Introduction
Ovarian cancer (OC) is a common tumor of the female
reproductive system, and ~80% of patients are diagnosed in the
first instance with advanced stage (stages III–IV) (1). In 2018, there were 22,240 newly
diagnosed OC cases in the United States alone, and 14,070 patients
died due to OC (2). The 5-year
survival rate of OC is only 40–45% (2). It is therefore crucial to determine the
underlying mechanisms of OC development and progression in order to
improve diagnosis and treatment.
Long non-coding RNAs (lncRNAs) are non-coding RNAs
of ≥200 nucleotides in length, which regulate a wide range of
physiological and pathological processes, including gene
transcription and translation, chromatin modification, cell cycle
progression, cell differentiation, carcinogenesis and cancer
progression (3). lncRNAs are
crucially involved in the progression of various types of cancer
(4–6). For example, small nucleolar RNA host
gene 1 is highly expressed in non-small cell lung cancer (NSCLC)
tissues and cells (4), and lncRNA
activated by transforming growth factor-beta inhibits the
proliferation and induces the cell cycle arrest of gastric cancer
cells (5). The lncRNA GATA6
antisense RNA1 (GATA6-AS1), whose gene is located on chromosome
18q11.2, functions as a tumor suppressor in several types of
cancer, such as gastric cancer (7).
However, to the best of our knowledge, the role and regulatory
mechanism of GATA6-AS1 in OC remain unknown.
MicroRNAs (miRNAs/miRs) are endogenous non-coding
RNA molecules of ~22 nucleotides in length, which can repress the
expression of target genes at the post-transcriptional level and
largely participate in multiple biological processes (8). miR-19a has been implicated in the
promotion of certain malignant biological behaviors, including the
proliferation, metastasis and drug resistance, of cancer cells in
thyroid cancer, NSCLC, colorectal cancer, osteosarcoma and OC
(9–16). However, the role of miR-19a-5p in OC
has not been extensively studied.
Tet methylcytosine dioxygenase 2 (TET2) is a member
of the DNA demethylase TET protein family, which promotes DNA
demethylation via conversion of 5-methylcytosine (5mC) into
5-hydroxymethylcytosine (5hmC). A decrease in 5-hmC levels can be
used as an epigenetic marker of OC progression (17–19).
However, the role of TET2 in OC remains unclear.
This study aimed to clarify the expression of
GATA6-AS1 in OC, its clinical significance and mechanism. The
findings may indicate that GATA6-AS1 has anticancer effects and
could provide potential novel therapeutic targets for the treatment
of OC.
Materials and methods
Tissue samples
A total of 40 patients with OC were recruited
between June 2017 and March 2019 at The Affiliated Yantai
Yuhuangding Hospital of Qingdao University (Yantai, China).
Patients were aged between 20 and 69 years (mean age, 50.32±7.28
years). A total of 26 cases had a tumor <3 cm in diameter and 32
cases were serous and 8 cases were mucinous. According to the
International Federation of Gynecology and Obstetrics staging
system, 5 cases were stage I, 7 cases were stage II and 28 cases
were stage III or IV. A total of 29 cases had lymph node
metastases, whereas 11 did not. The present study was approved by
the Research Ethics Committee of Yuhuangding Hospital of Qingdao
University (approval no. 20161228A) and written informed consent
was obtained from all participants. The cancerous and adjacent
normal ovarian tissues were obtained during surgery and were
immediately stored in liquid nitrogen until RNA extraction.
Adjacent normal ovarian tissues were taken from the same patient
>2 cm from the edge of the primary tumor.
Cell culture
The human ovarian surface epithelial HOSEPiC cell
line was purchased from ScienCell Research Laboratories. The OC
SKOV-3 and ES-2 cell lines were purchased from the American Type
Culture Collection. The OC COC1 and A2780 cell lines were purchased
from the China Center for Type Culture Collection. Cells were
cultured in RPMI-1640 medium (Invitrogen; Thermo Fisher Scientific,
Inc.) supplemented with 10% FBS (Thermo Fisher Scientific, Inc.)
and 100 U/ml penicillin, 100 µg/ml streptomycin (Hyclone; Cytiva)
and placed at 37°C in a humidified incubator containing 5%
CO2. The medium was replaced every 2–3 days. When cells
reached 70–80% confluence, they were routinely sub-cultured using
0.25% trypsin (Roche Diagnostics).
Cell transfection
Full-length GATA6-AS1 sequence lacking a poly-A tail
was synthesized based on the sequence obtained from the National
Center for Biotechnology Information database and sub-cloned into
pcDNA3.1 by Shanghai GenePharma Co., Ltd. Empty plasmid, GATA6-AS1
overexpression plasmid (pcDNA3.1-GATA6-AS1), negative control short
hairpin-RNA (sh-NC), shRNA targeting GATA6-AS1 (sh-GATA6-AS1),
miRNA control (miR-NC, 5′-UCACAACCUCCUAGAAAGAGUAGA-3′), miR-19a-5p
mimics (miR-19a-5p, 5′-AGTTTTGCATAGTTGCACTACA-3′), inhibitors
control (In-NC, 5′-CACUGGUACAAGGGUUGGGAGA-3′), miR-19a-5p inhibitor
(miR-19a-5p in, 5′-TGTAGTGCAACTATGCAAAACT-3′), TET2 overexpression
plasmid (pcDNA3.1-TET2) and shRNA targeting TET2 (sh-TET2) were
purchased from Shanghai GenePharma Co., Ltd.
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to transfect cells according to the
manufacturer's protocol. After 36 h transfection, overexpression or
knockdown efficiency was examined using reverse transcription
quantitative (RT-q)PCR.
RT-qPCR
Total RNA was extracted from cell lines or tissues
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and was reverse transcribed into cDNA using a
RevertAid™ First Strand DNA Synthesis kit (Thermo Fisher
Scientific, Inc.). QuantiFast SYBR-Green PCR kit (Roche
Diagnostics) was used to perform qPCR on an ABI 7300 Real-time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling protocol was as follows: Initial denaturation at 95°C
for 5 min, followed by 45 repeats of a three-step cycling program
consisting of 10 sec at 95°C (denaturation), 10 sec at 60°C (primer
annealing) and 10 sec at 72°C (elongation), and a final extension
step for 10 min at 72°C. The sequences of the primers were as
follows: GATA6-AS1, forward 5′-ACCACAACCACTACCTTATGGCGT-3′, reverse
5′-TGCCATCTGGACTGCTGGACAATA-3′; miR-19a-5p, forward
5′-GTTTGCTGGGAAGGCAAAG-3′, reverse 5′-TGTTTTGCTGGGAAGGCAAA-3′; U6,
forward 5′-CGCTTCGGCAGCACATATAC-3′, reverse
5′-TTCACGAATTTGCGTGTCAT-3′; TET2, forward
5′-GGACTGAGCTGCTGAATTCAACT-3′, reverse
5′-CCTCAACATGGTTGGTTCTATCC-3′; and GAPDH, forward
5′-TGTCCGTCGTGGATCTGA-3′ and reverse 5′-TTGCTGTTGAAGTCGCAGGAG-3′.
The relative expression levels of GATA6-AS1, miR-19a-5p and TET2
were normalized to endogenous controls GAPDH or U6 and were
expressed as 2−ΔΔCq (20).
Cell Counting Kit-8 (CCK-8) assay
The density of cell suspensions was adjusted to
1×104 cells/ml, and 100 µl of cell suspension was added
to each well of a 96-well plate. After 12, 24, 48, 72 and 96 h, 10
µl CCK-8 reagent (Beyotime Institute of Biotechnology) was added to
each well, and the cells were incubated for 1 h at 37°C. The
absorbance was read at 450 nm using a microplate reader (BioTek
Instruments, Inc.).
Transwell assay
OC cells were harvested using 0.25% trypsin,
centrifuged at 10,000 × g for 15 min at room temperature and
resuspended in serum-free medium. In the invasion assay only the
membranes of the Transwell chambers (Corning, Inc.) were pre-coated
with Matrigel (1:10; BD Biosciences) to mimic the extracellular
matrix. The cell suspension (200 µl) containing ~5×104
cells was added to the upper chamber of the Transwell chamber
whereas the lower chamber was filled with 400 µl medium
supplemented with 10% FBS. After incubation at 37°C for 24 h, cells
that had not migrated or invaded the lower chamber were removed.
The migrated or invaded cells were fixed with 4% paraformaldehyde
for 10 min at room temperature and stained with 0.5% crystal violet
for 5 min at room temperature. Chambers were subsequently immersed
in tap water and cells were visualized and counted using a light
microscope (Nikon Corporation) at ×200 magnification for five
random fields.
Bioinformatics analysis
The GEPIA database (http://gepia.cancer-pku.cn/) was used to assess the
expression of GATA6-AS1. LncBase Predicted version 2 (http://carolina.imis.athena-innovation.gr/index.php?r=lncbasev2)
and TargetScan online websites (http://www.targetscan.org/vert_71/) predicted
potential binding sites between GATA6-AS1 and miR-19a-5p, and
between miR-19a-5p and the 3′UTR of TET2.
Dual-luciferase reporter gene
assay
Bioinformatics analysis predicted potential binding
sites between GATA6-AS1 (5′-UUUAUGUUGGUUUAAUUUCGAAAAUAAACU-3′) and
miR-19a-5p, and between miR-19a-5p and the 3′UTR of TET2
(5′-ACUGGAGUCUCAUUUGCAAAACC-3′). The predicted fragment was
amplified and inserted into pmirGLO Vectors (Promega Corporation)
to construct the wild-type (WT) reporter vector
pmirGLO-GATA6-AS1-WT or pmirGLO-TET2-WT. The mutant (MUT) reporter
vectors were constructed using a GeneArt™ Site-Directed Mutagenesis
system (Thermo Fisher Scientific, Inc.). The reporter vectors and
miR-19a-5p mimics or miR-NC were co-transfected into OC cells, and
cells were subsequently cultured for 48 h. Luciferase activity of
the cells in each group was then measured using a dual-luciferase
reporter assay system (Promega Corporation). Renilla
luciferase activity was normalized to firefly luciferase
activity.
RNA immunoprecipitation (RIP)
assay
RIP assay was performed using a Magna RIP
RNA-Binding Protein Immunoprecipitation kit (EMD Millipore)
according to the manufacturer's protocol. In brief, cells were
centrifuged at 1,500 × g and 4°C for 10 min and incubated with RIP
lysis buffer, and the obtained cellular lysates were next probed
with magnetic beads conjugated with a human anti-AGO2 antibody
(cat. no. ab5072; rabbit polyclonal antibody; Abcam) or control IgG
(cat. no. 03-110; EMD Millipore) at 4°C for 6 h. The cell lysates
were then treated with proteinase K buffer (150 µl) at 55°C for 30
min to digest the protein. The magnetic beads were repeatedly
washed with RIP washing buffer to remove non-specific adsorption as
much as possible. The expression of GATA6-AS1 and miR-19a-5p in
RIP-derived immunoprecipitated RNA was measured by RT-qPCR.
Western blotting
Cells were lysed with RIPA buffer (Thermo Fisher
Scientific, Inc.) on ice, and the supernatant was collected after
high-speed centrifugation (at 12,000 × g for 15 min at 4°C). The
protein concentration was quantified using a BCA assay kit
(Beyotime Institute of Biotechnology). After mixing with loading
buffer, the samples were heated in a water bath at 100°C for 10 min
to denature the proteins. Proteins (30 µg per lane) were separated
by SDS-PAGE on 10% gels and transferred onto PVDF membranes (EMD
Millipore). Membranes were blocked with 5% skimmed milk in TBST
with 5% BSA at room temperature. Membranes were then incubated with
primary antibodies against TET2 (cat. no. ab243323; 1:500; Abcam)
and GAPDH (cat. no. ab181602; 1:2,000; Abcam) at 4°C for 8 h.
Membranes were washed with TBST and incubated with secondary
antibody (cat. no. ab150077; 1:1,000; Abcam) at room temperature
for 1 h. Enhanced chemiluminescence reagent (EMD Millipore) was
used to detect the signal on the membrane. Quantity One software
v.4.62 (Bio-Rad Laboratories, Inc.) was used for densitometry
analysis.
Statistical analysis
GraphPad Prism version 8 (GraphPad Software, Inc.)
and SPSS version 16.0 (SPSS Inc.) were used to analyze the data.
Data were presented as the means ± standard deviation. Whether the
data were normally distributed or not was examined using the
Kolmogorov-Smirnov test. For normally distributed data, an unpaired
or paired t-test was used to compare data between two 2 groups.
Comparisons among ≥3 groups were conducted with one-way ANOVA
followed by Tukey's post hoc test. For data that were not normally
distributed, comparisons between two groups were performed using
paired sample Wilcoxon signed-rank test. Comparison between OC and
adjacent normal tissue samples from patients with OC was performed
using a paired t-test, while comparison between experimental and
control groups was performed using an unpaired t-test. Correlation
analyses among the expression levels of GATA6-AS1, miR-19a-5p and
TET1 were performed using Pearson's correlation coefficient.
P<0.05 was considered to indicate a statistically significant
difference.
Results
GATA6-AS1 is downregulated in OC
tissues
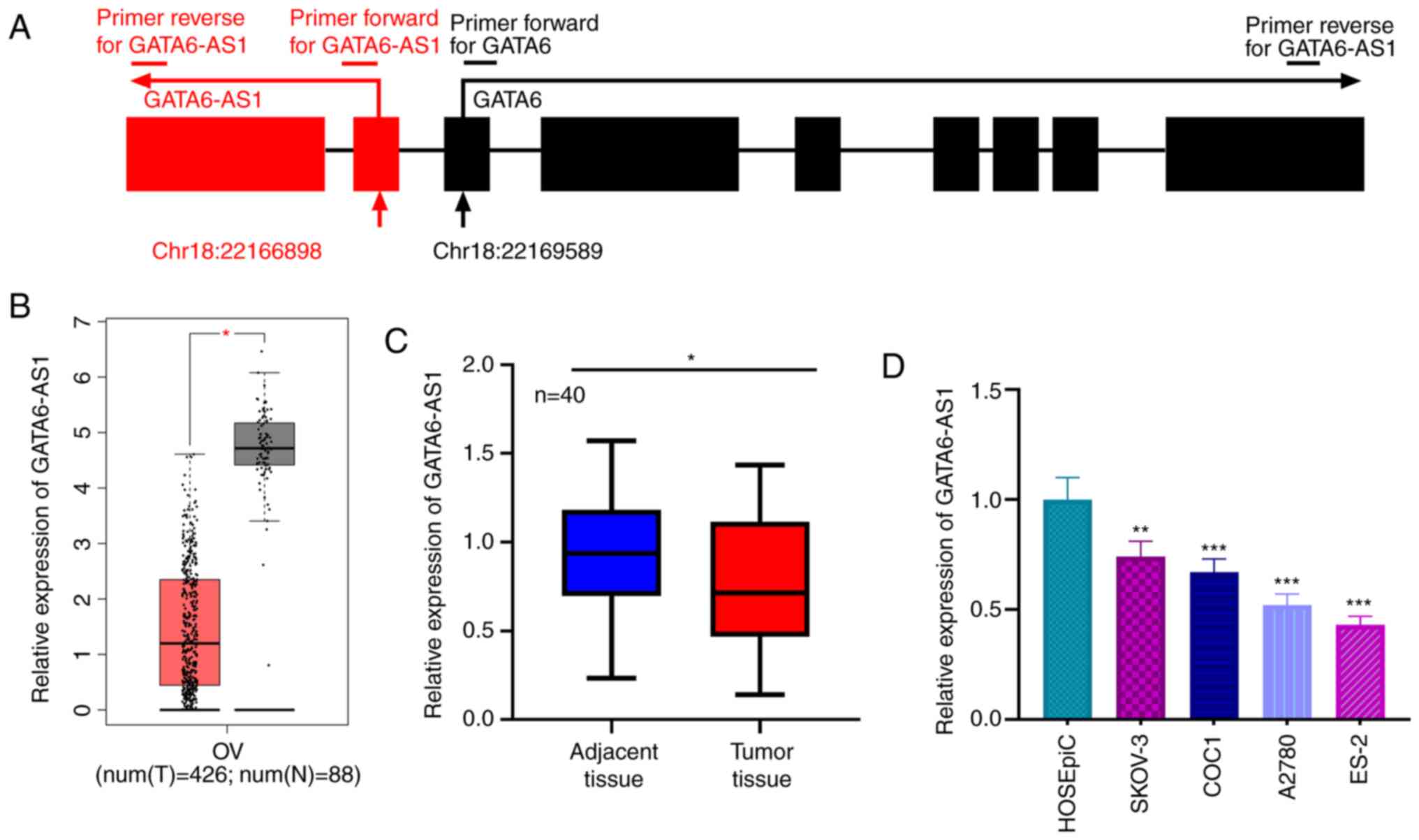
GATA6-AS1 has a 1,788-nucleotide long sequence
(accession no. NR_102763.1), whose gene is located on chromosome
18, next to the sequence for GATA6 (Fig.
1A). The GEPIA database was used to assess the expression of
GATA6-AS1, and its expression was found to be downregulated in OC
tissues relative to normal ovarian tissues (Fig. 1B). Consistent with this result, the
expression of GATA6-AS1 in 40 pairs of OC and adjacent ovarian
tissues was detected by RT-qPCR. The results demonstrated that
GATA6-AS1 was significantly downregulated in OC tissues compared
with adjacent normal tissues (Fig.
1C). Additionally, compared with HOSEpiC cells, GATA6-AS1
expression was also significantly lower in OC cell lines (SKOV-3,
COC1, A2780 and ES-2; Fig. 1D).
These data suggested that GATA6-AS1 may function as a tumor
suppressor in OC.
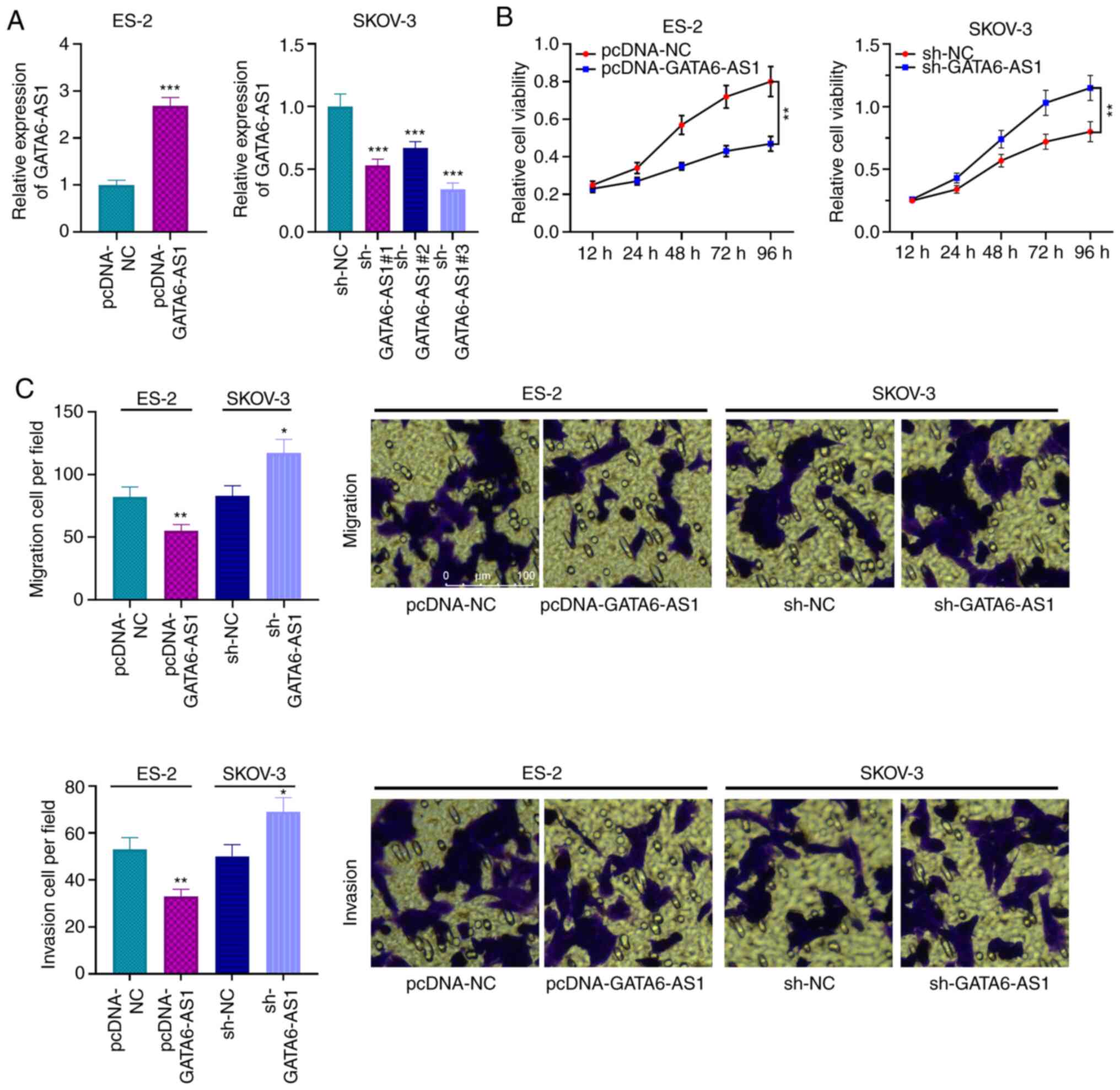
GATA6-AS1 inhibits the proliferation
and migratory and invasive abilities of OC cells
The results indicated that GATA6-AS1 expression in
OC cell lines (SKOV-3, COC1, A2780 and ES-2) was significantly
lower compared with HOSEpiC cells (Fig.
1D). Among these four types of cell, GATA6-AS1 expression was
the lowest in ES2 cells, while it was the highest in SKOV-3 cells.
Subsequently, the overexpression GATA6-AS1 plasmid was transfected
into ES-2 cells whereas GATA6-AS1 expression was knocked down in
SKOV-3 cells using shRNA. The results from RT-qPCR confirmed that
these transfections were successful (Fig. 2A). Furthermore, the results from
CCK-8 and Transwell assays showed that GATA6-AS1 overexpression
inhibited the proliferation and migratory and invasive abilities of
ES-2 cells compared with the control group, whereas GATA6-AS1
knockdown had the opposite effects in SKOV-3 cells (Fig. 2B and C). These findings indicated
that GATA6-AS1 may inhibit the malignant biological behaviors of OC
cells.
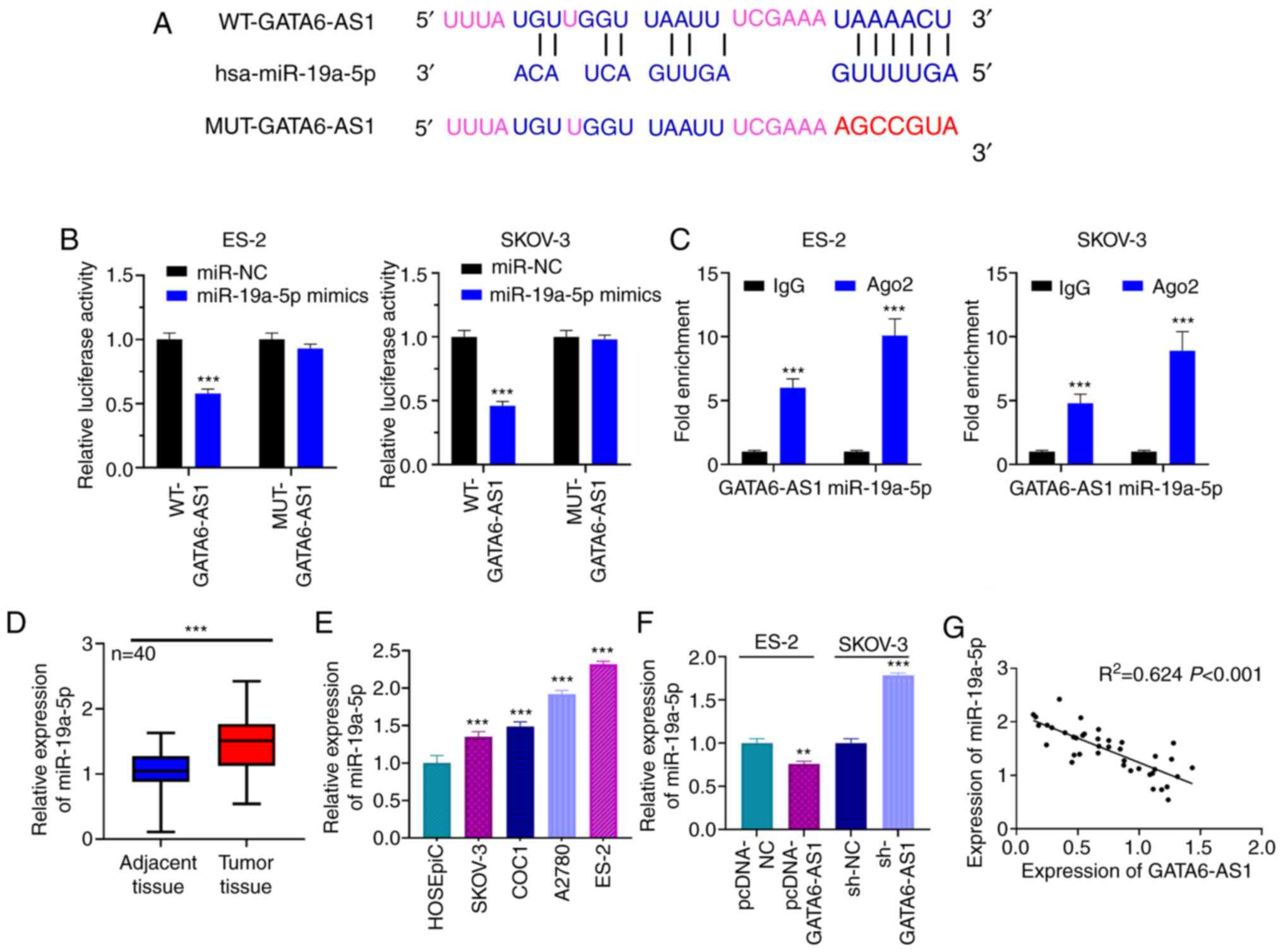
GATA6-AS1 sponges miR-19a-5p
Bioinformatics analysis was performed using LncBase
Predicted version 2, and it was predicted that GATA6-AS1 contained
a potential binding site for miR-19a-5p (Fig. 3A). The results from dual luciferase
reporter gene assay showed that the luciferase activity of the
WT-GATA6-AS1 reporter was decreased by miR-19a-5p mimics, whereas
that of MUT-GATA6-AS1 reporter was not significantly affected
(Fig. 3B). Furthermore, results from
RIP assay suggested that GATA6-AS1 and miR-19a-5p may directly
interact with each other in the immunoprecipitate containing Ago2
(Fig. 3C). In addition, miR-19a-5p
expression was assessed in OC and normal adjacent tissues using
RT-qPCR. The results demonstrated that miR-19a-5p expression was
significantly higher in OC tissues compared with normal adjacent
tissues (Fig. 3D). Furthermore,
miR-19a-5p expression was significantly increased in the four OC
cell lines compared with HOSEpiC cells (Fig. 3E). It was also demonstrated that
GATA6-AS1 upregulation in ES-2 cells could significantly inhibit
miR-19a-5p expression, whereas GATA6-AS1 knockdown in SKOV-3 cells
resulted in miR-19a-5p upregulation (Fig. 3F). Pearson's correlation analysis
showed that miR-19a-5p expression was negatively correlated with
GATA6-AS1 expression in OC tissue samples (Fig. 3G), suggesting that miR-19a-5p may be
a downstream target of GATA6-AS1.
TET2 is a target gene of miR-19a-5p in
OC
The candidate targets of miR-19a-5p were predicted
using TargetScan, and TET2 was found to be a candidate target of
miR-19a-5p. The binding site is presented in Fig. 4A. The results from RT-qPCR indicated
that miR-19a-5p expression was significantly increased in ES-2 and
SKOV-3 following transfection with miR-19a-5p mimics (Fig. S1). Furthermore, dual luciferase
reporter gene assay showed that the luciferase activity of WT-TET2
reporter was decreased by miR-19a-5p mimics; however, miR-19a-5p
mimics had no effect on the luciferase activity of MUT-TET2
reporter (Fig. 4B). In addition,
results from RT-qPCR and western blotting demonstrated that TET2
expression in OC tissues and cells was significantly lower compared
with that in adjacent tissues and normal ovarian epithelial cell
(Fig. 4C-E). Compared with miR-NC,
the expression level of miR-19a-5p was significantly increased in
ES-2 cells transfected with miR-19a-5p mimics (Fig. S1A). The expression of miR-19a-5p was
significantly decreased in SKOV-3 following transfection with
miR-19a-5p inhibitors compared with inh-NC (Fig. S2). In addition, transfection with
miR-19a-5p mimics significantly decreased TET2 mRNA and protein
expression in ES-2 cells, whereas miR-19a-5p inhibitors
significantly increased TET2 mRNA and protein expression in SKOV-3
cells (Fig. 4F and G). Pearson's
correlation analysis demonstrated that miR-19a-5p expression was
negatively correlated with TET2 mRNA expression in OC tissue
samples (Fig. 4H). These findings
indicated that miR-19a-5p could target and negatively regulate TET2
expression in OC.
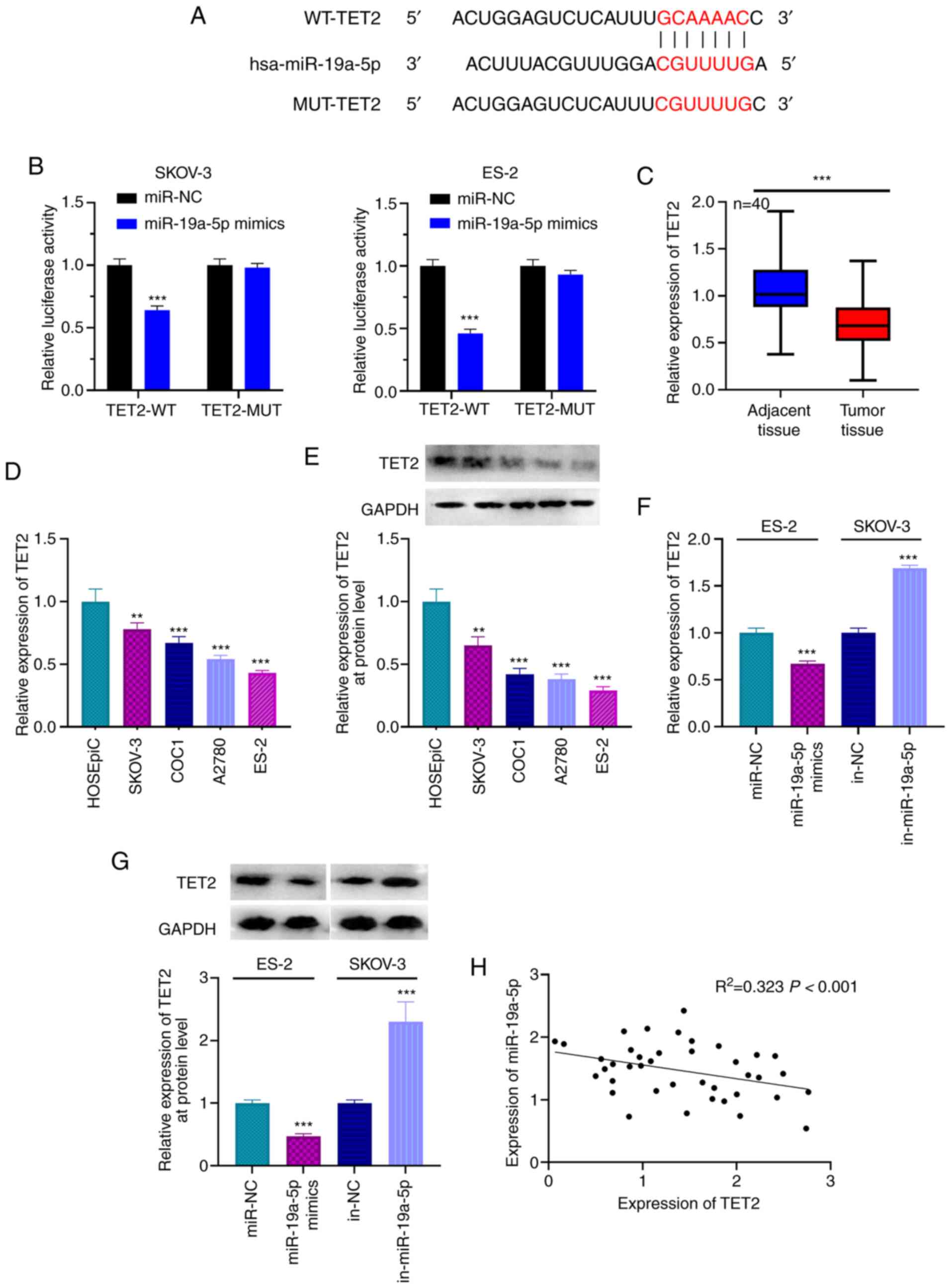 | Figure 4.TET2 is the target gene of miR-19a-5p
in OC cells. (A) TargetScan analysis predicted the presence of a
binding site between the 3′UTR of TET2 and miR-19a-5p. (B) Dual
luciferase reporter gene assay was used to confirm the binding
relationship between miR-19a-5p and TET2. (C) Expression of TET2 in
OC and adjacent normal tissues was detected using RT-qPCR. (D)
Expression of TET2 in cell lines was detected using RT-qPCR. (E)
Expression of TET2 in cell lines was detected using western
blotting. (F) miR-19a-5p mimics and inhibitors were transfected
into ES-2 and SKOV-3 cells, respectively, and the expression levels
of TET2 was detected using RT-qPCR. (G) miR-19a-5p mimics and
inhibitors were transfected into ES-2 and SKOV-3 cells,
respectively, and the expression levels of TET2 was detected using
western blotting. (H) Relationship between expression of miR-19a-5p
and TET2 was examined using Pearson's correlation analysis. n=40.
All experiments were performed in triplicate. **P<0.01 and
***P<0.001. miR, microRNA; OC, ovarian cancer; TET2, ten eleven
translocation 2; UTR, untranslated region; RT-qPCR, reverse
transcription-quantitative PCR; MUT, mutant; WT, wild-type; NC,
negative control; in, inhibitor. |
GATA6-AS1 alters OC cell phenotype via
the miR-19a-5p/TET2 axis
To determine the role of the
GATA6-AS1/miR-19a-5p/TET2 axis in the progression of OC, ES-2 cells
were transfected with pcDNA-NC, pcDNA-GATA6-AS1, pcDNA-GATA6-AS1 +
miR-19a-5p mimics or pcDNA-GATA6-AS1 + sh-TET2; and SKOV-3 cells
were transfected with sh-NC, sh-GATA6-AS1, sh-GATA6-AS1 +
miR-19a-5p inhibitors or pcDNA-GATA6-AS1 + pcDNA-TET2. The results
from RT-qPCR and western blotting demonstrated that GATA6-AS1
overexpression resulted in increased TET2 mRNA and protein
expression in ES-2 cells, and that this effect was attenuated by
miR-19a-5p mimics or sh-TET2. Furthermore, GATA6-AS1 knockdown
decreased TET2 expression in SKOV-3 cells, but this inhibitory
effect was partially reversed by co-transfection with miR-19a-5p
inhibitor or pcDNA-TET2 (Fig. 5A and
B). The results from CCK-8 and Transwell assays demonstrated
that GATA6-AS1 overexpression significantly inhibited the
proliferation and migratory and invasive abilities of ES-2 cells;
however, this inhibitory effect was partially compensated by
transfection with miR-19a-5p mimics or sh-TET2 (Fig. 5B-D). Furthermore, GATA6-AS1 knockdown
promoted the malignant properties of SKOV-3 cells, which was
partially attenuated by transfection with miR-19a-5p inhibitor or
pcDNA-TET2 (Fig. 5B-D). These
results suggested that GATA6-AS1 may regulate OC progression via
the miR-19a-5p/TET2 axis.
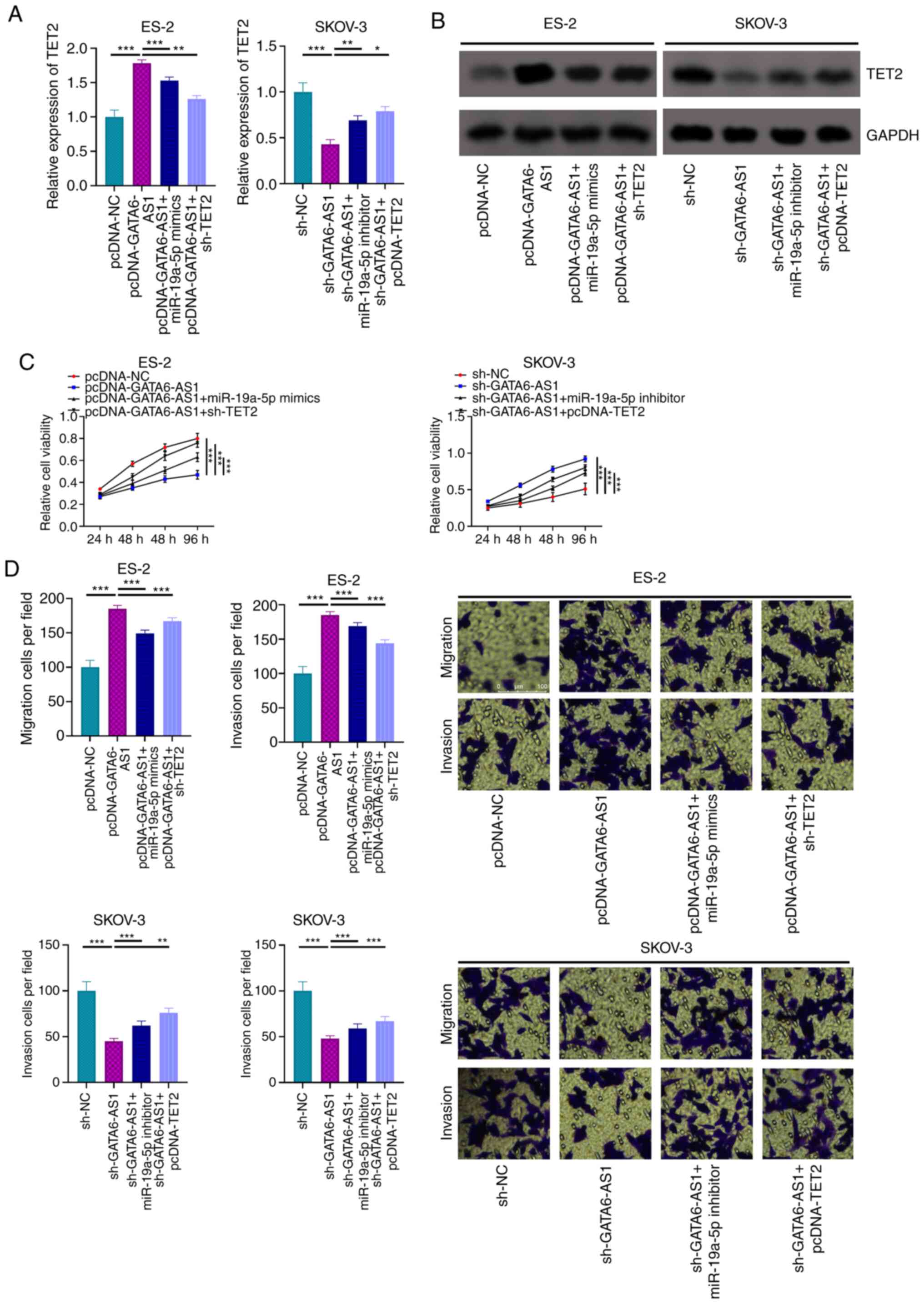 | Figure 5.GATA6-AS1 regulates OC via a
miR-19a-5p/TET2 axis. ES-2 cells were transfected with pcDNA-NC,
pcDNA-GATA6-AS1, pcDNA-GATA6-AS1 + miR-19a-5p mimics or
pcDNA-GATA6-AS1 + sh-TET2. SKOV-3 cells were transfected with
sh-NC, sh-GATA6-AS1, sh-GATA6-AS1 + miR-19a-5p inhibitors or
pcDNA-GATA6-AS1 + pcDNA-TET2. Subsequently, the mRNA and protein
expression levels of TET2 in OC cells were detected using (A)
RT-qPCR and (B) western blotting, respectively. (C) Proliferation
of OC cells was assessed using a Cell Counting Kit-8 assay. (D)
Transwell assays were used to evaluate OC cell migratory and
invasive abilities. All experiments were performed in triplicate.
*P<0.05, **P<0.01 and ***P<0.001. GATA6-AS1, GATA6
antisense RNA 1; OC, ovarian cancer; miR, microRNA; TET2, ten
eleven translocation 2; RT-qPCR, reverse transcription-quantitative
PCR; NC, negative control; sh, short hairpin. |
Discussion
lncRNAs are widely expressed in various types of
human tumor, and dysregulation of certain lncRNAs has been shown to
be associated with tumor progression, highlighting their potential
roles as biomarkers and/or therapeutic targets (21,22).
GATA6-AS1 expression has been reported to be downregulated in
gastric cancer, where it decreases frizzled class receptor 4 (FZD4)
expression by recruiting zeste homolog 2 and inducing
trimethylation at lysine 27 of histone H3 of the FZD4 promoter
region, inhibiting therefore the Wnt/β-catenin signaling pathway
and preventing epithelial-mesenchymal transition (7). Furthermore, downregulated expression of
GATA6-AS1 activates the PI3K/AKT/Snail signaling pathway via
regulation of miR-582/FOXO3 axis and subsequently promote the
proliferation and metastasis of gastric cancer cells (23). The present study demonstrated that
GATA6-AS1 expression was decreased in OC and that GATA6-AS1
significantly inhibited the proliferation and migratory and
invasive abilities of OC cells, confirming that GATA6-AS1 may be a
tumor-suppressive lncRNA in OC.
MiRNAs have been involved in the regulation of cell
proliferation, differentiation and apoptosis, amongst other cell
biological processes (24–29). Both miR-19a-3p and miR-19a-5p have
been reported to be crucial in the promotion or inhibition of
cancer progression. For example, miR-19a-3p was reported to promote
the proliferation of hepatocellular carcinoma cells via regulation
of the PIK3IP1/AKT pathway (30);
however, another study reported that miR-19-3p induces colorectal
cancer cell apoptosis by repressing the expression of Fas cell
surface death receptor (31).
Furthermore, miR-19a-5p expression is significantly decreased in
NSCLC, and miR-19a-5p can suppress the proliferation, migration and
invasion of NSCLC cells (28). In
the present study, miR-19a-5p expression was significantly
increased in OC tissues and cells compared with adjacent normal
tissues and normal cells, respectively. In addition, functional
experiments showed that miR-19a-5p reversed the tumor-suppressive
effect of GATA6-AS1, suggesting that miR-19a-5p may act as an
oncomiR involved in the promotion of OC progression. GATA6-AS1 was
also identified as a competitive endogenous RNA of miR-19a-5p,
which could sponge and reduce the expression of miR-19a-5p. This
demonstration may partly explain the mechanism leading to
miR-19a-5p dysregulation in OC.
TET proteins are a class of α-ketoglutarates and
Fe2+-dependent dioxygenases that consist of three
members, TET1, TET2 and TET3. All three TET proteins are capable of
transforming 5mC into 5hmC, and mutation or dysregulation of TET
contribute to tumorigenesis in several types of cancer (32). TET2 is the second most frequently
mutated gene in hematopoiesis, and TET2 mutations were demonstrated
to drive blood cell tumorigenesis (33). Defects in TET2 enzyme activity
contribute to the incidence of myeloid cancer, and measurement of
5-hmC levels in myeloid cancer is a promising strategy for
diagnostic and prognostic evaluation (34). As previously reported, TET2
expression is downregulated in OC and is crucial in inhibiting
cancer progression (19). TET2
regulates 5mC oxidation pathways to activate or inhibit the
expression of downstream genes through modulating transcription
repressors or associated activators (19). However, to the best of our knowledge,
there are only a few reports focusing on the underlying mechanism
of TET2 dysregulation in OC. Notably, it has been reported that
miR-19a-5p can negatively regulate the expression of TET2 in
glioblastoma (13). In the present
study, a similar regulatory mechanism between miR-19a-5p and TET2
was demonstrated in OC. In addition, GATA6-AS1 positively regulated
the expression of TET2 via repression of miR-19a-5p in OC cells.
The results from functional experiments suggested that the
tumor-suppressive role of GATA6-AS1 in OC was partly dependent on
its regulatory function on TET2. These results not only clarify the
mechanism by which GATA6-AS1 could repress OC progression, but also
propose a reasonable explanation for TET2 dysregulation in OC.
In summary, the present study demonstrated that
GATA6-AS1 expression is downregulated in OC tissues and cells.
Furthermore, the results from functional experiments showed that
GATA6-AS1 could inhibit the proliferation and migratory and
invasive abilities of OC cells via regulating the miR-19a-5p/TET2
axis. Restoration of GATA6-AS1 may therefore be considered as an
effective strategy for OC treatment. In future studies, in
vivo experiments should be performed to confirm the present
findings. In addition, the small cohort size and lack of survival
analysis were a limitation to the present study. The role of
GATA6-AS1 as a prognostic biomarker should therefore be
investigated in a larger cohort.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HX, XW and HZ conceived and designed the
experiments. HX, XW, YZ and WZ performed the experiments. YZ and WZ
performed statistical analyses. HX, XW and HZ wrote the paper. WZ
and HJZ confirm the authenticity of all the raw data. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
This study was approved by the Ethics Review Board
of The affiliated Yantai Yuhuangding Hospital of Qingdao University
(approval no. 20161228A).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stewart C, Ralyea C and Lockwood S:
Ovarian Cancer: An integrated review. Semin Oncol Nurs. 35:151–156.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Trabert B, DeSantis CE, Miller
KD, Samimi G, Runowicz CD, Gaudet MM, Jemal A and Siegel RL:
Ovarian cancer statistics, 2018. CA Cancer J Clin. 68:284–296.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Peng WX, Koirala P and Mo YY:
LncRNA-mediated regulation of cell signaling in cancer. Oncogene.
36:5661–5667. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li Z, Lu Q, Zhu D, Han Y, Zhou X and Ren
T: Lnc-SNHG1 may promote the progression of non-small cell lung
cancer by acting as a sponge of miR-497. Biochem Biophys Res
Commun. 506:632–640. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lei K, Liang X, Gao Y, Xu B, Xu Y, Li Y,
Tao Y, Shi W and Liu J: Lnc-ATB contributes to gastric cancer
growth through a miR-141-3p/TGFβ2 feedback loop. Biochem Biophys
Res Commun. 484:514–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ge YW, Gao HM and Wang ZM: Advances in
study of genus curcuma. Zhongguo Zhong Yao Za Zhi. 32:2461–2467.
2007.(In Chinese). PubMed/NCBI
|
|
7
|
Li ZT, Zhang X, Wang DW, Xu J, Kou KJ,
Wang ZW, Yong G, Liang DS and Sun XY: Overexpressed lncRNA
GATA6-AS1 Inhibits LNM and EMT via FZD4 through the Wnt/β-catenin
signaling pathway in GC. Mol Ther Nucleic Acids. 19:827–840. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Catela Ivkovic T, Voss G, Cornella H and
Ceder Y: microRNAs as cancer therapeutics: A step closer to
clinical application. Cancer Lett. 407:113–122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen QQ, Shi JM, Ding Z, Xia Q, Zheng TS,
Ren YB, Li M and Fan LH: Berberine induces apoptosis in
non-small-cell lung cancer cells by upregulating miR-19a targeting
tissue factor. Cancer Manag Res. 11:9005–9015. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Liu R, Yang F, Cheng R, Chen X, Cui
S, Gu Y, Sun W, You C, Liu Z, et al: miR-19a promotes colorectal
cancer proliferation and migration by targeting TIA1. Mol Cancer.
16:532017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Zhao S, Zhu L, Zhang Q and Ren Y:
miR-19a negatively regulated the expression of PTEN and promoted
the growth of ovarian cancer cells. Gene. 670:166–173. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang B, Liu Y and Zhang J: Silencing of
miR-19a-3p enhances osteosarcoma cells chemosensitivity by
elevating the expression of tumor suppressor PTEN. Oncol Lett.
17:414–421. 2019.PubMed/NCBI
|
|
13
|
Ren S and Xu Y: AC016405.3, a novel long
noncoding RNA, acts as a tumor suppressor through modulation of
TET2 by microRNA-19a-5p sponging in glioblastoma. Cancer Sci.
110:1621–1632. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Alven S and Aderibigbe BA: Nanoparticles
formulations of artemisinin and derivatives as potential
therapeutics for the treatment of cancer, leishmaniasis and
malaria. Pharmaceutics. 12:7482020. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sheng J, Wang L, Han Y, Chen W, Liu H,
Zhang M, Deng L and Liu YN: Dual roles of protein as a template and
a sulfur provider: A general approach to metal sulfides for
efficient photothermal therapy of cancer. Small. Nov 17–2017.(Epub
ahead of print). doi: 10.1002/smll.201702529.
|
|
16
|
Wang P, Wang J, Tan H, Weng S, Cheng L,
Zhou Z and Wen S: Acid- and reduction-sensitive micelles for
improving the drug delivery efficacy for pancreatic cancer therapy.
Biomater Sci. 6:1262–1270. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang H, Liu Y, Bai F, Zhang JY, Ma SH, Liu
J, Xu ZD, Zhu HG, Ling ZQ, Ye D, et al: Tumor development is
associated with decrease of TET gene expression and
5-methylcytosine hydroxylation. Oncogene. 32:663–669. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tucker DW, Getchell CR, McCarthy ET, Ohman
AW, Sasamoto N, Xu S, Ko JY, Gupta M, Shafrir A, Medina JE, et al:
Epigenetic reprogramming strategies to reverse global loss of
5-hydroxymethylcytosine, a prognostic factor for poor survival in
high-grade serous ovarian cancer. Clin Cancer Res. 24:1389–1401.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang LY, Li PL, Wang TZ and Zhang XC:
Prognostic values of 5-hmC, 5-mC and TET2 in epithelial ovarian
cancer. Arch Gynecol Obstet. 292:891–897. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zou MF, Ling J, Wu QY and Zhang CX: Long
non-coding RNA PVT1 functions as an oncogene in ovarian cancer via
upregulating SOX2. Eur Rev Med Pharmacol Sci.
24:75712020.PubMed/NCBI
|
|
22
|
Martini P, Paracchini L, Caratti G,
Mello-Grand M, Fruscio R, Beltrame L, Calura E, Sales G, Ravaggi A,
Bignotti E, et al: lncRNAs as novel indicators of patients'
prognosis in stage I epithelial ovarian cancer: A retrospective and
multicentric study. Clin Cancer Res. 23:2356–2366. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xie T, Wu D, Li S, Li X, Wang L, Lu Y,
Song Q, Sun X and Wang X: microRNA-582 potentiates liver and lung
metastasis of gastric carcinoma cells through the FOXO3-mediated
PI3K/Akt/Snail pathway. Cancer Manag Res. 12:5201–5212. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ni WJ and Leng XM: miRNA-dependent
activation of mRNA Translation. Microrna. 5:83–86. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deb B, Uddin A and Chakraborty S: miRNAs
and ovarian cancer: An overview. J Cell Physiol. 233:3846–3854.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiang G and Cheng Y: miR-126-3p inhibits
ovarian cancer proliferation and invasion via targeting PLXNB2.
Reprod Biol. 18:218–224. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu ZH, Yao TZ and Liu W: miR-378a-3p
sensitizes ovarian cancer cells to cisplatin through targeting
MAPK1/GRB2. Biomed Pharmacother. 107:1410–1417. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang CB, Xiao SW, Cheng SM and Zhang C:
LncRNA FAS-AS1 inhibits the progression of non-small cell lung
cancer through regulating miR-19a-5p. Eur Rev Med Pharmacol Sci.
24:3775–3785. 2020.PubMed/NCBI
|
|
29
|
Liu GM, Lu TC, Sun ML, Ji X, Zhao YA, Jia
WY and Luo YG: RP11-874J12.4 promotes oral squamous cell carcinoma
tumorigenesis via the miR-19a-5p/EBF1 axis. J Oral Pathol Med.
49:645–654. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sun HX, Yang ZF, Tang WG, Ke AW, Liu WR,
Li Y, Gao C, Hu B, Fu PY, Yu MC, et al: MicroRNA-19a-3p regulates
cell growth through modulation of the PIK3IP1-AKT pathway in
hepatocellular carcinoma. J Cancer. 11:2476–2484. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Su YF, Zang YF, Wang YH and Ding YL:
miR-19-3p induces tumor cell apoptosis via targeting FAS in rectal
cancer cells. Technol Cancer Res Treat. 19:15330338209179782020.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ko M, An J, Pastor WA, Koralov SB,
Rajewsky K and Rao A: TET proteins and 5-methylcytosine oxidation
in hematological cancers. Immunol Rev. 263:6–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chiba S: Dysregulation of TET2 in
hematologic malignancies. Int J Hematol. 105:17–22. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ko M, Huang Y, Jankowska AM, Pape UJ,
Tahiliani M, Bandukwala HS, An J, Lamperti ED, Koh KP, Ganetzky R,
et al: Impaired hydroxylation of 5-methylcytosine in myeloid
cancers with mutant TET2. Nature. 468:839–843. 2010. View Article : Google Scholar : PubMed/NCBI
|