 5% non-fat dry-milk in TBS-Tween-20
(TBST; 0.1% v/v Tween-20) for 1 h at room temperature, and then
incubated with the primary antibodies overnight at 4°C. After
washing three times with TBST, the membranes were incubated with
horseradish peroxidase-conjugated secondary antibodies for 1 h at
room temperature. The membranes were washed three times with TBST,
and incubated with chemiluminescent substrates (Santa Cruz
Biotechnology, Inc.). Protein semi-quantification was conducted
with ImageJ software (National Institutes of Health). Results are
representative of three independent experiments.
Statistical analysis
Data are presented as the mean ± SD. Differences
between two groups were determined using unpaired two-sample
Student's t-test. Comparisons between multiple groups were
determined using a one-way ANOVA with Bonferroni post hoc analysis.
Significance was analyzed using GraphPad Prism 6.0 (GraphPad
Software, Inc.). P<0.05 was considered to indicate a
statistically significant difference.
Results
Cambinol inhibits the proliferation of
RPMI8226 and U266 cells
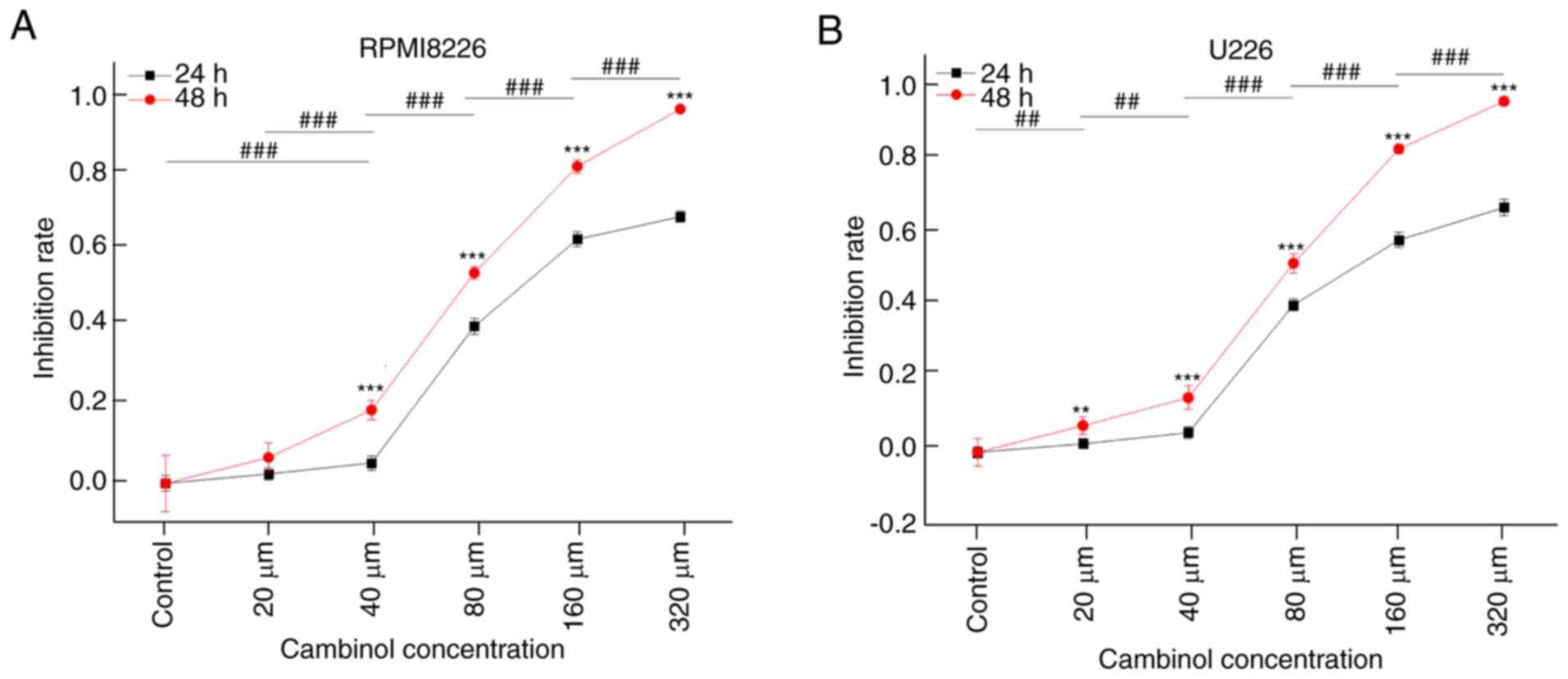
The inhibitive effect of cambinol on MM cell
proliferation was measured using a CCK-8 assay. RPMI8226 and U266
cell lines were treated with different concentrations (20, 40, 80,
160 and 320 µM) of cambinol for 24 and 48 h. It was demonstrated
that cambinol inhibited the proliferation of RPMI8226 and U266
cells in a dose- and time-dependent manner. IC50 values
of cambinol on RPMI8226 and U266 were ~77.24 and 79.23 µM,
respectively, following the 48 h treatment (Fig. 1A and B).
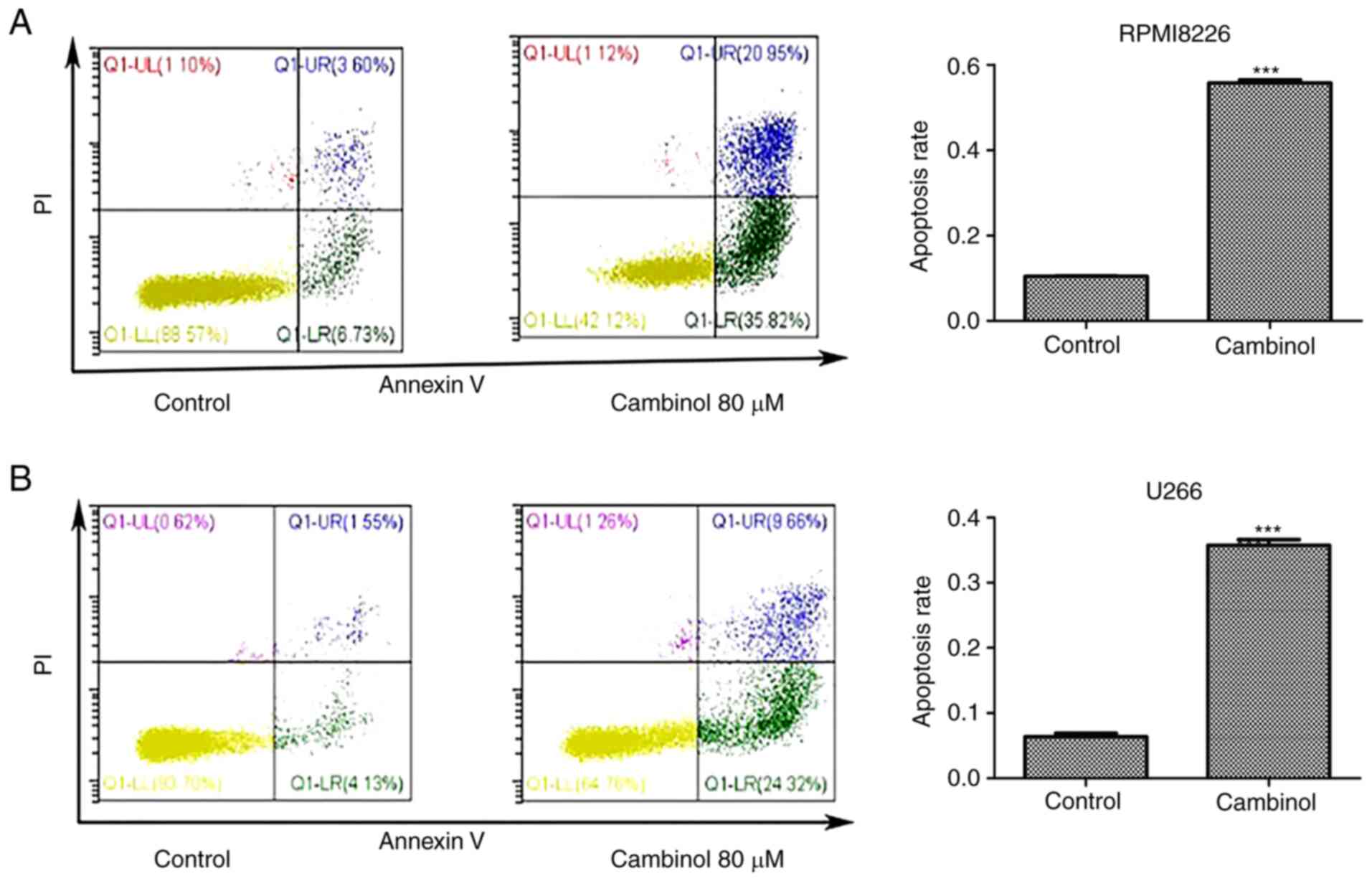
Cambinol induces apoptosis in RPMI8226
and U266 cells
The apoptosis-inducing effect of cambinol on
RPMI8226 and U266 cell lines was measured via FACS. MM cells were
cultured with 80 µM cambinol for 48 h. FACS data demonstrated that
the apoptotic rate of RPMI8226 cells increased from 11.43% in the
control group to 55.72% in the drug-treated group (Fig. 2A), and the rate of apoptosis of U266
cells increased from 6.3% in control group to 36.71% in the
drug-treated group (Fig. 2B). These
data indicate that cambinol significantly induced apoptosis in
RPMI8226 and U266 cells.
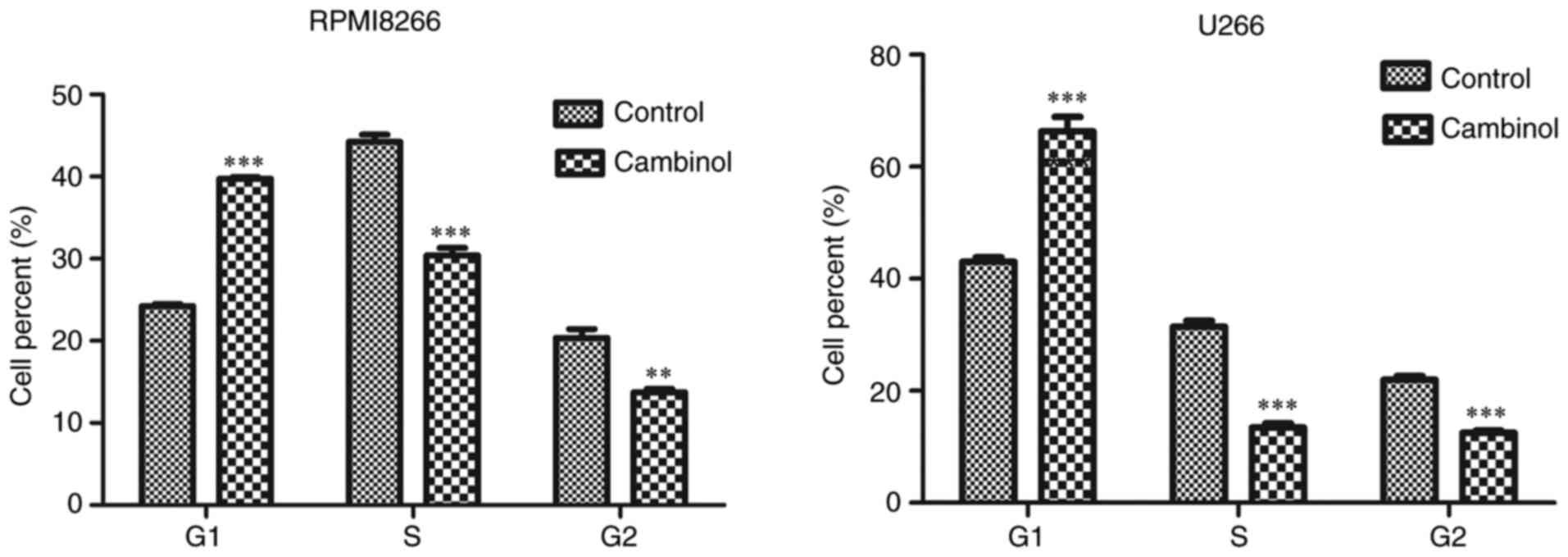
Cambinol induces cell cycle arrest in
RPMI8226 and U266 cells
To examine the effect of cambinol on the cell cycle
of RPMI8226 and U266 MM cells, cells were treated with cambinol for
48 h, and then analyzed via FACS after staining with PI. For
RPMI8226 cells, the percentage of cells in the G1 phase
was 24.23% in the control group, which increased to 39.73% in the
drug-treated group. For U266 cells, 43.03% of cells were in the
G1 phase in the control group, which increased to 66.33%
in the drug-treated group (Fig.
3).
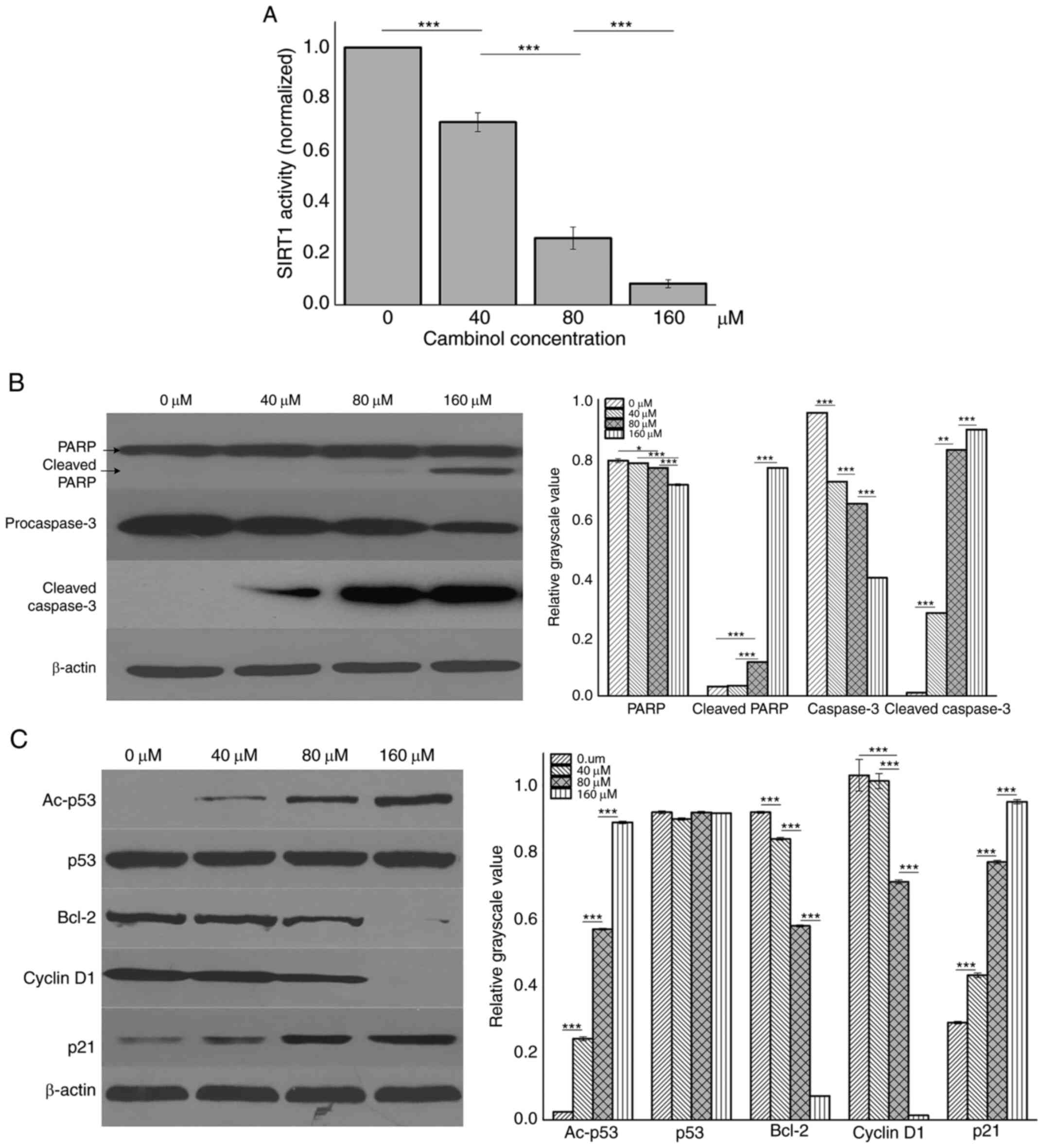
Inhibition of cambinol on SIRT1
activity and the effect on protein expression levels inRPMI8226
cells
RPMI8226 cells were treated with different
concentrations (0, 40, 80 and 160 µM) of cambinol for 48 h and then
lysed with RIPA buffer. SIRT1 activity was analyzed using an SIRT1
assay kit. It was demonstrated that cambinol decreased SIRT1
activity in a dose-dependent manner (P<0.001; Fig. 4A). Western blotting was used to
measure expression levels of PARP, caspase3, p53, Ac-p53, Bcl-2,
cyclin D1 and p21. It was identified that the expression levels of
cleaved PARP, cleaved caspase3, Ac-p53 and p21 were upregulated in
a dose-dependent manner, while the expression levels of
procaspase3, Bcl-2 and cyclin D1 were downregulated (Fig. 4B and C).
Discussion
SIRT1 belongs to class III histone deacetylases
(HDAC) and it is a homologue protein of yeast silent information
regulator 2 (Sir2). The de-acetylation activity of type III HDACs
depends on the cofactor NAD, which is dispensable for type I and
type II HDACs. The human SIRTUIN family is a homologue to Sir2, and
these are highly conservative from bacteria to humans. SIRT1 and
SIRT2 of the SIRTUIN family have highly similar sequences, and are
associated with cell apoptosis, the cell cycle, DNA repair, DNA
recombination and gene silencing (15,16).
SIRT1 deacetylates histone H3 and histone H4. It
also interacts with transcription factors or co-factors, including
p53, FOXO family member, NF-κB, myoblast determination protein 1,
nuclear receptor corepressor, p300 and peroxisome
proliferator-activated receptor γ coactivator 1-α, and regulates
their transcriptional activity. SIRT1 inhibitors exhibit antitumor
activity, which has been observed in cells from breast, thyroid,
lung, pancreatic, prostate, hepatitis and colon cancer (17–21).
Moreover, SIRT1 has effective antitumor activity against
hematological malignancies, such as T/B-cell lymphoma, chronic
myeloid leukemia (CML), leukemia and other hematological
malignancies (11,22–26).
Cambinol is an inhibitor of SIRT1/2 that has been identified to
exert antitumor activity and low toxicity in murine models
(27,28). The present results demonstrated that
cambinol exhibited anti-proliferative effects on RPMI8226 and U266
cell lines that were dose- and time-dependent. Previous studies
have reported that the antitumor activity of SIRT1 inhibitors was
the result of cell apoptosis and cell cycle arrest (27). In primary exudative lymphoma, SIRT1
inhibitors were found to induce cell apoptosis and cell cycle
arrest at the G1 phase (24). When treated with cambinol, the
proliferation of hepatic cancer cells is inhibited and the tumor
burden in mice is also significantly reduced (27). The present results demonstrated that
cambinol suppressed the activity of SIRT1, promoted cell apoptosis
and arrested the cell cycle at the G1 phase in MM
cells.
p53 is a tumor suppresser, which is actively
involved in tumor occurrence and development. The antitumor
function of p53 is mediated via two pathways:
Transcription-dependent or -independent signaling pathways. As a
transcription factor, p53 can activate the transcription and
expression of multiple target genes to regulate cell apoptosis,
including growth arrest and DNA damage inducible α, murine double
minute-2, p21, Bax, fas, insulin-like growth factor-binding protein
3 and TNF receptor superfamily member 10b. p53 can also activate
apoptosis by interacting with target proteins directly, which is
another effective cell-regulating mechanism (29). p53 can be activated by acetylation
(30). Using a SIRT1 inhibitor or
gene silencing methods, the protein level of acetylated p53 can be
increased in CML cells, and the elimination of CML stem cells can
be further potentiated when combined with imatinib (a tyrosine
kinase inhibitor) (31). The SIRT1
inhibitor regulates downstream-associated proteins and cyclins
through increasing the active form of p53 and decreasing Bcl-2
protein and cyclin D1. Also, the SIRT1 inhibitor reduces the cell
proliferation rate and induces apoptosis in breast cancer cell
(32). The present study identified
that cambinol increased the acetylation level of p53, which further
upregulates p21 protein levels, subsequently inhibiting cyclin D1.
In addition, the results indicated that increased levels of
acetylated p53 was a possible mechanism for the inhibitory effect
of cambinol on MM cells.
MM is currently recognized as an incurable tumor
type, due to drug resistance. The application of novel drugs is an
effective method to improve the survival of patients with MM.
Cambinol was investigated to inhibit cell viability in
hepatocellular carcinoma, breast cancer, prostate cancer and human
glioma cell lines (33–36), and also exhibits antitumor activity.
According to the literature, to the best of our knowledge, no
studies have examined the effect and mechanism of cambinol in MM
cells. The present study demonstrated the antitumor activity of
cambinol on MM cells and described the potential mechanism.
Moreover, further studies to increase the potency and selectivity
towards SIRT1 or SIRT2 for enhanced their antitumor efficacy are
underway. Cambinol analogs may be beneficial for sensitizing tumor
cells to other chemotherapeutic agents (37,38).
Thus, cambinol and its analogs are promising antitumor agents for
MM.
In conclusion, the present study investigated the
effect of cambinol on MM cell lines-RPMI8226 and U266, and found
that cambinol can inhibit proliferation, and induce apoptosis of MM
cells. The possible underlying mechanism of cambinol on MM cells
involves the upregulation of acetylation p53 protein through
inhibition of SIRT1. The results of this study provide evidence for
the further clinical use of cambinol or its analogs in treating
patients with MM.
Acknowledgements
Not applicable.
Funding
This study was supported by the Sanming Project of
Medicine in Shenzhen (grant no. SZSM201911004) and the hospital
research fund of The Seventh Affiliated Hospital Sun Yat-sen
University (grant no. ZSQYLCKYJJ202027).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XX and BL conceived and designed all the
experiments. BL and DZ performed the experiments. XW, YC and DL
analyzed the experimental data. BL and DZ contributed to manuscript
preparation, writing, editing and revision. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lynch HT, Watson P, Tarantolo S, Wiernik
PH, Quinn-Laquer B, Isgur Bergsagel K, Huiart L, Olopade OI, Sobol
H, Sanger W, et al: Phenotypic heterogeneity in multiple myeloma
families. J Clin Oncol. 23:685–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Terpos E; International Myeloma Society, :
Multiple myeloma: Clinical updates from the American Society of
hematology annual meeting, 2017. Clin Lymphoma Myeloma Leuk.
18:321–334. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Robak P, Drozdz I, Szemraj J and Robak T:
Drug resistance in multiple myeloma. Cancer Treat Rev. 70:199–208.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xu XH, Liu J, Shen CY, Ding LL, Zhong F,
Ouyang Y, Wang YC and He S: The role of ubiquitin-specific protease
14 (USP14) in cell adhesion-mediated drug resistance (CAM-DR) of
multiple myeloma cells. Eur J Haematol. 98:4–12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Liu M, Jiang L and Guan XY: The genetic
and epigenetic alterations in human hepatocellular carcinoma: A
recent update. Protein Cell. 5:673–691. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Khochbin S, Verdel A, Lemercier C and
Seigneurin-Berny D: Functional significance of histone deacetylase
diversity. Curr Opin Genet Dev. 11:162–166. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tang Y, Zhao W, Chen Y, Zhao Y and Gu W:
Acetylation is indispensable for p53 activation. Cell. 133:612–626.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Conte M, De Palma R and Altucci L: HDAC
inhibitors as epigenetic regulators for cancer immunotherapy. Int J
Biochem Cell Biol. 98:65–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jin X, Wei Y, Xu F, Zhao M, Dai K, Shen R,
Yang S and Zhang N: SIRT1 promotes formation of breast cancer
through modulating Akt activity. J Cancer. 9:2012–2023. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kumar A and Chauhan S: How much successful
are the medicinal chemists in modulation of SIRT1: A critical
review. Eur J Med Chem. 119:45–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Roth M, Wang Z and Chen WY: Sirtuins in
hematological aging and malignancy. Crit Rev Oncog. 18:531–547.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hwang ES and Song SB: Nicotinamide is an
inhibitor of SIRT1 in vitro, but can be a stimulator in cells. Cell
Mol Life Sci. 74:3347–3362. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li L and Bhatia R: Role of SIRT1 in the
growth and regulation of normal hematopoietic and leukemia stem
cells. Curr Opin Hematol. 22:324–329. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tae IH, Park EY, Dey P, Son JY, Lee SY,
Jung JH, Saloni S, Kim MH and Kim HS: Novel SIRT1 inhibitor
15-deoxy-Δ12,14-prostaglandin J2 and its derivatives exhibit
anticancer activity through apoptotic or autophagic cell death
pathways in SKOV3 cells. Int J Oncol. 53:2518–2530. 2018.PubMed/NCBI
|
|
15
|
Mellini P, Valente S and Mai A: Sirtuin
modulators: An updated patent review (2012–2014). Expert Opin Ther
Pat. 25:5–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hirsch BM and Zheng W: Sirtuin mechanism
and inhibition: Explored with N(ε)-acetyl-lysine analogs. Mol
Biosyst. 7:16–28. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Panathur N, Gokhale N, Dalimba U, Koushik
PV, Yogeeswari P and Sriram D: New indole-isoxazolone derivatives:
Synthesis, characterisation and in vitro SIRT1 inhibition studies.
Bioorg Med Chem Lett. 25:2768–2772. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Oon CE, Strell C, Yeong KY, Ostman A and
Prakash J: SIRT1 inhibition in pancreatic cancer models:
Contrasting effects in vitro and in vivo. Eur J Pharmacol.
757:59–67. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yao Y, Liu T, Wang X and Zhang D: The
contrary effects of Sirt1 on MCF7 cells depend on CD36 expression
level. J Surg Res. 238:248–254. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karbasforooshan H, Roohbakhsh A and Karimi
G: SIRT1 and microRNAs: The role in breast, lung and prostate
cancers. Exp Cell Res. 367:1–6. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ghosh A, Sengupta A, Seerapu GPK, Nakhi A,
Shivaji Ramarao EVV, Bung N, Bulusu G, Pal M and Haldar D: A novel
SIRT1 inhibitor, 4bb induces apoptosis in HCT116 human colon
carcinoma cells partially by activating p53. Biochem Biophys Res
Commun. 488:562–569. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou L, Wang Q, Chen X, Fu L, Zhang X,
Wang L, Deng A, Li D, Liu J, Lv N, et al: AML1-ETO promotes SIRT1
expression to enhance leukemogenesis of t(8;21) acute myeloid
leukemia. Exp Hematol. 46:62–69. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sharma VK, Raimondi V, Ruggero K,
Pise-Masison CA, Cavallari I, Silic-Benussi M, Ciminale V and
D'Agostino DM: Expression of miR-34a in T-cells infected by human
T-Lymphotropic Virus 1. Front Microbiol. 9:8322018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
He M, Tan B, Vasan K, Yuan H, Cheng F,
Ramos da Silva S, Lu C and Gao SJ: SIRT1 and AMPK pathways are
essential for the proliferation and survival of primary effusion
lymphoma cells. J Pathol. 242:309–321. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bhalla S and Gordon LI: Functional
characterization of NAD dependent de-acetylases SIRT1 and SIRT2 in
B-cell chronic lymphocytic leukemia (CLL). Cancer Biol Ther.
17:300–309. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim HB, Lee SH, Um JH, Oh WK, Kim DW, Kang
CD and Kim SH: Sensitization of multidrug-resistant human cancer
cells to Hsp90 inhibitors by down-regulation of SIRT1. Oncotarget.
6:36202–36218. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Portmann S, Fahrner R, Lechleiter A, Keogh
A, Overney S, Laemmle A, Mikami K, Montani M, Tschan MP, Candinas D
and Stroka D: Antitumor effect of SIRT1 inhibition in human HCC
tumor models in vitro and in vivo. Mol Cancer Ther. 12:499–508.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heltweg B, Gatbonton T, Schuler AD,
Posakony J, Li H, Goehle S, Kollipara R, Depinho RA, Gu Y, Simon JA
and Bedalov A: Antitumor activity of a small-molecule inhibitor of
human silent information regulator 2 enzymes. Cancer Res.
66:4368–4377. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang J: Current developments of targeting
the p53 signaling pathway for cancer treatment. Pharmacol Ther. Oct
29–2020.(Epub ahead of print). doi:
10.1016/j.pharmthera.2020.107720. View Article : Google Scholar
|
|
30
|
Brooks CL and Gu W: New insights into p53
activation. Cell Res. 20:614–621. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Li L, Wang L, Li L, Wang Z, Ho Y, McDonald
T, Holyoake TL, Chen W and Bhatia R: Activation of p53 by SIRT1
inhibition enhances elimination of CML leukemia stem cells in
combination with imatinib. Cancer Cell. 21:266–281. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Peck B, Chen CY, Ho KK, Di Fruscia P,
Myatt SS, Coombes RC, Fuchter MJ, Hsiao CD and Lam EW: SIRT
inhibitors induce cell death and p53 acetylation through targeting
both SIRT1 and SIRT2. Mol Cancer Ther. 9:844–855. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ceballos MP, Decandido G, Quiroga AD,
Comanzo CG, Livore VI, Lorenzetti F, Lambertucci F,
Chazarreta-Cifre L, Banchio C, Alvarez ML, et al: Inhibition of
sirtuins 1 and 2 impairs cell survival and migration and modulates
the expression of P-glycoprotein and MRP3 in hepatocellular
carcinoma cell lines. Toxicol Lett. 289:63–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dykes SS, Friday E, Pruitt K and Cardelli
JA: The histone deacetylase inhibitor cambinol prevents acidic
pHe-induced anterograde lysosome trafficking independently of
sirtuin activity. Biochem Biophys Rep. 3:83–93. 2015.PubMed/NCBI
|
|
35
|
Simmons GE Jr, Pandey S,
Nedeljkovic-Kurepa A, Saxena M, Wang A and Pruitt K: Frizzled 7
expression is positively regulated by SIRT1 and β-catenin in breast
cancer cells. PLoS One. 9:e988612014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Holloway KR, Barbieri A, Malyarchuk S,
Saxena M, Nedeljkovic-Kurepa A, Cameron Mehl M, Wang A, Gu X and
Pruitt K: SIRT1 positively regulates breast cancer associated human
aromatase (CYP19A1) expression. Mol Endocrinol. 27:480–490. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Botta L, Filippi S, Bizzarri BM, Meschinia
R, Caputoa M, Proietti-De-Santisa L, Isideb C, Nebbiosob A,
Gualandia G and Saladinoa R: Oxidative nucleophilic substitution
selectively produces cambinol derivatives with antiproliferative
activity on bladder cancer cell lines. Bioorg Med Chem Lett.
29:78–82. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mahajan SS, Scian M, Sripathy S, Posakony
J, Lao U, Loe TK, Leko V, Thalhofer A, Schuler AD, Bedalov A and
Simon JA: Development of pyrazolone and isoxazol-5-one cambinol
analogues as sirtuin inhibitors. J Med Chem. 57:3283–3294. 2014.
View Article : Google Scholar : PubMed/NCBI
|