Introduction
Cancer has been one of the leading causes of
disease-associated death worldwide since 2010, and the incidence
and mortality rates associated with various types of cancer are
increasing (1). Breast cancer is the
most commonly diagnosed type of cancer and the leading cause of
cancer-associated death in women, with ~17,000,000 new cases
occurring annually in the world (2),
and its incidence and mortality rates are expected to increase
significantly in the next 5–10 years. In developing countries, the
incidence rate will increase by 55% and the mortality rate will
increase by 58% within 20 years (2).
There are several causes of breast cancer, among which gene
mutations are closely associated with the development and/or
progression of breast cancer (3,4). The
diagnosis, treatment and prognosis of patients with breast cancer
has improved; however, the median survival of the majority of
patients with metastatic cancer remains low (24 months) (5). Additionally, a majority of patients
with breast cancer will experience a relapse following treatment.
Therefore, identifying effective prognostic biomarkers may assist
in predicting the prognosis of patients with cancer and the
curative effects of several therapeutics, consequently providing a
suitable treatment plan and ultimately improving the clinical
efficacy and survival outcomes.
MicroRNAs (miRNAs/miRs) are endogenous non-coding
RNAs 18–23 nucleotides in length that can regulate the expression
of non-coding and coding RNAs (6,7). In
human cells, a single miRNA can directly regulate a large number of
RNAs (8). Thus, dysregulated miRNA
expression can result in the aberrant expression levels of coding
RNAs in cancer cells (5). In
addition, dysregulation of miRNA expression may result in the
acquisition of malignant properties, leading to cancer progression,
metastasis and treatment resistance (9–12). Ke
et al (13) revealed that
miR-148b serves an inhibitory role in hepatocellular carcinoma
(HCC) and that the miR-148b-CSF1 axis induces tumor-associated
macrophage infiltration, thus promoting HCC. Han et al
(14) demonstrated that miR-338-5p
negatively regulates the inhibitor of DNA binding-1, altering 5-FU
chemoresistance and suppressing the metastasis of esophageal
squamous cell carcinoma. Additionally, it has been revealed that
miR-1231 expression is downregulated in pancreatic cancer and
serves a role in the TNM stage of pancreatic cancer, suggesting
that miR-1231 exhibits an inhibitory role on the metastasis and
development of pancreatic cancer (15).
Non-SMC condensin I complex subunit G (NCAPG) is a
subunit of the condensed protein complex, which is responsible for
the condensation and stabilization of chromosomes during mitosis
and meiosis (16). According to Gene
Ontology (GO) analysis, the pathways associated with NCAPG are
involved in cell cycle progression, mitosis and concentration of
cell cycle chromosomes in the prophase and metaphase (17). An increasing number of studies has
revealed increased NCAPG expression in prostate cancer, glioma and
lung cancer, indicating the notable involvement of NCAPG in various
biological functions (18–20). For example, Arai et al
(19) indicated that NCAPG was
regulated by miR-99a-3p and that its overexpression was involved in
the pathogenesis of castration-resistant prostate cancer. Liang
et al (20) revealed that
CENPE, KIF14 and NCAPG were direct targets of miR-137 or
miR-6500-3p, and that knockdown of CENPE, KIF14 or NCAPG combined
with temozolomide treatment resulted in a combined suppressive
effect on pediatric high-grade glioma cell proliferation. Studies
have revealed that upregulated NCAPG expression is associated with
a poor prognosis in patients with prostate cancer (21). Ke et al (13) demonstrated that small homologous
RNA-mediated knockdown of NCAPG significantly impaired cell
viability, caused aberrant mitotic division, fragmented the
mitochondrial network and increased cell death. Notably,
upregulated NCAPG expression is significantly associated with a
poor overall and disease-free survival in patients with HCC
(22). However, to the best of our
knowledge, previous studies on the role of NCAPG in breast cancer
and the potential mechanisms by which NCAPG affects breast cancer
are insufficient (23), and the
association between the expression levels of NCAPG and prognosis in
breast cancer has not been determined.
In the present study, NCAPG expression was assessed
in breast cancer. Subsequently, the prognostic effect of NCAPG in
patients with breast cancer based on their clinicopathological
characteristics was determined. Additionally, the underlying
mechanisms by which NCAPG regulated breast cancer development and
progression were determined.
Materials and methods
Human protein atlas database
analysis
Expression levels of NCAPG protein in different
human normal and cancer tissues were determined using the Human
Protein Atlas database (https://www.proteinatlas.org/) (24).
UALCAN database analysis
The UALCAN database (ualcan.path.uab.edu/) is an interactive network
resource that provides a convenient method to obtain open cancer
transcriptome data from The Cancer Genome Atlas (TCGA) (25). In the present study, the gene
expression levels and the correlations between two genes were
assessed using the UALCAN database. The UALCAN database performed
the statistical analysis using a log-rank test. P<0.05 was
considered to indicate a statistically significant difference.
Breast cancer gene expression
miner
Breast cancer gene expression miner (Bc-GenExMiner;
bcgenex.centregauducheau.fr/) is an online platform that can
analyze gene expression, prognosis and the associations in breast
cancer (26,27). Based on the different
clinicopathological characteristics of patients with breast cancer,
Bc-GenExMiner was used to detect the expression levels of NCAPG in
breast cancer. In addition, Bc-GenExMiner calculated the
correlation between NCAPG and cell division cyclin 25 homolog C
(CDC25C) expression. P<0.05 was considered to indicate a
statistically significant difference.
Kaplan-Meier Plotter database
analysis
Kaplan-Meier Plotter (kmplot.com/) is
based on data obtained from Gene Expression Omnibus, which includes
data on gene expression and survival information of patients with
cancer (28). Two probes
(218662_s_at and 218663_at) were used to analyze the association
between NCAPG expression and the overall survival, relapse-free
survival, distant metastasis-free survival and post-progression
survival rates in patients with breast cancer. Briefly, NCAPG was
selected in the database, and the Kaplan-Meier survival curves were
plotted. The cases were classified into low and high expression
groups based on the median NCAPG expression. Subsequently, the
cohorts were compared using Kaplan-Meier survival plots, and the
online tool calculated the hazard ratio (HR), 95% CI and log-rank
P-values. Additionally, the Kaplan Meier-Plotter database was used
to evaluate the prognostic values of the predicted miRNAs in breast
cancer. A log-rank P<0.05 was considered to indicate a
statistically significant difference.
PrognoScan database analysis
The PrognoScan database
(dna00.bio.kyutech.ac.jp/PrognoScan/) collates clinically annotated
publicly available cancer microarray datasets for bladder, brain,
breast, blood, esophageal, colorectal and head and neck cancer
(29). In the present study,
PrognoScan was used to evaluate the biological association between
the expression levels and prognosis of NCAPG in breast cancer. The
datasets were as follows: GSE19615 (30), GSE12276 (31), GSE6532-GPL570 (32), GSE9195 (33), GSE12093 (34), GSE11121 (35), GSE1378 (36), GSE1379 (37), GSE2034 (38), GSE1456-GPL96 (39), GSE7378 (40), E-TABM-158 (41), GSE3494-GPL96 (42), GSE4922-GPL96 (43), GSE2990 (33) and GSE7390 (44). A log-rank P<0.05 was considered to
indicate a statistically significant difference.
starBase database
The starBase database (starbase.sysu.edu.cn/) is an open-source platform that
systematically defines the RNA-RNA and protein-RNA interactions
from crosslinking-immunoprecipitation and high-throughput
sequencing (45). In the present
study, the upstream miRNAs of NCAPG were predicted using starBase,
and the negative correlations between each miRNA expression and
NCAPG expression were identified using starBase. R<-0.1 and
P<0.05 were used as the thresholds for identification of
potential miRNAs targeted by NCAPG for further investigation.
OncomiR database analysis
OncomiR (oncomir.org/)
is an online resource for exploring dysregulated miRNA expression
in several types of cancer. OncomiR acquires RNA-sequencing (seq),
miRNA-seq and clinical data from TCGA, then performs a systematic
statistical analysis to identify the dysregulated miRNAs associated
with the development and progression of several types of cancer
(46). OncomiR was used to explore
the expression levels of miRNAs in breast cancer. A log-rank
P<0.05 was considered to indicate a statistically significant
difference.
Gene expression profiling interactive
analysis (GEPIA) database
GEPIA (gepia.cancer-pku.cn/) is a database that
provides key interactive and customizable functions, including
correlation analysis, differential expression analysis, patient
survival analysis, dimensionality reduction and similar gene
detection analysis (47). In the
present study, GEPIA was used to determine the co-expressed genes
of NCAPG in breast cancer.
cBio cancer genomics portal
(cBioPortal) database analysis
cBioPortal (cbioportal.org/) is an open online access resource for
interactive analysis of multidimensional datasets for cancer
genomics, acquiring data from >5,000 tumor samples from 172
cancer studies (48). cBioPortal was
used to obtain the genes that were co-expressed with NCAPG in
breast cancer, and the correlated genes with a Pearson's r>0.3
were selected for further analysis.
Enrichr database analysis
Enrichr (amp.pharm.mssm.edu/Enrichr/) is a comprehensive
application, including new gene-set libraries, an optional approach
to rank the enriched terms and a JavaScript library. Data-driven
documents offer multiple interactive visualization approaches to
show the enrichment results. The top 10 enriched GO items and
pathways were displayed using Enrichr.
Human tissue samples and cell
culture
A total of 24 pairs of breast cancer and adjacent
normal tissues (>1 cm from the tumor) were collected from the
Tongji Medical College of Huazhong Science and Technology
University (Wuhan, China) between May 2019 and June 2019.
Additionally, 113 paired cancer samples and adjacent tissue data
were obtained from TCGA database. The 24 pairs of fresh breast
cancer samples were acquired from excision surgery and immediately
preserved in RNAlater Stabilization Solution (Qiagen GmbH)
overnight at 4°C, and subsequently stored at −80°C until total RNA
was extracted. All patients were female, with a median age of 57
years (range, 35–67 years). None of the patients had previously
received pre-operative chemotherapy or radiotherapy. All procedures
involving human participants in the present study were performed
according to the ethical standards of the Ethics Committee of
Tongji Medical College of Huazhong Science and Technology
University, and written informed consent was provided by each
participant. The human breast cancer MCF-7 cell line was obtained
from The Cell Bank of Type Culture Collection of the Chinese
Academy of Sciences. MCF-7 cells were cultured in DMEM (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% FBS
(Sigma-Aldrich; Merck KGaA) at 37°C with 5% CO2.
Cell transfection
Small interfering (si)RNAs targeting NCAPG and a
scrambled control siRNA used as the negative control (NC) were
purchased from Guangzhou RiboBio Co., Ltd. NCAPG siRNAs and the NC
(50 nM) were transfected using Lipofectamine® 3000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) into cells,
according to the manufacturer's protocol. Each well was transfected
with 2 µg siRNAs. After 48 h of transfection at 37°C, cells were
collected for subsequent experiments. Expression levels of NCAPG
following transfection were detected using reverse
transcription-quantitative (RT-qPCR). The sequences of siRNAs and
NC used are listed in Table SI.
RT-qPCR
Total RNA from the human samples or the cell lines
was extracted using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). Total RNA was reverse-transcribed
according to the manufacturer's protocol using
HiScript®II RT SuperMix (Vazyme Biotech Co., Ltd.). qPCR
was performed in a Roche LightCycle480 II Real-Time PCR Detection
System using SYBR Premix Ex Taq (cat. no. RR420A; Takara
Biotechnology Co., Ltd.). The following thermocycling conditions
were used for qPCR: Initial denaturation at 95°C for 10 min,
followed by 45 cycles at 92°C for 15 sec and 72°C for 5 min. All
the measurements were performed in triplicate. The sequences of the
primers used in the present study are listed in Table SI. Quantitative mRNA data were
normalized and presented as a ratio to GAPDH, calculated using the
2−ΔΔCq method (49).
Flow cytometry
For cell cycle distribution, transfected cells were
stained with propidium iodide (BD Biosciences) and fixed in
ice-cold 75% ethanol overnight at 4°C. After fixing, the cells were
washed and resuspended twice in PBS, and were then incubated with
propidium iodide according to the manufacturer's protocol (BD
Biosciences) and RNase for 30 min at room temperature. The cells
were then analyzed using a FACSCalibur flow cytometer (BD
Biosciences) with a laser beam at 488 nm. The data were analyzed
using FlowJo v7.6 (FlowJo LLC).
Western blotting
Cells were lysed using RIPA lysis buffer (Thermo
Fisher Scientific, Inc.) containing protease inhibitors. Total
protein concentration was measured using a Bradford assay.
Subsequently, 20 µg protein/lane was separated via 10% SDS-PAGE and
transferred to nitrocellulose membranes. Membranes were blocked at
room temperature in TBS-Tween (0.1% Tween-20) with 5% skimmed milk
for 1 h and then incubated overnight at 4°C with rabbit anti-CDC25C
(1:200; cat. no. ab32444; Abcam) and mouse anti-GAPDH (1:1,000;
cat. no. ab8245; Abcam) primary antibodies. Subsequently, the
membranes were washed with Tris-HCl solution + 0.1% Tween-20 three
times for 10 min and were incubated with horseradish
peroxidase-conjugated anti-rabbit (1:3,000; cat. no. ab150077;
Abcam) and anti-mouse (1:3,000; cat. no. ab150113; Abcam) secondary
antibodies for 1 h at room temperature. Signals were visualized
using an enhanced chemiluminescence reagent (Thermo Fisher
Scientific, Inc.) and imaged using a GelDocXR+ (Bio-Rad
Laboratories, Inc.). GAPDH was used as the loading control.
Statistical analysis
Data are presented as the mean ± standard error of
the mean. Statistical analysis was performed using GraphPad Prism
5.0 (GraphPad Software, Inc.). Unpaired Student's t-test was used
to analyze the differences between two groups and paired Student's
t-test was used to compare NCAPG expression between paired cancer
and normal tissues. Multiple comparisons were analyzed using a
one-way ANOVA with Tukey's post-hoc test. The χ2 test
was used to analyze the association between NCAPG expression and
clinicopathological features. The correlations between CDC25C and
NCAPG expression were analyzed using Spearman's and Pearson's
correlation analyses. Kaplan-Meier survival analysis and
univariate/multivariate Cox proportional hazards analyses with
log-rank method were used to compare the survival between the high
and low NCAPG expression groups. The enrichment pathways of the
co-expressed genes of NCAPG were analyzed using Fisher's exact test
to obtain the Cox P-value. P<0.05 was considered to indicate a
statistically significant difference.
Results
NCAPG expression is upregulated in
breast cancer tissues
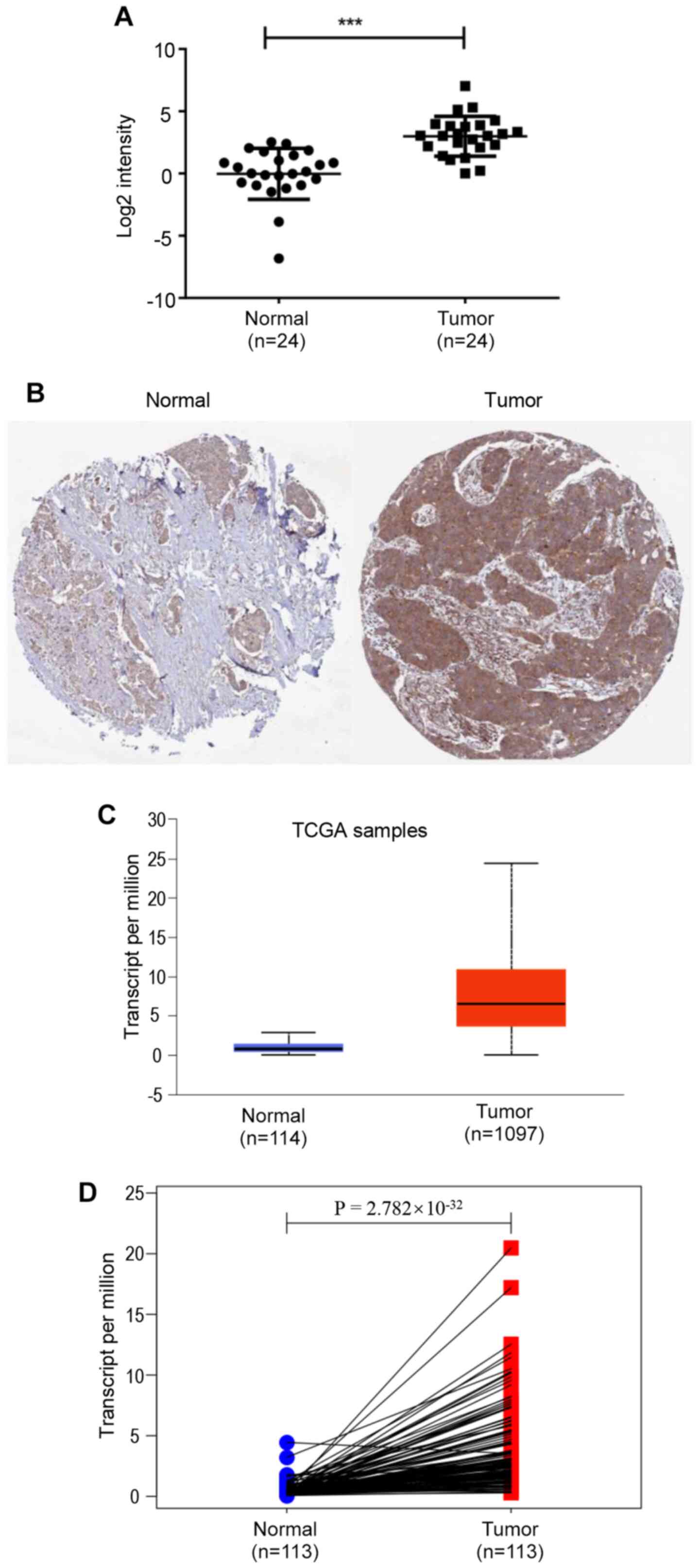
In the present study, the expression levels of NCAPG
in 24 pairs of breast cancer and adjacent normal breast tissues
were determined using RT-qPCR. NCAPG expression was significantly
upregulated in breast cancer tissues compared with in normal breast
tissues (Fig. 1A). Subsequently,
NCAPG expression in breast cancer was analyzed using the UALCAN
database based on data obtained from TCGA, revealing that NCAPG
mRNA expression was significantly higher in breast cancer tissues
compared with in normal tissues (Fig.
1C). Furthermore, the results from 113 paired cancer samples
and adjacent tissues from TCGA database demonstrated that NCAPG
mRNA expression was significantly upregulated in tumor tissues
compared with in para-cancerous tissues (Fig. 1D). Additionally, the protein
expression levels of NCAPG in breast cancer tissues were higher
compared with in normal breast tissues (Fig. 1B), analyzed using the GEPIA database.
Subsequently, the association between NCAPG expression and
clinicopathological characteristics of patients with breast cancer
was analyzed using TCGA breast cancer data. The results revealed
that NCAPG expression was significantly associated with estrogen
receptor (ER) status (P<0.0001), progesterone receptor (PR)
status (P<0.0001), HER2 status (P=0.0034), T stage (P<0.001)
and TNM pathological stage (50)
(P=0.0002) (Table I). The current
results indicated that NCAPG expression was significantly
upregulated in breast cancer and was associated with the
progression of breast cancer.
 | Table I.Association between NCAPG expression
and clinicopathological characteristics of patients with breast
cancer in The Cancer Genome Atlas. |
Table I.
Association between NCAPG expression
and clinicopathological characteristics of patients with breast
cancer in The Cancer Genome Atlas.
|
|
| NCAPG
expression |
|
|---|
|
|
|
|
|
|---|
| Features | Cases, n | High/low, n | P-value |
|---|
| Age at diagnosis,
years |
|
≤51 | 349 | 184/165 | 0.2035 |
|
>51 | 686 | 333/353 |
|
| Estrogen receptor
status |
|
Positive | 765 | 311/454 | <0.0001 |
|
Negative | 221 | 180/41 |
|
| Progesterone
receptor status |
|
Positive | 663 | 255/408 | <0.0001 |
|
Negative | 320 | 235/85 |
|
| HER2 status |
|
Positive | 173 | 104/69 | 0.0034 |
|
Negative | 721 | 344/377 |
|
| Pathological
stage |
| I | 171 | 63/108 | 0.0002 |
|
II/III/IV | 841 | 443/398 |
|
| T stage |
| 1 | 268 | 102/166 | <0.0001 |
|
2/3/4 | 764 | 413/351 |
|
| N stage |
|
N0/N1 | 823 | 413/410 | 0.9643 |
|
N2/N3 | 192 | 96/96 |
|
| M stage |
| M0 | 861 | 452/409 | 0.6740 |
| M1 | 21 | 12/9 |
|
Association between NCAPG expression
and clinicopathological characteristics of patients with breast
cancer
NCAPG expression and its association with the
different clinicopathological parameters of patients with breast
cancer were analyzed using the Bc-GenExMiner database. The average
expression levels of NCAPG were significantly upregulated in
patients <51 years old (Fig. 2A),
nodal-positive (Fig. 2B),
ER− (Fig. 2C),
PR− (Fig. 2D) and
HER2+ positive (Fig.
2E).
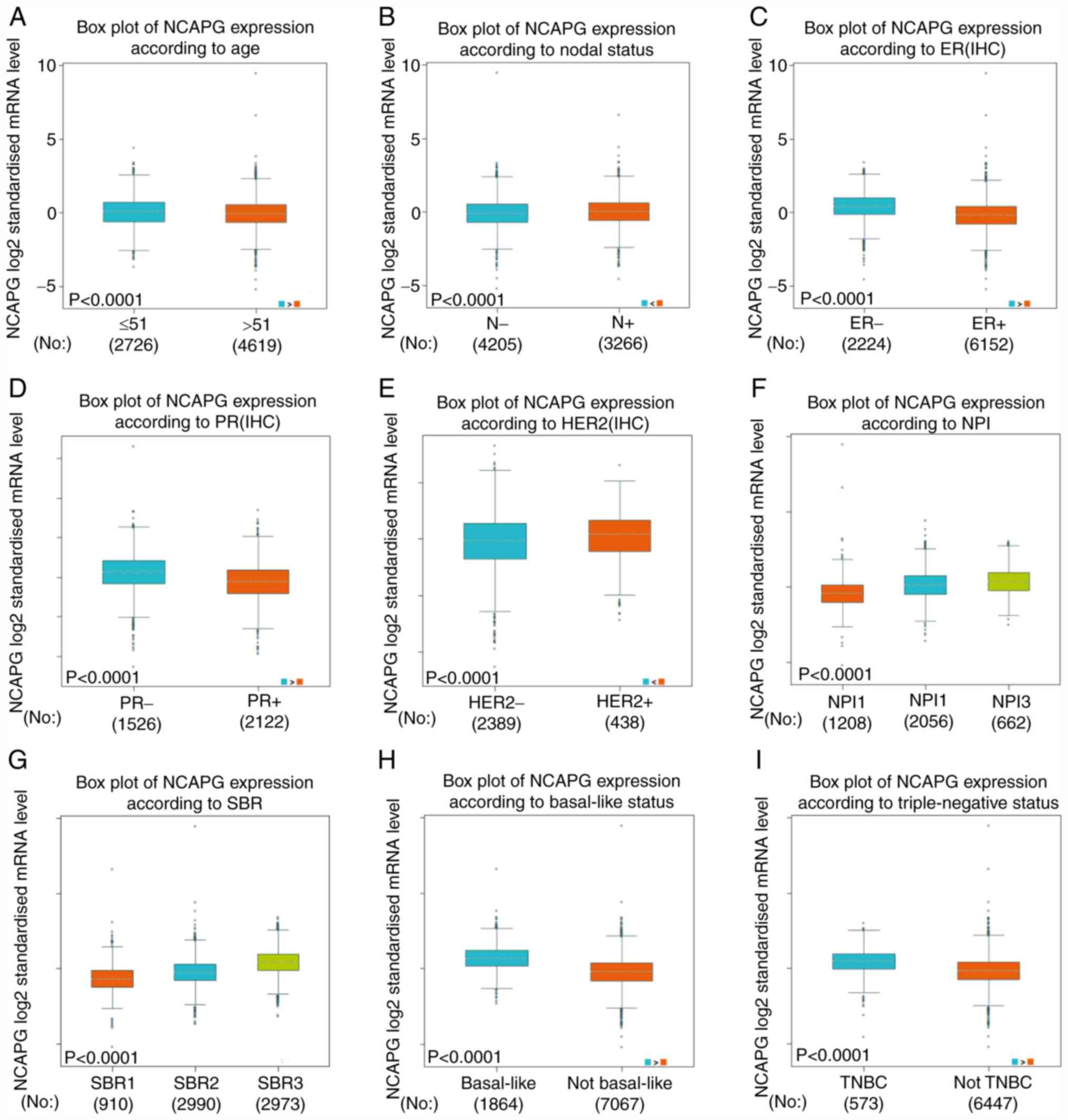 | Figure 2.Differences in NCAPG expression in
patients with breast cancer based on different clinicopathological
features from The Cancer Genome Atlas. NCAPG expression according
to (A) age, (B) N status, (C) ER status, (D) PR status, (E) HER2
status, (F) NPI score, (G) SBR grade, (H) basal-like status and (I)
triple-negative status. ER, estrogen receptor; PR, progesterone
receptor; NPI, Nottingham Prognostic Index; SBR,
Scarff-Bloom-Richardson; NCAPG, non-SMC condensin I complex subunit
G; IHC, immunohistochemistry; TNBC, triple-negative breast cancer;
N, nodal. |
There was a positive association between the
expression levels of NCAPG and the Nottingham Prognostic Index
(NPI) score (51) and
Scarff-Bloom-Richardson (SBR) grade (52), as shown in Fig. 2F and G. Additionally, the average
NCAPG expression was significantly increased in basal-like
(Fig. 2H) and triple negative breast
cancer cases compared with in non-basal-like and
non-triple-negative breast cancer cases, respectively (Fig. 2H and I).
The aforementioned results suggested that patients
with higher expression levels of NCAPG were more likely to develop
clinically advanced or more aggressive breast cancer types.
Patients with breast cancer with high
expression levels of NCAPG have a poor prognosis
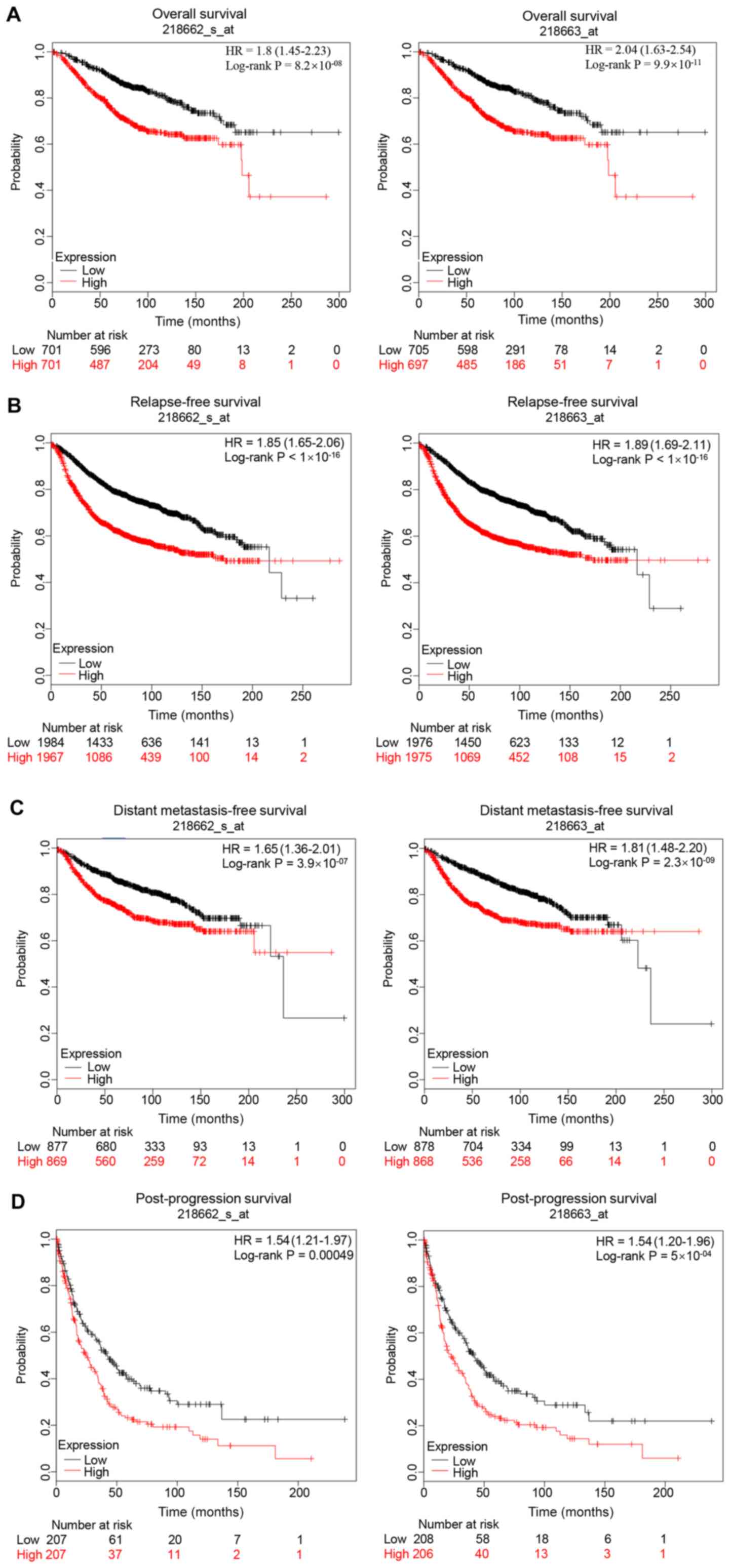
The prognostic value of NCAPG in breast cancer was
evaluated. The Kaplan Meier-Plotter database contained two probes
(218662_s_at and 218663_at) of NCAPG expression. As shown in
Fig. 3, high expression levels of
NCAPG resulted in a less favorable prognosis in patients with
breast cancer compared with low expression levels, including less
favorable overall survival, relapse-free survival, distant
metastasis-free survival and post-progression survival rates in
both probes. In addition, the association between NCAPG expression
in breast cancer tissues and prognosis was determined using the
PrognoScan database. The results revealed that the expression
levels of NCAPG were negatively associated with overall survival
(GSE1456-GPL96), disease-specific survival (GSE1456-GPL96),
relapse-free survival (GSE12276, GSE6532-GPL570 and GSE1456-GPL96),
disease-free survival (GSE7378) and distant metastasis-free
survival (GSE6532-GPL570, GSE12093, GSE11121 and GSE2034) (Table II).
| Table II.Association between NCAPG expression
and prognosis in patients with breast cancer using the PrognoScan
database. |
Table II.
Association between NCAPG expression
and prognosis in patients with breast cancer using the PrognoScan
database.
| Dataset | Endpoint | Patient number | Probe ID | Cox P-value | Hazard ratio (95%
CI) |
|---|
| GSE19615 | Distant
metastasis-free survival | 115 | 218663_at | 0.3759 | 1.41
(0.66–3.05) |
| GSE19615 | Distant
metastasis-free survival | 115 | 218662_s_at | 0.5339 | 1.32
(0.55–3.18) |
| GSE12276 | Relapse-free
survival | 204 | 218663_at | 0.0001 | 1.53
(1.24–1.90) |
| GSE12276 | Relapse-free
survival | 204 | 218662_s_at |
6.88×10−05 | 1.51
(1.23–1.85) |
| GSE6532-GPL570 | Distant
metastasis-free survival | 87 | 218662_s_at | 0.1155 | 1.37
(0.93–2.03) |
| GSE6532-GPL570 | Distant
metastasis-free survival | 87 | 218663_at | 0.0361 | 1.47
(1.03–2.11) |
| GSE6532-GPL570 | Relapse-free
survival | 87 | 218662_s_at | 0.1155 | 1.37
(0.93–2.03) |
| GSE6532-GPL570 | Relapse-free
survival | 87 | 218663_at | 0.0361 | 1.47
(1.03–2.11) |
| GSE9195 | Distant
metastasis-free survival | 77 | 218663_at | 0.4349 | 1.30
(0.67–2.50) |
| GSE9195 | Distant
metastasis-free survival | 77 | 218662_s_at | 0.1658 | 1.67
(0.81–3.43) |
| GSE9195 | Relapse-free
survival | 77 | 218662_s_at | 0.2247 | 1.49
(0.78–2.83) |
| GSE9195 | Relapse-free
survival | 77 | 218663_at | 0.4998 | 1.22
(0.68–2.21) |
| GSE12093 | Distant
metastasis-free survival | 136 | 218662_s_at | 0.0062 | 3.38
(1.41–8.09) |
| GSE12093 | Distant
metastasis-free survival | 136 | 218663_at | 0.0123 | 2.52
(1.22–5.19) |
| GSE11121 | Distant
metastasis-free survival | 200 | 218662_s_at |
4.52×10−05 | 3.01
(1.77–5.12) |
| GSE11121 | Distant
metastasis-free survival | 200 | 218663_at | 0.0099 | 1.78
(1.15–2.75) |
| GSE1378 | Relapse-free
survival | 60 | 21899 | 0.7551 | 1.07
(0.71–1.59) |
| GSE1379 | Relapse-free
survival | 60 | 21899 | 0.7284 | 1.08
(0.69–1.71) |
| GSE2034 | Distant
metastasis-free survival | 286 | 218662_s_at | 0.0330 | 1.43
(1.03–1.98) |
| GSE2034 | Distant
metastasis-free survival | 286 | 218663_at | 0.0025 | 1.67
(1.20–2.34) |
| GSE1456-GPL96 | Overall
survival | 159 | 218662_s_at | 0.0008 | 2.91
(1.56–5.43) |
| GSE1456-GPL96 | Disease-specific
survival | 159 | 218663_at | 0.0006 | 2.67
(1.52–4.71) |
| GSE1456-GPL96 | Overall
survival | 159 | 218663_at | 0.0020 | 2.06
(1.30–3.27) |
| GSE1456-GPL96 | Relapse-free
survival | 159 | 218662_s_at | 0.0003 | 3.16
(1.70–5.88) |
| GSE1456-GPL96 | Disease-specific
survival | 159 | 218662_s_at | 0.0002 | 3.99
(1.90–8.34) |
| GSE1456-GPL96 | Relapse-free
survival | 159 | 218663_at | 0.0005 | 2.26
(1.42–3.57) |
| GSE7378 | Disease-free
survival | 54 | 218663_at | 0.0335 | 1.92
(1.05–3.52) |
| GSE7378 | Disease-free
survival | 54 | 218662_s_at | 0.0709 | 1.74
(0.95–3.18) |
| E-TABM-158 | Overall
survival | 117 | 218662_s_at | 0.2037 | 0.80
(0.56–1.13) |
| E-TABM-158 | Distant
metastasis-free survival | 117 | 218662_s_at | 0.6812 | 1.09
(0.72–1.66) |
| E-TABM-158 | Relapse-free
survival | 117 | 218663_at | 0.3616 | 0.84
(0.58–1.22) |
| E-TABM-158 | Disease-specific
survival | 117 | 218663_at | 0.0900 | 0.67
(0.42–1.06) |
| E-TABM-158 | Distant
metastasis-free survival | 117 | 218663_at | 0.7572 | 1.07
(0.70–1.63) |
| E-TABM-158 | Overall
survival | 117 | 218663_at | 0.3616 | 0.84
(0.58–1.22) |
| E-TABM-158 | Disease-specific
survival | 117 | 218662_s_at | 0.0352 | 0.63
(0.42–0.97) |
| E-TABM-158 | Relapse-free
survival | 117 | 218662_s_at | 0.2038 | 0.80
(0.56–1.13) |
| GSE3494-GPL96 | Disease-specific
survival | 236 | 218662_s_at | 0.0003 | 2.45
(1.51–3.96) |
| GSE3494-GPL96 | Disease-specific
survival | 236 | 218663_at | 0.0016 | 1.80
(1.25–2.61) |
| GSE4922-GPL96 | Disease-free
survival | 249 | 218663_at | 0.0007 | 1.61
(1.22–2.13) |
| GSE4922-GPL96 | Disease-free
survival | 249 | 218662_s_at |
4.93×10−05 | 2.24
(1.52–3.32) |
| GSE2990 | Distant
metastasis-free survival | 125 | 218662_s_at | 0.0760 | 1.42
(0.96–2.10) |
| GSE2990 | Relapse-free
survival | 125 | 218662_s_at | 0.1234 | 1.27
(0.94–1.72) |
| GSE2990 | Relapse-free
survival | 62 | 218663_at | 0.1058 | 1.65
(0.90–3.03) |
| GSE2990 | Distant
metastasis-free survival | 54 | 218662_s_at | 0.0024 | 2.45
(1.37–4.37) |
| GSE2990 | Distant
metastasis-free survival | 125 | 218663_at | 0.0797 | 1.51
(0.95–2.39) |
| GSE2990 | Relapse-free
survival | 125 | 218663_at | 0.1058 | 1.37
(0.94–2.00) |
| GSE2990 | Distant
metastasis-free survival | 54 | 218663_at | 0.1420 | 1.74
(0.83–3.67) |
| GSE2990 | Relapse-free
survival | 62 | 218662_s_at | 0.0037 | 2.07
(1.27–3.38) |
| GSE7390 | Distant
metastasis-free survival | 198 | 218663_at | 0.1751 | 1.15
(0.94–1.42) |
| GSE7390 | Overall
survival | 198 | 218663_at | 0.0655 | 1.24
(0.99–1.55) |
| GSE7390 | Relapse-free
survival | 198 | 218662_s_at | 0.2174 | 1.15
(0.92–1.44) |
| GSE7390 | Relapse-free
survival | 198 | 218663_at | 0.1340 | 1.14
(0.96–1.34) |
| GSE7390 | Distant
metastasis-free survival | 198 | 218662_s_at | 0.2798 | 1.16
(0.88–1.54) |
| GSE7390 | Overall
survival | 198 | 218662_s_at | 0.2032 | 1.21
(0.90–1.63) |
Identification of miRNAs that
potentially regulate NCAPG
The starBase database was used to predict the
upstream miRNAs of mRNAs. The miRNAs that potentially regulated
NCAPG were predicted using starBase. A total of 27 miRNAs
(hsa-miR-488-3p, hsa-miR-181b-5p, hsa-miR-181a-5p, hsa-miR-15a-5p,
hsa-miR-494-3p, hsa-miR-543, hsa-miR-495-3p, hsa-miR-422a,
hsa-miR-497-5p, hsa-miR-27a-3p, hsa-miR-23a-3p, hsa-miR-181c-5p,
hsa-miR-181d-5p, hsa-miR-128-3p, hsa-miR-124-3p, hsa-miR-124-5p,
hsa-miR-15b-5p, hsa-miR-378a-3p, hsa-miR-340-5p, hsa-miR-340-3p,
hsa-miR-590-5p, hsa-miR-27b-3p, hsa-miR-374b-5p, hsa-miR-424-5p,
hsa-miR-506-3p, hsa-miR-374a-5p and hsa-miR-23b-3p) were identified
to potentially regulate NCAPG expression. It is well known that
certain miRNAs negatively regulate mRNA expression (53–55).
Thus, the correlations (using Pearson's correlation analysis)
between the expression levels of the predicted miRNAs and NCAPG
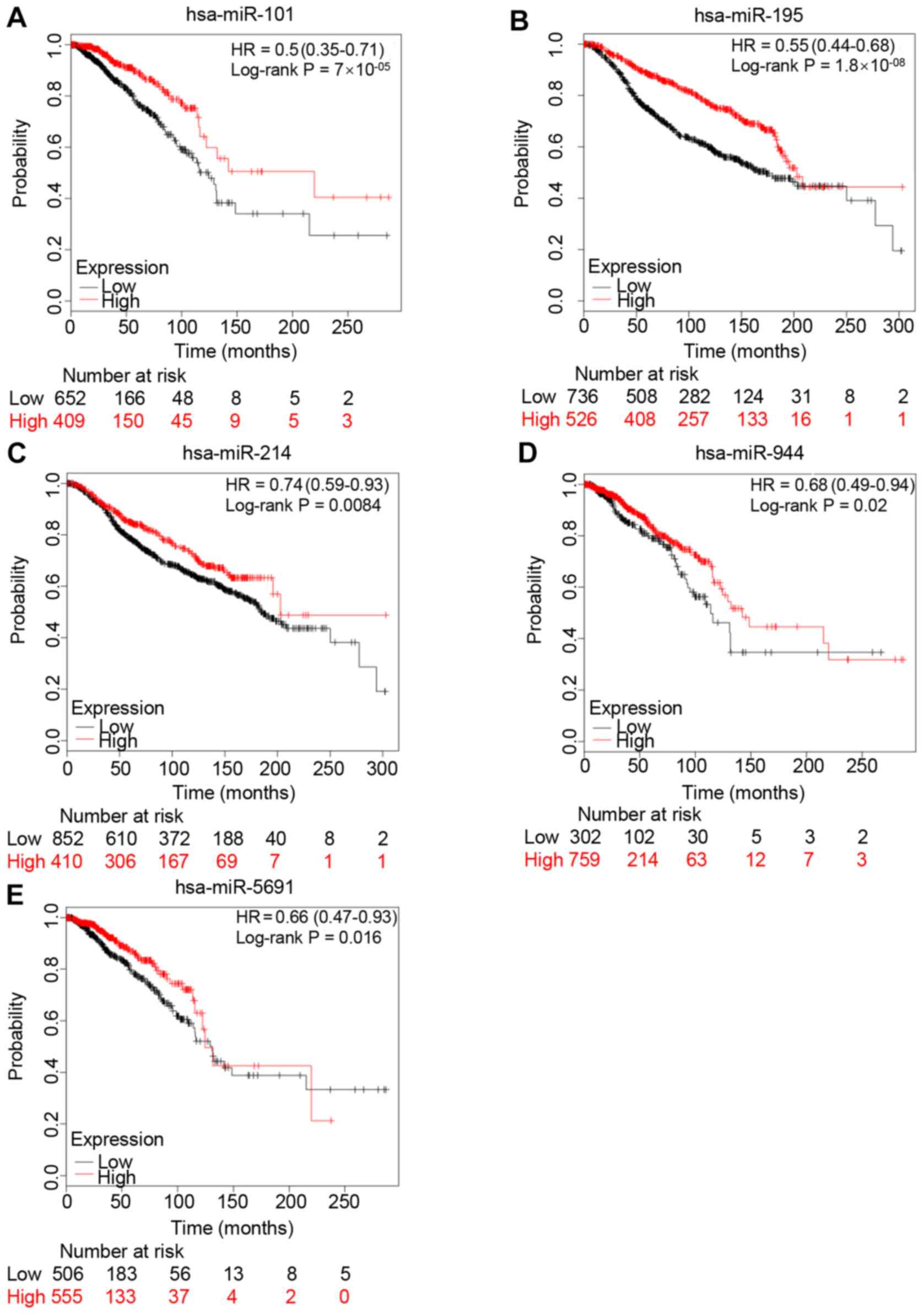
were explored using TCGA. The results revealed that 12 miRNAs were
inversely correlated with NCAPG expression (Table III). Subsequently, the prognostic
values of the 12 miRNAs in patients with breast cancer were
assessed using Kaplan-Meier Plotter. Low expression levels of
miR-101-3p, miR-195-5p, miR-214-3p, miR-944 and miR-5691 were
significantly associated with a poor prognosis in patients with
breast cancer, compared with high expression levels (Fig. 4). The prognostic analyses of the
other miRNAs are shown in Fig. S1.
Overall, the present results and previous studies revealed that the
expression levels of these 4 miRNAs (miR-101-3p, miR-195-5p,
miR-214-3p and miR-944) were downregulated in breast cancer,
resulting in a poor prognosis, and were also negatively correlated
with NCAPG expression.
| Table III.Pearson correlation between predicted
miRNAs and non-SMC condensin I complex subunit G expression. |
Table III.
Pearson correlation between predicted
miRNAs and non-SMC condensin I complex subunit G expression.
| Predicted
miRNA | R | P-value |
|---|
| miR-101-3p | −0.264 |
8.14×10−19 |
| miR-195-5p | −0.210 |
2.94×10−12 |
| miR-214-3p | −0.104 |
5.96×10−04 |
| miR-369-3p | −0.108 |
3.67×10−04 |
| miR-381-3p | −0.165 |
4.55×10−08 |
| miR-410-3p | −0.201 |
2.17×10−11 |
| miR-432-5p | −0.167 |
3.43×10−08 |
| miR-494-3p | −0.128 |
2.52×10−05 |
| miR-543 | −0.102 |
8.04×10−04 |
| miR-655-3p | −0.174 |
8.54×10−09 |
| miR-944 | −0.140 |
3.42×10−06 |
| miR-5691 | −0.234 |
6.31×10−15 |
GO functional annotation and pathway
enrichment analysis of co-expressed genes of NCAPG
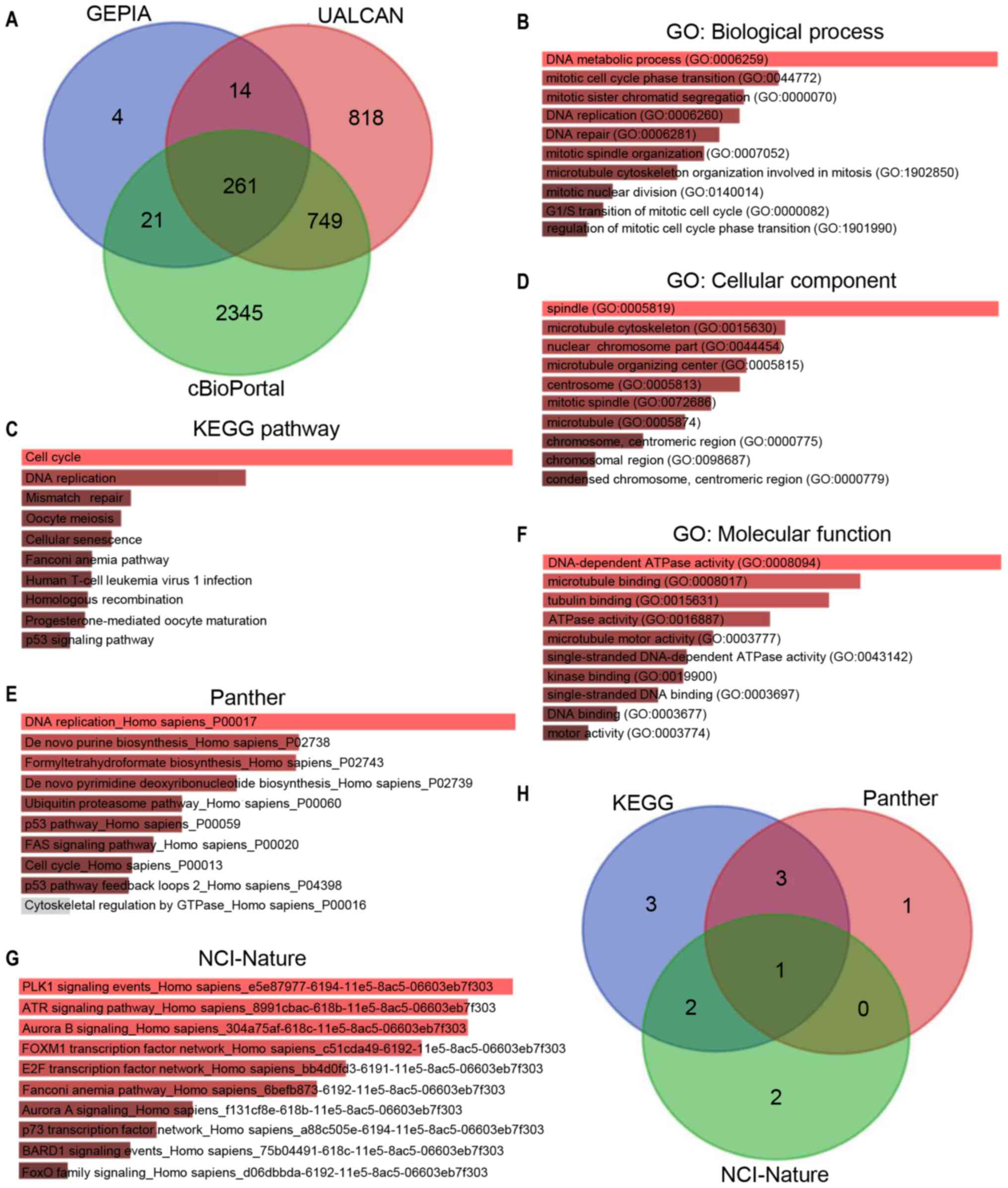
GEPIA, UALCAN and cBioPortal were used to analyze
the co-expressed genes of NCAPG. A total of 261 co-expressed genes
of NCAPG were identified in these three databases (Fig. 5A). To determine the functions of the
co-expressed genes, GO functional annotation and pathway enrichment
analysis was performed using Enrichr. The functional annotation was
stratified into molecular function, cellular component and
biological process. Additionally, the cell signaling pathways from
the National Cancer Institute (NCI)-Nature, KEGG and Panther were
analyzed for pathway enrichment using the Enrichr database. As
shown in Fig. 5B, D and F, the top
10 enriched GO terms in each category included ‘DNA metabolic
process’, ‘mitotic cell cycle phase transition’ and ‘mitotic sister
chromatid segregation’ in the biological process category;
‘spindle’, ‘microtubule cytoskeleton’ and ‘nuclear chromosome part’
in the cellular component category; and ‘DNA-dependent ATPase
activity’, ‘microtubule binding’ and ‘tubulin binding’ in the
molecular function category. Fig. 5C, E
and G presents the top 10 enriched signaling pathways (such as
‘cell cycle’, ‘DNA replication’ and ‘PLK1 signaling pathway’) in
the NCI-Nature, KEGG and Panther databases, respectively. The p53
signaling pathway was the intersection of the pathways that were
significantly enriched in the three databases (Table SII). Thus, the corresponding genes
of the p53 signaling pathway were analyzed in KEGG, Panther and
NCI-Nature pathways (Fig. 5H),
revealing a total of 12 genes [cyclin (CCN)A2, CCNB1, CCNB2, CCNE2,
CDC25C, CDK1, CDK2, checkpoint kinase (CHEK)1, CHEK2, G2
and S phase expressed 1 (GTSE1), ribonucleotide reductase
regulatory subunit M2 (RRM2) and S-phase kinase-associated protein
2 (SKP2)] enriched in the p53 signaling pathway.
Upregulated CDC25C expression is
associated with dysregulation of the p53 signaling pathway
In order to determine the effects of p53 signaling
on NCAPG-mediated progression of breast cancer, the UALCAN database
was used to assess the expression levels of the 12 genes enriched
in the p53 signaling pathway (Fig.
6A-L). The results revealed that SKP2 expression (Fig. 6L) was downregulated in breast cancer,
while CCNA2 (Fig. 6A), CCNB1
(Fig. 6B), CCNB2 (Fig. 6C), CCNE2 (Fig. 6D), CDC25C (Fig. 6E), CDK1 (Fig. 6F), CDK2 (Fig. 6G), CHEK1 (Fig. 6H), CHEK2 (Fig. 6I), GTSE1 (Fig. 6J) and RRM2 (Fig. 6K) expression was upregulated in
breast cancer compared with in normal breast tissue samples. The
prognostic value of these 12 genes was assessed using Kaplan-Meier
Plotter (Fig. 6M-X). High CDC25C
expression resulted in a significantly worse prognosis in patients
with breast cancer compared with low CDC25C expression (Fig. 6Q). For the other 11 genes, there were
no significant associations based on expression and survival.
Therefore, it was hypothesized that CDC25C expression was closely
associated with NCAPG expression in breast cancer. GEPIA, UALCAN,
bc-GenExMiner and cBioPortal were used to further detect the
positive correlation between CDC25C and NCAPG expression in breast
cancer (Fig. S2).
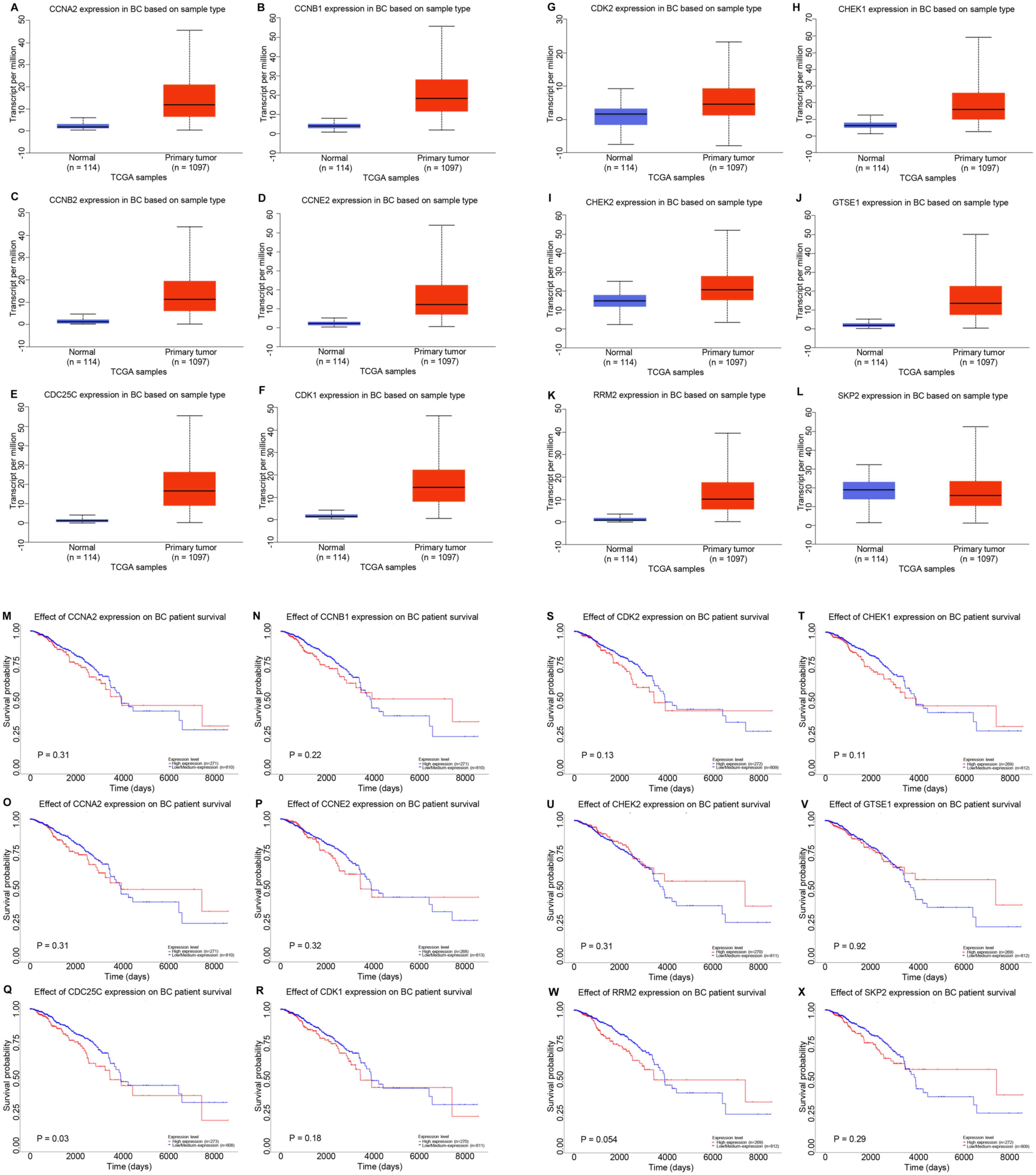 | Figure 6.Potential co-expressed genes of NCAPG
in breast cancer. Expression levels of (A) CCNA2, (B) CCNB1, (C)
CCNB2, (D) CCNE2, (E) CDC25C, (F) CDK1, (G) CDK2, (H) CHEK1, (I)
CHEK2, (J) GTSE1, (K) RRM2 and (L) SKP2 in breast cancer analyzed
using UALCAN. Prognostic roles of (M) CCNA2, (N) CCNB1, (O) CCNB2,
(P) CCNE2, (Q) CDC25C, (R) CDK1, (S) CDK2, (T) CHEK1, (U) CHEK2,
(V) GTSE1, (W) RRM2 and (X) SKP2 in breast cancer analyzed using
Kaplan-Meier Plotter. NCAPG, non-SMC condensin I complex subunit G;
BC, breast cancer; CCN, cyclin; CDC25C, cell division cyclin 25
homolog C; CHEK, checkpoint kinase; GTSE1, G2 and S
phase expressed 1; RRM2, ribonucleotide reductase regulatory
subunit M2; SKP2, S-phase kinase-associated protein 2; TCGA, The
Cancer Genome Atlas. |
To preliminarily assess the association between
CDC25C and NCAPG in breast cancer, the expression levels of CDC25C
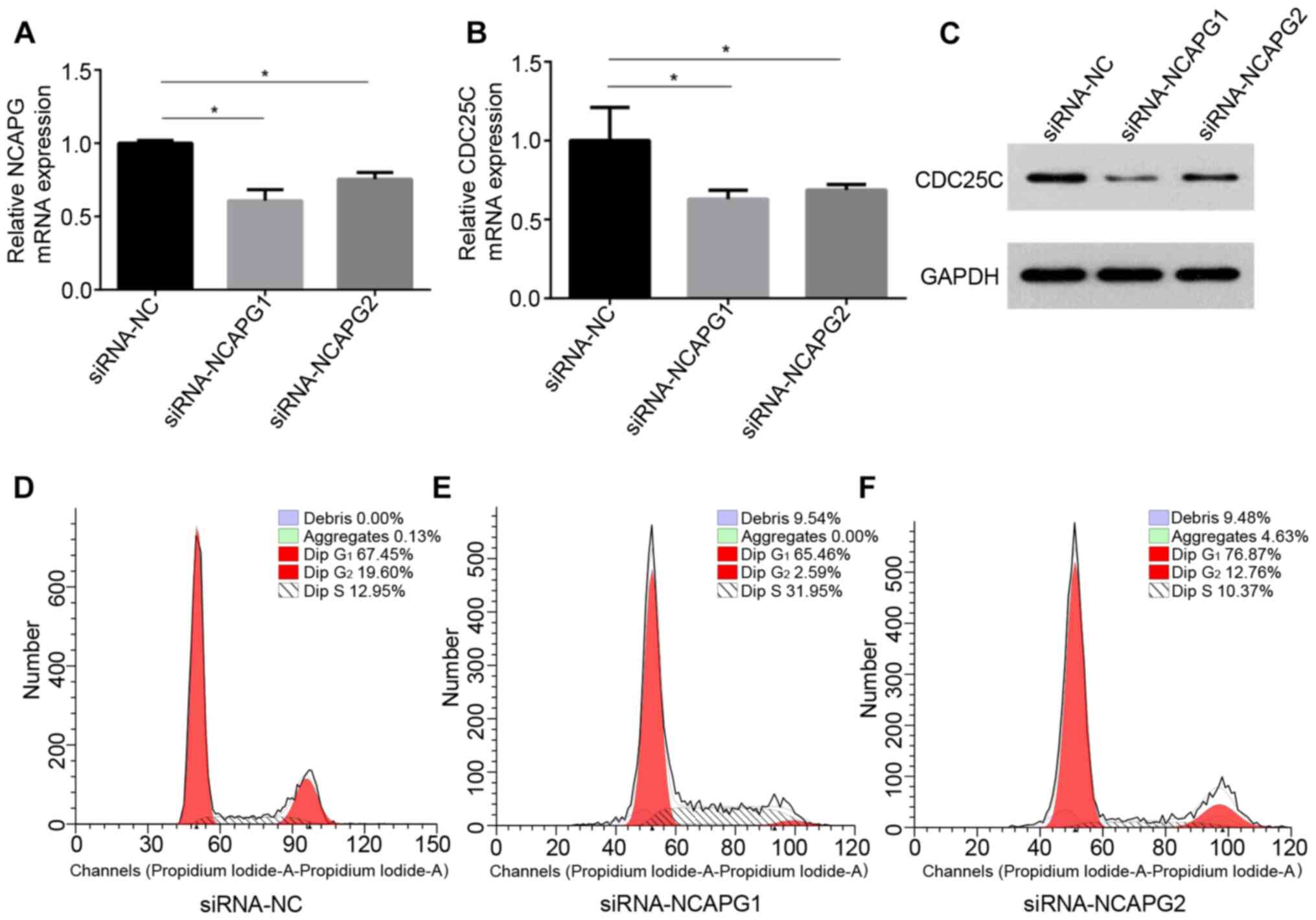
and NCAPG were determined following knockdown of NCAPG using
siRNA-NCAPG in the common breast cancer MCF-7 cell line. The
knockdown effects of siRNA-NCAPG are presented in Fig. 7A. Additionally, a significant
decrease in CDC25C mRNA and protein expression was observed
following NCAPG-knockdown (Fig. 7B and
C). Previous studies have demonstrated that CDC25C is a cell
cycle regulatory protein and serves an important role in the cell
cycle (56,57). Thus, the effects of NCAPG-knockdown
on cell cycle progression in MCF-7 cells were determined. Cell
cycle analysis revealed that NCAPG-knockdown resulted in an
increased proportion of cells in the S phase (31.95% with
siRNA-NCAPG1) and G1 phase (76.78% with siRNA-NCAPG2)
compared with the control group (S phase, 12.95% and G1
phase, 67.45%), suggesting that downregulation of NCAPG resulted in
S/M and G1/M arrest of MCF-7 cells (Fig. 7D-F). Overall, the current results
suggested that NCAPG may be associated with p53 signaling through
regulation of CDC25C, in turn promoting cell cycle progression in
breast cancer.
Discussion
NCAPG is a subunit of the condensin complex and is
hypothesized to serve important roles in the mitosis and meiosis
responsible for the condensation and stabilization of chromosomes
(16). Several studies have revealed
that higher expression levels of NCAPG serve a role in cell
proliferation, angiogenesis and tumor therapy resistance in cancer
(22,58,59).
Wang et al (22) revealed
that transient inhibition of NCAPG using specific siRNAs resulted
in a significant decrease in cell proliferation and migration in
vitro in hepatocellular carcinoma. Ai et al (59) reported that miR-181c induced
hepatocellular carcinoma suppression via the regulation of NCAPG
expression. However, at present, the specific mechanism by which
NCAPG exerts its effects in cancer remains unclear.
To the best of our knowledge, the role and mechanism
of NCAPG in the development and progression of breast cancer has
not been extensively studied. NCAPG was upregulated in breast
cancer clinical samples compared with in matched adjacent normal
clinical samples. Additionally, NCAPG expression in breast cancer
was upregulated based on analysis of the UALCAN database. The
average NCAPG expression was negatively associated with PR and ER
status, and positively associated with HER2 status, NPI score, SBR
grade, basal-like status and triple-negative status in breast
cancer. Thus, it was hypothesized that upregulated NCAPG expression
was associated with the progression of breast cancer. Kaplan-Meier
Plotter analysis results revealed that high NCAPG expression was
associated with a less favorable progression in patients with
breast cancer. Additionally, the prognostic role of NCAPG analyzed
by PrognoScan suggested that low NCAPG expression was associated
with a favorable prognosis in patients with breast cancer.
Subsequently, the potential mechanisms by which
NCAPG exerted its effects in breast cancer were studied. miRNAs can
regulate genes post-transcriptionally (60). Among the miRNAs predicted to
potentially regulate NCAPG, 4 miRNAs (miR-101-3p, miR-195-5p,
miR-214-3p and miR-944) were previously reported to function as
tumor suppressor genes in breast cancer (Table IV) (4,61–65),
combined with the prognosis analysis, text mining, expression
analysis and correlation analysis.
| Table IV.Text mining the roles of potential
miRNAs in breast cancer. |
Table IV.
Text mining the roles of potential
miRNAs in breast cancer.
| First author,
year | PubMed ID | miRNAs | Direct targets | Function | Overall effect | (Refs.) |
|---|
| Zhang et al,
2018 | 30583076 | miR-101-3p | Med19 | Inhibit tumor
growth and metastasis | Tumor
suppressor | (61) |
| Wang et al,
2020 | 32226515 | miR-101-3p | E2F8 | Inhibit tumor
growth and metastasis | Tumor
suppressor | (62) |
| Yang et al,
2019 | 30621700 | miR-195-5p | CCNE1 | Inhibit
proliferation, migration, invasion | Tumor
suppressor | (63) |
| Luo et al,
2014 | 24402230 | miR-195-5p | CCNE1 | Inhibit cell
proliferation, reduced cell colony formation, suppressed cell
migration | Tumor
suppressor | (4) |
| Han et al,
2019 | 31539134 | miR-214-3p | Survivin | Inhibit
proliferation | Tumor
suppressor | (64) |
| Flores-pérez et
al, 2016 | 27377268 | miR-944 | SIAH1, PTP4A1 | Inhibit migration
and invasion | Tumor
suppressor | (65) |
A total of 261 co-expressed genes of NCAPG were
identified by co-expression analysis, and pathway enrichment
analysis indicated that these co-expressed genes were significantly
enriched in the p53 signaling pathway. Numerous studies have
demonstrated that the activation of the p53 signaling pathway is
associated with the development of various types of cancer,
including breast cancer (66–68).
Furthermore, the association between NCAPG and p53 has been
previously reported. Liu et al (69) revealed that silencing NCAPG in
SMMC-7721 and BEL-7404 cells resulted in decreased cell
proliferation and increased apoptosis, and was associated with
increased mRNA expression levels of p53, p27 and Bad. Additionally,
6 crucial proteins (NDC80, ESR1, ZWINT, NCAPG, ENO3 and CENPF) in
HCC were primarily enriched in cell cycle regulation and p53
signaling pathway (70). A total of
12 genes (CCNA2, CCNB1, CCNB2, CCNE2, CDC25C, CDK1, CDK2, CHEK1,
CHEK2, GTSE1, RRM2 and SKP2) were enriched in the p53 signaling
pathway. TCGA breast cancer data, UALCAN and Kaplan-Meier Plotter
were used to further determine the roles of these 12 genes,
including their expression and prognostic value in breast cancer.
Among these genes, CDC25C expression was upregulated in breast
cancer, and high CDC25C expression resulted in a significantly
worse prognosis compared with low CDC25C expression. CDC25C is a
cell cycle regulatory protein that serves a role in cell cycle
progression (4,63). In the present study, CDC25C
expression was significantly decreased following knockdown of
NCAPG. Additionally, downregulation of NCAPG resulted in
G2/M arrest in MCF-7 breast cancer cells. It was
preliminarily confirmed that NCAPG inhibited cell cycle progression
in breast cancer by regulating CDC25C. Additionally, the present
results indicated that NCAPG was downregulated by four targeted
miRNAs, possibly by elevating CDC25C expression to promote the
development of breast cancer.
Although the present study revealed that NCAPG
expression was associated with the clinical prognosis of breast
cancer, due to the limitations of online databases, only a single
factor analysis was performed as opposed to a multifactor
comparison of NCAPG expression. The grouping mode of the
bc-GenExMiner database requires further optimization. Additionally,
the association between NCAPG, CDC25C, the four identified miRNAs
(mir-101-3p, mir-195-5p, mir-214-3p and mir-944) and the p53
signaling pathway in breast cancer samples and the potential
therapeutic application of NCAPG needs to be further clarified.
In conclusion, the present study revealed that NCAPG
expression was upregulated in breast cancer, and this upregulated
expression was associated with a less favorable prognosis. In
addition, NCAPG expression was targeted by four miRNAs and was
associated with the dysregulation of the p53 signaling pathway, via
increased expression levels of CDC25C. The current findings
suggested that NCAPG may serve a key role in the development of
breast cancer and may become a new therapeutic target for breast
cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant on. 81802676) and the
Wuhan Youth Cadre Project (grant nos. 2017zqnlxr01 and
2017zqnlxr02).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request. The datasets (GSE19615, GSE12276, GSE6532-GPL570, GSE9195,
GSE12093, GSE11121, GSE1378, GSE1379, GSE2034, GSE1456-GPL96,
GSE7378, E-TABM-158, GSE3494-GPL96, GSE4922-GPL96, GSE2990 and
GSE7390) generated and/or analyzed during the current study are
available in The Cancer Genome Atlas (https://www.cancer.gov/about-nci/organization/ccg/research/structural-genomics/tcga).
Authors' contributions
MD conceived the study. MD and TX designed the
study. XC and HL performed the experiments. MD analyzed the data
and wrote the manuscript. XL and WX made substantial contributions
to the conception of the study, secured funding and supervised the
study. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All procedures involving human participants in the
present study were performed according to the ethical standards of
the Ethics Committee of Tongji Medical College of Huazhong Science
and Technology University (Wuhan, China; approval no. S340), and
written informed consent was provided by each participant.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics for hispanics/latinos, 2012. CA Cancer J Clin.
62:283–298. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kong Q and Qiu M: Long noncoding RNA
SNHG15 promotes human breast cancer proliferation, migration and
invasion by sponging miR-211-3p. Biochem Biophys Res Commun.
495:1594–1600. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Luo Q, Wei C, Li X, Li J, Chen L, Huang Y,
Song H, Li D and Fang L: MicroRNA-195-5p is a potential diagnostic
and therapeutic target for breast cancer. Oncol Rep. 31:1096–1102.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Greaney ML, Sprunck-Harrild K, Ruddy KJ,
Ligibel J, Barry WT, Baker E, Meyer M, Emmons KM and Partridge AH:
Study protocol for young & strong: A cluster randomized design
to increase attention to unique issues faced by young women with
newly diagnosed breast cancer. BMC Public Health. 15:372015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bartel DP: MicroRNAs: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Friedman RC, Farh KK, Burge CB and Bartel
DP: Most mammalian mRNAs are conserved targets of microRNAs. Genome
Res. 19:92–105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nelson KM and Weiss GJ: MicroRNAs and
cancer: Past, present, and potential future. Mol Cancer Ther.
7:3655–3660. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wiemer EA: The role of microRNAs in
cancer: No small matter. Eur J Cancer. 43:1529–1544. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Iorio MV and Croce CM: MicroRNAs in
cancer: small molecules with a huge impact. J Clin Oncol.
27:5848–5856. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Esquela-Kerscher A and Slack FJ:
Oncomirs-microRNAs with a role in cancer. Nat Rev Cancer.
6:259–269. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ke M, Zhang Z, Cong L, Zhao S, Li Y, Wang
X, Lv Y, Zhu Y and Dong J: MicroRNA-148b-colony-stimulating
factor-1 signaling-induced tumor-associated macrophage infiltration
promotes hepatocellular carcinoma metastasis. Biomed Pharmacother.
120:1095232019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Han L, Cui D, Li B, Xu WW, Lam AKY, Chan
KT, Zhu Y, Lee NPY, Law SYK, Guan XY, et al: MicroRNA-338-5p
reverses chemoresistance and inhibits invasion of esophageal
squamous cell carcinoma cells by targeting Id-1. Cancer Sci.
110:3677–3688. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shang S, Wang J, Chen S, Tian R, Zeng H,
Wang L, Xia M, Zhu H and Zuo C: Exosomal miRNA-1231 derived from
bone marrow mesenchymal stem cells inhibits the activity of
pancreatic cancer. Cancer Med. 8:7728–7740. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Eberlein A, Takasuga A, Setoguchi K, Pfuhl
R, Flisikowski K, Fries R, Klopp N, Fürbass R, Weikard R and Kühn
C: Dissection of genetic factors modulating fetal growth in cattle
indicates a substantial role of the non-SMC condensin I complex,
subunit G (NCAPG) gene. Genetics. 183:951–964. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu M and Thomas PD: GO functional
similarity clustering depends on similarity measure, clustering
method, and annotation completeness. BMC Bioinformatics.
20:1552019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li S, Xuan Y, Gao B, Sun X, Miao S, Lu T,
Wang Y and Jiao W: Identification of an eight-gene prognostic
signature for lung adenocarcinoma. Cancer Manag Res. 10:3383–3392.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Arai T, Okato A, Yamada Y, Sugawara S,
Kurozumi A, Kojima S, Yamazaki K, Naya Y, Ichikawa T and Seki N:
Regulation of NCAPG by miR-99a-3p (passenger strand) inhibits
cancer cell aggressiveness and is involved in CRPC. Cancer Med.
7:1988–2002. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liang ML, Hsieh TH, Ng KH, Tsai YN, Tsai
CF, Chao ME, Liu DJ, Chu SS, Chen W, Liu YR, et al: Downregulation
of miR-137 and miR-6500-3p promotes cell proliferation in pediatric
high-grade gliomas. Oncotarget. 7:19723–19737. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goto Y, Kurozumi A, Arai T, Nohata N,
Kojima S, Okato A, Kato M, Yamazaki K, Ishida Y, Naya Y, et al:
Impact of novel miR-145-3p regulatory networks on survival in
patients with castration-resistant prostate cancer. Br J Cancer.
117:409–420. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Gao B, Tan PY, Handoko YA, Sekar
K, Deivasigamani A, Seshachalam VP, OuYang HY, Shi M, Xie C, et al:
Genome-wide CRISPR knockout screens identify NCAPG as an essential
oncogene for hepatocellular carcinoma tumor growth. FASEB J.
33:8759–8770. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang L, Ren L, Chen H, Pan J, Zhang Z,
Kuang X, Chen X, Bao W, Lin C, Zhou Z, et al: NCAPG confers
trastuzumab resistance via activating SRC/STAT3 signaling pathway
in HER2-positive breast cancer. Cell Death Dis. 11:5472020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pontén F, Schwenk JM, Asplund A and
Edqvist PH: The human protein atlas as a proteomic resource for
biomarker discovery. J Intern Med. 270:428–446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jézéquel P, Campone M, Gouraud W,
Guérin-Charbonnel C, Leux C, Ricolleau G and Campion L:
bc-GenExMiner: An easy-to-use online platform for gene prognostic
analyses in breast cancer. Breast Cancer Res Treat. 131:765–775.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jézéquel P, Frénel JS, Campion L,
Guérin-Charbonnel C, Gouraud W, Ricolleau G and Campone M:
bc-GenExMiner 3.0: New mining module computes breast cancer gene
expression correlation analyses. Database (Oxford).
2013:bas0602013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Győrffy B, Surowiak P, Budczies J and
Lánczky A: Online survival analysis software to assess the
prognostic value of biomarkers using transcriptomic data in
non-small-cell lung cancer. PLoS One. 8:e822412013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mizuno H, Kitada K, Nakai K and Sarai A:
PrognoScan: A new database for meta-analysis of the prognostic
value of genes. BMC Med Genomics. 2:182009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li Y, Zou L, Li Q, Haibe-Kains B, Tian R,
Li Y, Desmedt C, Sotiriou C, Szallasi Z, Iglehart JD, et al:
Amplification of LAPTM4B and YWHAZ contributes to chemotherapy
resistance and recurrence of breast cancer. Nat Med. 16:214–218.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bos PD, Zhang XH, Nadal C, Shu W, Gomis
RR, Nguyen DX, Minn AJ, van de Vijver MJ, Gerald WL, Foekens JA and
Massagué J: Genes that mediate breast cancer metastasis to the
brain. Nature. 459:1005–1009. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Loi S, Haibe-Kains B, Desmedt C, Lallemand
F, Tutt AM, Gillet C, Ellis P, Harris A, Bergh J, Foekens JA, et
al: Definition of clinically distinct molecular subtypes in
estrogen receptor-positive breast carcinomas through genomic grade.
J Clin Oncol. 25:1239–1246. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sotiriou C, Wirapati P, Loi S, Harris A,
Fox S, Smeds J, Nordgren H, Farmer P, Praz V, Haibe-Kains B, et al:
Gene expression profiling in breast cancer: Understanding the
molecular basis of histologic grade to improve prognosis. J Natl
Cancer Inst. 98:262–272. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang Y, Sieuwerts AM, McGreevy M, Casey
G, Cufer T, Paradiso A, Harbeck N, Span PN, Hicks DG, Crowe J, et
al: The 76-gene signature defines high-risk patients that benefit
from adjuvant tamoxifen therapy. Breast Cancer Res Treat.
116:303–309. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schmidt M, Böhm D, von Törne C, Steiner E,
Puhl A, Pilch H, Lehr HA, Hengstler JG, Kölbl H and Gehrmann M: The
humoral immune system has a key prognostic impact in node-negative
breast cancer. Cancer Res. 68:5405–5413. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Loi S, Haibe-Kains B, Desmedt C, Wirapati
P, Lallemand F, Tutt AM, Gillet C, Ellis P, Ryder K, Reid JF, et
al: Predicting prognosis using molecular profiling in estrogen
receptor-positive breast cancer treated with tamoxifen. BMC
Genomics. 9:2392008. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ma XJ, Wang Z, Ryan PD, Isakoff SJ,
Barmettler A, Fuller A, Muir B, Mohapatra G, Salunga R, Tuggle JT,
et al: A two-gene expression ratio predicts clinical outcome in
breast cancer patients treated with tamoxifen. Cancer Cell.
5:607–616. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang Y, Klijn JG, Zhang Y, Sieuwerts AM,
Look MP, Yang F, Talantov D, Timmermans M, Meijer-van Gelder ME, Yu
J, et al: Gene-expression profiles to predict distant metastasis of
lymph-node-negative primary breast cancer. Lancet. 365:671–679.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hall P, Ploner A, Bjöhle J, Huang F, Lin
CY, Liu ET, Miller LD, Nordgren H, Pawitan Y, Shaw P, et al:
Hormone-replacement therapy influences gene expression profiles and
is associated with breast-cancer prognosis: A cohort study. BMC
Med. 4:162006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhou Y, Yau C, Gray JW, Chew K, Dairkee
SH, Moore DH, Eppenberger U, Eppenberger-Castori S and Benz CC:
Enhanced NF kappa B and AP-1 transcriptional activity associated
with antiestrogen resistant breast cancer. BMC Cancer. 7:592007.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chin K, DeVries S, Fridlyand J, Spellman
PT, Roydasgupta R, Kuo WL, Lapuk A, Neve RM, Qian Z, Ryder T, et
al: Genomic and transcriptional aberrations linked to breast cancer
pathophysiologies. Cancer Cell. 10:529–541. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Miller LD, Smeds J, George J, Vega VB,
Vergara L, Ploner A, Pawitan Y, Hall P, Klaar S, Liu ET and Bergh
J: An expression signature for p53 status in human breast cancer
predicts mutation status, transcriptional effects, and patient
survival. Proc Natl Acad Sci USA. 102:13550–13555. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ivshina AV, George J, Senko O, Mow B,
Putti TC, Smeds J, Lindahl T, Pawitan Y, Hall P, Nordgren H, et al:
Genetic reclassification of histologic grade delineates new
clinical subtypes of breast cancer. Cancer Res. 66:10292–10301.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Desmedt C, Piette F, Loi S, Wang Y,
Lallemand F, Haibe-Kains B, Viale G, Delorenzi M, Zhang Y,
d'Assignies MS, et al: Strong time dependence of the 76-gene
prognostic signature for node-negative breast cancer patients in
the TRANSBIG multicenter independent validation series. Clin Cancer
Res. 13:3207–3214. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42((Database Issue)): D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Wong NW, Chen Y, Chen S and Wang X:
OncomiR: An online resource for exploring pan-cancer microRNA
dysregulation. Bioinformatics. 34:713–715. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45((W1)):
W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Fernandez-Martinez A, Krop IE, Hillman DW,
Polley MY, Parker JS, Huebner L, Hoadley KA, Shepherd J, Tolaney S,
Henry NL, et al: Survival, pathologic response, and genomics in
CALGB 40601 (alliance), a neoadjuvant phase III trial of
paclitaxel-trastuzumab with or without lapatinib in HER2-positive
breast cancer. J Clin Oncol. 38:4184–4193. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Elsharawy KA, Mohammed OJ, Aleskandarany
MA, Hyder A, El-Gammal HL, Abou-Dobara MI, Green AR, Dalton LW and
Rakha EA: The nucleolar-related protein Dyskerin pseudouridine
synthase 1 (DKC1) predicts poor prognosis in breast cancer. Br J
Cancer. 123:1543–1552. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kuba MG, Lester SC, Bowman T, Stokes SM,
Taneja KL, Garber JE and Dillon DA: Histopathologic features of
breast cancer in Li-Fraumeni syndrome. Mod Pathol. Jul
7–2020.(Online ahead of print).
|
|
53
|
Lou W, Chen J, Ding B, Chen D, Zheng H,
Jiang D, Xu L, Bao C, Cao G and Fan W: Identification of
invasion-metastasis-associated microRNAs in hepatocellular
carcinoma based on bioinformatic analysis and experimental
validation. J Transl Med. 16:2662018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Lou W, Liu J, Ding B, Xu L and Fan W:
Identification of chemoresistance-associated miRNAs in breast
cancer. Cancer Manag Res. 10:4747–4757. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Lou W, Liu J, Gao Y, Zhong G, Ding B, Xu L
and Fan W: MicroRNA regulation of liver cancer stem cells. Am J
Cancer Res. 8:1126–1141. 2018.PubMed/NCBI
|
|
56
|
Liao WL, Lin JY, Shieh JC, Yeh HF, Hsieh
YH, Cheng YC, Lee HJ, Shen CY and Cheng CW: Induction of G2/M phase
arrest by diosgenin via activation of Chk1 kinase and Cdc25C
regulatory pathways to promote apoptosis in human breast cancer
cells. Int J Mol Sci. 21:1722019. View Article : Google Scholar
|
|
57
|
Pan Z, Zhang X, Yu P, Chen X, Lu P, Li M,
Liu X, Li Z, Wei F, Wang K, et al: Cinobufagin induces cell cycle
arrest at the G2/M phase and promotes apoptosis in malignant
melanoma cells. Front Oncol. 9:8532019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang Q, Su R, Shan C, Gao C and Wu P:
Non-SMC condensin I complex, subunit G (NCAPG) is a novel mitotic
gene required for hepatocellular cancer cell proliferation and
migration. Oncol Res. 26:269–276. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ai J, Gong C, Wu J, Gao J, Liu W, Liao W
and Wu L: MicroRNA-181c suppresses growth and metastasis of
hepatocellular carcinoma by modulating NCAPG. Cancer Manag Res.
11:3455–3467. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bertoli G, Cava C and Castiglioni I: The
potential of miRNAs for diagnosis, treatment and monitoring of
breast cancer. Scand J Clin Lab Invest Suppl. 245:S34–S39. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang X, Gao D, Fang K, Guo Z and Li L:
Med19 is targeted by miR-101-3p/miR-422a and promotes breast cancer
progression by regulating the EGFR/MEK/ERK signaling pathway.
Cancer Lett. 444:105–115. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Wang H, Wang L, Tang L, Luo J, Ji H, Zhang
W, Zhou J, Li Q and Miao L: Long noncoding RNA SNHG6 promotes
proliferation and angiogenesis of cholangiocarcinoma cells through
sponging miR-101-3p and activation of E2F8. J Cancer. 11:3002–3012.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Yang R, Xing L, Zheng X, Sun Y, Wang X and
Chen J: The circRNA circAGFG1 acts as a sponge of miR-195-5p to
promote triple-negative breast cancer progression through
regulating CCNE1 expression. Mol Cancer. 18:42019. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Han LC, Wang H, Niu FL, Yan JY and Cai HF:
Effect miR-214-3p on proliferation and apoptosis of breast cancer
cells by targeting survivin protein. Eur Rev Med Pharmacol Sci.
23:7469–7474. 2019.PubMed/NCBI
|
|
65
|
Flores-Pérez A, Marchat LA,
Rodríguez-Cuevas S, Bautista VP, Fuentes-Mera L, Romero-Zamora D,
Maciel-Dominguez A, de la Cruz OH, Fonseca-Sánchez M, Ruíz-García
E, et al: Suppression of cell migration is promoted by miR-944
through targeting of SIAH1 and PTP4A1 in breast cancer cells. BMC
Cancer. 16:3792016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Gao J, Xia R, Chen J, Gao J, Luo X, Ke C,
Ren C, Li J and Mi Y: Inhibition of esophageal-carcinoma cell
proliferation by genistein via suppression of JAK1/2-STAT3 and
AKT/MDM2/p53 signaling pathways. Aging (Albany NY). 12:6240–6259.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Jiang W, Hou L, Wei J, Du Y, Zhao Y, Deng
X and Lin X: Hsa-miR-217 inhibits the proliferation, migration, and
invasion in non-small cell lung cancer cells via targeting SIRT1
and P53/KAI1 signaling. Balkan Med J. 37:208–214. 2020.PubMed/NCBI
|
|
68
|
Zheng X, Zhang J, Fang T, Wang X, Wang S,
Ma Z, Xu Y, Han C, Sun M, Xu L, et al: The long non-coding RNA
PIK3CD-AS2 promotes lung adenocarcinoma progression via
YBX1-mediated suppression of p53 pathway. Oncogenesis. 9:342020.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Liu K, Li Y, Yu B, Wang F, Mi T and Zhao
Y: Silencing non-SMC chromosome-associated polypeptide G inhibits
proliferation and induces apoptosis in hepatocellular carcinoma
cells. Can J Physiol Pharmacol. 96:1246–1254. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Liu ZK, Zhang RY, Yong YL, Zhang ZY, Li C,
Chen ZN and Bian H: Identification of crucial genes based on
expression profiles of hepatocellular carcinomas by bioinformatics
analysis. PeerJ. 7:e74362019. View Article : Google Scholar : PubMed/NCBI
|