Introduction
Cervical cancer is one of the most common types of
cancer worldwide and a leading cause of cancer-associated mortality
in women (1,2). In 2012, the incidence of cervical
cancer was 527,600 cases and nearly 265,700 mortalities worldwide
(3). According to its heterogeneity,
cervical cancer is histologically divided into three subtypes,
adenosquamous carcinoma, squamous cell carcinoma and adenocarcinoma
(4). Currently, the main treatment
strategies for patients with cervical cancer include pelvic lymph
node dissection, hysterectomy and radiotherapy combined with
chemotherapy (5). Although a certain
degree of progress has been made in the early diagnosis, surgical
treatment and application of the human papillomavirus (HPV)16, 18,
31, 33, 45, 52 and 58 vaccine for cervical cancer, the survival
rate of patients with advanced cervical cancer remains low
(6,7). The 5-year survival rate for cervical
cancer in China from 2012–2015 was only 59.8% (8). Thus, it is important to identify and
develop biomarkers for cervical cancer and determine its disease
mechanism.
Non-coding RNAs (ncRNAs) are distinctive RNA
molecules that are not translated into proteins (9). ncRNAs are divided into two types,
including housekeeping and regulatory ncRNAs (10). The regulatory ncRNAs include
microRNAs (miRNAs/miRs), circular RNAs (circRNAs) and long ncRNAs
(lncRNAs) (11). miRNAs are short
ncRNAs, ~22 nucleotides in length that regulate gene expression by
binding to mRNAs (12–14) or by adjusting their stability through
sponging circRNAs (15,16). lncRNAs are a group of ncRNAs with a
length of >200 nucleotides (17).
lncRNAs function as sponges that target miRNAs and subsequently
regulate gene expression (18,19).
Several studies have reported that miRNAs and lncRNAs play
important roles in certain types of cancer (20–22).
Increasing evidence suggest that lncRNAs are
involved in the occurrence of multiple tumors (23–25).
Several lncRNAs, such as growth arrest-specific 5 (26), nuclear paraspeckle assembly
transcript 1 (27),
metastasis-associated lung adenocarcinoma transcript 1 (28) and small nucleolar host gene 12
(29), have been identified as
important regulatory factors in cervical cancer. The lncRNA,
urothelial carcinoma-associated 1 (UCA1) is located on human
19p13.12 (30) and has been reported
to play an important role in different types of cancer, including
colorectal (31), prostate (32), gastric (33) and bladder cancer (34). Previous studies have reported that
UCA1 regulates the proliferation, migration and invasion of
cervical cancer cells (35–37). However, the mechanism of UCA1 in the
progression of cervical cancer remains unclear.
miRNAs have been reported to act as facilitators or
suppressors in different types of cancer (38–40).
According to previous studies, miR-299-3p participates in multiple
tumor processes. In hepatocellular carcinoma, miR-299-3p acts as a
tumor suppressor by regulating Sirtuin 5 (41). It also plays a tumor suppressive role
by targeting SHOC2 leucine-rich repeat scaffold protein in thyroid
cancer (42). miR-299-3p inhibits
cell proliferation and invasion by targeting vascular endothelial
growth factor A in human colon carcinoma (43). In cervical cancer, miR-299-3p
suppresses cell proliferation and invasion by binding to
transcription factor 4 (TCF4) (44).
However, the function of miR-299-3p, and the association between
UCA1 and miR-299-3p in cervical cancer remain unknown.
The present study aimed to investigate the role and
potential mechanism of UCA1 in cervical cancer. The association
between UCA1 and miR-299-3p in the occurrence of cervical cancer
was also investigated.
Materials and methods
Clinical specimens
Cervical cancer and paired adjacent normal tissues
(n=30) were surgically collected from the Department of Obstetrics
and Gynecology, Sanya People's Hospital (Sanya, China) from May
2018 to May 2020. The average age of all patients was 49 years (age
range, 36–69 years). The clinical information of patients with
cervical cancer is presented in Table
SI. No patients received adjuvant treatment prior to surgery.
In addition, the patients with other tumors or a history of
treatments for other gynecological tumors were excluded from the
present study. All tissue samples were immediately preserved in
liquid nitrogen and stored at −80°C until RNA extraction. The
present study was approved by the Ethics Committee of the Sanya
People's Hospital (approval no. 2017.106; Sanya, China) and written
informed consent was provided by all patients prior to the study
start.
Cell lines
The Ect1/E6E7 normal human cervical epithelial cell
line was purchased from the American Type Culture Collection, while
the SiHa, HeLa, CaSki and ME180 human cervical cancer cell lines
were purchased from the The Cell Bank of Type Culture Collection of
The Chinese Academy of Sciences. All cells were maintained in DMEM
(cat. no. SH30022.01B; Cytiva) supplemented with 10% fetal bovine
serum (cat. no. 16140071; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin (cat. no. 15140148; Thermo Fisher Scientific, Inc.) and
100 µg/ml streptomycin (cat. no. 15140122; Thermo Fisher
Scientific, Inc.), at 37°C with 5% CO2 and 100%
humidity.
Cell transfection
The overexpression vector pcDNA 3.1 was purchased
from Thermo Fisher Scientific, Inc. All sequences were synthesized
by Shanghai GenePharma Co., Ltd. The primers for the amplification
of the full-length UCA1 were as follows: Forward,
5′-CCGGAATTCTGACATTCTTCTGGACAATG-3′ and reverse,
5′-CCGCTCGAGCTGACTCTTTTAGGAAGATTTCT-3′. The overexpression vector
pcDNA-UCA1 was produced by cloning the full-length UCA1 into
pcDNA3.1. For the knockdown of UCA1, the small interfering (si)RNAs
targeting UCA1 (si-UCA1) were designed. The sequences was as
follows: 5′-GGACAACAGUACACGCAUATT-3′. The sequences used to
increase or decrease miR-299-3p expression were as follows:
miR-299-3p mimics, 5′-UCGCCAAAUGGUAGGGUGUAU-3′; miR-299-3p
inhibitor, 5′-AAGCGGUUUACCAUCCCACAU-3′; miR-NC mimics,
5′-UUCUCCGAACGUGUCACGUG-3′ and miR-NC inhibitor,
5′-CAGUACUUUUGUGUAGUACA-3′. The UCA1 overexpression vector
pcDNA-UCA1, specific siRNAs targeting UCA1 (si-UCA1), negative
control siRNA (si-NC), miR-299-3p mimics, NC mimics, miR-299-3p
inhibitor and NC inhibitor were transfected into SiHa cells. For
cell transfection, SiHa cells were seeded into a 6-well plate wat a
density of 1×105 cells/well. 1.5 ml of serum-free medium
containing 500 µl of Lipofectamine™ 3000 transfection solution
(cat. no. L3000015; Thermo Fisher Scientific, Inc.) were added in
each well at 37°C for 48 h. After cell culture for 1–2 weeks, the
positive colonies were harvested for amplification culture.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from normal and tumor
tissues and cell lines using TRIzol® reagent (cat. no.
15596018; Thermo Fisher Scientific, Inc.). The reaction mixture
containing 1 µg of total RNA was reverse transcribed into cDNA
using the PrimeScript™ RT reagent kit (cat. no. RR036A; Takara Bio,
Inc.). The temperature protocols for RT were as follows: 50°C for
15 min and 85°C for 5 sec. The reactions were performed using an
ABI 7500 Real-Time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.), and the RNA expression levels were measured
using a SYBR™ Green Master Mix (cat. no. RR430S; Takara Bio, Inc.).
The thermocycling conditions of qPCR were as follows: Initial
denaturation at 95°C for 3 min, followed by 45 cycles of
denaturation at 95°C for 15 sec and annealing/elongation at 60°C
for 20 sec. The following primer sequences were used for qPCR: UCA1
forward, 5′-GCCAGCCTCAGCTTAATCCA-3′ and reverse,
5′-CCCTGTTGCTAAGCCGATGA-3′; miR-299-3p forward,
5′-ACACTCCAGCTGGGTATGTGGGATGGTAAAC-3′ and reverse,
5′-GTGCAGGGTCCGAG-3′; U6 forward, 5′-CTCGCTTCGGCAGCACA-3′ and
reverse, 5′-AACGCTTCACGAATTTGCGT-3′ reverse; and GAPDH forward,
5′-ACCCACATCCCTCAGACAC-3′ and reverse, 5′-CCCCAATACGACCAAATCC-3′.
GAPDH and U6 were used as the internal controls. Relative
expression levels were calculated using the 2−ΔΔCq
method (45). The experiments were
performed in triplicate.
Cell Counting Kit-8 (CCK-8) assay
Cervical cancer cell proliferation was assessed via
the CCK-8 assay (cat. no. C0039; Beyotime Institute of
Biotechnology). The transfected cells were collected by
centrifugation at 1,000 × g for 5 min at 4°C, then the transfected
cells (2×103 cells/100 µl) were cultured in 96-well
plates. Following incubation for 0, 24, 48 and 72 h, 10 µl CCK-8
solution was added into each well and incubated for an additional 2
h at 37°C. Cell proliferation was subsequently analyzed at a
wavelength of 450 nm. The experiments were performed in
triplicate.
Invasion assay
Cell invasion was assessed via the Transwell assay.
The Transwell chambers were pre-coated with 100 µl Matrigel at 37°C
for 1 h. Transfected cells were collected (1,000 × g for 5 min at
4°C), then the cells (1×105 cells/well) were seeded into
the upper chambers of Matrigel-coated Transwell plates (8-µm pore
size; cat. no. 354483; Corning, Inc.) in serum-free DMEM. The DMEM
(cat. no. SH30022.01B; Cytiva) medium with 10% fetal bovine serum
(cat. no. 16140071; Thermo Fisher Scientific, Inc.) was plated in
the lower chambers. Following incubation for 24 h at 37°C, cells in
the lower chambers were fixed with 5% glutaraldehyde and stained
with 0.1% crystal violet dye (cat. no. 548-62-9; MilliporeSigma)
for 30 min at room temperature, respectively. Stained cells were
counted in five randomly selected fields using a BX63 microscope
(magnification, ×100, Olympus Corporation).
Dual-luciferase reporter assay
LncBase Predicted v.2 software (46) was used to predict the potential
binding sites between UCA1 and miR-299-3p.
The dual-luciferase reporter plasmids were
constructed by inserting a UCA1 wild-type (UCA1-WT) or UCA1 mutant
(UCA1-MUT) sequence into the psiCHECK-2 luciferase vector (Promega
Corporation). SiHa cells were co-transfected with UCA1-WT or -MUT
reporter and miR-299-3p or NC mimics, using
Lipofectamine® 3000 transfection reagent (cat. no.
L3000015; Thermo Fisher Scientific, Inc.). Transfected cells were
subsequently cultured in 24-well plates. Following incubation at
37°C for 24 h, luciferase activities were detected using a
Dual-Luciferase Reporter assay system (cat. no. E1910; Promega
Corporation). Firefly luciferase activity was normalized to
Renilla luciferase activity (cat. no. E2810; Promega
Corporation).
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 8 (GraphPad Software, Inc.). All experiments were performed
in triplicate and data are presented as the mean ± SD. Paired
Student's t-test was used to compare differences between tumor
tissues and adjacent normal tissues, while unpaired Student's
t-test was used to compare differences between unpaired groups.
One-way ANOVA followed by Tukey's or Dunnett's post hoc tests were
used to compare differences between multiple groups. Pearson's
correlation coefficient analysis was performed to assess the
correlation between UCA1 and miR-299-3p expression. P<0.05 was
considered to indicate a statistically significant difference.
Results
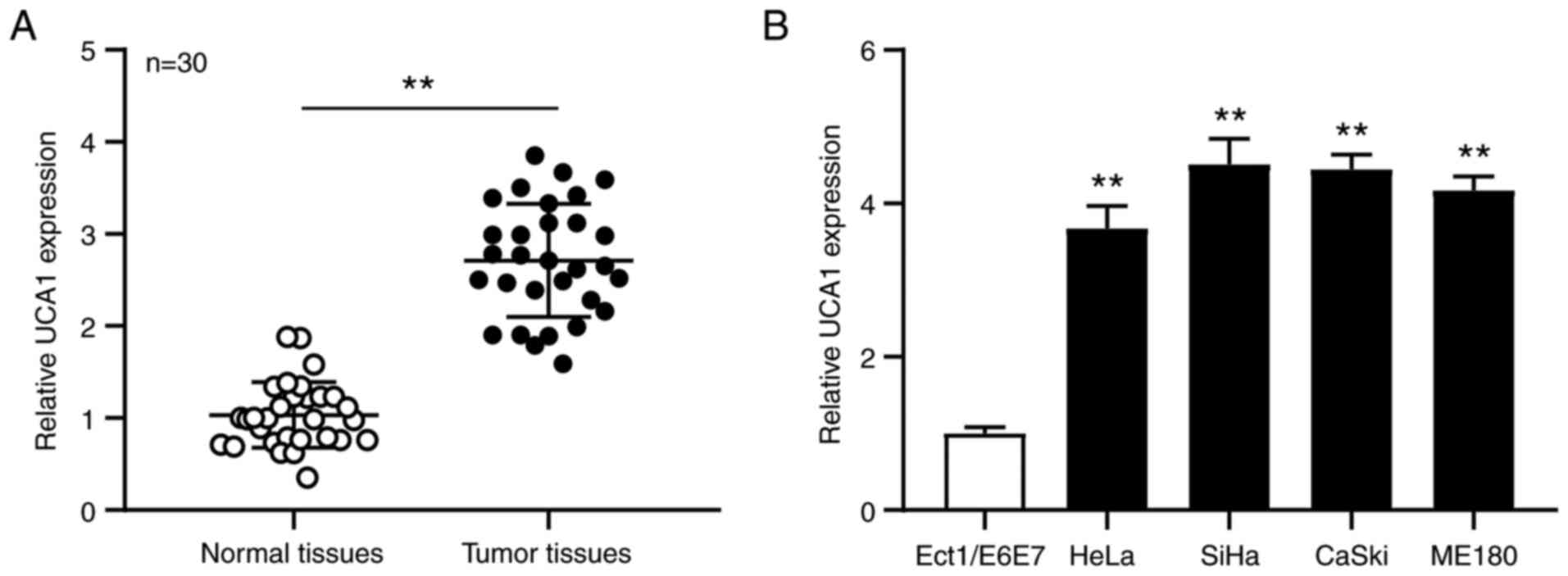
UCA1 expression is downregulated in
cervical cancer tissues and cell lines
RT-qPCR analysis was performed to detect UCA1
expression in cervical cancer tissues and adjacent normal tissues
(n=30). The results demonstrated that UCA1 expression was
significantly upregulated in cervical cancer tissues compared with
adjacent normal tissues (P<0.01; Fig.
1A). RT-qPCR analysis was also performed to detect UCA1
expression in the SiHa, HeLa, CaSki and ME180 cervical cancer cell
lines and Ect1/E6E7 human normal cervical epithelial cell line. The
results demonstrated that UCA1 expression was significantly
upregulated in the cervical cancer cell lines compared with the
normal cell line (all P<0.01; Fig.
1B). Taken together, these results suggest that UCA1 plays a
key role in the development of cervical cancer.
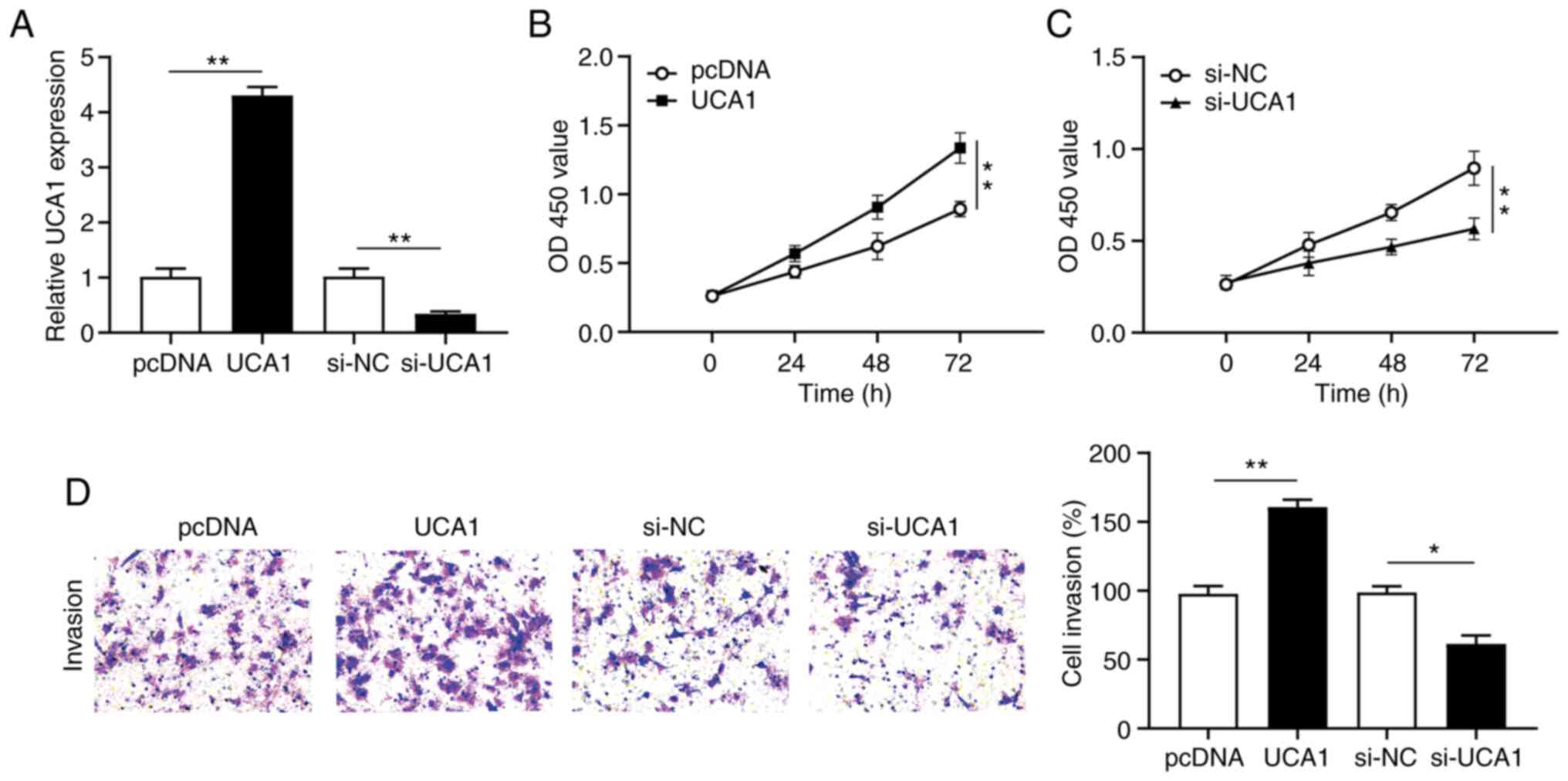
UCA1 positively regulates cervical
cancer cell proliferation and invasion
To further investigate the effect of UCA1 in
cervical cancer cell proliferation and invasion, SiHa cells were
transfected with UCA1 or si-UCA1 to construct a UCA1 overexpression
and knockdown model, respectively. The expression of UCA1 in
transfected SiHa cells was detected via RT-qPCR analysis. The
results demonstrated that UCA1 expression significantly increased
(P<0.01) following transfection with UCA1 and significantly
decreased (P<0.01) following transfection with si-UCA1 (Fig. 2A).
The CCK-8 assay was performed to assess the
proliferation of cervical cancer cells. The results demonstrated
that overexpression of UCA1 significantly promoted the
proliferation of SiHa cells (P<0.01; Fig. 2B), the effects of which were reversed
following UCA1 knockdown (P<0.01; Fig. 2C). Furthermore, the Transwell assay
demonstrated that overexpression of UCA1 significantly promoted the
invasive ability of SiHa cells (P<0.01; Fig. 2D), the effects of which were reversed
following UCA1 knockdown. Collectively, these results suggest that
UCA1 is involved in cervical cancer cell proliferation and
invasion.
miR-299-3p can bind with UCA1, and its
expression is negatively regulated by UCA1
To clarify the molecular mechanism of UCA1 in
cervical cancer, LncBase software was used to predict the binding
sites between miRNAs and UCA1. The results revealed a potential
binding site between UCA1 and miR-299-3p (Fig. 3A). The dual-luciferase reporter assay
was subsequently performed to confirm the association between UCA1
and miR-299-3p in SiHa cells. The results demonstrated that
transfection with miR-299-3p mimics significantly decreased
(P<0.01) the luciferase activity of UCA1-WT, but not UCA1-MUT in
SiHa cells (Fig. 3B). RT-qPCR
analysis was performed to detect miR-299-3p expression in cervical
cancer tissues and cell lines. The results demonstrated that
miR-299-3p expression was significantly downregulated in cervical
cancer tissues (P<0.01; Fig. 3C)
and cell lines (all P<0.01; Fig.
3D). Pearson's correlation analysis was performed to determine
the correlation between UCA1 and miR-299a-3p expression in cervical
cancer tissues. The results demonstrated that miR-299-3p expression
was negatively correlated with UCA1 expression (Fig. 3E). SiHa cells were transfected with
pcDNA, UCA1, si-NC or si-UCA1, and miR-299-3p expression was
detected. The results demonstrated that overexpression of UCA1
significantly decreased miR-299-3p expression (P<0.01), while
UCA1 knockdown significantly increased miR-299-3p expression
(P<0.01) in SiHa cells (Fig. 3F).
Taken together, these results suggest that UCA1 binds to miR-299-3p
and negatively regulates its expression in cervical cancer.
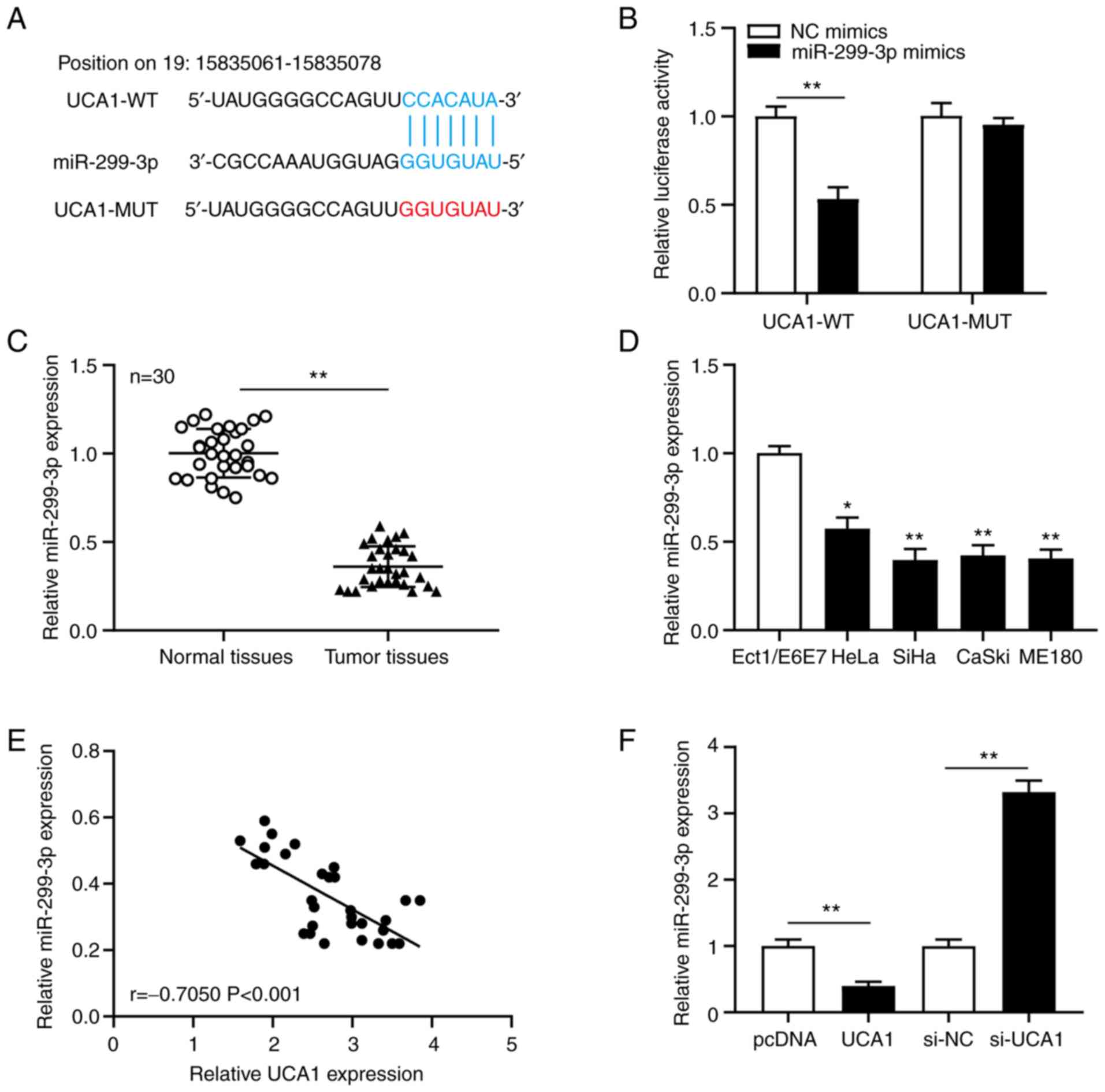 | Figure 3.miR-299-3p directly targets the
3′-untralsated region of UCA1 and is downregulated in cervical
cancer tissues and cell lines. (A) Binding sites between UCA1 and
miR-299-3p were predicted using LncBase software. (B) The
association between UCA1 and miR-299-3p was confirmed via the
dual-luciferase reporter assay. RT-qPCR analysis was performed to
detect miR-299-3p expression in (C) cervical cancer tissues and
normal tissues, and (D) cervical cancer cell lines and Ect1/E6E7.
(E) UCA1 expression was negatively correlated with miR-299-3p
expression in cervical cancer. (F) RT-qPCR analysis was performed
to detect miR-299-3p expression in SiHa cells transfected with
pcDNA, UCA1, si-NC or si-UCA1. *P<0.05, **P<0.01 vs.
Ect1/E6E7 cells. miR, microRNA; UCA1, urothelial cancer-associated
1; RT-qPCR, reverse transcription-quantitative PCR; si, small
interfering; NC, negative control; WT, wild-type; MUT, mutant. |
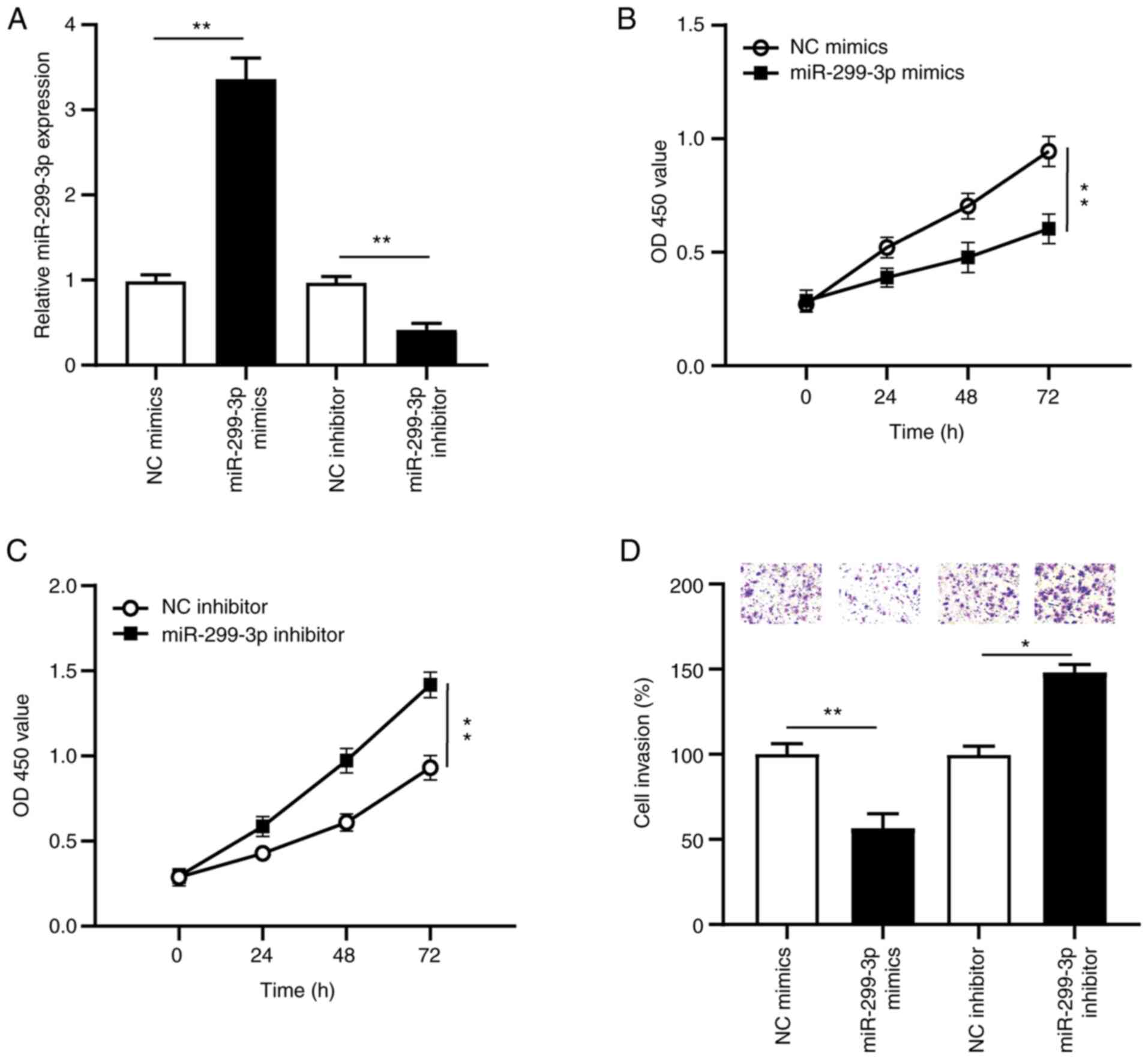
miR-299-3p suppresses cervical cancer
cell proliferation and invasion
To determine the biological function of miR-299-3p
in the proliferation and invasion of cervical cancer cells, SiHa
cells were transfected with miR-299-3p mimics, NC mimics,
miR-299-3p inhibitor or NC inhibitor. RT-qPCR analysis was
performed to detect miR-299-3p expression. The results demonstrated
that miR-229-3p expression significantly increased (P<0.01)
following transfection with miR-299-3p mimics, the effects of which
were reversed following transfection with miR-299-3p inhibitor
(P<0.01), compared with the NC groups (Fig. 4A).
The proliferation and invasion of cervical cancer
cells were assessed via the CCK-8 and Transwell assays,
respectively. The results confirmed that overexpression of
miR-299-3p significantly decreased cervical cancer cell
proliferation (P<0.01; Fig. 4B),
while miR-299-3p knockdown significantly increased cervical cancer
cell proliferation (P<0.01; Fig.
4C). The Transwell assay demonstrated that transfection with
miR-299-3p mimics significantly decreased cervical cancer cell
invasion (P<0.01; Fig. 4D), while
transfection with miR-299-3p inhibitor significantly promoted
cervical cancer cell invasion (P<0.05; Fig. 4D). Collectively, these results
suggest that miR-299-3p acts as a tumor suppressor in cervical
cancer.
Overexpression of UCA1 promotes cell
proliferation and invasion by suppressing miR-299-3p
expression
To further clarify the effect of the UCA1/miR-299-3p
axis in cervical cancer tumorigenesis, rescue experiments were
performed on SiHa cells. miR-299-3p mimics + UCA1 or miR-299-3p
inhibitor + si-UCA1 were transfected into SiHa cells. RT-qPCR
analysis demonstrated that overexpression of miR-299-3p partially
reversed the inhibitory effects of overexpressing UCA1 on SiHa
cells (P<0.05; Fig. 5A), while
transfection with miR-299-3p inhibitor reversed the promoting
effects of si-UCA1 on SiHa cells (P<0.01; Fig. 5B). Furthermore, transfection with
miR-299-3p mimics inhibited the proliferative ability of
UCA1-transfected SiHa cells (P<0.01; Fig. 5C), and si-UCA1-induced suppression in
cell proliferation was recovered following transfection with
miR-299-3p inhibitor (P<0.01; Fig.
5D). Furthermore, the invasive ability of UCA1-transfected SiHa
cells decreased following transfection with miR-299-3p mimics
(P<0.01; Fig. 5E), the effects of
which were reversed following transfection with miR-299-3p
inhibitor (P<0.05; Fig. 5F).
Collectively, these results suggest that UCA1 promotes cervical
cancer progression by regulating miR-299-3p expression.
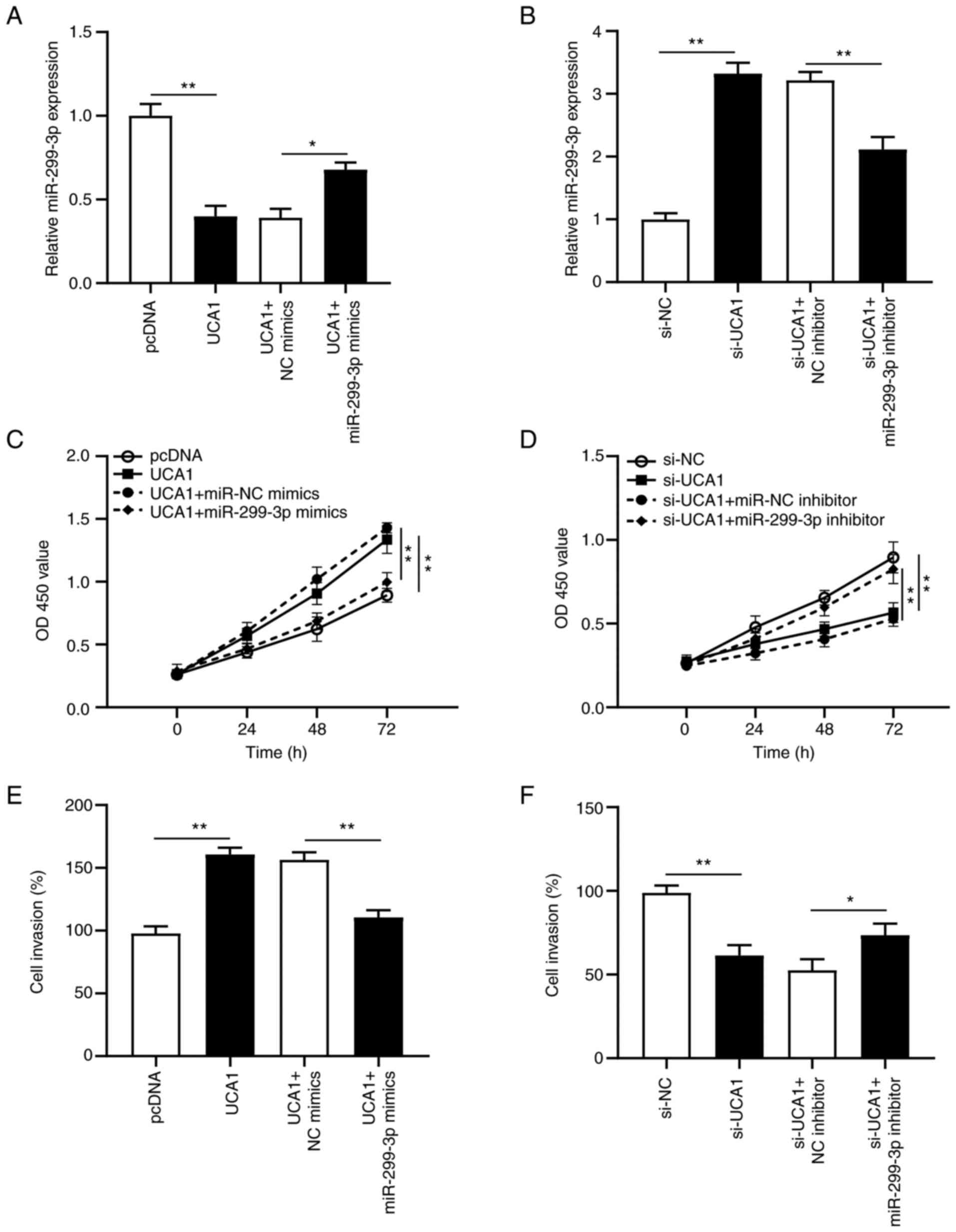 | Figure 5.UCA1 enhances proliferation and
invasion by inhibiting miR-299-3p expression in SiHa cells. SiHa
cells were transfected with pcDNA, UCA1, UCA1 + NC mimics, UCA1 +
miR-299-3p mimics, si-NC, si-UCA1, si-UCA1 + NC inhibitor and
si-UCA1 + miR-299-3p inhibitor. (A and B) Reverse
transcription-quantitative PCR analysis was performed to detect
miR-299-3p expression following transfection. (C and D) The Cell
Counting Kit-8 assay was performed to assess cell proliferation
following transfection. (E and F) The Transwell assay was performed
to assess cell invasion following transfection. *P<0.05;
**P<0.01. UCA1, urothelial cancer-associated 1; miR, microRNA;
NC, negative control; si, small interfering; OD, optical
density. |
Discussion
Cervical cancer is a common gynecological malignancy
and the second most serious threat to women's health worldwide
(47). Most cases of cervical cancer
are caused by HPV infection (48).
Although surgical treatment, chemotherapy and radiotherapy have a
therapeutic effect on cervical cancer, this effect is insufficient
(49). Recent studies have reported
that lncRNAs play important regulatory roles in cervical cancer
(50–52). Although several studies have
clarified the role of UCA1 in different tumors (31–33), to
the best of our knowledge, the molecular mechanism of UCA1 in
cervical cancer remains unclear. In the present study, the role of
UCA1 and its molecular mechanism in cervical cancer were further
explored.
The lncRNA, UCA1 acts as an oncogene in different
types of cancer, including pancreatic (53), gastric (54) and colorectal (26) cancers. Previous studies have
demonstrated that UCA1 is pregulated in cervical cancer, which
promotes cell proliferation, invasion and migration by regulating
miR-145 or miR-204 expression (35,36).
Consistent with these findings, the results of the present study
demonstrated that UCA1 was upregulated in cervical cancer tissues
and cell lines. Furthermore, overexpression of UCA1 promoted
cervical cancer cell proliferation and invasion, the effects of
which were reversed following UCA1 knockdown. Taken together, these
findings suggest that UCA1 may play a key role in the proliferation
and invasion of cervical cancer cells.
A recent study reported that miRNAs play important
roles in cervical cancer. For example, miR-216a-3p inhibits
cervical cancer cell proliferation and invasion by regulating
actin-like 6A (55). Furthermore,
miR-195-5p suppresses migration and invasion in cervical cancer by
targeting ADP ribosylation factor-like GTPase 2 (ARL2) (56). Overexpression of miR-139-5p promotes
cell proliferation and migration in cervical cancer by targeting
TCF4 (57). miR-299-3p has been
reported to act as a tumor suppressor in hepatocellular carcinoma,
thyroid cancer and colon carcinoma (41–43). In
addition, miR-299-3p inhibits cell proliferation and invasion by
regulating TCF4 in cervical cancer (44). In the present study, miR-299-3p
expression was significantly downregulated in cervical cancer, and
its overexpression inhibited cervical cancer cell proliferation and
invasion. Collectively, the results of the present study suggest
that miR-299-3p plays an inhibitory role in cervical cancer.
Previous studies have reported that lncRNAs bind to
specific miRNAs to regulate cancer progression (58,59).
lncRNA UCA1 has been reported to regulate the progression of
several tumors by targeting specific miRNAs. For example, UCA1
promotes cell proliferation in gastric cancer by regulating the
miR-495/phosphatase of regenerating liver 3 axis (54) and mitochondrial function in bladder
cancer via the miR-195/ARL2 signaling pathway (34). To further clarify the potential
molecular mechanism of UCA1 in cervical cancer, the target genes of
UCA1 were predicted using LncBase. The results revealed that UCA1
can bind to miR-299-3p. In addition, miR-299-3p expression was
negatively correlated with UCA1 expression in cervical cancer
tissues. Notably, overexpression of UCA1 decreased miR-299-3p
expression in SiHa cervical cancer cells. Furthermore, transfection
with miR-299-3p mimics inhibited cell proliferation and invasion,
and transfection with miR-299-3p inhibitors reversed the effects of
UCA1 knockdown on SiHa cells. Taken together, these results suggest
that UCA1 acts as an oncogene in cervical cancer and regulates cell
proliferation and invasion by inhibiting miR-299-3p expression.
The results of the present study provide novel
insight into the treatment of cervical cancer. Notably, a novel
target miRNA of UCA1 in cervical cancer, which has not been
previously reported, was identified in the present study. In
addition, the UCA1/miR-299-3p axis was revealed to regulate cell
proliferation and invasion in cervical cancer, providing a novel
target site for the treatment of cervical cancer. However, the
present study is not without limitations. The sample size assessed
was too small; thus, further studies with a larger sample size are
required to investigate the mechanism of the regulatory effect of
the UCA1/miR-299-3p axis on cell proliferation and invasion in
cervical cancer. In addition, the present only performed in
vitro experiments, and needs to be further investigated in
vivo.
In conclusion, the results of the present study
demonstrated that UCA1 promoted cell proliferation and invasion by
targeting miR-299-3p expression in cervical cancer. These results
suggest that the UCA1/miR-299-3p axis may present a potential
therapeutic target for cervical cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author upon reasonable
request.
Authors' contributions
MA and XX conceived and designed the present study,
analyzed the data and drafted the initial manuscript. TC
contributed to data collection, statistical analysis and manuscript
preparation. MA and XX confirmed the authenticity of all the raw
data. All authors have read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Sanya People's Hospital (approval no. 2017.106; Sanya,
China) and written informed consent was provided by all patients
prior to the study start.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cohen PA, Jhingran A, Oaknin A and Denny
L: Cervical cancer. Lancet. 393:169–182. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Huang X, Liu X, Du B, Liu X, Xue M, Yan Q,
Wang X and Wang Q: lncRNA LINC01305 promotes cervical cancer
progression through KHSRP and exosome-mediated transfer. Aging
(Albany NY). 13:19230–19242. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Agarwal R and Kaye SB: Ovarian cancer:
Strategies for overcoming resistance to chemotherapy. Nat Rev
Cancer. 3:502–516. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Burki TK: Cervical cancer: Screening and
risk with age. Lancet Oncol. 15:e1072014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zeng H, Chen W, Zheng R, Zhang S, Ji J,
Zou X, Xia C, Sun K, Yang Z, Li H, et al: Changing cancer survival
in China during 2003–15: A pooled analysis of 17 population-based
cancer registries. Lancet Glob Health. 6:e555–e567. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gong Z, Zhang S, Zhang W, Huang H, Li Q,
Deng H, Ma J, Zhou M, Xiang J, Wu M, et al: Long non-coding RNAs in
cancer. Sci China Life Sci. 55:1120–1124. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hemberg M, Gray JM, Cloonan N, Kuersten S,
Grimmond S, Greenberg ME and Kreiman G: Integrated genome analysis
suggests that most conserved non-coding sequences are regulatory
factor binding sites. Nucleic Acids Res. 40:7858–7869. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Anastasiadou E, Jacob LS and Slack FJ:
Non-coding RNA networks in cancer. Nat Rev Cancer. 18:5–18. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang M, Wang C and Mu L: miR-148a induces
apoptosis by upregulating BIM expression in gastric cancer cells.
Int J Clin Exp Med. 10:2791–2799. 2017.
|
|
13
|
Shen J, Zhang J, Xiao M, Yang J and Zhang
N: miR-203 suppresses bladder cancer cell growth and targets
Twist1. Oncol Res. 26:1155–1165. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tang L, Yang B, Cao X, Li Q, Jiang L and
Wang D: MicroRNA-377-3p inhibits growth and invasion through
sponging JAG1 in ovarian cancer. Genes Genomics. 41:919–926. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang X, Li Z, Zhang Q, Wang W, Li B, Wang
L, Xu Z, Zeng A, Zhang X, Zhang X, et al: Circular RNA AKT3
upregulates PIK3R1 to enhance cisplatin resistance in gastric
cancer via miR-198 suppression. Mol Cancer. 18:712019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu G, Wan Q, Li J, Hu X, Gu X and Xu S:
Circ_0038467 regulates lipopolysaccharide-induced inflammatory
injury in human bronchial epithelial cells through sponging
miR-338-3p. Thorac Cancer. 11:1297–1308. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bill M, Papaioannou D, Karunasiri M,
Kohlschmidt J, Pepe F, Walker CJ, Walker AE, Brannan Z,
Pathmanathan A, Zhang X, et al: Expression and functional relevance
of long non-coding RNAs in acute myeloid leukemia stem cells.
Leukemia. 33:2169–2182. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang Z, Huang D, Huang J, Nie K, Li X and
Yang X: lncRNA TMPO-AS1 exerts oncogenic roles in HCC through
regulating miR-320a/SERBP1 axis. Onco Targets Ther. 13:6539–6551.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li W, Zhang B, Jia Y, Shi H, Wang H, Guo Q
and Li H: lncRNA LOXL1-AS1 regulates the tumorigenesis and
development of lung adenocarcinoma through sponging miR-423-5p and
targeting MYBL2. Cancer Med. 9:689–699. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yan SY, Chen MM, Li GM, Wang YQ and Fan
JG: miR-32 induces cell proliferation, migration, and invasion in
hepatocellular carcinoma by targeting PTEN. Tumour Biol.
36:4747–4755. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sempere LF, Christensen M, Silahtaroglu A,
Bak M, Heath CV, Schwartz G, Wells W, Kauppinen S and Cole CN:
Altered MicroRNA expression confined to specific epithelial cell
subpopulations in breast cancer. Cancer Res. 67:11612–11620. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xie H, Liao X, Chen Z, Fang Y, He A, Zhong
Y, Gao Q, Xiao H, Li J, Huang W and Liu Y: lncRNA MALAT1 inhibits
apoptosis and promotes invasion by antagonizing miR-125b in bladder
cancer cells. J Cancer. 8:3803–3811. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang X, Wang L, Xie S, Chen Y, Song S, Lu
Y and Lu D: Long noncoding RNA MEG3 blocks telomerase activity in
human liver cancer stem cells epigenetically. Stem Cell Res Ther.
11:5182020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ma P, Pan Y, Yang F, Fang Y, Liu W, Zhao
C, Yu T, Xie M, Jing X, Wu X, et al: KLF5-modulated lncRNA NEAT1
contributes to tumorigenesis by acting as a scaffold for BRG1 to
silence GADD45A in gastric cancer. Mol Ther Nucleic Acids.
22:382–395. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shen SN, Li K, Liu Y, Yang CL, He CY and
Wang HR: Silencing lncRNAs PVT1 upregulates miR-145 and confers
inhibitory effects on viability, invasion, and migration in EC. Mol
Ther Nucleic Acids. 19:668–682. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang W, Xu X, Hong L, Wang Q, Huang J and
Jiang L: Upregulation of lncRNA GAS5 inhibits the growth and
metastasis of cervical cancer cells. J Cell Physiol.
234:23571–23580. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yuan LY, Zhou M, Lv H, Qin X, Zhou J, Mao
X, Li X, Xu Y, Liu Y and Xing H: Involvement of NEAT1/miR-133a axis
in promoting cervical cancer progression via targeting SOX4. J Cell
Physiol. 234:18985–18993. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shen F, Zheng H, Zhou L, Li W and Xu X:
Overexpression of MALAT1 contributes to cervical cancer progression
by acting as a sponge of miR-429. J Cell Physiol. 234:11219–11226.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lai SY, Guan HM, Liu J, Huang LJ, Hu XL,
Chen YH, Wu YH, Wang Y, Wu Q and Zhou JY: Long noncoding RNA SNHG12
modulated by human papillomavirus 16 E6/E7 promotes cervical cancer
progression via ERK/Slug pathway. J Cell Physiol. 235:7911–7922.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang XS, Zhang Z, Wang HC, Cai JL, Xu QW,
Li MQ, Chen YC, Qian XP, Lu TJ, Yu LZ, et al: Rapid identification
of UCA1 as a very sensitive and specific unique marker for human
bladder carcinoma. Clin Cancer Res. 12:4851–4858. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Luan Y, Li X, Luan Y, Zhao R, Li Y, Liu L,
Hao Y, Oleg Vladimir B and Jia L: Circulating lncRNA UCA1 promotes
malignancy of colorectal cancer via the miR-143/MYO6 axis. Mol Ther
Nucleic Acids. 19:790–803. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu Y, Gao F, He Q, Li G and Ding G: lncRNA
UCA1 functions as a ceRNA to promote prostate cancer progression
via sponging miR143. Mol Ther Nucleic Acids. 19:751–758. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
He X, Wang J, Chen J, Han L, Lu X, Miao D,
Yin D, Geng Q and Zhang E: lncRNA UCA1 predicts a poor prognosis
and regulates cell proliferation and migration by repressing p21
and SPRY1 expression in GC. Mol Ther Nucleic Acids. 18:605–616.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li HJ, Sun XM, Li ZK, Yin QW, Pang H, Pan
JJ, Li X and Chen W: lncRNA UCA1 promotes mitochondrial function of
bladder cancer via the miR-195/ARL2 signaling pathway. Cell Physiol
Biochem. 43:2548–2561. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wei H, Qiu YQ, Zeng QS, Wang SF and Yi CJ:
lncRNA UCA1 regulates proliferation, migration and invasion of
cervical cancer cells by targeting miR-145. Eur Rev Med Pharmacol
Sci. 24:3555–3564. 2020.PubMed/NCBI
|
|
36
|
He Q, Meng J, Liu S, Zeng Q, Zhu Q, Wei Z
and Shao Y: Long non-coding RNA UCA1 upregulates KIF20A expression
to promote cell proliferation and invasion via sponging miR-204 in
cervical cancer. Cell Cycle. 19:2486–2495. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang TJ, Wang L, Zhang Y, Zheng JD and Liu
L: lncRNA UCA1 regulates cervical cancer survival and EMT
occurrence by targeting miR-155. Eur Rev Med Pharmacol Sci.
24:9869–9879. 2020.PubMed/NCBI
|
|
38
|
Zhang Y, Pan Q and Shao Z:
Tumor-suppressive role of microRNA-202-3p in hepatocellular
carcinoma through the KDM3A/HOXA1/MEIS3 pathway. Front Cell Dev
Biol. 8:5560042021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang Y and Qin H: miR-338-3p targets RAB23
and suppresses tumorigenicity of prostate cancer cells. Am J Cancer
Res. 8:2564–2574. 2018.PubMed/NCBI
|
|
40
|
Wang R, Zuo X, Wang K, Han Q, Zuo J, Ni H,
Liu W, Bao H, Tu Y and Xie P: MicroRNA-485-5p attenuates cell
proliferation in glioma by directly targeting paired box 3. Am J
Cancer Res. 8:2507–2517. 2018.PubMed/NCBI
|
|
41
|
Dang S, Zhou J, Wang Z, Wang K, Dai S and
He S: miR-299-3p functions as a tumor suppressor via targeting
Sirtuin 5 in hepatocellular carcinoma. Biomed Pharmacother.
106:966–975. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen X, Qi M, Yang Q and Li J: miR-299-3p
functions as a tumor suppressor in thyroid cancer by regulating
SHOC2. Eur Rev Med Pharmacol Sci. 23:232–240. 2019.PubMed/NCBI
|
|
43
|
Wang JY, Jiang JB, Li Y, Wang YL and Dai
Y: MicroRNA-299-3p suppresses proliferation and invasion by
targeting VEGFA in human colon carcinoma. Biomed Pharmacother.
93:1047–1054. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yu Y, Zhao JD and Yang H: miR-299-3p
inhibits proliferation and invasion of cervical cancer cell via
targeting TCF4. Eur Rev Med Pharmacol Sci. 23:5621–5627.
2019.PubMed/NCBI
|
|
45
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Das T, Deb A, Parida S, Mondal S, Khatua S
and Ghosh Z: LncRBase V.2: An updated resource for multispecies
lncRNAs and ClinicLSNP hosting genetic variants in lncRNAs for
cancer patients. RNA Biol. 18:1136–1151. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Waggoner SE: Cervical cancer. Lancet.
361:2217–2225. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Roik E, Sharashova E, Kharkova O, Nieboer
E, Postoev V and Odland JØ: Sociodemographic characteristics,
sexual behaviour and knowledge about cervical cancer prevention as
risk factors for high-risk human papillomavirus infection in
Arkhangelsk, North-West Russia. Int J Circumpolar Health.
77:14986812018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang Q, Yan SP, Chu DX, Xie Y, Wang CF,
Zhang JY, Li WC and Guo RX: Silencing of long non-coding RNA
RP1-93H18.6 acts as a tumor suppressor in cervical cancer through
the blockade of the PI3K/Akt axis. Mol Ther Nucleic Acids.
19:304–317. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Chen DZ, Wang TF, Dai WC, Xu X and Chen
PF: lncRNA FOXD2-AS1 accelerates the progression of cervical cancer
via downregulating CDX1. Eur Rev Med Pharmacol Sci. 23:10234–10240.
2019.PubMed/NCBI
|
|
52
|
Zhang Q, Zheng J and Liu L: The long
noncoding RNA PCGEM1 promotes cell proliferation, migration and
invasion via targeting the miR-182/FBXW11 axis in cervical cancer.
Cancer Cell Int. 19:3042019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Guo Z, Wang X, Yang Y, Chen W, Zhang K,
Teng B, Huang C, Zhao Q and Liu Z: Hypoxic tumor-derived exosomal
long noncoding RNA UCA1 promotes angiogenesis via miR-96-5p/AMOTL2
in pancreatic cancer. Mol Ther Nucleic Acids. 22:179–195. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Cao Y, Xiong JB, Zhang GY, Liu Y, Jie ZG
and Li ZR: Long noncoding RNA UCA1 regulates PRL-3 expression by
sponging microRNA-495 to promote the progression of gastric cancer.
Mol Ther Nucleic Acids. 19:853–864. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhao J, Li L and Yang T: miR-216a-3p
suppresses the proliferation and invasion of cervical cancer
through downregulation of ACTL6A-mediated YAP signaling. J Cell
Physiol. 235:9718–9728. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pan SS, Zhou HE, Yu HY and Xu LH:
miR-195-5p inhibits the cell migration and invasion of cervical
carcinoma through suppressing ARL2. Eur Rev Med Pharmacol Sci.
23:10664–10671. 2019.PubMed/NCBI
|
|
57
|
Ji X, Guo H, Yin S and Du H: miR-139-5p
functions as a tumor suppressor in cervical cancer by targeting
TCF4 and inhibiting Wnt/β-catenin signaling. Onco Targets Ther.
12:7739–7748. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wang D and Hu Y: Long non-coding RNA PVT1
competitively binds microRNA-424-5p to regulate CARM1 in
radiosensitivity of non-small-cell lung cancer. Mol Ther Nucleic
Acids. 16:130–140. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhang G, He X, Ren C, Lin J and Wang Q:
Long noncoding RNA PCA3 regulates prostate cancer through sponging
miR-218-5p and modulating high mobility group box 1. J Cell
Physiol. 234:13097–13109. 2019. View Article : Google Scholar : PubMed/NCBI
|