Introduction
Melanoma is one of the most aggressive types of
cancer with an unpredictable course of disease. It accounts for 4%
of all skin cancer cases but is responsible for 75% of deaths
caused by these malignancies (1).
Surgical removal is the best treatment for localised melanoma. For
patients with metastatic disease, surgical treatment alone will not
be curative, and chemotherapy and targeted therapy are necessary
(2). However, the prognosis for
patients treated by chemotherapy is unsatisfactory, and the side
effects are serious, and the targeted therapy is limited to
specific genotype patients (2).
Therefore, effective drugs should be developed for the treatment of
melanoma.
Sanghuangporus spp., also called ‘Sanghuang’
in China, are a well-known medicinal mushroom (3). Sanghuang have been used in Chinese
traditional medicine for >2,000 years for the treatment of
various diseases such as haemorrhage, haemostasis and diseases
related to female menstruation (4).
In recent years, modern pharmacological researches have revealed
that Sanghuangporus spp. have multiple functions, such as
anti-carcinogenesis, anti-inflammatory, anti-oxidative, anti-fungal
and immunomodulatory activities (5,6). The
extracts of S. rimosus have antiproliferative and apoptotic
activities on human colorectal carcinoma HCT116 cells (7). The total ethanol extracts of S.
igniarius have antitumor activities against gastric cancer
SGC-7901 cells through a mitochondria-dependent pathway (8). Im et al (9) found that the extracts of S.
vaninii are cytotoxic to murine melanoma B16-F10 cells at
25–750 mg/ml. Bae et al (10,11)
demonstrated that polysaccharides from S. gilvus can inhibit
forestomach carcinogenesis in mice and the proliferation of murine
melanoma cells. Protocatechualdehyde induces S-phase arrest and
apoptosis by stimulating the
p27KIP1-CyclinA/D1-cyclin-dependent kinase 2 (CDK2) and
mitochondrial apoptotic pathways in HT-29 cells (12). 3,4-Dihydroxybenzalactone isolated
from S. linteus suppresses human non-small cell lung
carcinoma migration and invasiveness via the ROS-mediated
PI3K/AKT/MAPK/MMP and NFκB signalling pathways (13). 3,4-Dihydroxybenzalacetone isolated
from S. obliguus promotes anti-melanogenesis by
downregulating melanosome maturation and transportation in murine
melanoma B16F10 cells and human epidermal melanocytes (14). The extracts of S. baumii exert
antitumour effects on human melanoma A375 cells in vitro and
in vivo (15). S.
gilvus-derived protocatechualdehyde induces G0/G1-phase arrest
and apoptosis in murine B16-F10 cells (16). Therefore, Sanghuang has important
inhibitory effects on melanoma. However, the effect of S.
vaninii on human melanoma has not been reported. The present
study revealed that aqueous extracts of S. vaninii fruiting
body (referred as SH) acted as potential inhibitors of melanoma by
inhibiting the proliferation of A375 cells and inducing cell cycle
arrest and apoptosis.
Materials and methods
Cells and kits
The human melanoma A375 cell line was obtained from
the Institute of Biochemistry and Cell Biology of the Chinese
Academy of Sciences. The cells were cultured in RPMI-1640 medium
(Thermo Fisher Scientific, Inc.) at 37°C with 5% CO2. The medium
was supplemented with 10% FBS (Thermo Fisher Scientific, Inc.),
penicillin (100 µg/ml) and streptomycin (100 µg/ml). MTT was
obtained from Sigma-Aldrich (Merck KGaA). The Dead Cell Apoptosis
kit with FITC Annexin V and PI was obtained from Thermo Fisher
Scientific, Inc.
Preparation of aqueous extracts of S.
vaninii fruiting body
The fruiting body of S. vaninii was
cultivated at the Sericultural Research Institute of Zhejiang
Academy of Agricultural Sciences (Hangzhou, China). The dried
fruiting body powder was extracted with boiling water for 2 h. The
supernatant was collected after centrifugation at 8,000 × g at room
temperature for 10 min. The supernatant was lyophilised to obtain
extract powders. The content of total polysaccharides and
polyphenols was 5.51 and 23.00%, respectively. The major
polyphenols were 3,4-dihydroxybenzalacetone, phellibaumin D and
interfungin B (17).
Cell proliferation assay
Cell proliferation was determined by trypan blue
staining and the inhibitory ratio was measured by MTT-based
colorimetric assay as described by Zhong et al (16). Briefly, cells at the exponential
growth phase were dispensed into a 96-well microplate at 100
µl/well. After 24 h, 100 µl of different concentrations of SH were
added to prepare final concentrations of 50, 100, 200 and 400
µg/ml. Sterile water (0.1% DMSO) was used as control. The cells
were cultured at 37°C for 24, 36 and 48 h for cell counting. The
supernatants of each well were obtained, and 50 µl of 0.25% trypsin
was added into each well. After incubation at 37°C for 5 min, the
supernatants were re-added to the corresponding well to stop cell
digestion. Cell suspensions (~20 µl) were transferred into a new
1.5 ml centrifuge tube with 20 µl of 0.4% trypan blue solution.
After mixing, 20 µl cell suspension was added to a cell counting
plate for counting with an automatic cell counter (IC1000;
Countstar; ALIT Life Science Co., Ltd.). Cells cultured for 48 h
were used for the MTT assay. The cells were incubated at 37°C with
50 µl of MTT solution (1 mg/ml) for 2 h, and the resulting crystals
were dissolved in DMSO. The absorbance at 570 nm was recorded to
assess the formation of formazan. Inhibitory ratio (%) =
[1-absorbance (test)/absorbance (control)] × 100%, where the test
and control correspond to treated and untreated cells,
respectively.
Cell cycle and apoptosis analysis
The cell cycle phase was measured by assessing the
DNA content via flow cytometry as previously described by Zhong
et al (16). Briefly, A375
cells (1×105 cells/well) were incubated at 37°C with SH
at final concentrations of 0 (control), 200 and 400 µg/ml for 48 h.
The proportion of cells at different phases was determined using a
flow cytometer (Cytomics FC 500 MCL; Beckman Coulter, Inc.).
MultiCycle AV software (CXP V2.3 WIN7, C30309; Phoenix Flow
Systems, Inc.) was used for analysis. Apoptosis rate was measured
by staining with Annexin V-FITC (5 µl) and PI (5 µl) at 4°C for 15
min. Annexin V−/PI− (lower left quadrant)
cells were alive cells, Annexin V+/PI− (lower
right quadrant) cells were early apoptotic cells, Annexin
V+/PI+ (upper right quadrant) cells were late
apoptotic cells and Annexin V−/PI+ (upper
left quadrant) cells were necrotic cells. The proportion of cells
was determined using a flow cytometer (Cytomics FC 500 MCL; Beckman
Coulter, Inc.) with MultiCycle AV software (CXP V2.3 WIN7, C30309;
Phoenix Flow Systems, Inc.). All experiments were performed in
three biological duplicates.
Reverse transcription-quantitative
(RT-q)PCR analysis
Total RNA was isolated from A375 cells treated with
or without SH (400 µg/ml) for 6 and 48 h using the Takara MiniBEST
universal RNA extraction kit (Takara Bio, Inc.). PrimeScript RT
reagent kit with gDNA Eraser (Takara Bio, Inc.) and
SYBR® Fast qPCR Mix (Takara Bio, Inc.) were used for the
CFX96 real-time PCR system (Bio-Rad Laboratories, Inc.). All
operations were performed in accordance with the manufacturer's
instructions. The thermocycling conditions consisted of initial
denaturation at 95°C for 30 sec, followed by 40 cycles at 95°C for
5 sec and 60°C for 30 sec. GAPDH was used as an internal reference
gene, and gene expression was quantified using the comparative
2−∆∆Cq method (18). PCR
primer sequences are summarised in Table
I.
 | Table I.Primers used for reverse
transcription-quantitative PCR. |
Table I.
Primers used for reverse
transcription-quantitative PCR.
| Gene | Forward primer
(5–3) | Reverse primers
(5–3) |
|---|
| TGFB |
GGACCAGTGGGGAACACTAC |
AGAGTCCCTGCATCTCAGAGT |
| Smad2 |
CCATACCAAGGTCTCTTGATGGT |
ACTGTGAAGATCAGGCCAGC |
| Smad4 |
GCAGTCCTACTTCCAGTCCAG |
CCCATCTGAGTCTAATGCTACC |
| p21 |
AGCGACCTTCCTCATCCACC |
AAGACAACTACTCCCAGCCCCATA |
| p53 |
AGCTTTGAGGTGCGTGTTTGTG |
TCTCCATCCAGTGGTTTCTTCTTTG |
| p27 |
AAAAGCAACAGAAACCTATCCTCAC |
ATTCAAAACTCCCAAGCACCTC |
| Cyclin A |
TGGAAAGCAAACAGTAAACAGCC |
GGGCATCTTCACGCTCTATTT |
| Cyclin B |
GCAGCACCTGGCTAAGAATGT |
GCCTTGGCTAAATCTTGAACT |
| Cyclin C |
GCAGGACATGGGCCAAGAAGA |
TATCCACAGAAAGCTCAGCAAACCA |
| Cyclin D |
CCCTCGGTGTCCTACTTCAAATGT |
GGAAGCGGTCCAGGTAGTTCAT |
| Cyclin E |
GCCAGCCTTGGGACAATAATG |
CTTGCACGTTGAGTTTGGGT |
| CDK1 |
GGATGTGCTTATGCAGGATTCC |
CATGTACTGACCAGGAGGGATAG |
| CDK2 |
CCAGGAGTTACTTCTATGCCTGA |
TTCATCCAGGGGAGGTACAAC |
| CDK3 |
CCAGCTCTTTCGTATCTTTCGT |
TTCCTGGTCCACTTAGGGAAG |
| CDK4 |
ATGGCTACCTCTCGATATGAGC |
CATTGGGGACTCTCACACTCT |
| CDK5 |
GGAAGGCACCTACGGAACTG |
GGCACACCCTCATCATCGT |
| CDK6 |
TCTTCATTCACACCGAGTAGTGC |
TGAGGTTAGAGCCATCTGGAAA |
| CDK7 |
ATGGCTCTGGACGTGAAGTCT |
GCGACAATTTGGTTGGTGTTC |
| Rb |
TTGTAACGGGAGTCGGGAGA |
CAGCGAGCTGTGGAGGAG |
| E2F1 |
CACTTTCGGCCCTTTTGCTC |
GTGCTCTCACCGTCCTACAC |
| DP1 |
ATGGCTCAGGGAACTGTTGG |
GGTCACTGGCAGAGAACCTT |
| Bad |
TTGGGGTGAGACCTGTGCG |
CTCAGTCTCCCCTCAGAACCC |
| Bik |
CATTCCAAAGAATCGAAGTCC |
AATGTCTGAAGTAAGACCCCTCT |
| Bax |
CCCGAGAGGTCTTTTTCCGAG |
CCAGCCCATGATGGTTCTGAT |
| Bid |
AGGAAGCCAAACACCAGTAGG |
TGTGAACCAGGAGTGAGTCGG |
| Bim |
TTTCTTGCGTTTCTCAGTCCG |
CGCCACTACCACCACTTGATT |
| Bcl-xs |
TCCCCATGGCAGCAGTAAAG |
TCCACAAAAGTATCCTGTTCAAAGC |
| Bak |
GCAGGCTGATCCCGTCC |
CTGCGGAAAACCTCCTCTGT |
| Bcl-2 |
GGTGGGGTCATGTGTGTGG |
CGGTTCAGGTACTCAGTCATCC |
| Bcl-xl |
GCTGGGACACTTTTGTGGAT |
GAGCCCAGCAGAACTACACC |
| Bcl-w |
CTTTGTCTTTGGGGCTGCAC |
CCATCCACTCCTGCACTTGT |
| Ras |
AAGTGTGTGCTCTCCTGACG |
CACAAGGGAGGCTGCTGAC |
| Raf |
CAACTGATTGCACTGACTGCC |
CCTGGAATTGCTCTGGGGTT |
| MEK1 |
CTTCGCAGAGCGGCTAGG |
CAGCCCGGGATTCCCTTC |
| ERK1 |
GAGATGGATGTGGGTTCCAGTC |
TTCTCATCGTAGCCACTCACC |
| GAPDH |
GGACCTGACCTGCCGTCTAG |
GTAGCCCAGGATGCCCTTGA |
Western blot analysis
A375 cells treated with or without SH (400 µg/ml)
for 48 h were collected for western blot analysis as previously
described (12). Briefly, the total
proteins were extracted using the SD-001 buffer (Invent
Biotechnologies, Inc.) and supplemented with protein phosphatase
inhibitor (Beijing Solarbio Science & Technology Co., Ltd.),
and the protein concentrations were determined using a BCA protein
assay kit. A total of 40 µg protein/lane was separated via 4–12%
SDS-PAGE and then transferred to a polyvinylidene difluoride
membrane. After blocking with 5% BSA (Merck KGaA) for 1 h at room
temperature, membranes were incubated with the corresponding
primary antibodies at 4°C overnight. Subsequently, membranes were
washed with PBS three times and incubated with the corresponding
secondary antibodies at room temperature for 1 h. Primary
antibodies (1:1,000) against p21 (cat. no. ab188224), CyclinD (cat.
no. ab28283), CDK2 (cat. no. ab32147), CDK4 (cat. no. ab131469),
CDK6 (cat. no. ab131469), Bcl-2 (cat. no. ab32124) and β-actin
(cat. no. ab8226) were obtained from Abcam. The secondary antibody
was Alexa Fluor 790-conjugated goat anti-rabbit IgG H&L
(1:10,000; cat. no. ab175781; Abcam). The membranes were washed
with PBS three times at room temperature, and the protein signals
were visualised using the ChemiDoc touch imaging system (Bio-Rad
Laboratories, Inc.) with Image Lab touch software (version 1.2;
Chameleon Power).
Immunofluorescence analysis
A375 cells on a slide were fixed in 4%
paraformaldehyde in PBS for 20 min at room temperature, and then
washed with PBS for 3 times. Cells were blocked with 3% BSA (Merck
KGaA) for 30 min at room temperature, and then primary antibodies
against CyclinD (cat. no. ab28283) and Bcl-2 (cat. no. ab32124)
(both 1:200; Abcam) were added at 4°C overnight. Subsequently, the
slides were washed with PBS and incubated with DyLight
488-conjugated goat anti-rabbit IgG (1:200; cat. no. BA1127; Wuhan
Boster Biological Technology, Ltd.) for 1 h at room temperature.
The slides were mounted with DAPI-containing medium (C1002;
Beyotime Institute of Biotechnology) for counterstaining of nuclei
and then observed under a fluorescence microscope (Nikon Eclipse
80i; Nikon Corporation) at ×100 magnification.
Statistical analysis
Data were expressed as the mean ± SD. Statistical
analysis was performed using SPSS 16.0 software (SPSS, Inc.).
One-way ANOVA was used to analyse statistical differences between
groups under different conditions, followed by Tukey's post-hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
SH inhibits proliferation of A375
melanoma cells
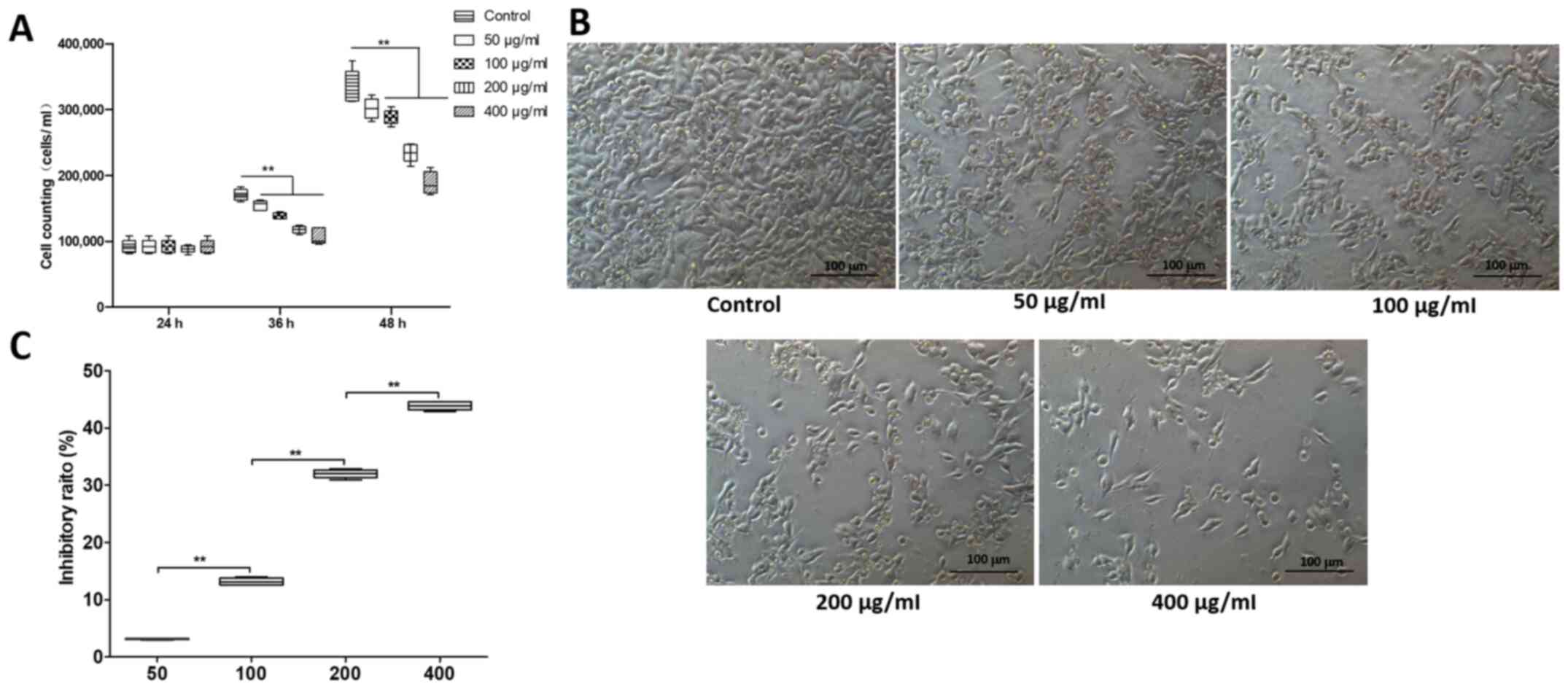
To examine the effects of SH on melanoma cells, A375
cells were treated with different concentrations of SH (0, 50, 100,
200 and 400 µg/ml) for 24, 36 and 48 h (Fig. 1). As shown in Fig. 1A and B, treatment of A375 cells with
SH for 36 and 48 h significantly decreased cell viability compared
with the control group in a dose-dependent manner. Particularly,
400 µg/ml SH effectively inhibited proliferation of A375 cells,
along with a significant inhibitory ratio of >43% (Fig. 1B and C). Hence, SH suppressed the
proliferation of A375 cells in vitro, suggesting that SH may
be used as a cell proliferation inhibitor for the treatment of
melanoma.
SH induces cell cycle arrest at the S
phase and apoptosis
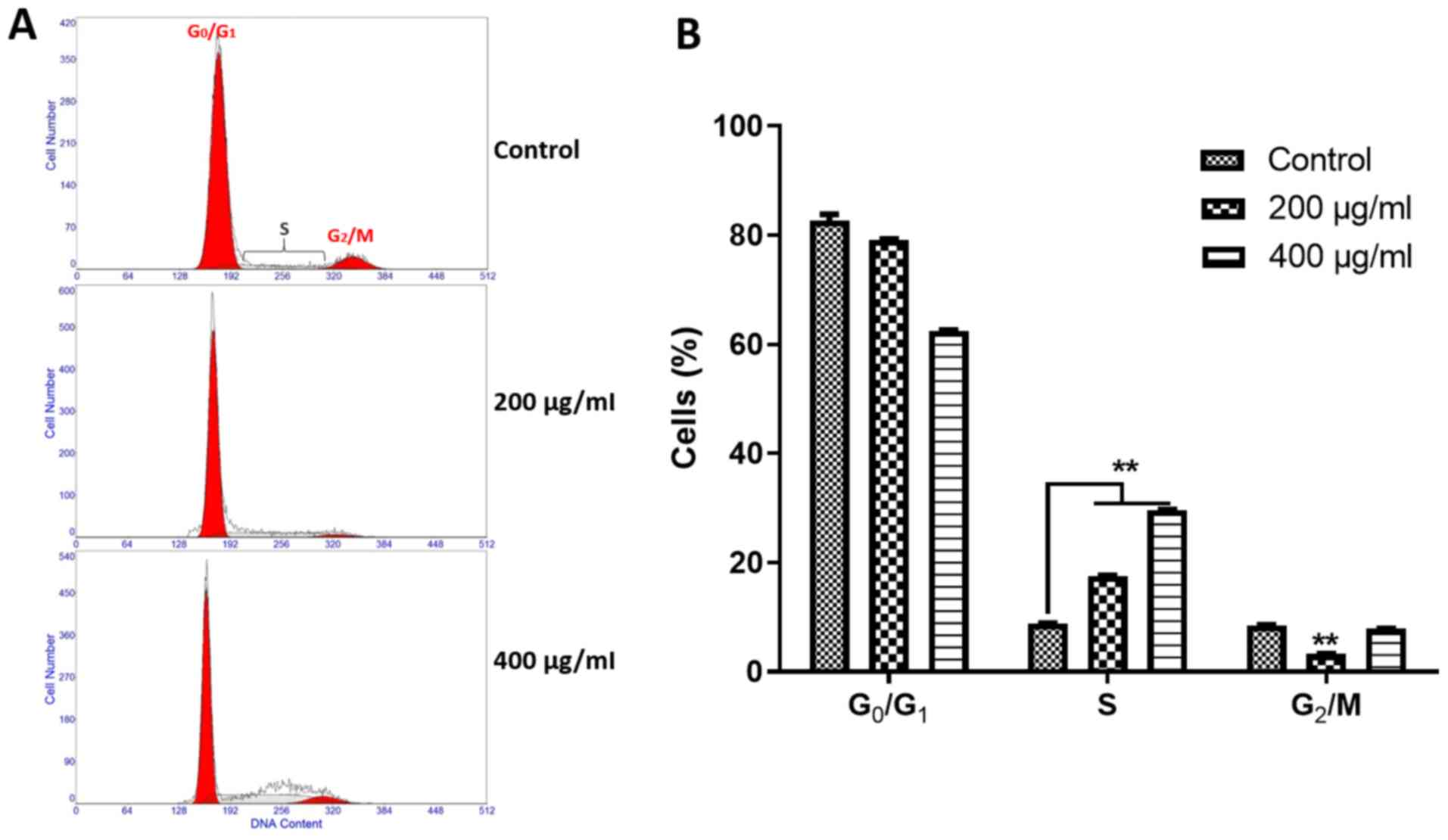
For further investigations on the inhibitory
mechanism of SH on A375 cells, flow cytometry was conducted to
assess whether the anti-proliferative effect of SH on A375 cells
was mediated by cell cycle arrest. Under the higher inhibitory
concentrations of 200 and 400 µg/ml, SH significantly increased the
number of cells in S phase (Fig. 2).
At 200 µg/ml, SH significantly decreased the number of cells at the
G2/M phase, while the number of cells at the G0/G1 phase
significantly decreased with 400 µg/ml SH (Fig. 2). Hence, SH mainly induced cell cycle
arrest of A375 cells at the S phase.
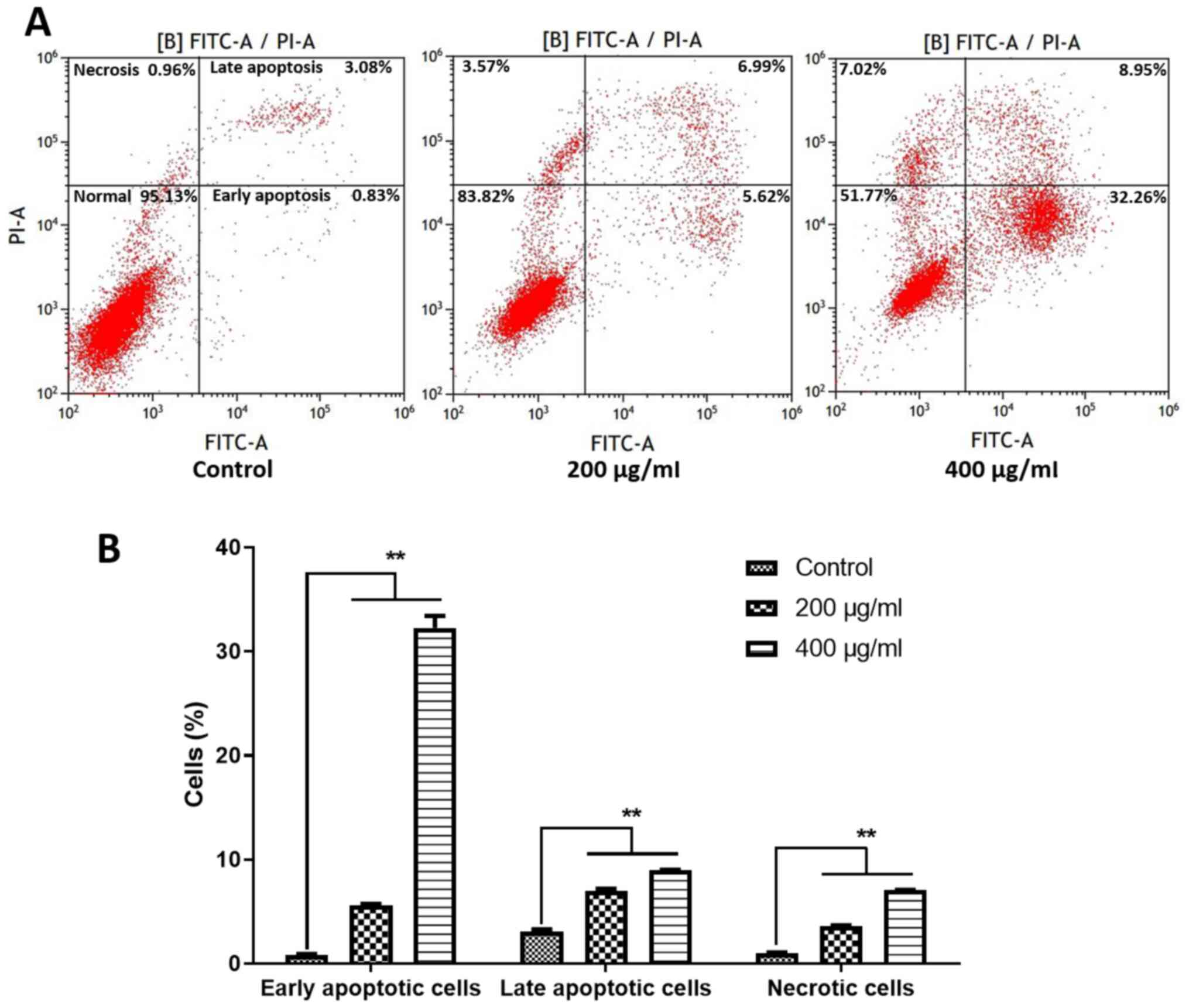
Flow cytometry with Annexin V-FITC and PI staining
was then used to examine the contribution of apoptosis to the
SH-mediated inhibition of cell proliferation. SH treatment
significantly increased early apoptotic, late apoptotic and
necrotic cells compared with the control group (Fig. 3). Hence, S-phase arrest and apoptosis
may be mainly involved in the anti-proliferative effect of SH on
A375 cells.
SH induces S phase arrest via
p21-cyclin-CDK complex signalling pathways
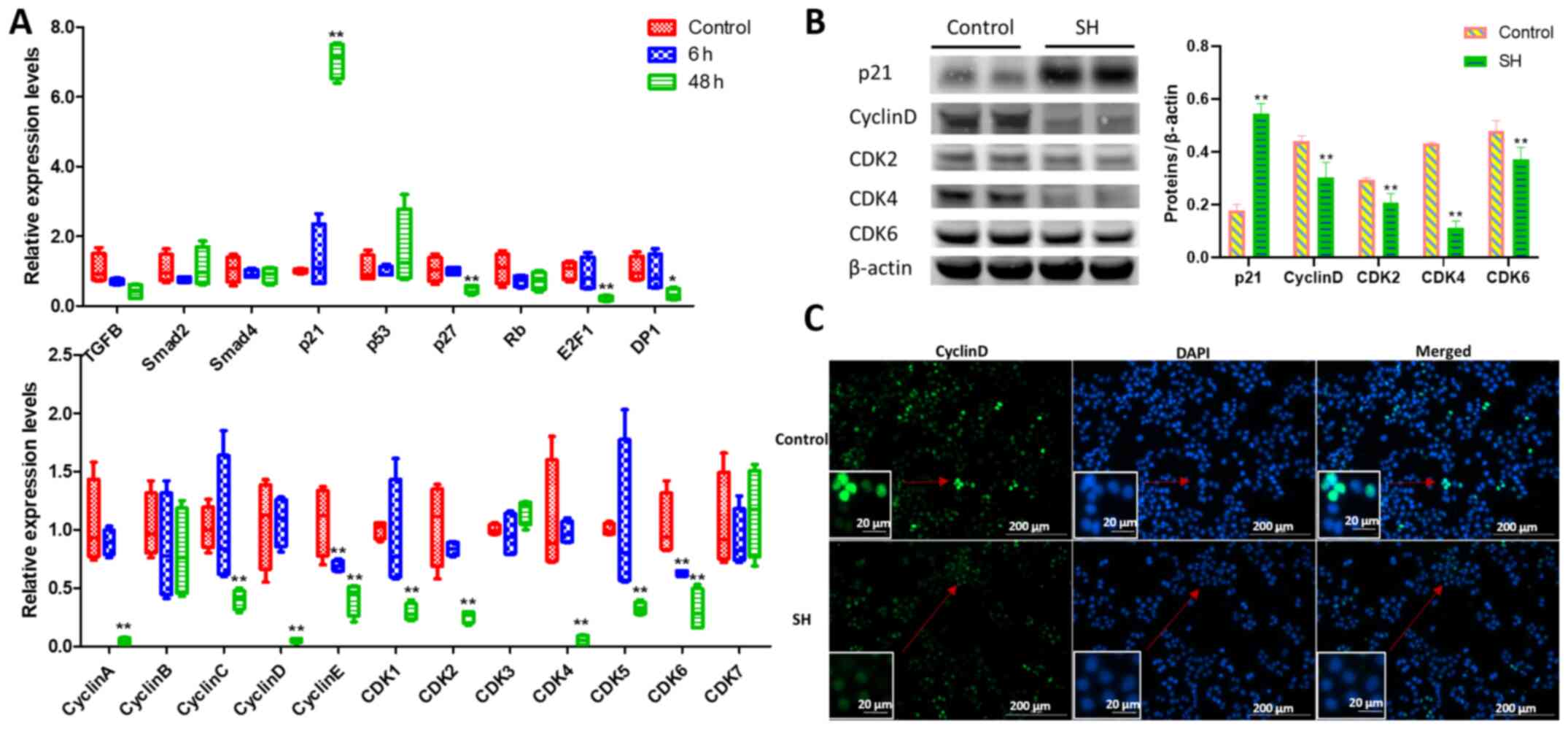
To further elaborate the molecular mechanism
underpinning the anti-proliferative action of SH, RT-qPCR analysis
was performed using A375 cells treated with 400 µg/ml SH for 6 and
48 h, since 400 µg/ml SH exhibited an increased ability of S phase
arrest. A number of key genes involved in cell cycle were analysed.
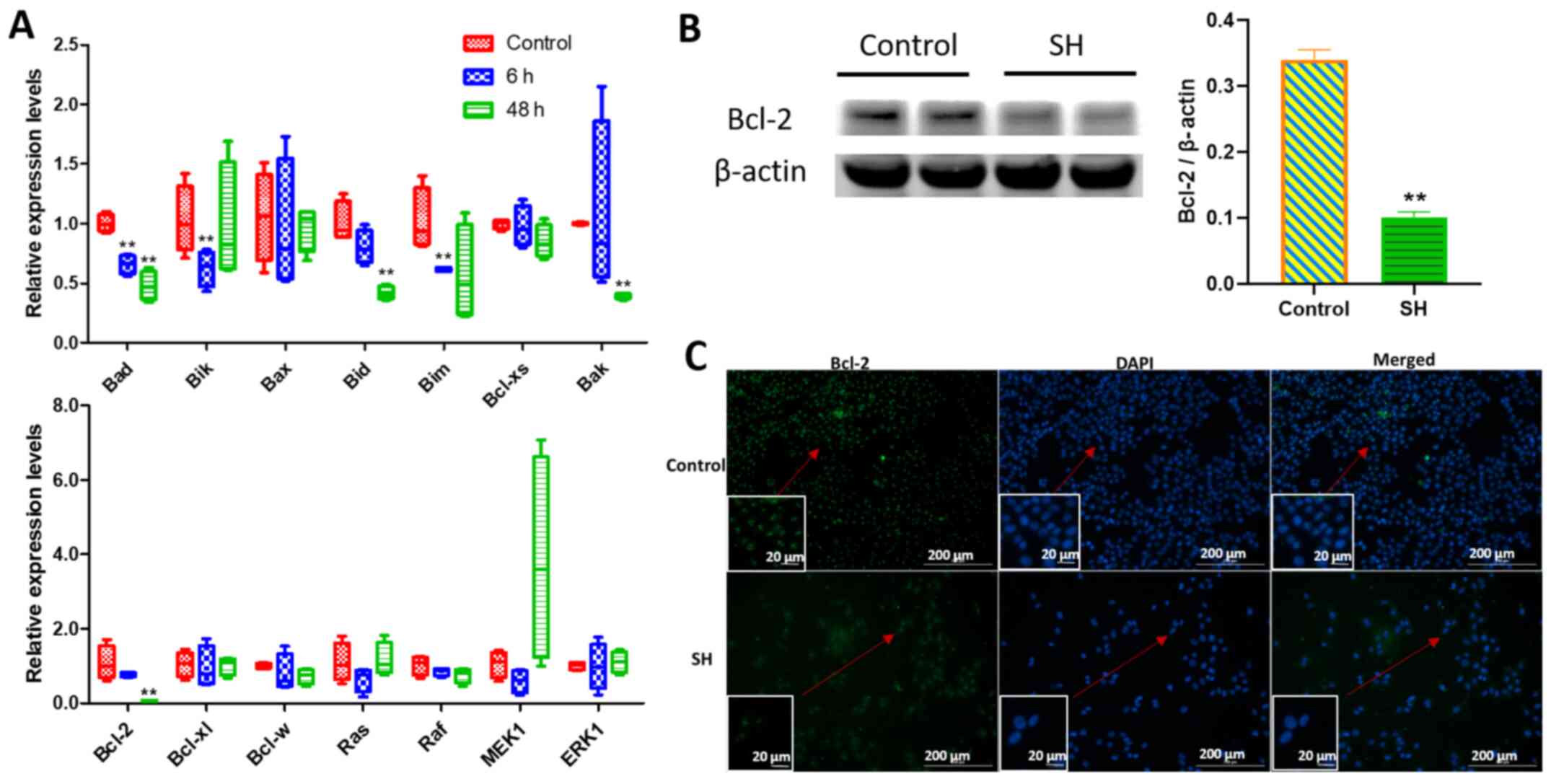
As shown in Fig. 4A, the expression
levels of p21 were significantly
upregulated, whereas those of p27, CyclinsA, C, D and
E, and CDK1, 2, 4, 5 and 6 were significantly
downregulated after 48 h of SH treatment. No significant difference
was observed in the expression levels of TGFB, Smad2, Smad4,
p53, CyclinB, CDK3, CDK7 and Rb. Additionally,
transcription factors E2F1 and DP1 were significantly
downregulated after 48 h of SH treatment.
To confirm the difference in gene expression,
western blot and immunofluorescence analyses were conducted. As
shown in Fig. 4B, the expression
levels of the proteins encoded by some of the aforementioned key
genes, such as p21, CyclinD, CDK2, CDK4 and CDK6, were consistent
with their gene expression levels after 48 h of SH treatment.
Immunofluorescence analysis of CyclinD-stained cells revealed that
the green fluorescence signals in nuclei were strong without SH
treatment, while after treatment with 400 µg/ml SH for 48 h, the
number of cells decreased markedly compared with the control, and
the green fluorescence signals were weaker (Fig. 4C). Therefore, SH may inhibit A375
cell proliferation by inducing cell cycle arrest at the S phase by
upregulating p21 expression, which inhibits the expression levels
of cyclin-CDK complexes.
SH induces apoptosis by downregulating
Bcl-2 expression
Several key genes involved in apoptosis were
analysed by RT-qPCR analysis using A375 cells treated with 400
µg/ml SH for 6 and 48 h, since 400 µg/ml SH strongly induced
apoptosis. The relative mRNA expression levels of pro-apoptotic
genes (Bad, Bik, Bid, Bim and Bak) decreased under SH
treatment. The expression levels of Bik and Bim
decreased at 6 h significantly, Bid and Bak were
downregulated at 48 h significantly, and the relative mRNA
expression levels of Bad decreased at 6 and 48 h
significantly (Fig. 5A). No
significant difference was observed in the expression levels of
MAPK signalling pathway-associated genes (Ras, Raf, MEK1 and
ERK1; Fig. 5A). The
anti-apoptotic gene Bcl-2 was significantly downregulated
after SH treatment at 48 h (Fig.
5A), consistent with the western blot analysis indicating
significantly decreased Bcl-2 protein expression (Fig. 5B). Immunofluorescence analysis of
Bcl-2-stained cells indicated that the cell density decreased
markedly after treatment with 400 µg/ml SH for 48 h, and the green
fluorescence signals were weaker than in the control group
(Fig. 5C). The current results
suggested that SH may induce apoptosis by downregulating Bcl-2
expression.
Discussion
Melanoma is one of the most aggressive types of
cancer. No specific treatment has been developed to cure patients
with metastasis (1). Natural
products are important sources for the identification of novel
drugs for the treatment of various diseases due to their low
toxicity and compelling efficacy (19). Sanghuang mushrooms grow mainly on
wild mulberry tree trunks and possess multifunctional
bioactivities, such as anti-carcinogenesis, anti-fungal and
immunomodulatory activities, as well as
anti-diabetic/hyperlipidaemic effects (5,6),
presenting Sanghuang as a potential natural product reservoir.
Extracts from S. gilvus and S. baumii inhibit murine
or human melanoma cell proliferation both in vitro and in
vivo (10,15,16).
However, to the best of our knowledge, whether S. vaninii
affects melanoma development has not been revealed. In the present
study, it was revealed that aqueous extracts of S. vaninii
(SH) exerted a proliferation inhibitory effect on human melanoma
cells, providing an alternative Sanghuang candidate for developing
effective interventions against melanoma.
In general, cancer is well-characterised by
continuous cell proliferative ability; hence, interruptions on
cancer cell cycle may hinder cancer development (20). In the present study, the effects of
SH on melanoma A375 cell proliferation were assessed by MTT assay,
revealing that SH exerted a prominent anti-proliferative action in
a dose-dependent manner. Perturbations of cell proliferation can be
largely attributed to abnormal modulations on cell cycle and
apoptosis. Therefore, flow cytometry assays were performed to
determine the underlying mechanism. Notably, SH treatment induced
cell cycle arrest at the S phase, as exhibited by the increased
cell number at the S phase and decreased cell number at the G0/G1
phase, suggesting that SH may act as a potential proliferation
inhibitor of melanoma cells. Consistently with the current results,
S. baumii can suppress the proliferation of human melanoma
A375 cells by promoting cell cycle arrest at the S phase (15). Moreover, protocatechualdehyde
isolated from S. gilvus can induce cell cycle arrest in HT29
and B16F10 cells (12,16). Overall, cell cycle arrest may be the
general regulatory mechanism of Sanghuang family members in
inhibiting the proliferation of cancer cells.
The cell cycle is highly orchestrated by a series of
events and signalling pathways, among which cyclins and CDKs are
essential in mediating the cell cycle (21). The activity of CDKs is mainly
regulated by binding of cyclin proteins/CDK inhibitors and
phosphorylation. High levels of p21, also known as CDK inhibitor
1A, can inhibit the functions of CyclinD-CDK4/6, CyclinE-CDK2 and
CyclinA-CDK2 complexes, leading to the interruption of the cell
cycle (20,22). Moreover, p21 participates in multiple
tumour suppressor pathways, including p53-dependent and
-independent mechanisms (20,22). p53
is a transcription factor that functions as a well-known tumour
suppressor and is involved in both the G0/G1 and G2/M cell cycle
phases (23). In our previous study,
it has been demonstrated that protocatechualdehyde isolated from
S. givlvus induces S-phase arrest in HT-29 cells via the
activation of the CyclinD1/E-CDK2 signalling pathway (12). To investigate the regulatory
mechanism of SH on cell cycle arrest in A375 cells, the expression
levels of cell cycle arrest-associated genes were assessed via
RT-qPCR, western blot and immunofluorescence analyses. p21
was significantly upregulated, and CyclinD-CDK4/6,
CyclinE-CDK2 and CyclinA-CDK2 complexes were
downregulated in A375 cells. No significant differences were
observed in the relative mRNA expression levels of p53 and
upstream genes (TGFB, Smad2 and Smad4) under SH
treatment, suggesting that SH may promote p21 expression
independently of p53 signalling. The expression levels of
E2F1 and DP1 were significantly decreased after 48 h
of treatment, and this may be caused by the downregulation of
CyclinA and CDK1. Therefore, the current results
suggested that SH treatment in A375 cells upregulated p21
expression to inhibit Cyclin-CDK complexes, resulting in cell cycle
arrest at the S phase.
Apoptosis is an evolutionarily conserved process
that serves an essential role in organism development and tissue
homeostasis (24). However, cancer
cells lose their ability to undergo apoptosis-induced death,
leading to uncontrolled proliferation. It has been previously
demonstrated that protocatechualdehyde isolated from S.
gilvus induces apoptosis in HT-29 and B16F10 cells (12,16). In
the present study, SH induced apoptosis in A375 cells. Under SH
treatment, the expression levels of pro-apoptotic genes, such as
Bad, Bik, Bid, Bim and Bak, decreased to varying
extents, and the anti-apoptotic gene Bcl-2 was significantly
inhibited. The Bcl-2 protein family is the key regulator of the
intrinsic apoptosis pathway (25).
Apoptosis is triggered by pro-apoptotic genes, such as Bim
and Bid. This pro-apoptotic chain of events is countered by
the anti-apoptotic genes Bcl-2, Bcl-xl and Bcl-w,
which can block apoptosis by binding and sequestering pro-apoptotic
proteins (25). Therefore, the
significant decrease of Bcl-2 may lead to apoptosis, even though
the expression levels of some pro-apoptotic genes were slightly
downregulated. These controversial but interesting results require
further research.
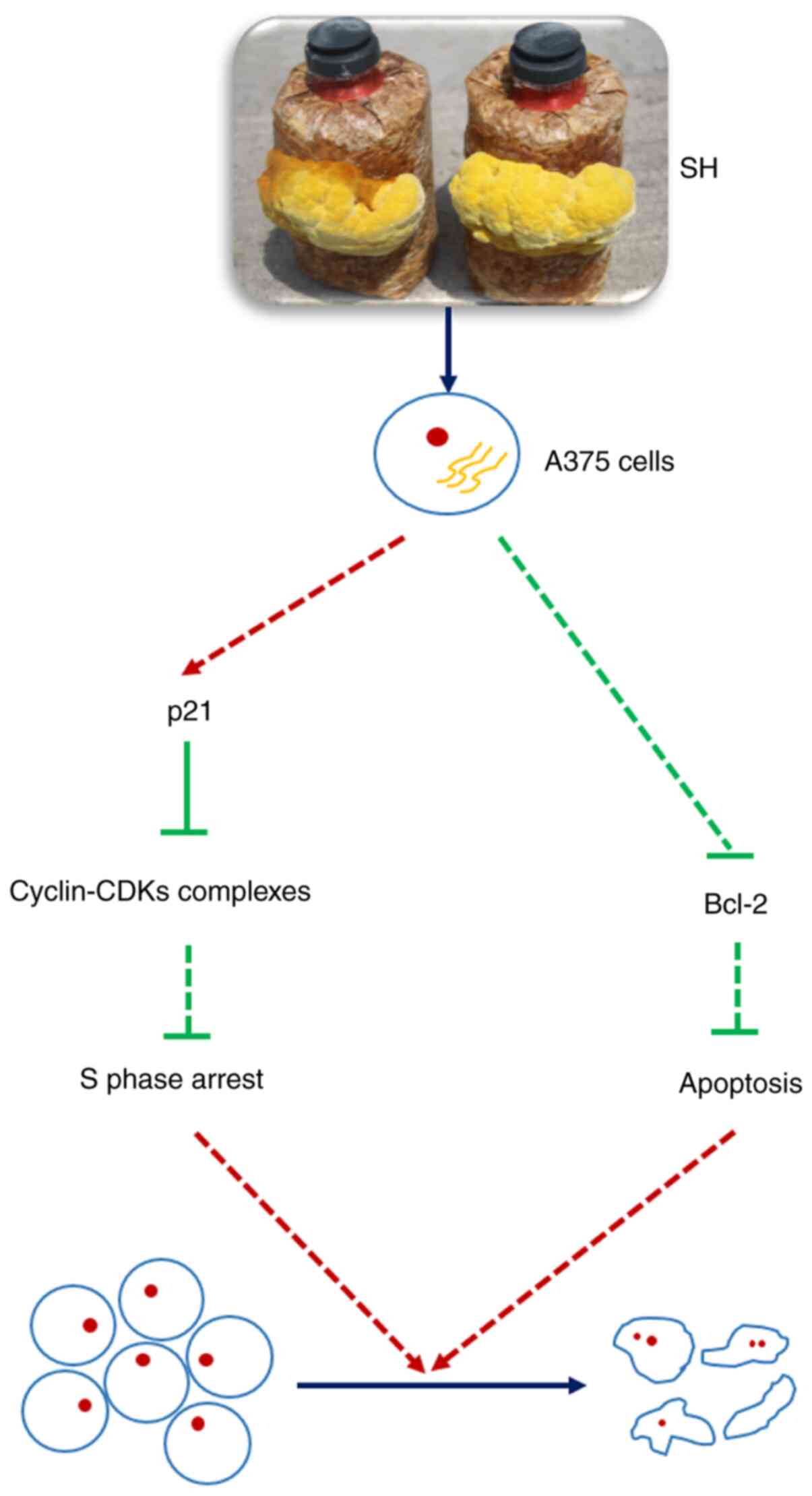
The current findings suggested that SH induced
S-phase arrest via p21-cyclin-CDKs complex pathways, and induced
apoptosis by inhibiting the expression levels of Bcl-2 (Fig. 6). Specific information about the
active component(s) in aqueous extracts of S. vaninii should
be obtained in future studies on isolation, identification and
illustration. In addition, the anti-proliferative effects of SH on
human melanoma cells in vivo should be further investigated
in future studies.
In conclusion, the present study revealed that the
aqueous extracts of S. vaninii fruiting body effectively
inhibited the proliferation of human melanoma cells by inducing
cell cycle arrest at the S phase, and this arrest may be mediated
by the activation of the p21-Cyclin-CDKs complexes signaling
pathways. Additionally, SH induced apoptosis of A375 cells via
significantly inhibiting the expression levels of the
anti-apoptotic gene Bcl-2 (Fig.
6). The current results suggested that S. vaninii
fruiting body may be used as a source of drug development for the
prevention and treatment of melanoma.
Acknowledgements
Not applicable.
Funding
The present study was supported financially by
Zhejiang Traditional Chinese Medicine Science and Technology
Project (grant no. 2020ZZ016), Science and Technology Department of
Zhejiang Province (grant nos. 2018C02003 and LGN18C170005),
Zhejiang Medical and Health Science and Technology Project (grant
no. 2018KY250) and Key Laboratory of Creative Agriculture, Ministry
of Agriculture and Rural Affairs.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TY, LL and JH contributed to the conception of the
study. SZ, YS, HS, WC, JZ and YL performed the experiments. JH and
LL revised the paper and gave final approval. TY, LL and JH are
responsible for confirming the authenticity of the data. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Olbryt M: Molecular background of skin
melanoma development and progression: Therapeutic implications.
Postepy Dermatol Alergol. 36:129–138. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Davis LE, Shalin SC and Tackett AJ:
Current state of melanoma diagnosis and treatment. Cancer Biol
Ther. 20:1366–1379. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lee JH, Cho SM, Kim HM, Hong ND and Yoo
ID: Immunostimulating activity of polysaccharides from mycelia of
Phellinus linteus grown under different culture conditions. J
Microbiol Biotechnol. 7:2–55. 1997.
|
|
4
|
Chen H, Tian T, Miao H and Zhao YY:
Traditional uses, fermentation, phytochemistry and pharmacology of
Phellinus linteus: A review. Fitoterapia. 113:6–26. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen W, Tan H, Liu Q, Zheng X, Zhang H,
Liu Y and Xu L: A review: The bioactivities and pharmacological
applications of Phellinus linteus. Molecules. 24:18882019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huo J, Zhong S, Du X, Cao Y, Wang W, Sun
Y, Tian Y, Zhu J, Chen J, Xuan L, et al: Whole-genome sequence of
Phellinus gilvus (mulberry Sanghuang) reveals its unique
medicinal values. J Adv Res. 24:325–335. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Meera CR, Janardhanan KK and Karunagaran
D: Antiproliferative and apoptotic activities of the medicinal
mushroom Phellinus rimosus (Agaricomycetes) on HCT116 human
colorectal carcinoma cells. Int J Med Mushrooms. 20:935–945. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang FF, Shi C, Yang Y, Fang Y, Sheng L
and Li N: Medicinal mushroom Phellinus igniarius induced
cell apoptosis in gastric cancer SGC-7901 through a
mitochondria-dependent pathway. Biomed Pharmacother. 102:18–25.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Im KH, Baek SA, Choi J and Lee TS:
Antioxidant, anti-melanogenic and anti-wrinkle effects of
Phellinus vaninii. Mycobiology. 47:494–505. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bae JS, Jang KH, Yim H and Jin HK:
Polysaccharides isolated from Phellinus gilvus inhibit
melanoma growth in mice. Cancer Lett. 218:43–52. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bae JS, Jang KH, Yim H, Park SC and Jin
HK: Inhibitory effects of polysaccharides isolated from
Phellinus gilvus on benzo(a)pyrene-induced forestomach
carcinogenesis in mice. World J Gastroenterol. 11:577–579. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhong S, Li YG, Ji DF, Lin TB and Lv ZQ:
Protocatechualdehyde induces S-phase arrest and apoptosis by
stimulating the p27KIP1-Cyclin A/D1-CDK2 and mitochondrial
apoptotic pathways in HT-29 Cells. Molecules. 21:9342016.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chao W, Deng JS, Li PY, Liang YC and Huang
GJ: 3,4-Dihydroxybenzalactone suppresses human non-small cell lung
carcinoma cells metastasis via suppression of epithelial to
mesenchymal transition, ROS-mediated PI3K/AKT/MAPK/MMP and NFκB
signaling pathways. Molecules. 22:5372017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu YJ, Lyu JL, Kuo YH, Chiu CY, Wen KC
and Chiang HM: The anti-melanogenesis effect of
3,4-dihydroxybenzalacetone through downregulation of melanosome
maturation and transportation in B16F10 and human epidermal
melanocytes. Int J Mol Sci. 22:28232021. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang Y, Zhang L, Chen Q, Lu WL and Li N:
Antitumor effects of extract of the oak bracket medicinal mushroom,
Phellinus baumii (Agaricomycetes), on human melanoma cells
A375 in vitro and in vivo. Int J Med Mushrooms. 22:197–209. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong S, Jin Q, Yu T, Zhu J and Li Y:
Phellinus gilvus derived protocatechualdehyde induces G0/G1
phase arrest and apoptosis in murine B16 F10 cells. Mol Med Rep.
21:1107–1114. 2020.PubMed/NCBI
|
|
17
|
Huo J, Sun Y, Zhong S, Li Y, Yang R, Xia
L, Wang J, Zhang M and Zhu J: Safety evaluation of aqueous extracts
of Sanghuangporus vaninii fruiting body in Sprague-Dawley
rats. Food Sci Nutr. 8:5107–5113. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Harvey AL, Edrada-Ebel R and Quinn RJ: The
re-emergence of natural products for drug discovery in the genomics
era. Nat Rev Drug Discov. 14:111–129. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Moussa RS, Park KC, Kovacevic Z and
Richardson DR: Ironing out the role of the cyclin-dependent kinase
inhibitor, p21 in cancer: Novel iron chelating agents to target p21
expression and activity. Free Radic Biol Med. 133:276–294. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Diaz-Moralli S, Tarrado-Castellarnau M,
Miranda A and Cascante M: Targeting cell cycle regulation in cancer
therapy. Pharmacol Ther. 138:255–271. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Al Bitar S and Gali-Muhtasib H: The role
of the cyclin dependent kinase inhibitor p21cip1/waf1 in targeting
cancer: Molecular mechanisms and novel therapeutics. Cancers
(Basel). 11:E14752019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Golias CH, Charalabopoulos A and
Charalabopoulos K: Cell proliferation and cell cycle control: A
mini review. Int J Clin Pract. 58:1134–1141. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du Toit A: Cell death: Balance through a
bivalent regulator. Nat Rev Mol Cell Biol. 14:546–547. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Singh R, Letai A and Sarosiek K:
Regulation of apoptosis in health and disease: The balancing act of
BCL-2 family proteins. Nat Rev Mol Cell Biol. 20:175–193. 2019.
View Article : Google Scholar : PubMed/NCBI
|