Ovarian cancer is one of the three primary
gynecological tumors, with a 5-year survival rate of 44%. Due to
its lack of specific clinical symptoms and practical measures for
early diagnosis, >75% patients are diagnosed in advanced stages
and quickly become prone to drug resistance during treatment
(1). Ovarian cancer cells can
simultaneously regulate immune activation and suppression by
presenting cancer antigens to immune cells, secreting cytokines and
a large number of soluble factors, as well as releasing exosomes to
the tumor microenvironment affecting the proximal and distal
tissues. These influencing factors together form a complex
interactome called the tumor immune microenvironment (2). Innate and adaptive immune cells can
stimulate an antitumor response by recognizing cancer antigens via
the antigen presenting cells (APCs) (3). In addition, immune cells, such as
lymphocytes, macrophages, dendritic cells (DCs), mast cells and
natural killer (NK) cells, can regulate angiogenesis, certain
tumorigenic metabolic pathways and metastasis within the tumor
microenvironment (4,5). Exosomes have also been shown to play an
important role in affecting the tumor immune microenvironment and
ensuing immune responses, such as antigen presentation, migration,
metastasis and tumor invasion. Previous research indicates that
exosomes carry a number of immunologically active molecules
[including major histocompatibility complex (MHC I), heat shock
protein (HSP) and CD81] that can stimulate an antitumor immune
response (6). Conversely, some
studies have shown that exosomes will weaken the antitumor immune
response to effect cancer progression by potentiating immune
evasion (7–9). Analyzing the immune microenvironment of
ovarian cancer and understanding the role of exosomes in cancer
progression could play a vital role in its early diagnosis and
designing an effective immunotherapy regimen.
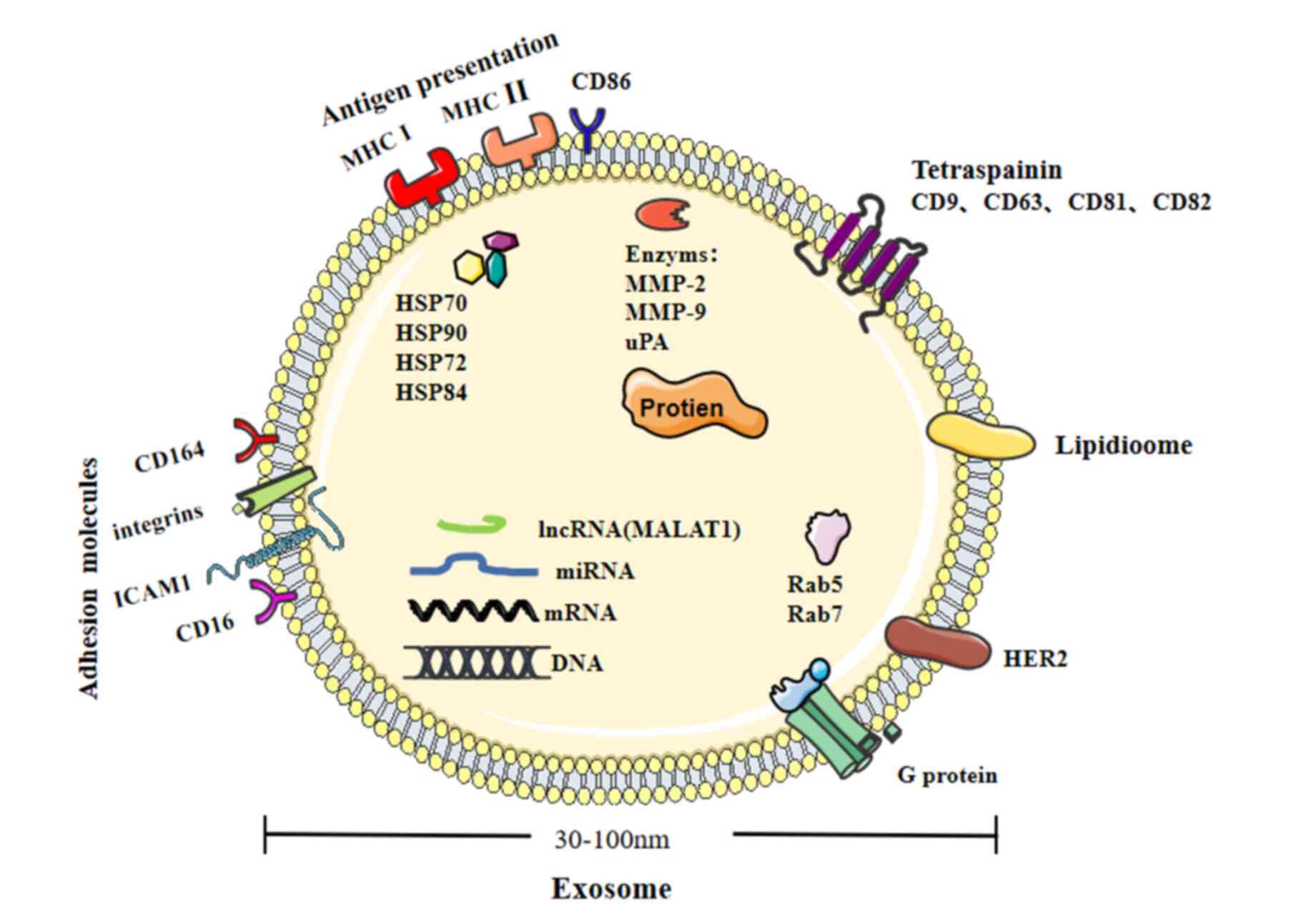
As endogenous cellular components, exosomes are
vesicles (30–100 nm) derived from the endosomal compartments called
multivesicular bodies (Fig. 1) that
are secreted by various cells into the extracellular
microenvironment. When exosomes are isolated by density gradient
centrifugation or ultracentrifugation, they appear as round
vesicles in solution. The ultrastructure is resolved by
dehydration, where it appears cup-shaped under an electron
microscope (10). Exosomes are
double-membraned organelles formed by periodic endocytosis of
intracellular fluid throughout the life cycle of eukaryotic cells.
As early endosomes mature and develop into late endosomes, the
inner membrane sprouts inward to form intraluminal vesicles (ILVs),
which contain randomly engulfed parts of the cytoplasmic content,
rich with mRNAs, microRNAs (miRNAs/miRs), proteins and lipids. ILVs
that are released to the extracellular environment are called
exosomes (11). Studies show that
exosomes can be found in various extracellular fluids, such as
blood, urine, ascites, semen and cerebrospinal fluid.
Exosomes can be used as delivery vehicles for a
variety of bioactive molecules, for example proteins, lipids, mRNA,
miRNA, long non-coding RNA (lncRNA), genomic DNA and cDNA. This
unique composition is also occasionally used as an identifier for a
particular exosome (12). Previous
research indicates that the common proteins in exosomes include
tetraspanins, co-stimulatory molecule CD86 and adhesion molecules,
such as integrins, ICAM1, CD166 and CD146. Besides specialized
proteins, exosomes may also carry common proteins, such as HSP-70,
HSP84 and HSP90 (11). Exosomes can
accelerate peptide loading onto MHC I and II, thereby mediating a
rapid immune response. Exosomes also carry signal transduction
proteins, for example, receptor tyrosine kinases and membrane
transport and fusion proteins, such as the GTPases Rab5 and Rab7
(11). Studies have also reported
that nucleic acids are carried by exosomes, comprising of a diverse
mix of DNA (13,14), RNA (15), lncRNA (16) and miRNA (17,18)
molecules. Exosomal miRs (miR-15, −16, 151 and −375) can promote
angiogenesis and tumor progression in the TME (11). The bioactive cargo carried by
exosomes can participate in the modification of immune response of
an ovarian cancer microenvironment (19).
In infectious and non-infectious pathological
conditions, both non-tumorous and tumorous cells tend to release
exosomes more actively, whereas the number of exosomes quantified
from the blood of patients with ovarian cancer is 3–4 times higher
compared with in healthy individuals (11). The exosomes extracted from two human
ovarian carcinoma cell lines OVCAR-3 and IGROV1 have a density
ranging from 1.09–1.15 g/ml, while ~2,230 proteins are detected in
the exosomal cargo, including other significant exosomal protein
markers (20). Andre et al
(6) in 2002 detected human epidermal
growth factor (EGF) receptor (Her2/neu gene) signaling in
exosomes of patients with ovarian cancer via western blotting.
Activated matrix metalloproteinase (MMP)-2, MMP-9 and urokinase
plasminogen activator are found in exosomes derived from the
ascites of patients with ovarian cancer, which promotes protease
activation to increase degradation of the extracellular matrices
(ECMs) and tumor cell invasion and metastasis (21).
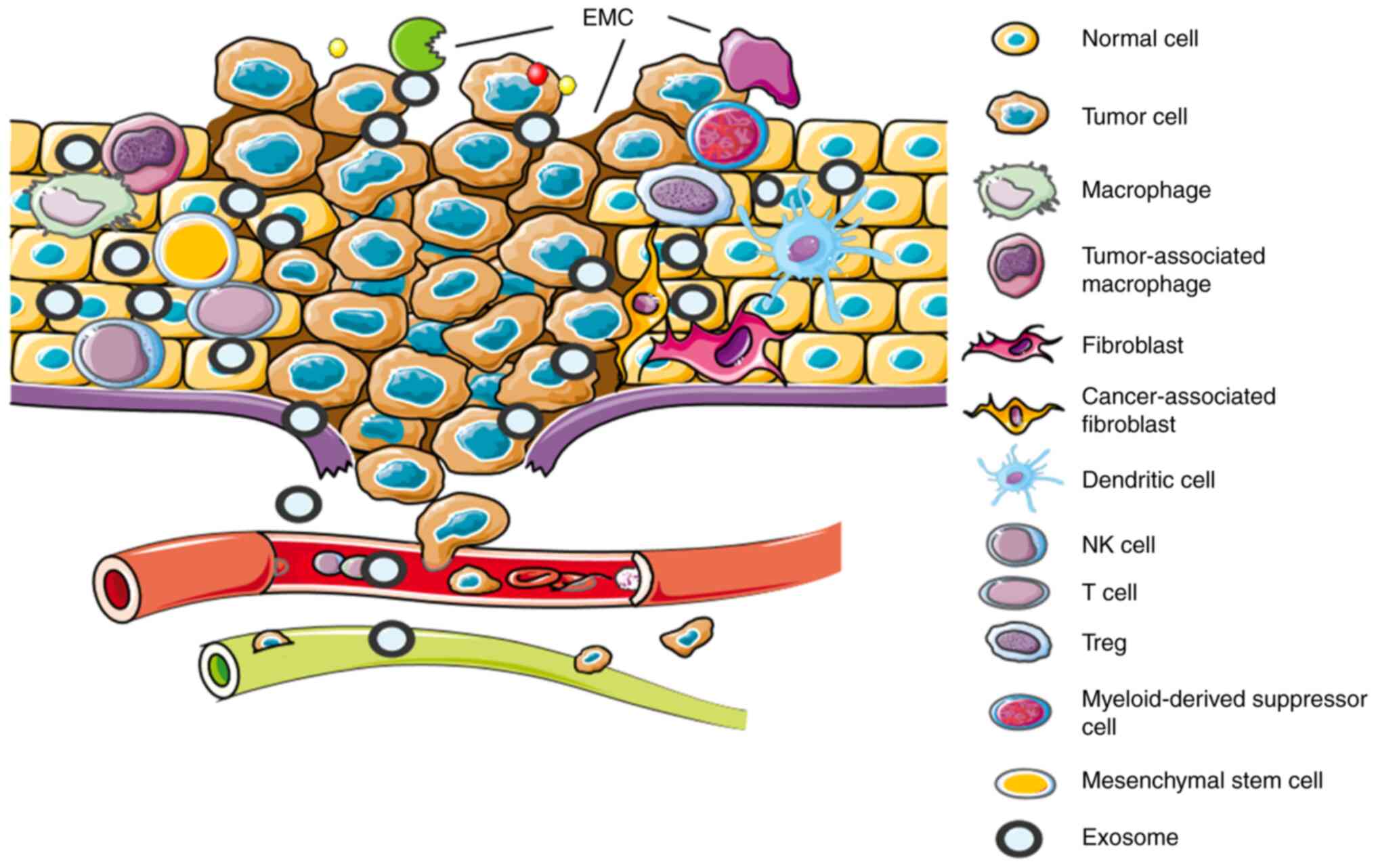
The tumor microenvironment is the product of a
number of cells and their accompanying extracellular matrix
component (EMCs) jointly contributing towards the development of a
distinct microenvironment surrounding the tumor mass. The cells
comprise of stromal cells, including: Fibroblasts, macrophages,
myeloid-derived suppressor cells, endothelial cells and mesenchymal
stem cells. EMCs comprise of inflammatory cytokines, chemokines,
MMPs, integrins and exosomes (22)
(Fig. 2). Tumor cells interact with
stromal cells to promote angiogenesis, infiltration and metastasis
that cause the tumor to grow and invade other tissues (23). TNF-α is a pro-inflammatory cytokine
that is secreted primarily by macrophages along with other cells of
the stroma, which promotes tumor necrosis or apoptosis. In total,
~28% of all cancer types are affected by TNF-mediated necrosis
(24). 5-Lipoxygenase (5-LOX)
is a member of the lipoxygenase family of genes that is a key
enzyme in the conversion of arachidonic acid to leukotrienes. In
ovarian cancer, upregulation of 5-LOX metabolites and TNF-α can
promote recruitment of macrophages to the tumor site (25). In addition, the pro-inflammatory
interleukin-6 (IL-6) has been shown to be an important cytokine in
the ovarian cancer tumor microenvironment. IL-6 can mediate the
maturation of macrophages into M2 macrophages, which enhance tumor
vascular stability by VEGF and TGF-β1, thus promoting tumor
progression (26–29). Upregulation of miR-217 in ovarian
cancer downregulates the IL-6-dependent JAK3/STAT3 signaling
pathway, thereby potentially inhibiting the maturation of
macrophages from M1 to M2 stage (30). Ovarian cancer-associated mesenchymal
stem cells (MSCs) highly express IL-6 and leukemia inhibitor factor
that activate the JAK3/STAT3 signaling pathway to increase the
tumorigenicity of ovarian cancer stem cells (31). Wang et al (32) have found that cancer-associated
fibroblasts (CAFs) can also secrete IL-6 and promote the
accumulation of ovarian cancer stem cells in residual tumors by
activating the STAT3 signaling pathway. Exosomes derived from
ascites in patients with ovarian cancer can promote the release of
more IL-6 from monocytes (THP-1 cells) and activate the NF-kB and
STAT3 signaling pathways, which leads to a cytokine environment
conducive for immune evasion of tumor cells (33). In addition, IL-6 has been associated
with chemotherapy resistance and poor prognosis in patients with
ovarian cancer. Studies have shown that the level of IL-6 in the
serum of patients with cancer is significantly higher compared with
that of normal individuals (34,35).
IL-6 can upregulate the expression of resistance-related genes
multi-drug resistance-1 and glutathione S-transferase
π; in addition to the expression of apoptosis inhibitor protein.
Moreover, IL-6 can activate the Ras/MEK/ERK and PI3K/Akt signaling
pathways that jointly induce chemotherapy resistance (34,36). The
value of IL-6 as a prognostic and diagnostic indicator of ovarian
cancer has been confirmed (37,38).
Studies suggest that a higher ratio of M2:M1
macrophages is associated with poor prognosis in patients with
ovarian cancer, whereas a higher ratio of M1:M2 macrophages is
associated with good prognosis (39,40).
Some investigations have shown that tumor-associated macrophages
(TAMs) can activate the MMP9/HB-EGF pathway along with the
production of EGF to promote ovarian cancer and breast cancer
progression (41,42). TGF-β can promote the transformation
of epithelial cells to mesenchymal cells, promoting angiogenesis
and inducing immunosuppression, subsequently promoting tumor
progression (43). TAMs release
TGF-β1 and tenascin-C to promote tumor metastasis in ovarian cancer
(44). CAFs can also promote
invasion and metastasis of ovarian cancer in the tumor
microenvironment (45,46). Studies have shown that TGF-β1
secreted by CAFs can notably potentiate the mechanism of
epithelial-mesenchymal transition (EMT), thereby promoting bladder
cancer to metastasize (47).
Similarly, CAFs highly express the TGF-β gene in ovarian
cancer (48). In addition, studies
have shown that CAFs in ascites can promote the production of
multicellular aggregates, thereby promoting peritoneal metastasis
(46,49). CAFs highly express X-linked sushi
repeat-containing protein, which are peroxiredoxin enzymes that
control cytokine-induced peroxide levels. Similarly, CAFs also
highly express hemicentin-1 genes in ovarian cancer tissue
samples. Sequential knock-down of these two genes can weaken the
ability of CAFs to promote ovarian cancer metastasis (50). Myeloid-derived suppressor cells
(MDSCs) and Tregs are important components of the tumor immune
evasion mechanism (51,52). A previous study confirmed that the
co-culture of MDSCs and ovarian cancer cells can promote the
formation of tumor spheres, cell colonies and the accumulation of
cancer stem cells, thus a strong indication that MDSCs can induce
tumor progression (53).
VEGF-induced MDSCs inhibit the activity of CD8+ T cells
in ovarian cancer, weakening the host's antitumor immune response
and leading to a poor prognosis (54). A study showed that a higher count of
Treg cells could be detected in the peripheral blood of patients
with ovarian cancer and thus is associated with poor prognosis
(55).
In addition to stromal cells, non-cellular
components are also included in the tumor microenvironment. TGF-β
is a major cytokine in the tumor microenvironment. TGF-β combines
with SMADs, the main signal transducers for TGF-β receptors, to
activate cells, which can promote transformation of fibroblasts and
regulate cell proliferation and apoptosis (56). The high expression of TGF-β3 is
associated with poor prognosis of high-grade serous carcinoma, and
it is a potential indicator for the evaluation of ovarian cancer
prognosis (57). Recent studies have
reported that in ovarian cancer stem cells, inhibition of
TGF-β/SMAD pathway activation can further inhibit EMT (58–60). In
2012, Kulbe et al (61) first
described the ‘TNF’ network in the ovarian cancer microenvironment.
‘TNF’ network means that TNF, CXCL12 and IL-6 have a paracrine
effect in the tumor microenvironment, affecting angiogenesis and
immune cell infiltration. In addition, cytokine-induced tumor cells
can release guanylate binding-protein-1 and have an antitumor
effect (62). The role of cytokines
in the tumor microenvironment provides new possibilities for the
treatment of ovarian cancer.
As an essential constituent of the tumor
microenvironment, the role of exosomes in the tumor
microenvironment can be summarized into two aspects, namely
tumor-promoting and tumor-inhibiting. Exosomes modulate immune
regulation to reshape the tumor microenvironment through
metabolism-regulation, stimulation signal upregulation and
inhibition signal evasion (63,64).
Exosomes induce angiogenesis by changing the biological
characteristics of endothelial cells and regulating pro-angiogenic
factors (65). In addition, exosomes
may induce human hepatocellular carcinoma metastasis and invasion
through EMT, ECM degradation and vascular leakage (66). Ras-like in rat brain (Rab) protein is
a member of the GTPase family and plays an important role in
regulating the budding, movement and fusion of microvesicles.
Studies have shown that ovarian cancer cells can increase the
release of exosomes by upregulating Rab27a, downregulating Rab7,
lysosome-associated membrane protein-1, neuraminidase-1 mRNA and
therefore promoting the secreted lysosomal phenotype (67,68). In
addition, the hypoxia-induced exosomes carry oncogenic proteins
STAT3 and FAS, which can significantly increase ovarian cancer cell
migration, invasion and chemotherapy resistance (67). Epithelial ovarian cancer cells
transfer metastasis-associated lung adenocarcinoma transcript-1 a
lncRNA to human umbilical vein endothelial cells through exosomes,
activating the expression of genes related to angiogenesis
(69). Tang et al (70) demonstrated that ascites-derived
exosomes highly express soluble E-cadherin, which can promote
angiogenesis and ovarian cancer progression. The immune response
regulated by exosomes also plays a role in suppressing tumor
progression. Exosomes derived from ascites in patients with ovarian
cancer can detect T cell receptors, CD20 and human leukocyte
antigen-DR isotype (HLA-DR), in addition to histones H2A, B7-2 and
HER2/neu gene, in order to participate in immune modulation
(21). In addition, exosomes
extracted from ascites of patients with ovarian cancer can induce
apoptosis, inhibit proliferation, invasion and metastasis of tumor
cells as they exert an antitumorigenic effect (21,71).
As aforementioned, the tumor immune microenvironment
is a product of immune cells and immune molecules that inhibit the
proliferation of tumor cells (3).
There are also a variety of components that can promote the
proliferation and invasion of tumor cells, including
immunosuppressive cells such as Tregs, TAMs, CAFs, MDSCs and some
immunosuppressive signaling factors (72).
In the tumor immune microenvironment, the host gives
an innate immune response against the tumor mass while an adaptive
immune response is given against the tumor antigens, thus
preventing tumor progression. Innate immune cells include NK cells,
macrophages and DCs. Adaptive immune effectors include
CD8+, cytotoxic T lymphocyte (CTL) and CD4+
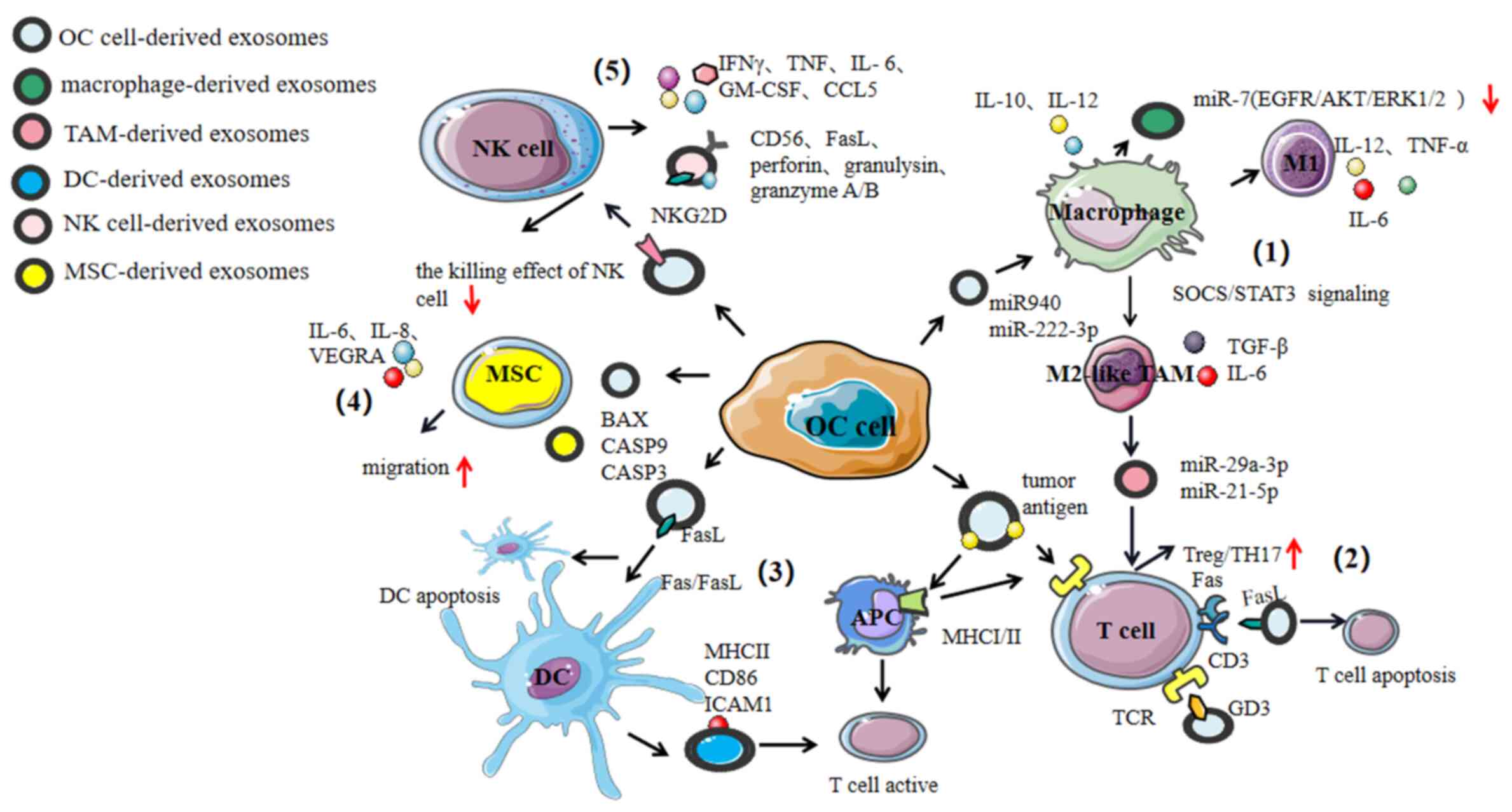
Th cells (72). Exosomes play a role
in mediating cross-talk with immune cells to exert an
antitumorigenic and/or a pro-tumorigenic effect (Fig. 3).
Macrophages are the first line of defense against
foreign pathogens and key effectors of innate immunity, they are
the key cells to bridge innate and adaptive immunities (73). According to their activation
pathways, macrophages can be divided into two types:
Classically-activated macrophages (named M1) and
alternatively-activated macrophages (named M2) (74). Macrophages are usually polarized into
M1 phenotype after being induced by IFN-γ, TNF-α, IL-6 and
lipopolysaccharides. Their surface highly expresses MHC II and
co-stimulatory proteins, such as CD80 and CD86 (75). M1 macrophages are generally
considered pro-inflammatory and release IL-6, IL-12, TNF-α and
reactive oxygen species (ROS), which are considered intermediates
that are associated with cytotoxicity and anti-tumorigenic
properties (76,77). M2 macrophages are polarized by IL-4,
IL-13, IL-10 and IL-33 (56). The
markers on the surface of M2 macrophages include found in
inflammatory zone 1, mannose receptor 1 and MHC II (74). M2 macrophages have the capacity to
secrete TGF-β, IL-6 and arginase-1 to facilitate
neovascularization, inhibit the adaptive immune response, ensure
tumor cell survival and remodel the ECM, which are all generally
considered as tumor-promoting functions (78,79). In
addition, TAMs are the main cells in the tumor immune
microenvironment that have two phenotypes: M1-Like TAMs and M2-like
TAMs (80). In the immune
microenvironment of ovarian cancer TAMs usually manifest as the
M2-like phenotype. The variety of biomarkers on the surface are
scavenging receptor B (CD163), mannose receptor (CD204), IL-10 and
chemotactic factor ligands CCL18 and CCL22 (81,82).
IL-10 secreted by TAMs can activate Treg cells and promote tumor
progression (83). miR-29a-3p and
−21-5p, which are abundant in TAM-derived exosomes, can be
transferred to CD4+T cells; thereby inhibiting STAT3
from regulating the ratio of Treg:Th17 and creating an
immunosuppressive microenvironment that is necessary for the
ovarian tumor to evade an active immune response, thus helping in
tumor progression (84).
Tumor-derived exosomes can induce macrophages to
differentiate into the M2 phenotype and TAMs, which have been
confirmed in extracts from various organs, including: Ovaries,
colorectal regions, endometrium, pancreas, melanoma, liver, breast
and lung cancer cells (85–91). In a hypoxic microenvironment, high
expression of miR-940 in exosomes has been derived from epithelial
ovarian cancer cells, which induces macrophages to differentiate
into the M2 phenotype, thus promoting proliferation and metastasis
of epithelial ovarian cancer (92).
Similar research shows that under hypoxic conditions, exosomes
derived from epithelial ovarian cancer activate hypoxia-inducible
factors that induce macrophages to highly express miR-21-3p,
−125b-5p and −181d-5p and promotes their polarization to the M2
phenotype through the cytokine signal transduction 4/5/STAT3
signaling pathway, which was also verified in vivo. The
JAK-STAT pathway mediates inflammatory immune response by
converting cytokine signals, and SOCS is the key regulator of the
pathway (76). Through microarray
analyses, some researchers found that the expression of miR-221-3p
was upregulated in exosomes that were derived from M2 macrophages.
Additionally, miR-221-3p can target the inhibition of
cyclin-dependent kinase inhibitor 1B, thus promoting the
proliferation of ovarian cancer cells via transition from
G1 to S (93). In
addition, epithelial ovarian cancer-derived exosomes overexpress
miR-222-3p and transfer it to macrophages to induce them into
M2-like polarization by the SOCS3/STAT3 pathway. miR-222-3p targets
downregulation of SOCS3 gene expression and activates STAT3
expression (94). This transfer of
miR-222-3p can facilitate the progression of ovarian cancer
(94). Wu et al (95) have shown that TAM-derived exosomes
inhibit endothelial cell migration by targeting the
miR-146b-5p/TRAF6/NF-kB/MMP2 pathway, whereas ovarian
cancer-derived exosomes can reverse the role of TAMs in endothelial
cells by transferring lncRNAs.
In order for macrophages to differentiate into TAMs,
macrophage-derived exosomes are important components involved in
the antitumor immune response (84,93,96). The
TNF-related weak inducer of apoptosis (TNSFS12 or TWEAK)-stimulated
macrophage-derived exosomes can be internalized by the tumor cells,
which can inhibit ovarian cancer metastasis. A study revealed that
TWEAK-stimulation increased the expression of miR-7 (a tumor
suppressor) in exosomes released by macrophages, which
downregulates the activity of the EGFR/AKT/ERK1/2 signaling pathway
and inhibits ovarian cancer metastasis (96). This was performed in mouse models
where TWEAK-stimulated macrophage-derived exosomes blocked the
metastasis of epithelial ovarian cancer (96). In addition, Baj-Krzyworzeka et
al (97) have shown that
tumor-derived exosomes can activate monocytes by increasing HLA-DR
expression, upregulating reactive oxygen intermediates and TNF and
by accumulating and secreting IL-10 and IL-12 mRNA. Exosomes
activate monocytes and induce them to differentiate into
macrophages (97–99). At present, there are few studies on
macrophage-derived exosomes in ovarian cancer, which is an area
that needs to be explored further. Exosomes derived from breast
cancer cells have upregulated levels of miR-130 and miR-33, which
can alter the polarization of macrophages from M2 to M1 phenotype
and inhibit tumor progression (100). The exosomes derived from TAMs of
progranulin (PGRN)-negative tumor tissues have upregulated
expression of miR-5100, which inhibits the invasion, migration and
EMT of breast cancer cells by targeting the CXCL12/CXCR4 axis
(101). Although a similar study in
ovarian cancer has not been performed, studies have shown that
expression of PGRN protein relates to poor ovarian cancer prognosis
(102,103). Moreover, high expression of PGRN
can induce EMT in ovarian cancer cells (104). This suggests that exosomes are a
potential therapeutic target for ovarian cancer.
In a tumor environment, NK cells are the first line
of defense within the immune system. NK cells mainly kill target
cells in four ways: i) Antibody-dependent cell-mediated
cytotoxicity via the Fas/FasL pathway (105), ii) the perforin-granzyme pathway
(106), iii) binding to target
cells through adhesion molecules (107) and iv) releasing cytokines to attack
target cells (108,109). NK cells are important effectors in
the cancer immune surveillance (110,111).
Upon activation, they secrete pro-inflammatory factors and
chemokines, for example IFN-γ, TNF, IL-6, GM-CSF and chemotactic
cytokine ligand 5 to mediate antitumor immune responses, affect
antitumor activity and promote formation of the tumor
microenvironment (112). IL-15
enhances the antitumor activity of NK cell-derived exosomes and has
been validated in mouse models (113). Exosomes derived from NK cells
express the killer protein (CD56), FasL, perforin, granulysin, and
granzyme A and B, which show antitumor activity and play a role in
immune surveillance (114). Killer
proteins expressed by NK cell-derived exosomes can participate in
NK cell-mediated cytotoxic killing effects (115), and the expressed DNAX accessory
molecule 1 (DNAM-1/CD226) receptor can bind to DNAM-1 ligands on
the cell membrane of tumor cells to exert a cytotoxic tumor cell
killing effect (116). This is an
important role of NK cells in cancer immune surveillance. NK
cell-derived exosomes express FasL and perforin and exert cytotoxic
effects in melanoma (117). NK
cell-derived exosomes carry miR-186, which can inhibit
neuroblastoma growth (118). The
antitumor effect of NK cell-derived exosomes on invasive melanoma
and neuroblastoma has been confirmed, exosomes derived from NK
cells can potentially be used in cancer treatment (117,118).
Although NK cells are a part of the innate immune
system and are capable of killing tumor cells, tumor
microenvironment also affects the cytotoxicity of NK cells
(119). Exosomes released by
ovarian cancer cells highly express KLRK1/ natural killer group 2
(NKG2D) ligands in the manner of MHC I chain-related protein A and
B and UL16-binding protein. This downregulates the expression of
NKG2D receptors on peripheral blood mononuclear cells, affecting
the activation of NK cells and suppressing their natural killing
effect (120).
Research shows that T cell-derived exosomes can
regulate gene expression and extracellular signal transduction of
DCs by carrying miRNA and T cell receptor protein (TCR)-rich
vesicles (125). MHC I, MHC II and
T cell co-stimulatory molecules on the surface of B
lymphocyte-derived exosomes can stimulate the proliferation of T
lymphocytes and thus prevent tumor progression (126). T follicular helper cell-derived
exosomes facilitate the proliferation of B lymphocytes (127). CD4+T cell-derived
exosomes regulate the gene expression of B lymphocytes by
transporting their miRNA cargo to B lymphocytes, which therefore
decreases the level of antibodies produced by B cells (128).
However, a previous study confirmed that tumor
cell-derived exosomes recruit lymphocytes to suppress the antitumor
immune response, promoting tumor progression by increasing tumor
invasion, angiogenesis and upregulating proinflammatory cytokines,
such as IL-6 and VEGFA (129).
Exosomes can induce T cell apoptosis and weaken the immune response
to achieve immune escape. FasL is a transmembrane protein in the
TNF family of proteins that is increased in malignant tumors. FasL
is overexpressed in tumor-derived exosomes (11,130).
FasL recognizes the Fas receptor on the surface of target cells and
triggers intrinsic apoptosis of target cells via the
FADD-procaspase-8 pathway, to induce T cell apoptosis and suppress
the immune response (11). In
addition, Fas can also mediate the apoptosis of lymphocytes through
antibody-dependent direct cytotoxicity. A previous study has showed
41 kDa FasL and HLA class I antigens are upregulated in ovarian
cancer-derived exosomes and inhibit T cell antitumor immune
function through FasL/CD3-ζ ligand-receptor binding (131). This inhibitory function has also
been observed in other tumors, such as exosomes derived from human
prostate cancer cells that induce CD8+T cell apoptosis
by FasL/Fas (132). FasL is also
found in exosomes from melanoma and colorectal cancer, which is
considered a new immune escape pathway (133). The expression of circ-0001068 (a
novel biomarker for ovarian cancer and inducer of PD1 expression in
T cells) in exosomes derived from ovarian cancer cells is
upregulated and can be delivered to T cells as well. Once in T
cells, circ-0001068 downregulates the expression level of miR-28-5p
and thus induces the expression of PD1 (134). Exosomes released by ovarian cancer
cells can carry plasma gelsolin and induce CD8+ T cell
apoptosis, which weakens immune surveillance (135).
Exosomes from the ascites of patients with ovarian
cancer can inhibit T cell signaling molecules, inhibit the
expression of CD3-ζ and JAK 3, and induce T cell apoptosis
(136). In the ascites of patients
with ovarian cancer, the ganglioside GD3 expressed on the surface
of exosomes binds with the TCR and actively arrest T cell function,
which greatly reduces the antitumorigenic effect of T cells
(137,138). Moreover, a previous study showed
that this inhibitory effect mediated by GD3 and TCR is related to
the sialic acid group in exosomes (137). When the sialic acid group is
hydrolyzed, this inhibitory effect disappears, which indicates that
GD3 can be a new target for ovarian cancer immunotherapy (137). In addition, the ascites-derived
exosomes of patients with ovarian cancer can effectively block T
cells in a reversible manner due to the early translocation of
NF-kB and later functional activation of IFN-γ. Researchers have
demonstrated that this can be reversed within 24–48 h by the
removal of exosomes. Therefore, targeted removal of exosomes will
increase the antitumorigenic effect of the host T cells (139).
Tumor-derived exosomes can also inhibit T cell
activation to cause immunosuppression. Functional CD39 and CD73
expressed by exosomes can dephosphorylate exogenous ATP and cAMP to
form adenosine, and inhibit T cell activation through the adenosine
A2A receptor. Therefore, exosomes increase the production of
extracellular adenosine to regulate the antitumor immune effect of
the T cells (140). In addition,
phosphatidylserine (PS)-positive exosomes derived from the ascites
of patients with ovarian cancer block the NF-kB and NFAT pathway
signaling cascade in T cells, and reversibly inhibit T cell
activation (141). Thus, depletion
of anti-PS antibodies or blocking PS can notably eliminate the
inhibition of T cells, which could be another new treatment method
for patients with ovarian cancer (142).
DCs are unique, in that they can activate T cells.
They can also activate immune responses or induce immune tolerance
(143). Mast cell-derived exosomes
contain HSP60 and HSP70, which can promote DC maturation and exert
antitumorigenic immune effects in a mouse model (144). The ovarian cancer microenvironment
is rich in cytokines and angiogenic factors, which can change the
phenotype and function of DCs. Most studies corroborate that the
ability of exosomes to stimulate T cells can be enhanced through
the interaction of exosomes with DCs (145–147).
Exosomes derived from the ascites of patients with ovarian cancer
present tumor antigens and can induce differentiation of dendritic
cells and tumor-specific cytotoxic T lymphocytes (6). Exosomes isolated from the ascites of
patients with ovarian cancer express MHC I molecules, HSP70 and
HSP90. DCs treated with these exosomes can promote T cell
activation and produce cytotoxicity (148). DC-derived exosomes present antigens
to DCs, and then these DCs can activate T cells. These results
suggest that the exosome is a potential safe and feasible
immunotherapy for advanced tumors (149). At present, the antitumor
immunotherapy of exosomes derived from DCs is in phase II clinical
trials of advanced malignant tumors.
Although DCs have antitumorigenic activity, their
function may be inhibited in tumor immune microenvironment. The
ovarian cancer microenvironment is rich in factors that inhibit
monocyte differentiation into DCs. Ascites-derived exosomes from
patients with ovarian cancer induce apoptosis in DCs by activation
of the Fas/FasL pathway and mediating TRAIL apoptosis-inducing
signal molecules in mature DC precursors. In one investigation,
ovarian cancer-derived exosomes were cultured with dendritic
precursor cells for 48 h. Exosome co-cultured DCs had an apoptotic
rate of 12.6% while the control group had an apoptotic rate of 8.6%
(21). Overall, exosomes may induce
apoptosis of DCs and stimulate precursors of mature DCs.
MSCs are an important member of the stem cell
family, which play an important role in cancer progression. They
are present, albeit in small numbers, in a variety of tissues (bone
marrow, umbilical cord blood, umbilical cord, placenta and adipose)
and are reported to have multidirectional differentiation and
regeneration properties (150). In
addition, MSCs also have the capability of immune modulation, which
in a number of cases can cause immunosuppressive effects. De Miguel
et al (151) demonstrated
in vitro that MSCs can inhibit the proliferation of immune
cells (lymphocytes, NK cells and DCs) and inhibit secretion of
cytokines, thereby inhibiting the cytotoxic effect of T and NK
cells via indoleamine 2,3-dioxygenase, while also activating and
inducing the maturation of DCs. A co-culture of ovarian cancer cell
lines (SKOV-3 and OVCAR-3) and MSCs demonstrates that adhesion,
migration, invasion, proliferation and chemical resistance of
ovarian cancer cells is enhanced, leading to accelerated
tumorigenicity (152). In the tumor
microenvironment, exosomes derived from MSCs also play an
immunoregulatory role. Bone marrow MSC-derived exosomes have
anti-inflammatory, anti-apoptotic, pro-angiogenic and
immune-regulating effects (153).
Bone marrow MSC-derived exosomes inhibit the proliferation of T and
B cells and affect mRNA function; downregulating the expression of
CXCL8 and marginal zone B and B1 cell-specific protein the level of
IgM to affect the anti-tumorigenic function of B cells (154). MSC-derived exosomes upregulate
MMP-2 and activate ecto-5′-nucleases, causing tumor cells to become
more malignant and thus altering the tumor microenvironment, as
well as enhancing tumor heterogeneity (155). In addition, cancer cell-derived
exosomes affect the tumorigenicity of MSCs. In vitro
analysis demonstrates that SKOV-3 and OVCAR-3 cell line-derived
exosomes can enhance the migration capacity of MSCs (156). In the microenvironment of ovarian
cancer, cancer stem cells are associated with creating drug
resistance and making the tumor mass refractory to a specific drug
(147,157). Vera et al (158) have revealed that upon treatment
with cisplatin, exosomes released by ovarian cancer that are rich
in cancer stem cells can upregulate IL-6, IL-8 and VEGFA,
increasing the migration capacity of cancer cells. In addition,
factors secreted by MSCs can induce endothelial cell angiogenesis
and accelerate the migration of low-invasive ovarian cancer cells.
Exosomes have also been shown to enhance the oncogenicity of MSCs,
leading to drug resistance and tumor progression (158). The expression of miR-146a in
MSC-derived exosomes is upregulated, which targets laminin γ-2 to
regulate the phosphoinositide 3-kinase (PI3K/Akt) signaling
pathway. This subsequently inhibits the proliferation of ovarian
cancer cells and induces chemotherapy resistance (159).
Previous studies have confirmed the antitumor effect
of MSC-derived exosomes. Human adipose MSC-derived exosomes can
induce apoptotic signals by upregulating pro-apoptotic signaling
via BAX, CASP9, CASP3 and downregulating the anti-apoptotic protein
BCL2 to inhibit A2780 and SKOV-3 cell proliferation, wound-repair
and colony-forming ability (160).
In mouse models of ovarian cancer, paclitaxel-loaded MSC-derived
exosomes have strong antitumor effects, which suggests that they
can be used as drug carriers to target ovarian cancer (161).
The host immune system recognizes the tumor during
progression, allowing immune cells to enter the tumor
microenvironment under the action of chemokines. Subsequently,
immune cells such as CD4+T, CD8+T, B
lymphocytes, NK cells, macrophages and DCs are recruited to
suppress the tumor in vivo. However, during the course of
tumor development, immune monitoring through cancer immune editing
is less selective for cancer cells that are less immunogenic,
allowing them to escape the immune attack and thus achieve immune
escape (Fig. 4). The clinical
manifestations of tumor immune editing trigger the establishment of
an immunosuppressive tumor microenvironment (112).
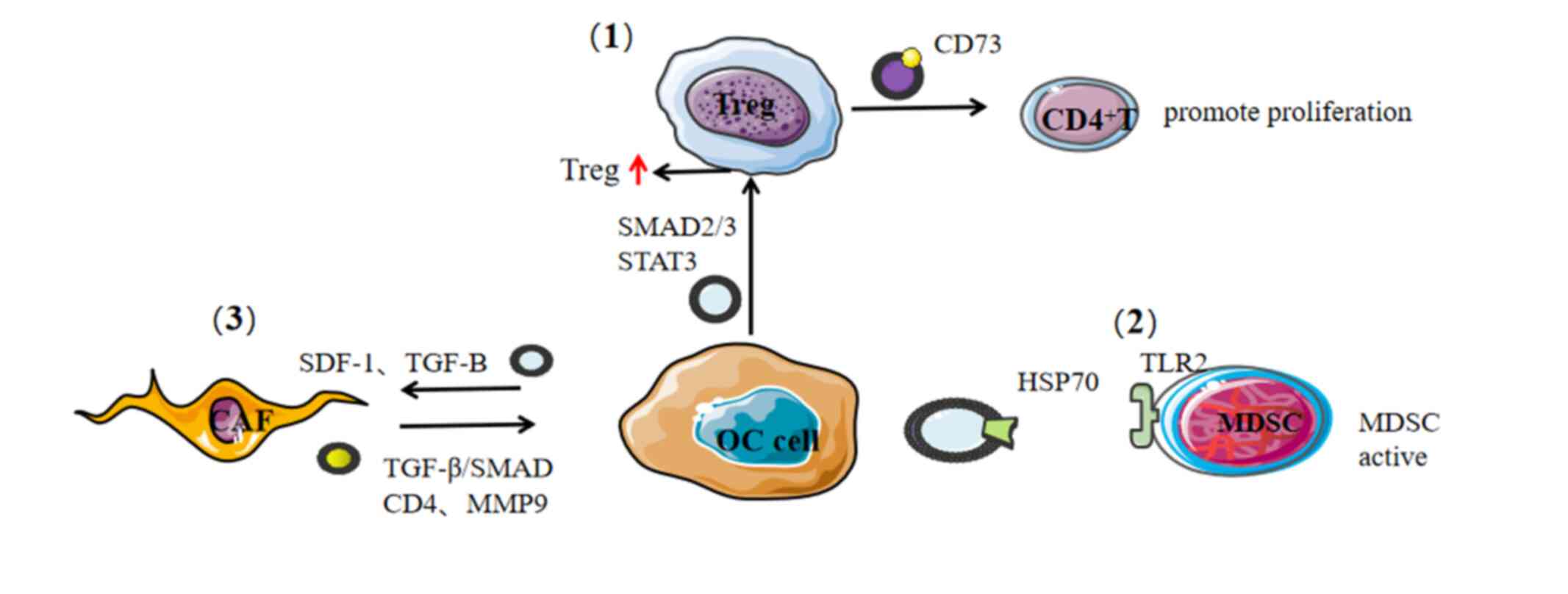
Tregs negatively regulate the antitumor response in
both a direct and indirect manner while also playing a key role in
immune escape (162). The increase
of Tregs in the tumor microenvironment of patients is related to
poor prognosis and shortened overall survival (OS) time (55). Treg-derived exosomes can exert
immunosuppressive effects by expressing CD73 and inhibiting the
proliferation of CD4+T cells (107). Curiel et al (163) in 2004 analyzed 104 specimens of
epithelial ovarian cancer and found that
CD4+CD25+FOXP3+ Tregs inhibited T
cells in vivo and promoted tumor development. After
tumor-derived exosomes activate Tregs, the expression levels of
STAT3/SMAD2/3/IL-10/TGF-B increase and the expression of granzyme
B, perforin and FasL are upregulated, thereby reducing the
antitumor immune response. In addition, exosomes act on the SMAD2/3
and STAT3 signaling pathways to convert CD4+CD25T cells
into CD4+CD25+FOXP3+ Tregs, which
upregulates their immunosuppressive function and anti-apoptotic
potential (164).
MDSC is an immunosuppressive cell of marrow-derived
cells, which are induced to differentiate into DCs, macrophages and
granulocytes. These MDSCs can depress the activity of T and NK
cells, which can significantly suppress immune cell response
(165). IL-6 is produced by
autocrine activation in a Toll-like receptor 2 (TLR2)/myeloid
differentiation primary response 88-dependent manner, triggering
STAT3 activation and promoting the immunosuppressive function of
MDSCs (166). A similar study found
that HSP70 is highly expressed on the surface of ovarian
cancer-derived exosomes, which fuses with and activates MDSCs by
binding TLR2 to promote cancer progression (166,167).
Notably, the study found that the A8 peptide blocked this
HSP70/TLR2 binding to weaken the ability of tumor-derived exosomes
to activate MDSCs in a mouse model. Drugs, such as cisplatin and
5-fluorouracil, cause tumor cells to release more exosomes with
HSP70 surface expression to activate MDSCs. When cisplatin or
5-fluorouracil is used in combination with A8 peptide, it can
effectively antagonize the activation of MDSCs caused by cisplatin
or 5-fluorouracil, which greatly enhances the antitumor effect of
these drugs. Overall, this study has notable implications for novel
ovarian cancer therapies (168).
As an important element of the tumor
microenvironment, CAFs secrete a variety of growth factors and
pro-inflammatory cytokines (TGF-β, VEGF, IL-6 and CXCL12) which
promotes angiogenesis and recruits immunosuppressive cells into the
tumor microenvironment to assist in immune evasion (169). Exosomes derived from epithelial
ovarian cancer induce adipose tissue-derived mesenchymal stem cells
to differentiate into tumor-associated myofibroblasts and
upregulate tumorigenic factors such as stromal cell-derived factor
1 and TGF-β (170). TGF-β receptor
and SMAD signaling can regulate the expression of multifunctional
proteoglycan VERISCAN protein, encoded by the VCAN gene.
Upregulating VCAN activates the NF-kB signal pathway, which
upregulates the expression of CD44, MMP-9 and hyaluronic
acid-mediated motor receptors that collectively promote the
migration and invasion of ovarian cancer cells (171). Further the CAF-derived exosomes can
be internalized by SKOV-3 and CAOV-3 cell lines leading to a more
aggressive tumorous phenotype, promoting the EMT of ovarian tumors.
This evidence suggests that CAF-derived exosomes have the potential
to provide a breakthrough in the treatment of ovarian cancer
(172).
Exosomes can carry a variety of biologically active
molecules. miRNA is a type of non-coding RNA molecule (range, 9–25
nucleotides in length) encoded by an endogenous gene, that
specifically binds to the 3′-untranslated region of target mRNA to
effectively repress gene expression after it has been transcribed
(173,174). Cancer-associated adipocytes and
CAFs transfer miR-21 to cancer cells via exosomes, thereby
inhibiting ovarian cancer cell apoptosis (175). Releasing exosomal miRNA into the
tumor microenvironment is a mechanism for reprogrammed gene
expression at the epigenetic level. Ovarian cancer cells excrete
unnecessary genetic material by releasing exosomes to maintain
their aggressiveness and tumor immunogenicity (176–178).
As a tumor-inhibiting factor, miR-6126 inhibits tumor progression
by decreasing integrin β1 mRNA level to promote metastatic behavior
(177). miR-940 can inhibit ovarian
cancer cell proliferation, colony formation, invasion and
migration, and is highly expressed in exosomes derived from
SKOV3-IP1, HeyA8 and HeyA8-MDR cell lines. Ovarian cancer cells
enhance the tumorigenicity of cells through miRNA excretion
mechanisms (178). A recent study
provided supporting evidence that exosomes derived from epithelial
ovarian cancer cells carry miR-141-3p, which activates the
JAK/STAT3 and NF-kB signaling pathways in endothelial cells. This
increases the level of VEGFR-2 in endothelial cells and enhances
migration and angiogenesis (179).
Exosomal miR-99a-5p derived from epithelial ovarian cancer cells
affects human peritoneal mesothelial cells (HPMCs) by upregulating
fibronectin and vitronectin to promote ovarian cancer progression
(180). miRNAs carried by exosomes
play a pro-tumorigenic role in the immune microenvironment of
ovarian cancer (Table I).
In different ovarian cancer cell lines, the miRNA
profile of exosomes varies. The miR-200 family inhibits EMT, which
is only detected in the exosomes of poorly-invasive cell line
OVCAR-3 (181). In a study of 109
patients with ovarian cancer and eight with ovarian cystadenoma,
exosomal miRNA analysis revealed that miR-200b and miR-320 have a
positive correlation with cellular proliferation and apoptosis.
Additionally, the levels of exosomal miR-200b is related to cancer
antigen 125 (CA125) and the OS rate of patients. Exosomal miR-200b
has the potential to become a new prognostic indicator. The
expression of miR-23a and miR-92a in ovarian cystadenoma-derived
exosomes is lower compared with that of ovarian cancer-derived
exosomes and exosomes derived from healthy individuals (182). Therefore, exosomal miRNAs can be
used as biomarkers of ovarian cancer. In 2018 Kobayashi et
al (183) found that miR-1290
is a potential biomarker for high-grade serous ovarian cancer and
can be used to distinguish patients with other histologically
malignant tumor types. This means that studying miRNAs carried by
exosomes can provide new directions for the early diagnosis of
ovarian cancer and in the search for novel and improved tumor
markers for targeted therapy.
Proteomic analysis of ovarian cancer-derived
exosomes revealed that these exosomes are rich in proteins related
to antigen processing, and that they can effectively initiate
antitumor immune responses (20).
Exosomes from different types of malignant tumors show varying
protein and lipid mass spectra. By comparing the proteome and lipid
profiles of exosomes derived from SKOV-3 cell line and ovarian
surface epithelial cells, it becomes clear that collagen α-2(V)
(also known as COL5A2) and lipoprotein lipase are highly expressed
in SKOV-3 derived exosomes (184).
Plus, CD44 is commonly found in the ovarian cancer-derived exosomes
that become internalized by HPMCs. Increased expression of CD44 in
HPMCs induces HPMCs to secrete MMP9 and allows HPMCs to clear the
mesothelial barrier thus promoting cancer cell invasion and
peritoneal metastasis (185). The
ovarian cancer-derived exosomes promote tumor progression, and the
proteins they carry have a role in malignancy of the tumor. These
proteins include membrane proteins such as programmed cell death
6-interacting protein, tumor susceptibility gene 101, tetraspanins,
HSPs and a variety of enzymes such as phosphate isomerase,
peroxidase, aldehyde reductase and fatty acid synthase (186). Ovarian cancer cell-derived exosomes
that overexpress LIN28 (an RNA-binding protein that promotes
pluripotency) can enhance cell invasion and migration (187). HSP27 can also enhance the
invasiveness and drug resistance of ovarian cancer and is a
potential biological marker of ovarian cancer. Stope et al
(188) demonstrated that exosomes
can carry HSP27 secreted by OVCAR-3 and SKOV-3 cell lines to the
tumor microenvironment, thereby promoting tumor progression.
Cancer immunotherapy is a relatively new treatment
option. By understanding the exosome profile and signal
transduction mechanism, it can be better applied to cancer
treatment. The advantages of exosomes are summarized as follows: i)
Tumor-derived exosomes have the heterogeneity profile of tumor
cells, ii) exosomes derived from homologous or allogeneic cells can
reduce unnecessary immune responses, iii) exosomes have good
stability, iv) the bio-distribution of exosomes can be adjusted by
modifying the surface of exosomes to target a specific tumor
location, v) exosomes have a long half-life and can improve the
efficacy of drugs loaded into them as cargo and vi) exosomes have
multiple types of internalization methods and can avoid the
degradation of lysosomes, so as to efficiently transport drugs to
the recipient cells.
At present, numerous achievements have been made in
applying exosomes to cancer treatment. Small interfering RNAs and
miRNAs carried by exosomes target and inhibit tumor cell
proliferation and drug resistance (175). The reorganization of the exosomal
membrane can improve the efficiency of drug loading and the
sustained release of drugs (189).
Exosomes can overcome the weak immunogenicity of tumor antigens
that are likely to be used in a cancer vaccine (190). DC-derived exosomes can stimulate T
cells by transporting MHC molecular complexes to the surface and
facilitating T cells binding to tumor cells. At present, the
antitumor immunotherapy of DC-derived exosomes has undergone II
clinical trials in advanced non-smooth cell lung carcinoma, showing
the feasibility and safety of antitumor exosome immunotherapy. It
also has a new importance in ovarian cancer therapy (131). Exosomes derived from NK cells have
a natural killing effect on melanomas, which is a potential cancer
immunotherapy strategy (117) and
exosomes have potential for ovarian cancer treatment.
In addition, exosomes can be used as carriers of
antitumor drugs. In mouse models, a combination of mesenchymal stem
cell-derived exosomes and paclitaxel increases the antitumor effect
of paclitaxel (191). Previous
studies have clarified the mechanism of miRNA generated resistance
when carried by exosomes in ovarian cancer (192–194).
Exosomes released by macrophages carry miR-223, which downregulates
the PTEN-PI3K/AKT signaling pathway that can make ovarian cancer
drug resistant (193). In addition,
Kanlikilicer et al (194)
demonstrated that miR-1246 expressed by ovarian cancer-derived
exosomes can make ovarian cancer resistant to paclitaxel via the
Cav1/multidrug resistance protein 1 (p-gp)/M2 phenotype macrophage
axis, miR-1246 targets the Cav1 gene and acts though
platelet-derived growth factor receptor target recipient cells,
induces polarization of M2 macrophages. Exosomes derived from CAFs
carry miR-98-5p to promote the resistance of cisplatin in ovarian
cancer (195). Using specific
exosome inhibitors can effectively prevent this mechanism of drug
resistance. Ovarian cancer-derived exosomes are enriched with DNA
methyltransferase 1 that makes cancer cells resistant to cisplatin,
but the exosomal inhibitor gw4869 can reverse this resistance and
restore their drug sensitivity (196). This information will provide new
avenues of exploration for targeted therapies against ovarian
cancer. Cancer-derived exosomes can carry CRISPR/Cas9 to other
ovarian cancer cells, inhibit PARP-1 expression, cause ovarian
cancer cell apoptosis and enhance the sensitivity to cisplatin
(197). Until now, there have been
no reports on the application of exosomes to ovarian cancer
immunotherapy, but it is an area should continue to be explored in
the future.
As outlined in this article, the role of exosomes
in the immune microenvironment of ovarian cancer can be described
as a double-edged sword. Exosomes derived from immune cells can
target tumor cells to exert antitumor immune effects. NK
cell-derived exosomes mediate NK cell cytotoxicity through their
surface receptors NKG2D and DNAX accessory molecule-1 (115,116);
NK cell-derived exosomes can carry killer protein (CD56), FasL,
perforin, granulysin, granzymes A and B to the tumor
microenvironment of distant tumors (114). DC-derived exosomes can activate T
cells to exert antitumor effects (145). Exosomes can also mediate cellular
communication between immune cells. Mast cell-derived exosomes can
promote the maturation of DCs (144). T cell-derived exosomes can regulate
miRNA and TCR-rich vesicles to regulate gene expression and
extracellular signal transduction of DCs (125). Treg-derived exosomes can inhibit
CD4+T cell proliferation by expressing CD73 (140).
However, tumor cell-derived exosomes exert
immunosuppression and immune escape through a variety of pathways
in the tumor microenvironment. Exosomes derived from epithelial
ovarian cancer can induce macrophages to differentiate into TAMs,
downregulate the killing effect of NK cells on tumors, induce T
cell apoptosis through Fas/FasL interactions and induce DC
apoptosis. Ovarian cancer-derived exosomes also upregulate the
functions of Tregs and MDSCs, induce the differentiation of CAFs
and induce the tumorigenic activity of mesenchymal stem cells;
forming a microenvironment that is beneficial to tumor
proliferation, invasion, metastasis and tumor progression.
A study has shown that the tumor microenvironment
contains functionally heterogeneous B lymphocytes and regulates
tumor immunity by producing immunoglobulins and presenting
costimulatory molecules (198).
However, in ovarian cancer, research on the interaction between
exosomes and B lymphocytes is rare, and is thus an area that
requires further exploration. The regulation of the immune system
by exosomes highlights the great potential of exosomes in cancer
immunotherapy. The release of exosomes in patients with ovarian
cancer is 3–4 times higher compared with in individuals without
ovarian cancer. If the production of tumor-derived exosomes can be
reduced, this could theoretically weaken the impact on immune
suppression and thus would make immune escape more difficult. At
present, preventing the excessive production of cancer cell-derived
exosome has shown significant antitumor and anti-metastatic effects
in breast cancer (199). If this
technology can be applied to ovarian cancer, it will become a new
strategy for ovarian cancer treatment.
Chemoresistance is common during the treatment of
ovarian cancer and is usually associated with poor prognosis.
Current biological techniques can effectively load chemotherapeutic
drugs into exosomes through co-culture, electroporation or
ultrasound (200). Studies have
verified that exosomes loaded with paclitaxel and cisplatin can
induce apoptosis of ovarian cancer cells (175,192,194).
If the exosomes loaded with paclitaxel and cisplatin can be used in
clinical treatment, it will be a novel strategy for the treatment
of drug-resistant ovarian cancer. Of course, this requires a large
number of clinical trials for the verification of the treatment
efficacy, and would require the joint efforts of various research
centers and hospitals. At present, Clinicaltrials.gov (https://clinicaltrials.gov/) has reported 198 studies
on exosomes, including three studies on exosomes as biomarkers of
ovarian cancer, and one study on polycystic ovary syndrome and
exosomes. There are no clinical trials using exosomes in the
treatment of ovarian cancer, to the best of our knowledge. Before
clinical trials, large-scale separation and purification of
exosomes is still a huge challenge. Fortunately, research on the
production of exosome mimics has made preliminary progress. Pisano
et al (201) used monocytes
as raw materials to produce exosome mimics through filters of
different porosity and size exclusion chromatography columns. The
development of immune-derived exosome mimics is expected to solve
the problems of yield and reproducibility, which greatly improves
the feasibility of applying exosomes to clinical trials (201).
The RNA, protein and lipid profiles of exosomes
derived from different ovarian tumors are different, and the
circular RNAs carried by ovarian tumor cell-derived exosomes are
also different from healthy volunteers (134). In addition, serum exosomal
piwi-interacting RNAs are considered to be a promising biomarker
for patients with gastric cancer (202). Researchers have found that the
detection level of CA125 in exosomes is higher compared with that
in serum, which significantly improves the sensitivity of ovarian
cancer diagnosis (203). This
suggests that exosomes have the potential to become biomarkers for
clinical analysis of ovarian cancer. With the development of
biochips, microfluidic Raman biochips have been successfully used
to monitor exosomes in prostate clinical serum samples (204). These devices can be similarly
applied for ovarian cancer investigations based on exosomes.
Not applicable.
This study was supported by The National Natural
Science Foundation of China (grant no. 81472761) and The Natural
Science Foundation of Tianjin City (grant no. 14JCYBJC25300).
GYL designed the review. XL drafted the manuscript
and prepared the figures. YL, TYZ, SSZ, JZZ, JW, and YS helped to
modify the manuscript. All authors read and approved the final
manuscript.
Not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Drakes ML and Stiff PJ: Regulation of
ovarian cancer prognosis by immune cells in the tumor
microenvironment. Cancers (Basel). 10:3022018. View Article : Google Scholar
|
|
3
|
Gajewski TF, Schreiber H and Fu YX: Innate
and adaptive immune cells in the tumor microenvironment. Nat
Immunol. 14:1014–1022. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Albini A, Bruno A, Noonan DM and Mortara
L: Contribution to tumor angiogenesis from innate immune cells
within the tumor microenvironment: Implications for immunotherapy.
Front Immunol. 9:5272018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cassim S and Pouyssegur J: Tumor
microenvironment: A metabolic player that shapes the immune
response. Int J Mol Sci. 21:1572019. View Article : Google Scholar
|
|
6
|
Andre F, Schartz NE, Movassagh M, Flament
C, Pautier P, Morice P, Pomel C, Lhomme C, Escudier B, Le Chevalier
T, et al: Malignant effusions and immunogenic tumour-derived
exosomes. Lancet. 360:295–305. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Giordano C, La Camera G, Gelsomino L,
Barone I, Bonofiglio D, Ando S and Catalano S: The biology of
exosomes in breast cancer progression: Dissemination, immune
evasion and metastatic colonization. Cancers (Basel). 12:21792020.
View Article : Google Scholar
|
|
8
|
Kulkarni B, Kirave P, Gondaliya P, Jash K,
Jain A, Tekade RK and Kalia K: Exosomal miRNA in chemoresistance,
immune evasion, metastasis and progression of cancer. Drug Discov
Today. 24:2058–2067. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lundholm M, Schroder M, Nagaeva O, Baranov
V, Widmark A, Mincheva-Nilsson L and Wikström P: Prostate
tumor-derived exosomes down-regulate NKG2D expression on natural
killer cells and CD8+ T cells: Mechanism of immune evasion. PLoS
One. 9:e1089252014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheng L, Wu S, Zhang K, Qing Y and Xu T: A
comprehensive overview of exosomes in ovarian cancer: Emerging
biomarkers and therapeutic strategies. J Ovarian Res. 10:732017.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Beach A, Zhang HG, Ratajczak MZ and Kakar
SS: Exosomes: An overview of biogenesis, composition and role in
ovarian cancer. J Ovarian Res. 7:142014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Doyle LM and Wang MZ: Overview of
extracellular vesicles, their origin, composition, purpose, and
methods for exosome isolation and analysis. Cells. 8:7272019.
View Article : Google Scholar
|
|
13
|
Sharma A and Johnson A: Exosome DNA:
Critical regulator of tumor immunity and a diagnostic biomarker. J
Cell Physiol. 235:1921–1932. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Iliev D, Strandskog G, Nepal A, Aspar A,
Olsen R, Jørgensen J, Wolfson D, Ahluwalia BS, Handzhiyski J and
Mironova R: Stimulation of exosome release by extracellular DNA is
conserved across multiple cell types. FEBS J. 285:3114–3133. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tang YT, Huang YY, Zheng L, Qin SH, Xu XP,
An TX, Xu Y, Wu YS, Hu XM, Ping BH and Wang Q: Comparison of
isolation methods of exosomes and exosomal RNA from cell culture
medium and serum. Int J Mol Med. 40:834–844. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shyu KG, Wang BW, Pan CM, Fang WJ and Lin
CM: Hyperbaric oxygen boosts long noncoding RNA MALAT1 exosome
secretion to suppress microRNA-92a expression in therapeutic
angiogenesis. Int J Cardiol. 274:271–278. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hannafon BN, Trigoso YD, Calloway CL, Zhao
YD, Lum DH, Welm AL, Zhao ZJ, Blick KE, Dooley WC and Ding WQ:
Plasma exosome microRNAs are indicative of breast cancer. Breast
Cancer Res. 18:902016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kobayashi M, Sawada K, Miyamoto M, Shimizu
A, Yamamoto M, Kinose Y, Nakamura K, Kawano M, Kodama M, Hashimoto
K and Kimura T: Exploring the potential of engineered exosomes as
delivery systems for tumor-suppressor microRNA replacement therapy
in ovarian cancer. Biochem Biophys Res Commun. 527:153–161. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mittal S, Gupta P, Chaluvally-Raghavan P
and Pradeep S: Emerging role of extracellular vesicles in immune
regulation and cancer progression. Cancers (Basel). 12:35632020.
View Article : Google Scholar
|
|
20
|
Liang B, Peng P, Chen S, Li L, Zhang M,
Cao D, Yang J, Li H, Gui T, Li X and Shen K: Characterization and
proteomic analysis of ovarian cancer-derived exosomes. J
Proteomics. 80:171–182. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Peng P, Yan Y and Keng S: Exosomes in the
ascites of ovarian cancer patients: Origin and effects on
anti-tumor immunity. Oncol Rep. 25:749–762. 2011.PubMed/NCBI
|
|
22
|
Luo Z, Wang Q, Lau WB, Lau B, Xu L, Zhao
L, Yang H, Feng M, Xuan Y, Yang Y, et al: Tumor microenvironment:
The culprit for ovarian cancer metastasis? Cancer Lett.
377:174–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Da Silva AC, Jammal MP, Crispim PCA, Murta
EFC and Nomelini RS: The role of stroma in ovarian cancer. Immunol
Invest. 49:406–424. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Josephs SF, Ichim TE, Prince SM, Kesari S,
Marincola FM, Escobedo AR and Jafri A: Unleashing endogenous
TNF-alpha as a cancer immunotherapeutic. J Transl Med. 16:2422018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wen Z, Liu H, Li M, Li B, Gao W, Shao Q,
Fan B, Zhao F, Wang Q, Xie Q, et al: Increased metabolites of
5-lipoxygenase from hypoxic ovarian cancer cells promote
tumor-associated macrophage infiltration. Oncogene. 34:1241–1252.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Browning L, Patel MR, Horvath EB, Tawara K
and Jorcyk CL: IL-6 and ovarian cancer: Inflammatory cytokines in
promotion of metastasis. Cancer Manag Res. 10:6685–6693. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sanmarco LM, Ponce NE, Visconti LM,
Eberhardt N, Theumer MG, Minguez AR and Aoki MP: IL-6 promotes M2
macrophage polarization by modulating purinergic signaling and
regulates the lethal release of nitric oxide during Trypanosoma
cruzi infection. Biochim Biophys Acta Mol Basis Dis. 1863:857–869.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yin Z, Ma T, Lin Y, Lu X, Zhang C, Chen S
and Jian Z: IL-6/STAT3 pathway intermediates M1/M2 macrophage
polarization during the development of hepatocellular carcinoma. J
Cell Biochem. 119:9419–9432. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fu XL, Duan W, Su CY, Mao FY, Lv YP, Teng
YS, Yu PW, Zhuang Y and Zhao YL: Interleukin 6 induces M2
macrophage differentiation by STAT3 activation that correlates with
gastric cancer progression. Cancer Immunol Immunother.
66:1597–1608. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jiang B, Zhu SJ, Xiao SS and Xue M:
miR-217 Inhibits M2-like macrophage polarization by suppressing
secretion of interleukin-6 in ovarian cancer. Inflammation.
42:1517–1529. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McLean K, Tan L, Bolland DE, Coffman LG,
Peterson LF, Talpaz M, Neamati N and Buckanovich RJ: Leukemia
inhibitory factor functions in parallel with interleukin-6 to
promote ovarian cancer growth. Oncogene. 38:1576–1584. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Y, Zong X, Mitra S, Mitra AK, Matei D
and Nephew KP: IL-6 mediates platinum-induced enrichment of ovarian
cancer stem cells. JCI Insight. 3:e1223602018. View Article : Google Scholar
|
|
33
|
Bretz NP, Ridinger J, Rupp AK, Rimbach K,
Keller S, Rupp C, Marmé F, Umansky L, Umansky V, Eigenbrod T, et
al: Body fluid exosomes promote secretion of inflammatory cytokines
in monocytic cells via Toll-like receptor signaling. J Biol Chem.
288:36691–3702. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
De Marco M, Falco A, Festa M, Raffone A,
Sandullo L, Rosati A, Reppucci F, Cammarota AL, Esposito F, Matassa
DS, et al: Different mechanisms underlie IL-6 release in
chemosensitive and chemoresistant ovarian carcinoma cells. Am J
Cancer Res. 10:2596–2602. 2020.PubMed/NCBI
|
|
35
|
Kumari N, Dwarakanath BS, Das A and Bhatt
AN: Role of interleukin-6 in cancer progression and therapeutic
resistance. Tumour Biol. 37:11553–11572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang Y, Niu XL, Qu Y, Wu J, Zhu YQ, Sun WJ
and Li LZ: Autocrine production of interleukin-6 confers cisplatin
and paclitaxel resistance in ovarian cancer cells. Cancer Lett.
295:110–123. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kampan NC, Madondo MT, Reynolds J, Hallo
J, McNally OM, Jobling TW, Stephens AN, Quinn MA and Plebanski M:
Pre-operative sera interleukin-6 in the diagnosis of high-grade
serous ovarian cancer. Sci Rep. 10:22132020. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Isobe A, Sawada K, Kinose Y, Ohyagi-Hara
C, Nakatsuka E, Makino H, Ogura T, Mizuno T, Suzuki N, Morii E, et
al: Interleukin 6 receptor is an independent prognostic factor and
a potential therapeutic target of ovarian cancer. PLoS One.
10:e01180802015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yuan X, Zhang J, Li D, Mao Y, Mo F, Du W
and Ma X: Prognostic significance of tumor-associated macrophages
in ovarian cancer: A meta-analysis. Gynecol Oncol. 147:181–187.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Maccio A, Gramignano G, Cherchi MC, Tanca
L, Melis L and Madeddu C: Role of M1-polarized tumor-associated
macrophages in the prognosis of advanced ovarian cancer patients.
Sci Rep. 10:60962020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Carroll MJ, Kapur A, Felder M, Patankar MS
and Kreeger PK: M2 macrophages induce ovarian cancer cell
proliferation via a heparin binding epidermal growth factor/matrix
metalloproteinase 9 intercellular feedback loop. Oncotarget.
7:86608–86620. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou ZN, Sharma VP, Beaty BT, Roh-Johnson
M, Peterson EA, Van Rooijen N, Kenny PA, Wiley HS, Condeelis JS and
Segall JE: Autocrine HBEGF expression promotes breast cancer
intravasation, metastasis and macrophage-independent invasion in
vivo. Oncogene. 33:3784–3793. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Haque S and Morris JC: Transforming growth
factor-β: A therapeutic target for cancer. Hum Vaccin Immunother.
13:1741–1750. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Steitz AM, Steffes A, Finkernagel F, Unger
A, Sommerfeld L, Jansen JM, Wagner U, Graumann J, Müller R and
Reinartz S: Tumor-associated macrophages promote ovarian cancer
cell migration by secreting transforming growth factor beta induced
(TGFBI) and tenascin C. Cell Death Dis. 11:2492020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dasari S, Fang Y and Mitra AK: Cancer
associated fibroblasts: Naughty neighbors that drive ovarian cancer
progression. Cancers (Basel). 10:4062018. View Article : Google Scholar
|
|
46
|
Gao Q, Yang Z, Xu S, Li X, Yang X, Jin P,
Liu Y, Zhou X, Zhang T, Gong C, et al: Heterotypic CAF-tumor
spheroids promote early peritoneal metastatis of ovarian cancer. J
Exp Med. 216:688–703. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhuang J, Lu Q, Shen B, Huang X, Shen L,
Zheng X, Huang R, Yan J and Guo H: TGFβ1 secreted by
cancer-associated fibroblasts induces epithelial-mesenchymal
transition of bladder cancer cells through lncRNA-ZEB2NAT. Sci Rep.
5:119242015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kan T, Wang W, Ip PP, Zhou S, Wong AS,
Wang X and Yang M: Single-cell EMT-related transcriptional analysis
revealed intra-cluster heterogeneity of tumor cell clusters in
epithelial ovarian cancer ascites. Oncogene. 39:4227–4240. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Han Q, Huang B, Huang Z, Cai J, Gong L,
Zhang Y, Jiang J, Dong W and Wang Z: Tumor cell-fibroblast
heterotypic aggregates in malignant ascites of patients with
ovarian cancer. Int J Mol Med. 44:2245–2255. 2019.PubMed/NCBI
|
|
50
|
Liu CL, Pan HW, Torng PL, Fan MH and Mao
TL: SRPX and HMCN1 regulate cancer-associated fibroblasts to
promote the invasiveness of ovarian carcinoma. Oncol Rep.
42:2706–2715. 2019.PubMed/NCBI
|
|
51
|
De Cicco P, Ercolano G and Ianaro A: The
new era of cancer immunotherapy: Targeting myeloid-derived
suppressor cells to overcome immune evasion. Front Immunol.
11:16802020. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Sasidharan Nair V and Elkord E: Immune
checkpoint inhibitors in cancer therapy: A focus on T-regulatory
cells. Immunol Cell Biol. 96:21–33. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li X, Wang J, Wu W, Gao H, Liu N, Zhan G,
Li L, Han L and Guo X: Myeloid-derived suppressor cells promote
epithelial ovarian cancer cell stemness by inducing the
CSF2/p-STAT3 signalling pathway. FEBS J. 287:5218–5235. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Horikawa N, Abiko K, Matsumura N,
Hamanishi J, Baba T, Yamaguchi K, Yoshioka Y, Koshiyama M and
Konishi I: Expression of vascular endothelial growth factor in
ovarian cancer inhibits tumor immunity through the accumulation of
myeloid-derived suppressor cells. Clin Cancer Res. 23:587–599.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Dutsch-Wicherek MM, Szubert S, Dziobek K,
Wisniewski M, Lukaszewska E, Wicherek L, Jozwicki W, Rokita W and
Koper K: Analysis of the treg cell population in the peripheral
blood of ovarian cancer patients in relation to the long-term
outcomes. Ginekol Pol. 90:179–184. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Batlle E and Massague J: Transforming
growth Factor-beta signaling in immunity and cancer. Immunity.
50:924–940. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhou J, Jiang W, Huang W, Ye M and Zhu X:
Prognostic values of transforming growth Factor-Beta subtypes in
ovarian cancer. Biomed Res Int. 2020:21706062020.PubMed/NCBI
|
|
58
|
Wen H, Qian M, He J, Li M, Yu Q and Leng
Z: Inhibiting of self-renewal, migration and invasion of ovarian
cancer stem cells by blocking TGF-beta pathway. PLoS One.
15:e02302302020. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bai Y, Li LD, Li J, Chen RF, Yu HL, Sun
HF, Wang JY and Lu X: A FXYD5/TGFβ/SMAD positive feedback loop
drives epithelial-to-mesenchymal transition and promotes tumor
growth and metastasis in ovarian cancer. Int J Oncol. 56:301–314.
2020.PubMed/NCBI
|
|
60
|
Fukui S, Nagasaka K, Miyagawa Y,
Kikuchi-Koike R, Kawata Y, Kanda R, Ichinose T, Sugihara T, Hiraike
H, Wada-Hiraike O, et al: The proteasome deubiquitinase inhibitor
bAP15 downregulates TGF-β/Smad signaling and induces apoptosis via
UCHL5 inhibition in ovarian cancer. Oncotarget. 10:5932–5948. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kulbe H, Chakravarty P, Leinster DA,
Charles KA, Kwong J, Thompson RG, Coward JI, Schioppa T, Robinson
SC, Gallagher WM, et al: A dynamic inflammatory cytokine network in
the human ovarian cancer microenvironment. Cancer Res. 72:66–75.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Carbotti G, Petretto A, Naschberger E,
Sturzl M, Martini S, Mingari MC, Filaci G, Ferrini S and Fabbi M:
Cytokine-Induced Guanylate Binding Protein 1 (GBP1) release from
human ovarian cancer cells. Cancers (Basel). 12:4882020. View Article : Google Scholar
|
|
63
|
Zhao H, Yang L, Baddour J, Achreja A,
Bernard V, Moss T, Marini JC, Tudawe T, Seviour EG, San Lucas FA,
et al: Tumor microenvironment derived exosomes pleiotropically
modulate cancer cell metabolism. Elife. 5:e102502016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Whiteside TL: The effect of tumor-derived
exosomes on immune regulation and cancer immunotherapy. Future
Oncol. 13:2583–2592. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Gong M, Yu B, Wang J, Wang Y, Liu M, Paul
C, Millard RW, Xiao DS, Ashraf M and Xu M: Mesenchymal stem cells
release exosomes that transfer miRNAs to endothelial cells and
promote angiogenesis. Oncotarget. 8:45200–45212. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Wu Q, Zhou L, Lv D, Zhu X and Tang H:
Exosome-mediated communication in the tumor microenvironment
contributes to hepatocellular carcinoma development and
progression. J Hematol Oncol. 12:532019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Dorayappan KDP, Wanner R, Wallbillich JJ,
Saini U, Zingarelli R, Suarez AA, Cohn DE and Selvendiran K:
Hypoxia-induced exosomes contribute to a more aggressive and
chemoresistant ovarian cancer phenotype: A novel mechanism linking
STAT3/Rab proteins. Oncogene. 37:3806–3821. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Matias Ostrowski NBC, Sophie Krumeich,
Isabelle Fanget, Graça Raposo, Ariel Savina, et al: Rab27a and
Rab27b control different steps of the exosome secretion pathway.
Nat Cell Biol. 12:19–30. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Qiu JJ, Lin XJ, Tang XY, Zheng TT, Lin YY
and Hua KQ: Exosomal metastasis associated lung adenocarcinoma
Transcript 1 promotes angiogenesis and predicts poor prognosis in
epithelial ovarian cancer. Int J Biol Sci. 14:1960–1973. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Tang MKS, Yue PYK, Ip PP, Huang RL, Lai
HC, Cheung ANY, Tse KY, Ngan HYS and Wong AST: Soluble E-cadherin
promotes tumor angiogenesis and localizes to exosome surface. Nat
Commun. 9:22702018. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Runz S, Keller S, Rupp C, Stoeck A, Issa
Y, Koensgen D, Mustea A, Sehouli J, Kristiansen G and Altevogt P:
Malignant ascites-derived exosomes of ovarian carcinoma patients
contain CD24 and EpCAM. Gynecol Oncol. 107:563–571. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Lei X, Lei Y, Li JK, Du WX, Li RG, Yang J,
Li J, Li F and Tan HB: Immune cells within the tumor
microenvironment: Biological functions and roles in cancer
immunotherapy. Cancer Lett. 470:126–133. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Xu SJ, Hu HT, Li HL and Chang S: The role
of miRNAs in immune cell development, immune cell activation, and
tumor immunity: With a focus on macrophages and natural killer
cells. Cells. 8:11402019. View Article : Google Scholar
|
|
74
|
Rhee I: Diverse macrophages polarization
in tumor microenvironment. Arch Pharm Res. 39:1588–1596. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tashiro-Yamaji J, Kubota T and Yoshida R:
Macrophage MHC receptor 2: A novel receptor on allograft
(H-2D(d)K(d))-induced macrophage (H-2D(b)K(b)) recognizing an MHC
class I molecule, H-2K(d), in mice. Gene. 384:1–8. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Chen X, Zhou J, Li X and Wang X, Lin Y and
Wang X: Exosomes derived from hypoxic epithelial ovarian cancer
cells deliver microRNAs to macrophages and elicit a tumor-promoted
phenotype. Cancer Lett. 435:80–91. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Ruffell B, Affara NI and Coussens LM:
Differential macrophage programming in the tumor microenvironment.
Trends Immunol. 33:119–126. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Shapouri-Moghaddam A, Mohammadian S,
Vazini H, Taghadosi M, Esmaeili SA, Mardani F, Seifi B, Mohammadi
A, Afshari JT and Sahebkar A: Macrophage plasticity, polarization,
and function in health and disease. J Cell Physiol. 233:6425–6440.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Funes SC, Rios M, Escobar-Vera J and
Kalergis AM: Implications of macrophage polarization in
autoimmunity. Immunology. 154:186–195. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Madeddu C, Gramignano G, Kotsonis P, Coghe
F, Atzeni V, Scartozzi M and Macciò A: Microenvironmental M1
tumor-associated macrophage polarization influences cancer-related
anemia in advanced ovarian cancer: Key role of interleukin-6.
Haematologica. 103:e388–e391. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Reinartz S, Schumann T, Finkernagel F,
Wortmann A, Jansen JM, Meissner W, Krause M, Schwörer AM, Wagner U,
Müller-Brüsselbach S and Müller R: Mixed-polarization phenotype of
ascites-associated macrophages in human ovarian carcinoma:
Correlation of CD163 expression, cytokine levels and early relapse.
Int J Cancer. 134:32–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Nowak M and Klink M: The role of
tumor-associated macrophages in the progression and chemoresistance
of ovarian cancer. Cells. 9:12992020. View Article : Google Scholar
|
|
83
|
Zhu Q, Wu X, Wu Y and Wang X: Interaction
between Treg cells and tumor-associated macrophages in the tumor
microenvironment of epithelial ovarian cancer. Oncol Rep.
36:3472–3478. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhou J, Li X, Wu X, Zhang T, Zhu Q and
Wang X, Wang H, Wang K, Lin Y and Wang X: Exosomes released from
tumor-associated macrophages transfer miRNAs that induce a
Treg/Th17 cell imbalance in epithelial ovarian cancer. Cancer
Immunol Res. 6:1578–1592. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Huang YJ, Huang TH, Yadav VK, Sumitra MR,
Tzeng DT, Wei PL, Shih JW and Wu AT: Preclinical investigation of
ovatodiolide as a potential inhibitor of colon cancer stem cells
via downregulating sphere-derived exosomal
beta-catenin/STAT3/miR-1246 cargoes. Am J Cancer Res. 10:2337–2354.
2020.PubMed/NCBI
|
|
86
|
Xiao L, He Y, Peng F, Yang J and Yuan C:
Endometrial cancer cells promote M2-like macrophage polarization by
delivering exosomal miRNA-21 under hypoxia condition. J Immunol
Res. 2020:97310492020. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wang X, Luo G, Zhang K, Cao J, Huang C,
Jiang T, Liu B, Su L and Qiu Z: Hypoxic tumor-derived exosomal
miR-301a Mediates M2 macrophage polarization via PTEN/PI3Kgamma to
promote pancreatic cancer metastasis. Cancer Res. 78:4586–4598.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Bardi GT, Smith MA and Hood JL: Melanoma
exosomes promote mixed M1 and M2 macrophage polarization. Cytokine.
105:63–72. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Li X, Lei Y, Wu M and Li N: Regulation of
macrophage activation and polarization by HCC-derived exosomal
lncRNA TUC339. Int J Mol Sci. 19:29582018. View Article : Google Scholar
|
|
90
|
Piao YJ, Kim HS, Hwang EH, Woo J, Zhang M
and Moon WK: Breast cancer cell-derived exosomes and macrophage
polarization are associated with lymph node metastasis. Oncotarget.
9:7398–7410. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Pritchard A, Tousif S, Wang Y, Hough K,
Khan S, Strenkowski J, Chacko BK, Darley-Usmar VM and Deshane JS:
Lung tumor cell-derived exosomes promote M2 macrophage
polarization. Cells. 9:13032020. View Article : Google Scholar
|
|
92
|
Chen X, Ying X and Wang X, Wu X, Zhu Q and
Wang X: Exosomes derived from hypoxic epithelial ovarian cancer
deliver microRNA-940 to induce macrophage M2 polarization. Oncol
Rep. 38:522–528. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Li X and Tang M: Exosomes released from M2
macrophages transfer miR-221-3p contributed to EOC progression
through targeting CDKN1B. Cancer Med. 9:5976–5988. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Ying X, Wu Q, Wu X, Zhu Q and Wang X,
Jiang L, Chen X and Wang X: Epithelial ovarian cancer-secreted
exosomal miR-222-3p induces polarization of tumor-associated
macrophages. Oncotarget. 7:43076–43087. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Wu Q, Wu X, Ying X, Zhu Q and Wang X,
Jiang L, Chen X, Wu Y and Wang X: Suppression of endothelial cell
migration by tumor associated macrophage-derived exosomes is
reversed by epithelial ovarian cancer exosomal lncRNA. Cancer Cell
Int. 17:622017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Hu Y, Li D, Wu A, Qiu X, Di W, Huang L and
Qiu L: TWEAK-stimulated macrophages inhibit metastasis of
epithelial ovarian cancer via exosomal shuttling of microRNA.
Cancer Lett. 393:60–67. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Baj-Krzyworzeka M, Szatanek R, Weglarczyk
K, Baran J and Zembala M: Tumour-derived microvesicles modulate
biological activity of human monocytes. Immunol Lett. 113:76–82.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Baj-Krzyworzeka M, Baran J, Weglarczyk K,
Szatanek R, Szaflarska A, Siedlar M and Zembala M: Tumour-derived
microvesicles (TMV) mimic the effect of tumour cells on monocyte
subpopulations. Anticancer Res. 30:3515–3520. 2010.PubMed/NCBI
|
|
99
|
Baj-Krzyworzeka M, Mytar B, Szatanek R,
Surmiak M, Weglarczyk K, Baran J and Siedlar M: Colorectal
cancer-derived microvesicles modulate differentiation of human
monocytes to macrophages. J Transl Med. 14:362016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Moradi-Chaleshtori M, Bandehpour M, Soudi
S, Mohammadi-Yeganeh S and Hashemi SM: In vitro and in vivo
evaluation of anti-tumoral effect of M1 phenotype induction in
macrophages by miR-130 and miR-33 containing exosomes. Cancer
Immunol Immunother. 2020.(Epub ahead of print). View Article : Google Scholar
|
|
101
|
Yue S, Ye X, Zhou T, Gan D, Qian H, Fang
W, Yao M, Zhang D, Shi H and Chen T: PGRN−/−
TAMs-derived exosomes inhibit breast cancer cell invasion and
migration and its mechanism exploration. Life Sci. 264:1186872020.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Han JJ, Yu M, Houston N, Steinberg SM and
Kohn EC: Progranulin is a potential prognostic biomarker in
advanced epithelial ovarian cancers. Gynecol Oncol. 120:5–10. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Carlson AM, Maurer MJ, Goergen KM, Kalli
KR, Erskine CL, Behrens MD, Knutson KL and Block MS: Utility of
progranulin and serum leukocyte protease inhibitor as diagnostic
and prognostic biomarkers in ovarian cancer. Cancer Epidemiol
Biomarkers Prev. 22:1730–1735. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Dong T, Yang D, Li R, Zhang L, Zhao H,
Shen Y, Zhang X, Kong B and Wang L: PGRN promotes migration and
invasion of epithelial ovarian cancer cells through an epithelial
mesenchymal transition program and the activation of cancer
associated fibroblasts. Exp Mol Pathol. 100:17–25. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Dou R, Hong Z, Tan X, Hu F, Ding Y, Wang
W, Liang Z, Zhong R, Wu X and Weng X: Fas/FasL interaction mediates
imbalanced cytokine/cytotoxicity responses of iNKT cells against
Jurkat cells. Mol Immunol. 99:145–153. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Wallin RP, Screpanti V, Michaelsson J,
Grandien A and Ljunggren HG: Regulation of perforin-independent NK
cell-mediated cytotoxicity. Eur J Immunol. 33:2727–2735. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Tanaka H, Kai S, Yamaguchi M, Misawa M,
Fujimori Y, Yamamoto M and Hara H: Analysis of natural killer (NK)
cell activity and adhesion molecules on NK cells from umbilical
cord blood. Eur J Haematol. 71:29–38. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Konjevic GM, Vuletic AM, Mirjacic
Martinovic KM, Larsen AK and Jurisic VB: The role of cytokines in
the regulation of NK cells in the tumor environment. Cytokine.
117:30–40. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Pahl JHW, Cerwenka A and Ni J: Memory-Like
NK Cells: Remembering a previous activation by cytokines and NK
cell receptors. Front Immunol. 9:27962018. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Gianchecchi E, Delfino DV and Fierabracci
A: NK cells in autoimmune diseases: Linking innate and adaptive
immune responses. Autoimmun Rev. 17:142–154. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Zitti B and Bryceson YT: Natural killer
cells in inflammation and autoimmunity. Cytokine Growth Factor Rev.
42:37–46. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Rodriguez GM, Galpin KJC, McCloskey CW and
Vanderhyden BC: The tumor microenvironment of epithelial ovarian
cancer and its influence on response to immunotherapy. Cancers
(Basel). 10:2422018. View Article : Google Scholar
|
|
113
|
Zhu L, Kalimuthu S, Oh JM, Gangadaran P,
Baek SH, Jeong SY, Lee SW, Lee J and Ahn BC: Enhancement of
antitumor potency of extracellular vesicles derived from natural
killer cells by IL-15 priming. Biomaterials. 190-191:38–50. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Jong AY, Wu CH, Li J, Sun J, Fabbri M,
Wayne AS and Seeger RC: Large-scale isolation and cytotoxicity of
extracellular vesicles derived from activated human natural killer
cells. J Extracell Vesicles. 6:12943682017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Lugini L, Cecchetti S, Huber V, Luciani F,
Macchia G, Spadaro F, Paris L, Abalsamo L, Colone M, Molinari A, et
al: Immune surveillance properties of human NK cell-derived
exosomes. J Immunol. 189:2833–2842. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Di Pace A, Tumino N, Besi F, Alicata C,
Conti LA, Munari E, Maggi E, Vacca P and Moretta L:
Characterization of human NK cell-derived exosomes: Role of DNAM1
receptor in exosome-mediated cytotoxicity against tumor. Cancers
(Basel). 12:6612020. View Article : Google Scholar
|
|
117
|
Zhu L, Kalimuthu S, Gangadaran P, Oh JM,
Lee HW, Baek SH, Jeong SY, Lee SW, Lee J and Ahn BC: Exosomes
derived from natural killer cells exert therapeutic effect in
melanoma. Theranostics. 7:2732–2745. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Neviani P, Wise PM, Murtadha M, Liu CW, Wu
CH, Jong AY, Seeger RC and Fabbri M: Natural killer-derived
exosomal miR-186 inhibits neuroblastoma growth and immune escape
mechanisms. Cancer Res. 79:1151–1164. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Terrén I, Orrantia A, Vitallé J,
Zenarruzabeitia O and Borrego F: NK cell metabolism and tumor
microenvironment. Front Immunol. 10:22782019. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Labani-Motlagh A, Israelsson P, Ottander
U, Lundin E, Nagaev I, Nagaeva O, Dehlin E, Baranov V and
Mincheva-Nilsson L: Differential expression of ligands for NKG2D
and DNAM-1 receptors by epithelial ovarian cancer-derived exosomes
and its influence on NK cell cytotoxicity. Tumour Biol.
37:5455–5466. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Ishii S and Koziel MJ: Immune responses
during acute and chronic infection with hepatitis C virus. Clin
Immunol. 128:133–147. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Kaech SM and Cui W: Transcriptional
control of effector and memory CD8+ T cell differentiation. Nat Rev
Immunol. 12:749–761. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Thommen DS and Schumacher TN: T cell
dysfunction in cancer. Cancer Cell. 33:547–562. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Hiltbrunner S, Larssen P, Eldh M,
Martinez-Bravo MJ, Wagner AK, Karlsson MC and Gabrielsson S:
Exosomal cancer immunotherapy is independent of MHC molecules on
exosomes. Oncotarget. 7:38707–38717. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Torralba D, Baixauli F, Villarroya-Beltri
C, Fernández-Delgado I, Latorre-Pellicer A, Acín-Pérez R,
Martín-Cófreces NB, Jaso-Tamame ÁL, Iborra S, Jorge I, et al:
Priming of dendritic cells by DNA-containing extracellular vesicles
from activated T cells through antigen-driven contacts. Nat Commun.
9:26582018. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Raposo G, Nijman HW, Stoorvogel W,
Liejendekker R, Harding CV, Melief CJ and Geuze HJ: B lymphocytes
secrete antigen-presenting vesicles. J Exp Med. 183:1161–1172.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Yang J, Bi L, He X, Wang Z, Qian Y, Xiao L
and Shi B: Follicular helper T cell derived exosomes promote B cell
proliferation and differentiation in antibody-mediated rejection
after renal transplantation. Biomed Res Int.
2019:63879242019.PubMed/NCBI
|
|
128
|
Fernandez-Messina L, Rodriguez-Galan A, de
Yebenes VG, Gutierrez-Vazquez C, Tenreiro S, Seabra MC, Ramiro AR
and Sánchez-Madrid F: Transfer of extracellular vesicle-microRNA
controls germinal center reaction and antibody production. EMBO
Rep. 21:e489252020. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Li Y, Yang Y, Xiong A, Wu X, Xie J, Han S
and Zhao S: Comparative gene expression analysis of lymphocytes
treated with exosomes derived from ovarian cancer and ovarian
cysts. Front Immunol. 8:6072017. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Xu HY, Li N, Yao N, Xu XF, Wang HX, Liu XY
and Zhang Y: CD8+ T cells stimulated by exosomes derived from RenCa
cells mediate specific immune responses through the FasL/Fas
signaling pathway and, combined with GM-CSF and IL-12, enhance the
anti-renal cortical adenocarcinoma effect. Oncol Rep. 42:866–879.
2019.PubMed/NCBI
|
|
131
|
Taylor DD, Gerçel-Taylor C, Lyons KS,
Stanson J and Whiteside TL: T-Cell apoptosis and suppression of
T-Cell Receptor/CD3-zeta by Fas Ligand-Containing membrane vesicles
shed from ovarian tumors. Clin Cancer Res. 9:5113–5119.
2003.PubMed/NCBI
|
|
132
|
Abusamra AJ, Zhong Z, Zheng X, Li M, Ichim
TE, Chin JL and Min WP: Tumor exosomes expressing Fas ligand
mediate CD8+ T-cell apoptosis. Blood Cells Mol Dis. 35:169–173.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Filipazzi P, Burdek M, Villa A, Rivoltini
L and Huber V: Recent advances on the role of tumor exosomes in
immunosuppression and disease progression. Semin Cancer Biol.
22:342–349. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Wang X, Yao Y and Jin M: Circ-0001068 is a
novel biomarker for ovarian cancer and inducer of PD1 expression in
T cells. Aging (Albany NY). 12:19095–19106. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Asare-Werehene M, Communal L, Carmona E,
Han Y, Song YS, Burger D, Mes-Masson AM and Tsang BK: Plasma
gelsolin inhibits CD8+ T-cell function and regulates
glutathione production to confer chemoresistance in ovarian cancer.
Cancer Res. 80:3959–3971. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Taylor DD and Gercel-Taylor C:
Tumour-derived exosomes and their role in cancer-associated T-cell
signalling defects. Br J Cancer. 92:305–311. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Shenoy GN, Loyall J, Berenson CS, Kelleher
RJ Jr, Iyer V, Balu-Iyer SV, Odunsi K and Bankert RB: Sialic
Acid-Dependent inhibition of T Cells by Exosomal Ganglioside GD3 in
ovarian tumor microenvironments. J Immunol. 201:3750–3758. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Webb TJ, Li X, Giuntoli RL II, Lopez PH,
Heuser C, Schnaar RL, Tsuji M, Kurts C, Oelke M and Schneck JP:
Molecular identification of GD3 as a suppressor of the innate
immune response in ovarian cancer. Cancer Res. 72:3744–3752. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Shenoy GN, Loyall J, Maguire O, Iyer V,
Kelleher RJ Jr, Minderman H, Wallace PK, Odunsi K, Balu-Iyer SV and
Bankert RB: Exosomes Associated with human ovarian tumors harbor a
reversible checkpoint of T-cell Responses. Cancer Immunol Res.
6:236–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Clayton A, Al-Taei S, Webber J, Mason MD
and Tabi Z: Cancer exosomes express CD39 and CD73, which suppress T
cells through adenosine production. J Immunol. 187:676–683. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Kelleher RJ Jr, Balu-Iyer S, Loyall J,
Sacca AJ, Shenoy GN, Peng P, Iyer V, Fathallah AM, Berenson CS,
Wallace PK, et al: Extracellular vesicles present in human ovarian
tumor microenvironments induce a Phosphatidylserine-Dependent
arrest in the T-cell signaling cascade. Cancer Immunol Res.
3:1269–1278. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Li X and Wang X: The emerging roles and
therapeutic potential of exosomes in epithelial ovarian cancer. Mol
Cancer. 16:922017. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Wang Y, Xiang Y, Xin VW, Wang XW, Peng XC,
Liu XQ, Wang D, Li N, Cheng JT, Lyv YN, et al: Dendritic cell
biology and its role in tumor immunotherapy. J Hematol Oncol.
13:1072020. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Skokos D, Botros HG, Demeure C, Morin J,
Peronet R, Birkenmeier G, Boudaly S and Mécheri S: Mast
cell-derived exosomes induce phenotypic and functional maturation
of dendritic cells and elicit specific immune responses in vivo. J
Immunol. 170:3037–3045. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Robbins PD and Morelli AE: Regulation of
immune responses by extracellular vesicles. Nat Rev Immunol.
14:195–208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Lindenbergh MFS, Koerhuis DGJ, Borg EGF,
van't Veld EM, Driedonks TAP, Wubbolts R, Stoorvogel W and Boes M:
Bystander T-Cells Support Clonal T-Cell activation by controlling
the release of dendritic Cell-Derived Immune-Stimulatory
extracellular vesicles. Front Immunol. 10:4482019. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Zheng L, Li Z, Ling W, Zhu D, Feng Z and
Kong L: Exosomes derived from dendritic cells attenuate liver
injury by modulating the balance of Treg and Th17 Cells after
ischemia reperfusion. Cell Physiol Biochem. 46:740–756. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Li QL, Bu N, Yu YC, Hua W and Xin XY:
Exvivo experiments of human ovarian cancer ascites-derived exosomes
presented by dendritic cells derived from umbilical cord blood for
immunotherapy treatment. Clin Med Oncol. 2:461–467. 2008.PubMed/NCBI
|
|
149
|
Pitt JM, André F, Amigorena S, Soria JC,
Eggermont A, Kroemer G and Zitvogel L: Dendritic cell-derived
exosomes for cancer therapy. J Clin Invest. 126:1224–1232. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Ridge SM, Sullivan FJ and Glynn SA:
Mesenchymal stem cells: Key players in cancer progression. Mol
Cancer. 16:312017. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
De Miguel MP, Fuentes-Julián S,
Blázquez-Martínez A, Pascual CY, Aller MA, Arias J and
Arnalich-Montiel F: Immunosuppressive properties of mesenchymal
stem cells: Advances and applications. Curr Mol Med. 12:574–591.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Lis R, Touboul C, Raynaud CM, Malek JA,
Suhre K, Mirshahi M and Rafii A: Mesenchymal cell interaction with
ovarian cancer cells triggers pro-metastatic properties. PLoS One.
7:e383402012. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Zhang B, Tian X, Hao J, Xu G and Zhang W:
Mesenchymal Stem Cell-Derived extracellular vesicles in tissue
regeneration. Cell Transplant. 29:9636897209085002020. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Khare D, Or R, Resnick I, Barkatz C,
Almogi-Hazan O and Avni B: Mesenchymal stromal Cell-Derived
exosomes affect mRNA expression and function of B-Lymphocytes.
Front Immunol. 9:30532018. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Yang Y, Bucan V, Baehre H, von der Ohe J,
Otte A and Hass R: Acquisition of new tumor cell properties by
MSC-derived exosomes. Int J Oncol. 47:244–252. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Sharma S, Alharbi M, Kobayashi M, Lai A,
Guanzon D, Zuniga F, Ormazabal V, Palma C, Scholz-Romero K, Rice
GE, et al: Proteomic analysis of exosomes reveals an association
between cell invasiveness and exosomal bioactivity on endothelial
and mesenchymal cell migration in vitro. Clin Sci (Lond).
132:2029–2044. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Dean M, Fojo T and Bates S: Tumour stem
cells and drug resistance. Nat Rev Cancer. 5:275–284. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Vera N, Acuna-Gallardo S, Grunenwald F,
Caceres-Verschae A, Realini O, Acuna R, Lladser A, Illanes SE and
Varas-Godoy M: Small extracellular vesicles released from ovarian
cancer spheroids in response to Cisplatin Promote the
Pro-Tumorigenic activity of mesenchymal stem cells. Int J Mol Sci.
20:49722019. View Article : Google Scholar
|
|
159
|
Qiu L, Wang J, Chen M, Chen F and Tu W:
Exosomal microRNA-146a derived from mesenchymal stem cells
increases the sensitivity of ovarian cancer cells to docetaxel and
taxane via a LAMC2-mediated PI3K/Akt axis. Int J Mol Med.
46:609–620. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Reza AMMT, Choi YJ, Yasuda H and Kim JH:
Human adipose mesenchymal stem cell-derived exosomal-miRNAs are
critical factors for inducing anti-proliferation signalling to
A2780 and SKOV-3 ovarian cancer cells. Sci Rep. 6:384982016.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Melzer C, Rehn V, Yang Y, Bahre H, von der
Ohe J and Hass R: Taxol-Loaded MSC-Derived exosomes provide a
therapeutic vehicle to target metastatic breast cancer and other
carcinoma cells. Cancers (Basel). 11:7982019. View Article : Google Scholar
|
|
162
|
Tanaka A and Sakaguchi S: Regulatory T
cells in cancer immunotherapy. Cell Res. 27:109–118. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Curiel TJ, Coukos G, Zou L, Alvarez X,
Cheng P, Mottram P, Evdemon-Hogan M, Conejo-Garcia JR, Zhang L,
Burow M, et al: Specific recruitment of regulatory T cells in
ovarian carcinoma fosters immune privilege and predicts reduced
survival. Nat Med. 10:942–949. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
164
|
Szajnik M, Czystowska M, Szczepanski MJ,
Mandapathil M and Whiteside TL: Tumor-derived microvesicles induce,
expand and up-regulate biological activities of human regulatory T
cells (Treg). PLoS One. 5:e114692010. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Gabrilovich DI: Myeloid-Derived suppressor
cells. Cancer Immunol Res. 5:3–8. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Chalmin F, Ladoire S, Mignot G, Vincent J,
Bruchard M, Remy-Martin JP, Boireau W, Rouleau A, Simon B, Lanneau
D, et al: Membrane-associated Hsp72 from tumor-derived exosomes
mediates STAT3-dependent immunosuppressive function of mouse and
human myeloid-derived suppressor cells. J Clin Invest. 120:457–471.
2010.PubMed/NCBI
|
|
167
|
Balaburski GM, Leu JI, Beeharry N, Hayik
S, Andrake MD, Zhang G, Herlyn M, Villanueva J, Dunbrack RL Jr, Yen
T, et al: A modified HSP70 inhibitor shows broad activity as an
anticancer agent. Mol Cancer Res. 11:219–229. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Gobbo J, Marcion G, Cordonnier M, Dias
AMM, Pernet N, Hammann A, Richaud S, Mjahed H, Isambert N, Clausse
V, et al: Restoring anticancer immune response by targeting
Tumor-Derived exosomes with a HSP70 Peptide Aptamer. J Natl Cancer
Inst. 108:2015.(Epub ahead of print). PubMed/NCBI
|
|
169
|
Liu T, Han C, Wang S, Fang P, Ma Z, Xu L
and Yin R: Cancer-associated fibroblasts: An emerging target of
anti-cancer immunotherapy. J Hematol Oncol. 12:862019. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Cho JA, Park H, Lim EH, Kim KH, Choi JS,
Lee JH, Shin JW and Lee KW: Exosomes from ovarian cancer cells
induce adipose tissue-derived mesenchymal stem cells to acquire the
physical and functional characteristics of tumor-supporting
myofibroblasts. Gynecol Oncol. 123:379–386. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Yeung TL, Leung CS, Wong KK, Samimi G,
Thompson MS, Liu J, Zaid TM, Ghosh S, Birrer MJ and Mok SC: TGF-β
modulates ovarian cancer invasion by upregulating CAF-derived
versican in the tumor microenvironment. Cancer Res. 73:5016–5028.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Li W, Zhang X, Wang J, Li M, Cao C, Tan J,
Ma D and Gao Q: TGFβ1 in fibroblasts-derived exosomes promotes
epithelial-mesenchymal transition of ovarian cancer cells.
Oncotarget. 8:96035–96047. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Lu TX and Rothenberg ME: MicroRNA. J
Allergy Clin Immunol. 141:1202–1207. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Zhang J, Li S, Li L, Li M, Guo C, Yao J
and Mi S: Exosome and exosomal microRNA: Trafficking, sorting, and
function. Genomics Proteomics Bioinformatics. 13:17–24. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Au Yeung CL, Co NN, Tsuruga T, Yeung TL,
Kwan SY, Leung CS, Li Y, Lu ES, Kwan K, Wong KK, et al: Exosomal
transfer of stroma-derived miR21 confers paclitaxel resistance in
ovarian cancer cells through targeting APAF1. Nat Commun.
7:111502016. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Dickman CT, Lawson J, Jabalee J, MacLellan
SA, LePard NE, Bennewith KL and Garnis C: Selective extracellular
vesicle exclusion of miR-142-3p by oral cancer cells promotes both
internal and extracellular malignant phenotypes. Oncotarget.
8:15252–15266. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Kanlikilicer P, Rashed MH, Bayraktar R,
Mitra R, Ivan C, Aslan B, Zhang X, Filant J, Silva AM,
Rodriguez-Aguayo C, et al: Ubiquitous release of exosomal tumor
suppressor miR-6126 from ovarian cancer cells. Cancer Res.
76:7194–7207. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Rashed MH, Kanlikilicer P,
Rodriguez-Aguayo C, Pichler M, Bayraktar R, Bayraktar E, Ivan C,
Filant J, Silva A, Aslan B, et al: Exosomal miR-940 maintains
SRC-mediated oncogenic activity in cancer cells: A possible role
for exosomal disposal of tumor suppressor miRNAs. Oncotarget.
8:20145–20164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Masoumi-Dehghi S, Babashah S and
Sadeghizadeh M: MicroRNA-141-3p-containing small extracellular
vesicles derived from epithelial ovarian cancer cells promote
endothelial cell angiogenesis through activating the JAK/STAT3 and
NF-κB signaling pathways. J Cell Commun Signal. 14:233–244. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Yoshimura A, Sawada K, Nakamura K, Kinose
Y, Nakatsuka E, Kobayashi M, Miyamoto M, Ishida K, Matsumoto Y,
Kodama M, et al: Exosomal miR-99a-5p is elevated in sera of ovarian
cancer patients and promotes cancer cell invasion by increasing
fibronectin and vitronectin expression in neighboring peritoneal
mesothelial cells. BMC Cancer. 18:10652018. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Kobayashi M, Salomon C, Tapia J, Illanes
SE, Mitchell MD and Rice GE: Ovarian cancer cell invasiveness is
associated with discordant exosomal sequestration of Let-7 miRNA
and miR-200. J Transl Med. 12:42014. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Pan C, Stevic I, Muller V, Ni Q,
Oliveira-Ferrer L, Pantel K and Schwarzenbach H: Exosomal microRNAs
as tumor markers in epithelial ovarian cancer. Mol Oncol.
12:1935–1948. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Kobayashi M, Sawada K, Nakamura K,
Yoshimura A, Miyamoto M, Shimizu A, Ishida K, Nakatsuka E, Kodama
M, Hashimoto K, et al: Exosomal miR-1290 is a potential biomarker
of high-grade serous ovarian carcinoma and can discriminate
patients from those with malignancies of other histological types.
J Ovarian Res. 11:812018. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Cheng L, Zhang K, Qing Y, Li D, Cui M, Jin
P and Xu T: Proteomic and lipidomic analysis of exosomes derived
from ovarian cancer cells and ovarian surface epithelial cells. J
Ovarian Res. 13:92020. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Nakamura K, Sawada K, Kinose Y, Yoshimura
A, Toda A, Nakatsuka E, Hashimoto K, Mabuchi S, Morishige KI,
Kurachi H, et al: Exosomes promote ovarian cancer cell invasion
through transfer of CD44 to peritoneal mesothelial cells. Mol
Cancer Res. 15:78–92. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Dorayappan KDP, Wallbillich JJ, Cohn DE
and Selvendiran K: The biological significance and clinical
applications of exosomes in ovarian cancer. Gynecol Oncol.
142:199–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
187
|
Enriquez VA, Cleys ER, Da Silveira JC,
Spillman MA, Winger QA and Bouma GJ: High LIN28A expressing ovarian
cancer cells secrete exosomes that induce invasion and migration in
HEK293 cells. Biomed Res Int. 2015:7013902015. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Stope MB, Klinkmann G, Diesing K, Koensgen
D, Burchardt M and Mustea A: Heat Shock Protein HSP27 secretion by
ovarian cancer cells is linked to intracellular expression levels,
occurs independently of the endoplasmic reticulum pathway and
HSP27's phosphorylation status, and is mediated by exosome
liberation. Dis Markers. 2017:15753742017. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Nam GH, Choi Y, Kim GB, Kim S, Kim SA and
Kim IS: Emerging prospects of exosomes for cancer treatment: From
conventional therapy to immunotherapy. Adv Mater. 32:e20024402020.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Koyama Y, Ito T, Hasegawa A, Eriguchi M,
Inaba T, Ushigusa T and Sugiura K: Exosomes derived from tumor
cells genetically modified to express Mycobacterium tuberculosis
antigen: A novel vaccine for cancer therapy. Biotechnol Lett.
38:1857–1866. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Kalimuthu S, Gangadaran P, Rajendran RL,
Zhu L, Oh JM, Lee HW, Gopal A, Baek SH, Jeong SY, Lee SW, et al: A
new approach for loading anticancer drugs into mesenchymal stem
Cell-Derived exosome mimetics for cancer therapy. Front Pharmacol.
9:11162018. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Feng Y, Hang W, Sang Z, Li S, Xu W, Miao
Y, Xi X and Huang Q: Identification of exosomal and non-exosomal
microRNAs associated with the drug resistance of ovarian cancer.
Mol Med Rep. 19:3376–3392. 2019.PubMed/NCBI
|
|
193
|
Zhu X, Shen H, Yin X, Yang M, Wei H, Chen
Q, Feng F, Liu Y, Xu W and Li Y: Macrophages derived exosomes
deliver miR-223 to epithelial ovarian cancer cells to elicit a
chemoresistant phenotype. J Exp Clin Cancer Res. 38:812019.
View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Kanlikilicer P, Bayraktar R, Denizli M,
Rashed MH, Ivan C, Aslan B, Mitra R, Karagoz K, Bayraktar E, Zhang
X, et al: Exosomal miRNA confers chemo resistance via targeting
Cav1/p-gp/M2-type macrophage axis in ovarian cancer. EBioMedicine.
38:100–112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Guo H, Ha C, Dong H, Yang Z, Ma Y and Ding
Y: Cancer-associated fibroblast-derived exosomal microRNA-98-5p
promotes cisplatin resistance in ovarian cancer by targeting
CDKN1A. Cancer Cell Int. 19:3472019. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Cao YL, Zhuang T, Xing BH, Li N and Li Q:
Exosomal DNMT1 mediates cisplatin resistance in ovarian cancer.
Cell Biochem Funct. 35:296–303. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Kim SM, Yang Y, Oh SJ, Hong Y, Seo M and
Jang M: Cancer-derived exosomes as a delivery platform of
CRISPR/Cas9 confer cancer cell tropism-dependent targeting. J
Control Release. 266:8–16. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Tsou P, Katayama H, Ostrin EJ and Hanash
SM: The emerging role of B Cells in tumor immunity. Cancer Res.
76:5597–5601. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Im EJ, Lee CH, Moon PG, Rangaswamy GG, Lee
B, Lee JM, Lee JC, Jee JG, Bae JS, Kwon TK, et al: Sulfisoxazole
inhibits the secretion of small extracellular vesicles by targeting
the endothelin receptor A. Nat Commun. 10:13872019. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Walker S, Busatto S, Pham A, Tian M, Suh
A, Carson K, Quintero A, Lafrence M, Malik H, Santana MX and
Wolfram J: Extracellular vesicle-based drug delivery systems for
cancer treatment. Theranostics. 9:8001–8017. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Pisano S, Pierini I, Gu J, Gazze A,
Francis LW, Gonzalez D, Conlan RS and Corradetti B: Immune (Cell)
Derived Exosome Mimetics (IDEM) as a treatment for ovarian cancer.
Front Cell Dev Biol. 8:5535762020. View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Ge L, Zhang N, Li D, Wu Y, Wang H and Wang
J: Circulating exosomal small RNAs are promising non-invasive
diagnostic biomarkers for gastric cancer. J Cell Mol Med.
24:14502–14513. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Chen Z, Liang Q, Zeng H, Zhao Q, Guo Z,
Zhong R, Xie M, Cai X, Su J, He Z, et al: Exosomal CA125 as a
promising biomarker for ovarian cancer diagnosis. J Cancer.
11:6445–6453. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Wang Y, Li Q, Shi H, Tang K, Qiao L, Yu G,
Ding C and Yu S: Microfluidic Raman biochip detection of exosomes:
A promising tool for prostate cancer diagnosis. Lab Chip.
20:4632–4637. 2020. View Article : Google Scholar : PubMed/NCBI
|