Breast cancer is a malignant tumor that occurs in
the lobule and the ductal epithelium of the breast (1). It ranks first in terms of incidence
among malignancies in female patients worldwide (2). In 2005, ~1 million new cases of breast
cancer were reported worldwide and cases were growing at a rate of
5–20% per year (3). With the
development of medical technology and the increasing awareness of
cancer prevention, the mortality rate of breast cancer is
decreasing every year. In 2019, ~268,600 new cases of invasive
breast cancer and 41,760 deaths associated with breast cancer were
reported in USA women (2). Numerous
studies have focused on the pathogenesis, progression, prognosis
and treatment of breast cancer. Notably, increasing attention has
been paid to the role of metabolic oxidase in breast cancer.
Cytochrome P450 (CYP) enzymes are a large family of
self-oxidizing heme proteins belonging to the class of
mono-oxygenases, and these compounds are involved in the synthesis
of hormones, second messengers and other endogenous substances in
the body, and in the regulation of detoxification and substance
metabolism (4). Under normal
conditions, CYPs are widely expressed in multiple human organs and
are mainly located in the liver (5).
During the course of the disease, such as breast cancer, CYPs are
selectively expressed in different types of neoplasms (6). Notably, this phenomenon demonstrates
the biological importance of exploring tumor-inherent metabolic
pathways. Therefore, the present review focuses on the effects of
the CYP metabolic pathway and its gene polymorphism on the
occurrence, progression, treatment and prognosis of breast
cancer.
CYPs are involved in the biosynthesis and oxidative
metabolism of sex hormones. Estradiol is gradually synthesized from
cholesterol by the catalytic action of enzymes, such as CYP11,
CYP17 and CYP19 family (29).
Subsequently, estradiol is converted into various hydroxyl products
by enzymatic oxidation, and the hydroxyl products undergo
glucuronidation, sulfation, esterification and O-methylation
metabolism, leading to the production of carcinogens (30). CYP1B1 is highly expressed in breast
carcinoma tissues, and this compound metabolizes estrogen to
4-hydroxyestradiol and simultaneously binds to and activates ER,
thereby promoting cell mitosis in breast tissues (29). This excessive growth stimulation may
promote breast cancer occurrence (31). In addition, reactive estrogen
semiquinone/quinone intermediates are the metabolic redox products
of 4-hydroxyestradiol (32), causing
DNA destruction, induction of cell transformation and initiation of
tumorigenesis (33,34). Bradlow et al (35) demonstrated that the metabolite
16α-hydroxyestradiol of CYP2C9 and CYP3A4 is positively associated
with the incidence of breast cancer. Furthermore, based on an
epidemiological case-control study (36) and prospective study (37), elevated levels of
16α-hydroxyestradiol are associated with increased risk of breast
cancer. Conversely, ERs also regulate CYP expression in breast
tumors (38). CYP2B6 expression is
markedly increased in ER-positive breast tumors because ERα
regulates CYP2B6 gene expression in human breast cancer cells by
directly binding to functional estrogen response elements located
in the upstream regulatory region of CYP2B6 (38). Notably, Fukasawa et al
(39) revealed a novel compound,
NK150460, which inhibits 17β-estradiol (E2)-dependent transcription
without affecting binding of E2 to ER. Contrary to expectations,
NK150460 inhibits the proliferation of not only most ER-positive,
but also some ER-negative breast cancer cell lines, such as T-47D,
MCF-7, and SK-BR-3; however, it never inhibits the proliferation of
non-breast cancer cell lines (39).
At present, inhibiting the synthesis of estrogen is still an
important regimen for the treatment of luminal-type breast cancer.
Although estrogen is considered to serve a causal role in breast
cancer, as aforementioned, there is increasing evidence that the
way estrogen is metabolized is related to the risk of breast
cancer. This may explain why the effect of endocrine therapy is not
always satisfactory. Therefore, studies clarifying the mechanism of
estrogen metabolism in patients with breast cancer are an exciting
domain of investigation. This could provide patients with precise
endocrine targeted therapy in the future.
In addition to participating in estrogen metabolism,
the gene polymorphisms of CYPs carry huge implications for the risk
and prognosis of breast cancer. CYP3A4 mRNA expression is
negatively associated with the morbidity of breast cancer (40). Additionally, Johnson et al
(41) reported that the CYP3A
polymorphism site rs10235235 is negatively associated with the
morbidity of patients with breast carcinoma with late menarche.
Johnson et al (42) further
revealed that CYP3A4 SNP (rs10273424), a non-coding variant at the
CYP3A locus, is associated with reduced risk of breast cancer in
younger women below the age of 50 years at the time of diagnosis.
However, polymorphisms in the CYP17, CYP19 and CYP1A1 genes are
closely associated with breast cancer susceptibility (43–45).
Women with CYP1A1 T6235C and A4889G genotypes rather than AA and TT
are more likely to develop low-grade tumors; 85.9% of tumors in the
AA and TT genotype groups are grade III; however, only 76.1% of
polymorphism carriers are grade III (46). There was no significant difference in
survival related to CYP1A1 gene status (46). Sangrajrang et al (29) performed genetic testing on the breast
tissue of 1,067 Thai women and revealed that CYP1A2, CYP2C19 and
CYP17 polymorphisms serve a crucial role in estrogen metabolism and
affect the individual susceptibility of Thai women to breast
cancer. Furthermore, CYP1A2 rs2470890 is prominently associated
with the prognosis of patients with breast cancer and might serve
as a novel genetic indicator of prognosis of patients with breast
carcinoma (47). Raskin et al
(48) demonstrated that CYP19 Val
(80) polymorphisms and its
haplotypes are associated with increased risk of breast cancer in
young women with breast cancer susceptibility gene mutations. In
addition, the association between six SNPs of CYP8A1 and breast
cancer risk has been reported in detail (49,50).
Patients with homozygotes of minor alleles of rs5602, rs477627 and
rs6125671 exhibit increased risk of breast cancer compared with
normal alleles (49). Furthermore,
the minor allele homozygotes of rs477627 have a protective effect
in Caucasian populations (49).
Among Caucasian women with progesterone receptor-positive breast
cancer, the cancer risk is associated with the rs6095541 and
rs6095543 alleles (50). Notably,
genetic polymorphisms in CYPs vary among different ethnicities
(49). Justenhoven et al
(51) reported that the CYP2C19*17
variant is associated with reduced risk of breast cancer in the
German population; however, this variation is common in Europeans
but rare in Asians. Furthermore, Ruiter et al (52) suggested that CYP2C19*2 polymorphisms
are associated with increased survival time in European patients
with breast cancer who have been treated with tamoxifen. Among
female Chinese Han patients, the carriers of the A allele of
CYP2C19*3 are 2.19 times more likely to be attacked by breast
cancer than those of the G allele (53). However, this variant of CYP2C19*3 is
rare among the European population (53). The relationship between the gene
polymorphisms of CYPs and breast cancer has been clarified, thus
contributing to the prediction of populations with high-risk tumor
and determine individuals susceptible to tumors. However, at
present, numerous conflicting results have been reported, and this
may be associated with various reasons, such as region, ethnicity
and sex. To the best of our knowledge, limited information is
available regarding the interaction between the CYP gene and other
genes or environmental factors. Therefore, an in-depth study on the
association between CYP gene polymorphisms and tumorigenesis is
important for early tumor screening, targeted therapy and
therapeutic efficacy.
Angiogenesis is one of the major characteristics of
malignant tumors, which are rich in blood supply. Angiogenesis is
regulated by pro-angiogenic and anti-angiogenic factors via a
complicated process. Under normal conditions, the dynamic balance
is maintained between pro-angiogenic and anti-angiogenic factors
during physiological angiogenesis. When this balance is broken by
pathogenic factors, pathological angiogenesis, such as tumor
angiogenesis, might occur. Breast cancer cells flourish in the
tumor microenvironment. Diverse components of the breast cancer
microenvironment, such as pathological neovascular structures, may
promote breast cancer progression and metastasis (54). The CYP4 family can hydroxylate
arachidonic acid (AA), and the product of this metabolism is a
novel lipid mediator, which may be involved in the proliferation of
breast cancer cells and tumor angiogenesis (55). 20-Hydroxy-eicosatetraenoic acid
(20-HETE) is converted from AA by the ω-hydroxylase enzymes from
the CYP family 4 and subfamily A (CYP4A) genes (55). 20-HETE is the main pro-inflammatory
metabolite that regulates vascular remodeling and
neovascularization under ischemic or hypoxic conditions (55–57).
Additionally, 20-HETE serves an important role in epidermal growth
factor and vascular endothelial growth factor (VEGF) activation,
pro-angiogenic effects and the stimulation of endothelial cell
proliferation, migration and cell survival (55,58). In
tumors, the CYP4A/20-HETE axis promotes endothelial cell migration
and neovascularization (59,60). When
N-hydroxy-N′-(4-butyl-2methylphenyl) formamidine(HET0016), a highly
selective inhibitor of 20-HETE synthesis, is used alone in
tumor-bearing animals, tumor neovascularization is reduced
(61,62). When the level of different pro- and
anti-angiogenic factors in tumor lysates is detected, prominent
changes occur after HET0016 treatment compared with placebo-treated
tumors (63,64). When certain indicators, including
extravascular cell space (EES), vascular parameters and tumor
angiogenesis, are analyzed, HET0016 treatment reduces EES, tumor
blood volume, permeability and tumor angiogenesis (63,64). In
the field of cancer, triple-negative breast cancer is not sensitive
to chemotherapeutics. The inhibition of the synthesis of 20-HETE is
expected to be a breakthrough in the treatment of triple-negative
breast cancer. Further investigation is required to explore how
20-HETE inhibitors could be more effective in decreasing tumor
growth and neovascularization.
In addition, compared with other members of the
large CYP family 4, CYP4Z1 is unique and vital in the development
of breast cancer. A 52% increase in CYP4Z1 mRNA expression was
identified in breast cancer tissues compared with non-cancerous
tissues (65). Yu et al
(66) demonstrated that CYP4Z1
overexpression activates the PI3K/Akt and ERK1/2 signaling pathways
and induces human breast cancer angiogenesis and tumor growth.
Additionally, Wang et al (67) demonstrated that CYP4Z1 3′untranslated
region may be involved in the regulation of E-cadherin protein,
thus affecting the migration of breast cancer cells. In addition,
in ER-positive breast cancer cells, the regulation of the CYP4Z1
RNA network may be associated with tamoxifen resistance during
breast cancer treatment (68).
Further in vivo experiments are required to confirm whether
CYP4Z1 could be a therapeutic target in restraining cancer
progression.
The tumor interstitial microenvironment serves an
essential role in the occurrence, development and metastasis of
breast cancer. The breast carcinoma microenvironment has numerous
components, including suppressive immune cells, reprogrammed
fibroblasts, pathological neovascular structures, altered
extracellular matrix and certain soluble factors, which together
facilitate a pro-tumorigenic environment (54). Tumor-associated macrophages (TAMs)
are the main component of the tumor microenvironment and are among
the most abundant inflammatory stromal cells (69). According to different functional
characteristics, TAMs are usually divided into the M1 and M2
subtype. The M1 type is equipped with antitumor effects, while M2
activates the generation of tumor growth factors, such as VEGF, and
promotes tumor growth, invasion and metastasis (70–72).
Furthermore, the role of TAMs that construct a protective niche of
tumor cells in distant organs is slowly being recognized (73). Notably, targeting TAMs in part
prevents tumor metastasis by inhibiting the production of multiple
endogenous factors, including chemokines, inflammatory factors and
growth factors (74–76). CYP4A, a key inducible cytochrome P450
enzyme, catalyzes the synthesis of 20-HETE from AA in human tissues
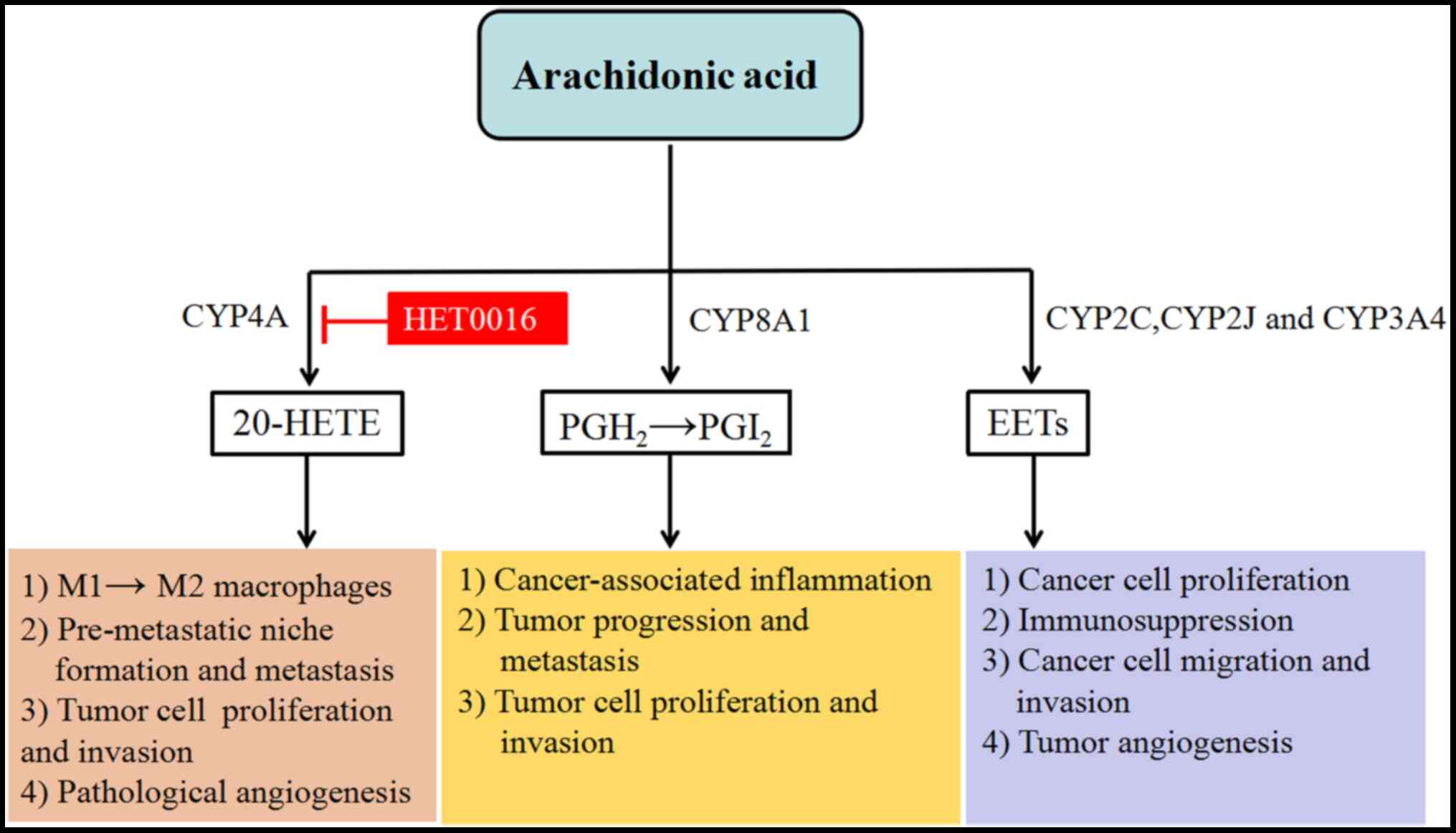
(77), as shown in Fig. 1. 20-HETE is a novel lipid mediator,
which promotes tumor growth and angiogenesis (78) and induces the transformation of the
tumor stromal microenvironment into the pro-tumorigenic environment
(79,80). The expression levels of
20-HETE-producing enzymes of the CYP4A family are upregulated in
invasive breast carcinoma tissues (73). CYP4A/20-HETE induces the
transformation of macrophage to M2 phenotype via the STAT3
signaling pathway, and CYP4A/20-HETE activation is an important
medium for promoting niche formation before tumor cell metastasis,
leading to distant colonization of tumor cells (73). Therefore, downregulation of
CYP4A/20-HETE expression has therapeutic potential in the tumor
microenvironment.
In human tissues, CYP4A11 and CYP4A22 are the most
important isozyme types of CYP4A for catalyzing the synthesis of
20-HETE (80). Alexanian et
al (81) revealed that the
expression levels of CYP4A11 and CYP4A22 in triple negative breast
cancer tissues are higher than those in normal breast tissues,
indicating an increase in CYP4A/20-HETE activity after the
malignant transformation of breast ductal epithelium. The
downregulation of cyclooxygenase-2 and CYP4A signaling in human
breast carcinoma cells inhibits cancer metastasis by preventing
anoikis resistance, migration and invasion (82). The inhibition of 20-HETE synthesis
with chemical inhibitor HET0016 reduces proangiogenic factors and
inhibits breast cancer growth (62).
Borin et al (61)
demonstrated that targeting the AA signaling pathway by inhibiting
the synthesis of 20-HETE reduces the migration and invasion of
metastatic breast cancer cells. Furthermore, an animal study
indicated that the incidence of lung metastases in breast cancer
can be reduced by inhibiting CYP4A/20-HETE activity (61). Notably, CYP8A1 also is involved in
the metabolism of AA. CYP8A1, also known as prostacyclin I2
synthase, is an isomerase, which converts prostaglandin H2 into
prostacyclin (PGI2) (83). PGI2 is a
type of prostanoid, which exerts a crucial role in
cancer-associated inflammation, tumor progression and metastasis
(84,85). Notably, CYP8A1 is expressed in breast
carcinoma and adjacent tissues and is involved in the inflammation
mechanism of breast cancer cells through the aforementioned
eicosanoid metabolites (83). In
addition, CYP8A1 affects tumor cell survival signaling via AA
metabolism pathways, including cell proliferation and apoptosis,
tumor cell invasion, metastasis, and angiogenesis (83). Similarly, CYP2C19 is a key enzyme for
the synthesis of epoxy-eicosatrienoic acids (EETs) in the AA
metabolic pathway (86). The
exogenous supplementation of EETs promotes the proliferation of
cancer cells in vitro and in vivo (86). In a number of tumor cell lines, such
as breast cancer cell lines, EETs stimulate the activation of the
MAPK and PI3K/Akt signaling pathways, promote phosphorylation of
EGFR (86), alter the tumor
microenvironment, and induce immunosuppression in an autocrine and
paracrine manner (80).
Additionally, EET inhibits cancer cell apoptosis by upregulating
the anti-apoptotic proteins Bcl-2 and Bcl-xl and downregulating the
pro-apoptotic protein Bax (86).
Therefore, CYP2C19 serves a vital role in promoting the malignant
manifestation of tumors and in the pathogenesis of breast cancer
via the EET anabolic pathway. In humans, CYP2C8, 2C9 and 2J2
subfamily members are also involved in the synthesis of EETs.
CYP2C8, 2C9 and 2J2 are highly expressed in breast cancer tissues,
thus indirectly promoting cancer cell proliferation, migration,
angiogenesis and invasion (87).
CYP3A4, an activated AA epoxygenase, accelerates STAT3-mediated
cell proliferation of breast cancer via EET biosynthesis (88). Furthermore, EETs induce tumor cell
proliferation and survival via multiple signaling pathways and
molecular mechanisms, including EGFR/PI3K/Akt, EGFR/MAPK, TNF-α and
prometastatic matrix metalloproteinases (86,89,90).
Therefore, novel therapeutics that target the AA metabolic pathway
should be the focus for cancer chemoprevention and treatments.
CYPs are selectively expressed in breast cancer
tissues, and their expression levels affect clinicopathological
factors and patient prognosis (91).
Based on the analysis of the expression levels of CYPs in 393
patients with breast cancer, Haas et al (92) revealed that the expression levels of
CYP3A4/5 are markedly associated with lymph node metastasis rate,
and high expression levels of CYP1B1 are associated with poor tumor
differentiation. Murray et al (91) demonstrated that CYP2S1, CYP3A4,
CYP4V2 and CYP26A1 are associated with survival in patients with
breast cancer, indicating that CYP can be used as a marker of
prognosis for patients with breast cancer. CYP4Z1 is a novel member
of the CYP4 family that is upregulated in human breast cancer, and
CYP4Z1 expression is positively associated with high-grade
malignancy tumors and poor prognosis (66). Patients with breast cancer with high
CYP4A22 expression often experience shortened relapse-free
survival, which is a negative prognostic factor (73). The downregulation of CYP4A signal
transduction may inhibit tumor cell migration and invasion, thereby
inhibiting breast cancer metastasis (82). However, the expression of several
CYPs may imply favorable clinical outcomes (93,94).
CYP2E1 expression is elevated in breast cancer tissues and
negatively associated with tumor size (93). High CYP2E1 expression contributes to
the production of reactive oxygen species and the occurrence of the
oxidative stress reaction in breast cancer cells, resulting in
damaged mitochondria and DNA modification, leading to accelerated
death of necrotic cancer cells, which may be beneficial to patients
with breast cancer to a certain extent (93,94).
CYP2E1 could inhibit breast cancer cell metastasis by regulating
tumor cell autophagy and stimulating endoplasmic reticulum stress
(95). Other mechanisms involved in
the adverse prognostic factors mediated by CYPs in patients with
breast carcinoma need to be explored further. However, the present
review suggests that the inhibition of CYPs may be a novel
therapeutic target for improving the prognosis of patients with
breast cancer.
Inter-individual genetic variation in the metabolism
of carcinogens is a determinant factor of susceptibility to various
types of cancer (96). CYPs serve a
profound role in the metabolism of carcinogens (97). The difference in the enzyme activity
of CYPs determines the susceptibility to chemical carcinogens
(96). Numerous CYPs are involved in
catalyzing the metabolism of potential breast cancer-related
carcinogens (96). Polycyclic
aromatic hydrocarbons (PAHs) are common environmental carcinogens
that induce tumorigenesis when they are activated by CYPs (98). They accumulate in breast tissues
(99), and are metabolized and
activated by CYP1A1 to produce electrophilic epoxy compounds with
strong carcinogenic activity, which change the base pairing of DNA,
cause codon changes, introduce mutations and eventually lead to
cancer (100). Therefore, the
activity of CYP1A1 isoenzyme in breast tissues is the key to
determine the carcinogenicity of PAHs. The aryl hydrocarbon
receptor (AhR) is a transcriptional regulator of CYP1A1, thereby
regulating its protein expression (101). To the best of our knowledge,
Al-Dhfyan et al (101) were
the first to report that the AhR/CYP1A1 signaling pathway regulates
the proliferation, development, self-renewal and chemoresistance of
breast cancer stem cells by inhibiting phosphatase and tensin
homolog and activating β-catenin and Akt signaling pathways. AhR
may be a potential drug target for treatment of ER-negative breast
cancer in the future. In addition, CYP1B1 can convert the
heterocyclic amine 2-amino-1-methyl-6-phenylimidazole[4,5-b]
pyridine in food into N2-hydroxylated derivatives with DNA
mutagenicity, which has been linked to the incidence of colon
cancer and breast cancer (102).
Additionally, CYP2W1 expression is upregulated in breast cancer
tissues and is involved in the biological activation of PAH
carcinogens (83). CYPs have
hallmark effects in the metabolic activation and elimination of
numerous carcinogens. A number of chemical carcinogens are mostly
indirect carcinogens, which need to be activated by metabolic
activation in vivo to interact with cellular biomolecules to
cause cancer. The genetic polymorphism of CYPs determines the
discrepancy in the metabolism of carcinogens and the susceptibility
of tumors in patients (96). In
future research, attention should be paid to the metabolic function
and genotypes of CYPs in patients to carry out individualized
administration for patients with tumors and optimize the clinical
therapeutic schedule.
The genetic polymorphisms of CYP enzymes in breast
tumors have an effect on the drug treatment outcomes of patients
with breast cancer. They lead to changes in the response to drugs
ranging from adverse reactions to lack of efficacy. The discovery
of genetic markers for susceptibility to breast carcinoma has led
to a growing body of epidemiological research examining relatively
common genetic polymorphisms. Among the 57 identified CYP
isoenzymes that catalyze drug metabolism, >20 CYP genes,
including CYP1B1, CYP2B6, CYP2C9, CYP2C19 and CYP2D6, have
functional polymorphisms. CYP1B1 is a metabolic enzyme for numerous
anticancer drugs, including cyclophosphamide, paclitaxel,
doxorubicin, docetaxel, cisplatin and 5-fluorouracil (103). Furthermore, the CYP1B1 4326G allele
is associated with a decreased response rate, reduced
progression-free survival and shorter overall survival in patients
with breast cancer treated with taxanes (104). CYP2B6 serves a pivotal role in the
efficacy of doxorubicin and cyclophosphamide (105). Compared with that in patients with
wild-type alleles, the efficacy of neoadjuvant chemotherapy is
reduced in patients with breast cancer with CYP2C9*2 heterozygotes
(106). Furthermore, the genotypes
of CYP2C19 and CYP2D6 influence the therapeutic actions of
tamoxifen in patients with breast cancer (107). Similarly, the majority of CYP genes
are associated with the clinical efficacy of chemotherapy drugs in
patients with breast cancer; these genes include CYP1A2, CYP2A6,
CYP2B, CYP2C8, CYP2C9, CYP2E1, CYP2S1, CYP2W1, CYP3A4 and CYP3A5
(97,106,108–110).
Furthermore, the genetic polymorphisms of CYPs are closely
associated with the hematological adverse reactions caused by
chemotherapy drugs in patients with breast cancer (105). Bray et al (105) revealed that CYP2B6*2 and CYP2B6*5
mutant genes are closely associated with a high incidence of drug
dose delay and adverse hematological reactions. Nakajima et
al (111) demonstrated that
leukopenia is associated with CYP2B6 gene polymorphism g.
−2320T>C, g. −750T>C, g. 18492T>C. Therefore, this may
explain the unsatisfactory therapeutic effect in some patients with
breast cancer. Whole genome sequencing of CYPs in patients with
breast cancer is important to provide the necessary guarantee to
patients to implement precise treatment. Although next-generation
sequencing technology is relatively mature now, its high cost
limits the actions of patients. In the future, more patients are
expected to benefit from it.
CYPs serve a multi-faceted role in contributing to
carcinogenesis, tumor growth, invasion and metastasis.
Additionally, they catalyze phase 1 metabolism of xenobiotics, such
as drugs and carcinogens. These CYP gene polymorphisms are
associated with drug responses and susceptibility to breast cancer.
Notably, the CYP4A/20-HETE axis serves a key role in promoting
tumor growth, neovascularization, migration and invasion, and may
be a potential target for prevention and therapy of metastasis.
Evidence that glioma and breast tumor growth and metastasis can be
successfully controlled by HET0016 has been provided by some
research groups. Therefore, novel therapeutics targeting the
CYP4A/20-HETE metabolic pathway should deserve more attention in
translational medicine, either as monotherapy or in combination
with first-line chemotherapy drugs and radiotherapeutic approaches.
An increased understanding of CYPs will provide novel therapeutic
targets for clinical precision treatment of patients with breast
cancer.
Not applicable.
This project was funded by Science and Technology
Planning Project of Wuhan (grant no. 2017060201010172) and Guidance
Foundation of Renmin Hospital of Wuhan University (grant no.
RMYD2018M27).
Data sharing is not applicable to this article, as
no datasets were generated or analyzed during the current
study.
JY designed and supervised the study. BL drafted the
manuscript and prepared the figures. HY performed the literature
analysis. DY revised the manuscript. Data authentication is not
applicable. All authors have read and approved the manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Lakhani SR, Ellis IO, Schnitt SJ, Pan PH
and van de Vijver MJ: WHO Classifcation of Tumours of the Breast.
WHO Classifcation of Tumours. 4th edition. 4. IARC Press; Lyon: pp.
82–134. 2012
|
|
2
|
Desantis CE, Ma J, Gaudet MM, Newman LA,
Miller KD, Goding SA, Jemal A and Siegel RL: Breast cancer
statistics, 2019. CA Cancer J Clin. 69:438–451. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dumitrescu RG and Cotarla I: Understanding
breast cancer risk-where do we stand in 2005? J Cell Mol Med.
9:208–221. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gonzalez FJ and Gelboin HV: Role of human
cytochromes P450 in the metabolic activation of chemical
carcinogens and toxins. Drug Metab Rev. 26:165–183. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ding X and Kaminsky LS: Human extrahepatic
cytochromes P450: Function in xenobiotic metabolism and
tissue-selective chemical toxicity in the respiratory and
gastrointestinal tracts. Annu Rev Pharmacol Toxicol. 43:149–173.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Murray GI: The role of cytochrome P450 in
tumour development and progression and its potential in therapy. J
Pathol. 192:419–426. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gajjar K, Martin-Hirsch PL and Martin FL:
CYP1B1 and hormone-induced cancer. Cancer Lett. 324:13–30. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nebert DW and Russell DW: Clinical
importance of the cytochromes P450. Lancet. 360:1155–1162. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luthra A, Denisov IG and Sligar SG:
Spectroscopic features of cytochrome P450 reaction intermediates.
Arch Biochem Biophys. 507:26–35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nelson DR: Cytochrome P450 nomenclature,
2004. Methods Mol Biol. 320:1–10. 2006.PubMed/NCBI
|
|
11
|
Pelkonen O, Turpeinen M, Hakkola J,
Honkakoski P, Hukkanen J and Raunio H: Inhibition and induction of
human cytochrome P450 enzymes: Current status. Arch Toxicol.
82:667–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nelson DR, Zeldin DC, Hoffman SM, Maltais
LJ, Wain HM and Nebert DW: Comparison of cytochrome P450 (CYP)
genes from the mouse and human genomes, including nomenclature
recommendations for genes, pseudogenes and alternative-splice
variants. Pharmacogenetics. 14:1–18. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Thelen K and Dressman JB: Cytochrome
P450-mediated metabolism in the human gut wall. J Pharm Pharmacol.
61:541–558. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Renaud HJ, Cui JY, Khan M and Klaassen CD:
Tissue distribution and gender-divergent expression of 78
cytochrome P450 mRNAs in mice. Toxicol Sci. 124:261–277. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Guengerich FP: Mechanisms of cytochrome
P450 substrate oxidation: MiniReview. J Biochem Mol Toxicol.
21:163–168. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Furge LL and Guengerich FP: Cytochrome
P450 enzymes in drug metabolism and chemical toxicology: An
introduction. Biochem Mol Biol Educ. 34:66–74. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guengerich FP: Cytochrome p450 and
chemical toxicology. Chem Res Toxicol. 21:70–83. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bozina N, Bradamante V and Lovric M:
Genetic polymorphism of metabolic enzymes P450 (CYP) as a
susceptibility factor for drug response, toxicity, and cancer risk.
Arh Hig Rada Toksikol. 60:217–242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Missmer SA, Eliassen AH, Barbieri RL and
Hankinson SE: Endogenous estrogen, androgen, and progesterone
concentrations and breast cancer risk among postmenopausal women. J
Natl Cancer Inst. 96:1856–1865. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chan MF, Dowsett M, Folkerd E, Bingham S,
Wareham N, Luben R, Welch A and Khaw KT: Usual physical activity
and endogenous sex hormones in postmenopausal women: The European
prospective investigation into cancer-norfolk population study.
Cancer Epidemiol Biomarkers Prev. 16:900–905. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baglietto L, Severi G, English DR,
Krishnan K, Hopper JL, Mclean C, Morris HA, Tilley WD and Giles GG:
Circulating steroid hormone levels and risk of breast cancer for
postmenopausal women. Cancer Epidemiol Biomarkers Prev. 19:492–502.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Rod NH, Hansen AM, Nielsen J, Schnohr P
and Gronbaek M: Low-risk factor profile, estrogen levels, and
breast cancer risk among postmenopausal women. Int J Cancer.
124:1935–1940. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pearce ST and Jordan VC: The biological
role of estrogen receptors alpha and beta in cancer. Crit Rev Oncol
Hematol. 50:3–22. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen GG, Zeng Q and Tse GM: Estrogen and
its receptors in cancer. Med Res Rev. 28:954–974. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guillette TC, Jackson TW and Belcher SM:
Duality of estrogen receptor β action in cancer progression. Curr
Opin Pharmacol. 41:66–73. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Frech MS, Torre KM, Robinson GW and Furth
PA: Loss of cyclin D1 in concert with deregulated estrogen receptor
alpha expression induces DNA damage response activation and
interrupts mammary gland morphogenesis. Oncogene. 27:3186–3193.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Giuliano M, Trivedi MV and Schiff R:
Bidirectional Crosstalk between the estrogen receptor and human
epidermal growth factor receptor 2 signaling pathways in breast
cancer: Molecular basis and clinical implications. Breast Care
(Basel). 8:256–262. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Levin ER: Bidirectional signaling between
the estrogen receptor and the epidermal growth factor receptor. Mol
Endocrinol. 17:309–317. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sangrajrang S, Sato Y, Sakamoto H, Ohnami
S, Laird NM, Khuhaprema T, Brennan P, Boffetta P and Yoshida T:
Genetic polymorphisms of estrogen metabolizing enzyme and breast
cancer risk in Thai women. Int J Cancer. 125:837–843. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Samavat H and Kurzer MS: Estrogen
metabolism and breast cancer. Cancer Lett. 356:231–243. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liehr JG: 4-hydroxylation of oestrogens as
a marker for mammary tumours. Biochem Soc Trans. 27:318–323. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liehr JG, Ulubelen AA and Strobel HW:
Cytochrome P-450-mediated redox cycling of estrogens. J Biol Chem.
261:16865–16870. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yager JD and Liehr JG: Molecular
mechanisms of estrogen carcinogenesis. Annu Rev Pharmacol Toxicol.
36:203–232. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cavalieri EL, Stack DE, Devanesan PD,
Todorovic R, Dwivedy I, Higginbotham S, Johansson SL, Patil KD,
Gross ML, Gooden JK, et al: Molecular origin of cancer: Catechol
estrogen-3,4-quinones as endogenous tumor initiators. Proc Natl
Acad Sci USA. 94:10937–10942. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bradlow HL, Hershcopf RJ, Martucci CP and
Fishman J: Estradiol 16 alpha-hydroxylation in the mouse correlates
with mammary tumor incidence and presence of murine mammary tumor
virus: A possible model for the hormonal etiology of breast cancer
in humans. Proc Natl Acad Sci USA. 82:6295–6299. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kabat GC, Chang CJ, Sparano JA, Sepkovie
DW, Hu XP, Khalil A, Rosenblatt R and Bradlow HL: Urinary estrogen
metabolites and breast cancer: A case-control study. Cancer
Epidemiol Biomarkers Prev. 6:505–509. 1997.PubMed/NCBI
|
|
37
|
Meilahn EN, De Stavola B, Allen DS,
Fentiman I, Bradlow HL, Sepkovic DW and Kuller LH: Do urinary
oestrogen metabolites predict breast cancer? Guernsey III cohort
follow-up. Br J Cancer. 78:1250–1255. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lo R, Burgoon L, Macpherson L, Ahmed S and
Matthews J: Estrogen receptor-dependent regulation of CYP2B6 in
human breast cancer cells. Biochim Biophys Acta. 1799:469–479.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fukasawa K, Kagaya S, Maruyama S, Kuroiwa
S, Masuda K, Kameyama Y, Satoh Y, Akatsu Y, Tomura A, Nishikawa K,
et al: A novel compound, NK150460, exhibits selective antitumor
activity against breast cancer cell lines through activation of
aryl hydrocarbon receptor. Mol Cancer Ther. 14:343–354. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mcdaniel DO, Thurber T, Lewis-Traylor A,
Berry C, Barber WH, Zhou X, Bigler S and Vance R: Differential
association of cytochrome P450 3A4 genotypes with onsets of breast
tumors in African American versus Caucasian patients. J Investig
Med. 59:1096–1103. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Johnson N, Dudbridge F, Orr N, Gibson L,
Jones ME, Schoemaker MJ, Folkerd EJ, Haynes BP, Hopper JL, Southey
MC, et al: Genetic variation at CYP3A is associated with age at
menarche and breast cancer risk: A case-control study. Breast
Cancer Res. 16:R512014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Johnson N, Walker K, Gibson LJ, Orr N,
Folkerd E, Haynes B, Palles C, Coupland B, Schoemaker M, Jones M,
et al: CYP3A variation, premenopausal estrone levels, and breast
cancer risk. J Natl Cancer Inst. 104:657–669. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Mao C, Wang XW, He BF, Qiu LX, Liao RY,
Luo RC and Chen Q: Lack of association between CYP17 MspA1
polymorphism and breast cancer risk: A meta-analysis of 22,090
cases and 28,498 controls. Breast Cancer Res Treat. 122:259–265.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kuo SH, Yang SY, Lien HC, Lo C, Lin CH, Lu
YS, Cheng AL, Chang KJ and Huang CS: CYP19 genetic polymorphism
haplotype AASA is associated with a poor prognosis in premenopausal
women with lymph node-negative, hormone receptor-positive breast
cancer. Biomed Res Int. 2013:5621972013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sergentanis TN and Economopoulos KP: Four
polymorphisms in cytochrome P450 1A1 (CYP1A1) gene and breast
cancer risk: A meta-analysis. Breast Cancer Res Treat. 122:459–469.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cardoso-Filho C, Sarian LO, de Oliveira
CB, Da SBL, Lourenco GJ, Lima CS and Gurgel MS: Clinical effects of
A4889G and T6235C polymorphisms in cytochrome P-450 CYP1A1 for
breast cancer patients treated with tamoxifen: Implications for
tumor aggressiveness and patient survival. Cancer Chemother
Pharmacol. 72:529–535. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bai X, Xie J, Sun S, Zhang X, Jiang Y and
Pang D: The associations of genetic polymorphisms in CYP1A2 and
CYP3A4 with clinical outcomes of breast cancer patients in northern
China. Oncotarget. 8:38367–38377. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Raskin L, Lejbkowicz F, Barnett-Griness O,
Dishon S, Almog R and Rennert G: BRCA1 breast cancer risk is
modified by CYP19 polymorphisms in Ashkenazi Jews. Cancer Epidemiol
Biomarkers Prev. 18:1617–1623. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Abraham JE, Harrington P, Driver KE, Tyrer
J, Easton DF, Dunning AM and Pharoah PD: Common polymorphisms in
the prostaglandin pathway genes and their association with breast
cancer susceptibility and survival. Clin Cancer Res. 15:2181–2191.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mavaddat N, Dunning AM, Ponder BA, Easton
DF and Pharoah PD: Common genetic variation in candidate genes and
susceptibility to subtypes of breast cancer. Cancer Epidemiol
Biomarkers Prev. 18:255–259. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Justenhoven C, Hamann U, Pierl CB, Baisch
C, Harth V, Rabstein S, Spickenheuer A, Pesch B, Bruning T, Winter
S, et al: CYP2C19*17 is associated with decreased breast cancer
risk. Breast Cancer Res Treat. 115:391–396. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ruiter R, Bijl MJ, van Schaik RH, Berns
EM, Hofman A, Coebergh JW, van Noord C, Visser LE and Stricker BH:
CYP2C19*2 polymorphism is associated with increased survival in
breast cancer patients using tamoxifen. Pharmacogenomics.
11:1367–1375. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gan CQ, Wang XY, Cao YD, Ye WX, Liu H and
Sun YY: Association of CYP2C19*3 gene polymorphism with breast
cancer in Chinese women. Genet Mol Res. 10:3514–3519. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yu T and Di G: Role of tumor
microenvironment in triple-negative breast cancer and its
prognostic significance. Chin J Cancer Res. 29:237–252. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen L, Joseph G, Zhang FF, Nguyen H,
Jiang H, Gotlinger KH, Falck JR, Yang J, Schwartzman ML and Guo AM:
20-HETE contributes to ischemia-induced angiogenesis. Vascul
Pharmacol. 83:57–65. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Garcia V, Joseph G, Shkolnik B, Ding Y,
Zhang FF, Gotlinger K, Falck JR, Dakarapu R, Capdevila JH,
Bernstein KE and Schwartzman ML: Angiotensin II receptor blockade
or deletion of vascular endothelial ACE does not prevent vascular
dysfunction and remodeling in 20-HETE-dependent hypertension. Am J
Physiol Regul Integr Comp Physiol. 309:R71–R78. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Seki T, Wang MH, Miyata N and
Laniado-Schwartzman M: Cytochrome P450 4A isoform inhibitory
profile of N-hydroxy-N′-(4-butyl-2-methylphenyl)-formamidine
(HET0016), a selective inhibitor of 20-HETE synthesis. Biol Pharm
Bull. 28:1651–1654. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Garcia V, Shkolnik B, Milhau L, Falck JR
and Schwartzman ML: 20-HETE activates the transcription of
angiotensin-converting enzyme via nuclear Factor-κB translocation
and promoter binding. J Pharmacol Exp Ther. 356:525–533. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen L, Ackerman R, Saleh M, Gotlinger KH,
Kessler M, Mendelowitz LG, Falck JR, Arbab AS, Scicli AG,
Schwartzman ML, et al: 20-HETE regulates the angiogenic functions
of human endothelial progenitor cells and contributes to
angiogenesis in vivo. J Pharmacol Exp Ther. 348:442–451. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Guo AM, Janic B, Sheng J, Falck JR, Roman
RJ, Edwards PA, Arbab AS and Scicli AG: The cytochrome P450
4A/F-20-hydroxyeicosatetraenoic acid system: A regulator of
endothelial precursor cells derived from human umbilical cord
blood. J Pharmacol Exp Ther. 338:421–429. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Borin TF, Shankar A, Angara K, Rashid MH,
Jain M, Iskander A, Ara R, Lebedyeva I, Korkaya H, Achyut BR and
Arbab AS: HET0016 decreases lung metastasis from breast cancer in
immune-competent mouse model. PLoS One. 12:e1788302017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Borin TF, Zuccari DA, Jardim-Perassi BV,
Ferreira LC, Iskander AS, Varma NR, Shankar A, Guo AM, Scicli G and
Arbab AS: HET0016, a selective inhibitor of 20-HETE synthesis,
decreases pro-angiogenic factors and inhibits growth of triple
negative breast cancer in mice. PLoS One. 9:e1162472014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shankar A, Borin TF, Iskander A, Varma NR,
Achyut BR, Jain M, Mikkelsen T, Guo AM, Chwang WB, Ewing JR, et al:
Combination of vatalanib and a 20-HETE synthesis inhibitor results
in decreased tumor growth in an animal model of human glioma. Onco
Targets Ther. 9:1205–1219. 2016.PubMed/NCBI
|
|
64
|
Jain M, Gamage NH, Alsulami M, Shankar A,
Achyut BR, Angara K, Rashid MH, Iskander A, Borin TF, Wenbo Z, et
al: Intravenous formulation of HET0016 decreased human glioblastoma
growth and implicated survival benefit in rat xenograft models. Sci
Rep. 7:418092017. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Rieger MA, Ebner R, Bell DR, Kiessling A,
Rohayem J, Schmitz M, Temme A, Rieber EP and Weigle B:
Identification of a novel mammary-restricted cytochrome P450,
CYP4Z1, with overexpression in breast carcinoma. Cancer Res.
64:2357–2364. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Yu W, Chai H, Li Y, Zhao H, Xie X, Zheng
H, Wang C, Wang X, Yang G, Cai X, et al: Increased expression of
CYP4Z1 promotes tumor angiogenesis and growth in human breast
cancer. Toxicol Appl Pharmacol. 264:73–83. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wang B, Zheng L, Chou J, Li C, Zhang Y,
Meng X and Xi T: CYP4Z1 3′UTR represses migration of human breast
cancer cells. Biochem Biophys Res Commun. 478:900–907. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zheng L, Li X, Meng X, Chou J, Hu J, Zhang
F, Zhang Z, Xing Y, Liu Y and Xi T: Competing endogenous RNA
networks of CYP4Z1 and pseudogene CYP4Z2P confer tamoxifen
resistance in breast cancer. Mol Cell Endocrinol. 427:133–142.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Chen P and Bonaldo P: Role of macrophage
polarization in tumor angiogenesis and vessel normalization:
Implications for new anticancer therapies. Int Rev Cell Mol Biol.
301:1–35. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Tariq M, Zhang J, Liang G, Ding L, He Q
and Yang B: Macrophage polarization: Anti-cancer strategies to
target tumor-associated macrophage in breast cancer. J Cell
Biochem. 118:2484–2501. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Lao L, Fan S and Song E: Tumor associated
macrophages as therapeutic targets for breast cancer. Adv Exp Med
Biol. 1026:331–370. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Choi J, Gyamfi J, Jang H and Koo JS: The
role of tumor-associated macrophage in breast cancer biology.
Histol Histopathol. 33:133–145. 2018.PubMed/NCBI
|
|
73
|
Chen XW, Yu TJ, Zhang J, Li Y, Chen HL,
Yang GF, Yu W, Liu YZ, Liu XX, Duan CF, et al: CYP4A in
tumor-associated macrophages promotes pre-metastatic niche
formation and metastasis. Oncogene. 36:5045–5057. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Jia X, Yu F, Wang J, Iwanowycz S, Saaoud
F, Wang Y, Hu J, Wang Q and Fan D: Emodin suppresses pulmonary
metastasis of breast cancer accompanied with decreased macrophage
recruitment and M2 polarization in the lungs. Breast Cancer Res
Treat. 148:291–302. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Ding L, Liang G, Yao Z, Zhang J, Liu R,
Chen H, Zhou Y, Wu H, Yang B and He Q: Metformin prevents cancer
metastasis by inhibiting M2-like polarization of tumor associated
macrophages. Oncotarget. 6:36441–36455. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lewis CE and Pollard JW: Distinct role of
macrophages in different tumor microenvironments. Cancer Res.
66:605–612. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Hoopes SL, Garcia V, Edin ML, Schwartzman
ML and Zeldin DC: Vascular actions of 20-HETE. Prostaglandins Other
Lipid Mediat. 120:9–16. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Alexanian A and Sorokin A: Targeting
20-HETE producing enzymes in cancer-rationale, pharmacology, and
clinical potential. Onco Targets Ther. 6:243–255. 2013.PubMed/NCBI
|
|
79
|
Edson KZ and Rettie AE: CYP4 enzymes as
potential drug targets: Focus on enzyme multiplicity, inducers and
inhibitors, and therapeutic modulation of
20-hydroxyeicosatetraenoic acid (20-HETE) synthase and fatty acid
omega-hydroxylase activities. Curr Top Med Chem. 13:1429–1440.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Borin TF, Angara K, Rashid MH, Achyut BR
and Arbab AS: Arachidonic acid metabolite as a novel therapeutic
target in breast cancer metastasis. Int J Mol Sci. 18:26612017.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Alexanian A, Miller B, Roman RJ and
Sorokin A: 20-HETE-producing enzymes are up-regulated in human
cancers. Cancer Genomics Proteomics. 9:163–169. 2012.PubMed/NCBI
|
|
82
|
Zheng H, Li Y, Wang Y, Zhao H, Zhang J,
Chai H, Tang T, Yue J, Guo AM and Yang J: Downregulation of COX-2
and CYP 4A signaling by isoliquiritigenin inhibits human breast
cancer metastasis through preventing anoikis resistance, migration
and invasion. Toxicol Appl Pharmacol. 280:10–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Bandala C, Floriano-Sanchez E,
Cardenas-Rodriguez N, Lopez-Cruz J and Lara-Padilla E: RNA
expression of cytochrome P450 in Mexican women with breast cancer.
Asian Pac J Cancer Prev. 13:2647–2653. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Casos K, Siguero L, Fernandez-Figueras MT,
Leon X, Sarda MP, Vila L and Camacho M: Tumor cells induce COX-2
and mPGES-1 expression in microvascular endothelial cells mainly by
means of IL-1 receptor activation. Microvasc Res. 81:261–268. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Greene ER, Huang S, Serhan CN and
Panigrahy D: Regulation of inflammation in cancer by eicosanoids.
Prostaglandins Other Lipid Mediat. 96:27–36. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Jiang JG, Chen CL, Card JW, Yang S, Chen
JX, Fu XN, Ning YG, Xiao X, Zeldin DC and Wang DW: Cytochrome P450
2J2 promotes the neoplastic phenotype of carcinoma cells and is
up-regulated in human tumors. Cancer Res. 65:4707–4715. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Wei X, Zhang D, Dou X, Niu N, Huang W, Bai
J and Zhang G: Elevated 14,15-epoxyeicosatrienoic acid by
increasing of cytochrome P450 2C8, 2C9 and 2J2 and decreasing of
soluble epoxide hydrolase associated with aggressiveness of human
breast cancer. BMC Cancer. 14:8412014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Mitra R, Guo Z, Milani M, Mesaros C,
Rodriguez M, Nguyen J, Luo X, Clarke D, Lamba J, Schuetz E, et al:
CYP3A4 mediates growth of estrogen receptor-positive breast cancer
cells in part by inducing nuclear translocation of phospho-Stat3
through biosynthesis of (±)-14,15-epoxyeicosatrienoic acid (EET). J
Biol Chem. 286:17543–17559. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yang S, Lin L, Chen JX, Lee CR, Seubert
JM, Wang Y, Wang H, Chao ZR, Tao DD, Gong JP, et al: Cytochrome
P-450 epoxygenases protect endothelial cells from apoptosis induced
by tumor necrosis factor-alpha via MAPK and PI3K/Akt signaling
pathways. Am J Physiol Heart Circ Physiol. 293:H142–H151. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Jiang JG, Ning YG, Chen C, Ma D, Liu ZJ,
Yang S, Zhou J, Xiao X, Zhang XA, Edin ML, et al: Cytochrome p450
epoxygenase promotes human cancer metastasis. Cancer Res.
67:6665–6674. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Murray GI, Patimalla S, Stewart KN, Miller
ID and Heys SD: Profiling the expression of cytochrome P450 in
breast cancer. Histopathology. 57:202–211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Haas S, Pierl C, Harth V, Pesch B,
Rabstein S, Bruning T, Ko Y, Hamann U, Justenhoven C, Brauch H and
Fischer HP: Expression of xenobiotic and steroid hormone
metabolizing enzymes in human breast carcinomas. Int J Cancer.
119:1785–1791. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Gonzalez FJ: Role of cytochromes P450 in
chemical toxicity and oxidative stress: Studies with CYP2E1. Mutat
Res. 569:101–110. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Vaclavikova R, Hubackova M,
Stribrna-Sarmanova J, Kodet R, Mrhalova M, Novotny J, Gut I and
Soucek P: RNA expression of cytochrome P450 in breast cancer
patients. Anticancer Res. 27:4443–4450. 2007.PubMed/NCBI
|
|
95
|
Leung T, Rajendran R, Singh S, Garva R,
Krstic-Demonacos M and Demonacos C: Cytochrome P450 2E1 (CYP2E1)
regulates the response to oxidative stress and migration of breast
cancer cells. Breast Cancer Res. 15:R1072013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Cardenas-Rodriguez N, Lara-Padilla E,
Bandala C, Lopez-Cruz J, Uscanga-Carmona C, Lucio-Monter PF and
Floriano-Sanchez E: CYP2W1, CYP4F11 and CYP8A1 polymorphisms and
interaction of CYP2W1 genotypes with risk factors in Mexican women
with breast cancer. Asian Pac J Cancer Prev. 13:837–846. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Tan BS, Tiong KH, Muruhadas A, Randhawa N,
Choo HL, Bradshaw TD, Stevens MF and Leong CO: CYP2S1 and CYP2W1
mediate 2-(3,4-dimethoxyphenyl)-5-fluorobenzothiazole (GW-610, NSC
721648) sensitivity in breast and colorectal cancer cells. Mol
Cancer Ther. 10:1982–1992. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Moorthy B, Chu C and Carlin DJ: Polycyclic
aromatic hydrocarbons: From metabolism to lung cancer. Toxicol Sci.
145:5–15. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Goth-Goldstein R, Stampfer MR, Erdmann CA
and Russell M: Interindividual variation in CYP1A1 expression in
breast tissue and the role of genetic polymorphism. Carcinogenesis.
21:2119–2122. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Nilsson R, Antic R, Berni A, Dallner G,
Dettbarn G, Gromadzinska J, Joksic G, Lundin C, Palitti F,
Prochazka G, et al: Exposure to polycyclic aromatic hydrocarbons in
women from Poland, Serbia and Italy-relation between PAH metabolite
excretion, DNA damage, diet and genotype (the EU DIEPHY project).
Biomarkers. 18:165–173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Al-Dhfyan A, Alhoshani A and Korashy HM:
Aryl hydrocarbon receptor/cytochrome P450 1A1 pathway mediates
breast cancer stem cells expansion through PTEN inhibition and
beta-Catenin and Akt activation. Mol Cancer. 16:142017. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Hsu MH, Baer BR, Rettie AE and Johnson EF:
The crystal structure of cytochrome P450 4B1 (CYP4B1) monooxygenase
complexed with octane discloses several structural adaptations for
omega-Hydroxylation. J Biol Chem. 292:5610–5621. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Mcfadyen MC, Mcleod HL, Jackson FC, Melvin
WT, Doehmer J and Murray GI: Cytochrome P450 CYP1B1 protein
expression: A novel mechanism of anticancer drug resistance.
Biochem Pharmacol. 62:207–212. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Marsh S, Somlo G, Li X, Frankel P, King
CR, Shannon WD, Mcleod HL and Synold TW: Pharmacogenetic analysis
of paclitaxel transport and metabolism genes in breast cancer.
Pharmacogenomics J. 7:362–365. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Bray J, Sludden J, Griffin MJ, Cole M,
Verrill M, Jamieson D and Boddy AV: Influence of pharmacogenetics
on response and toxicity in breast cancer patients treated with
doxorubicin and cyclophosphamide. Br J Cancer. 102:1003–1009. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Seredina TA, Goreva OB, Talaban VO,
Grishanova AY and Lyakhovich VV: Association of cytochrome P450
genetic polymorphisms with neoadjuvant chemotherapy efficacy in
breast cancer patients. BMC Med Genet. 13:452012. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Schroth W, Antoniadou L, Fritz P, Schwab
M, Muerdter T, Zanger UM, Simon W, Eichelbaum M and Brauch H:
Breast cancer treatment outcome with adjuvant tamoxifen relative to
patient CYP2D6 and CYP2C19 genotypes. J Clin Oncol. 25:5187–5193.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Daraei B, Aghvami M, Pourahmad J and
Dinarvand R: A comparison of hepatocyte cytotoxic mechanisms for
docetaxel and PLGA-docetaxel Nanoparticls. Iran J Pharm Res.
16:249–265. 2017.PubMed/NCBI
|
|
109
|
Jenkins P, Scaife J and Freeman S:
Validation of a predictive model that identifies patients at high
risk of developing febrile neutropaenia following chemotherapy for
breast cancer. Ann Oncol. 23:1766–1771. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Hlavac V, Brynychova V, Vaclavikova R,
Ehrlichova M, Vrana D, Pecha V, Trnkova M, Kodet R, Mrhalova M,
Kubackova K, et al: The role of cytochromes p450 and aldo-keto
reductases in prognosis of breast carcinoma patients. Medicine
(Baltimore). 93:e2552014. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Nakajima M, Komagata S, Fujiki Y, Kanada
Y, Ebi H, Itoh K, Mukai H, Yokoi T and Minami H: Genetic
polymorphisms of CYP2B6 affect the
pharmacokinetics/pharmacodynamics of cyclophosphamide in Japanese
cancer patients. Pharmacogenet Genomics. 17:431–445. 2007.
View Article : Google Scholar : PubMed/NCBI
|