The human body contains numerous different cell
types, which make up tissues and organs with specific functions
that play a role in ensuring sustainability. It was discovered long
ago that differentiated cells in some tissues, e.g., skin,
intestinal epithelium and blood, have a short lifecycle and are
incapable of self-renewal (1). Stem
cells are able to self-renew and possess developmental potency to
differentiate into numerous cell types of an organism. This finding
led to the concept of stem cells as small unspecialized cells in
the human body devoid of a number of phenotypic traits commonly
found in cells from adult tissues for maintaining static and
transient cell types (2). Potency
with each differentiation step classifies stem cells into
totipotent, pluripotent, multipotent, oligopotent and unipotent
stem cells (3). As potency
decreases, the possible cell types that stem cells can
differentiate into also decrease accordingly.
Stem cells are generally categorized into two main
groups: embryonic and nonembryonic (somatic stem cells). Embryonic
stem cells (ESCs) are pluripotent, while somatic stem cells, e.g.,
mesenchymal stem cells (MSCs), are multipotent (4,5). ESCs
were first isolated from mouse embryos (6), while MSCs were discovered in monolayer
cultures of guinea pig bone marrow (7). Following their initial discovery, human
stem cells were isolated and cultured, whereby ESCs were derived
from human blastocysts (8) while
MSCs were derived from human bone marrow (9). These achievements in isolating and
culturing human stem cells opened new possibilities to better
understand the basic molecular mechanisms behind human development
and differentiation, leading to potential new treatments for
various diseases. While the potential benefits of research on human
ESCs are immense, there is a major ethical issue to address, e.g.,
the derivation of human ESCs results in the destruction of an
embryo. In addition, reliance on human embryos may also lead to the
commodification and exploitation of women (10–12).
Indeed, the potential exploitation of women involving the donation
or sales of oocytes or embryos for research and the purposeful
creation of embryos for research remain huge ethical issues that
need to be addressed. This ethical dilemma negatively impacts the
benefit-to-risk ratio, and hence, research has moved towards
somatic stem cells instead. Despite the focus on ESCs, MSCs have
been extensively researched in clinical settings during the past
decade (13–24) because MSCs can be easily obtained and
cultured for clinical use from multiple tissue sources that are
easily accessible using minimally intrusive methods, reducing the
ethical dilemmas surrounding human stem cell research (25). Additionally, MSCs can differentiate
into a variety of cell types that confer pleiotropic effects when
used for therapeutic purposes (26).
MSCs were initially discovered in bone marrow, and studies have
reported that these stem cells can also be found in other postnatal
organs and tissues, e.g., brain, kidney, liver, lung, spleen,
adipose tissue, muscle, hair follicles, teeth, placenta, and
umbilical cord (27,28). The International Society for Cellular
Therapy (ISCT) defines three minimal criteria that need to be
fulfilled for MSCs to overcome the issue of different
characteristics due to isolation from different tissue types
(29):
1. MSCs must adhere to
plastic surfaces when cultured in vitro.
2. The surface anti-genes
CD73, CD90, and CD105 must be expressed by MSCs, while CD34, CD45,
CD14 or CD11b, CD79α or CD19, and HLA-DR surface molecules should
be absent.
3. MSCs must be able to
differentiate into different mesodermal cell types, e.g.,
adipocytes, chondrocytes, and osteoblasts, when cultured in
vitro under certain conditions.
In addition to these criteria, the ISCT recommended
three additional conditions in 2019 to further clarify the
nomenclature of MSCs to avoid confusion between mesenchymal stem
cells and mesenchymal stromal cells (30). The tissue-source origin of MSCs
should be documented to highlight tissue-specific properties, e.g.,
phenotypic, functional and secretome behaviour. Comprehensive in
vitro and in vivo data demonstrate the stemness of MSCs
associated with a robust matrix of functional assays that test the
functionality of MSCs in vitro and in vivo based on
their proposed utility.
Previous studies have reported that MSCs are
multipotent and capable of differentiating into cells of
mesodermal, ectodermal, and endodermal lineages (29,31–33).
This plasticity of MSCs and their self-renewal capacity make these
cells promising therapeutic targets for various diseases, including
cancer treatment and tissue regeneration. MSCs undeniably offer
immense potential in the field of medicine; however, the cells also
present potential danger due to their ability to differentiate into
tumour-associated fibroblasts (34–36),
which support tumour growth through their secretome (37,38) and
resistance to apoptosis (39). Due
to their conflicting role in cancer progression and regression,
efforts to utilize MSCs in anticancer therapies have been
unsuccessful. Therefore, it is important to understand the
underlying molecular mechanisms of MSCs to fully utilize their
therapeutic potential.
Significant advancements in DNA sequencing,
computational biology, and bioinformatics have been made to
identify transcriptional processes associated with the multipotency
of MSCs. Based on previous studies, cyclin L2 (CCNL2), stromal
cell-derived factor 1 (CXCL12), podocalyxin-like protein (PODXL),
and ubiquitin carboxyl-terminal hydrolase 1 (USP1) were identified
as four genes responsible for maintaining multipotency, chromosomal
integrity, and MSC functions (40–42).
CCNL2 was reported to inhibit proliferation and cell specialization
while promoting apoptosis upon upregulation in mouse embryonic
carcinoma P19 cells. In the same study, CCNL2-overexpressing P19
cells had a remarkably decreased S phase and reduced expression
levels of myocardial cell differentiation-related genes, e.g.,
cardiac actin, GATA binding protein 4 (GATA4), myocyte-specific
enhancer factor 2C (Mef2C), homeobox protein Nkx-2.5 (Nkx2.5), and
B-type natriuretic peptide (BNP) (43). On the other hand, CXCL12 is a
chemokine protein that induces the migration of stem cells. It
functions by binding to CXC chemokine receptor (CXCR) 4, CXCR7 and
atypical chemokine receptor 3 (ACKR3) (44,45).
CXCL12 has been reported to be responsible for cell survival,
growth and migration during tissue/organ development (46). While the exact mechanism by which
CXCL12 helps maintain the stemness of MSCs has not been elucidated,
there are numerous reports on its function in other stem cells. The
CXCL12-CXCR4 axis was found to be responsible for cell migration,
while the CXCL12-CXCR7 axis promotes cell adhesion in cardiac stem
cells. Similar findings also reported the importance of
CXCL12-mediated CXCR4 signalling in controlling the position of
haematopoietic stem cells in bone marrow niches, which contain
limiting lymphoid-instructive cytokines that are responsible for
the multipotency of HSCs and their maintenance (47). A study confirmed that CXCL12-mediated
CXCR4 signalling promotes the proliferation, survival, and
migration of mesenchymal stromal cells in vitro (48). It is also likely that CXCL12 acts
through a similar mechanism to help MSCs maintain their
stemness.
The niche microenvironment strongly influences the
behaviour of stem cells. As mentioned, CXCL12 maintains
multipotency by directing MSCs to specific niches, where secreted
factors influence their self-renewal and stemness (53). This phenomenon indicates that the
behaviour of MSCs is determined by the interaction between
intrinsic transcriptional genes and extrinsic factors of the
environment. It has been established that the protein kinase B
(Akt) and extracellular-signal-regulated kinase (Erk) signalling
pathways control both stem cell proliferation and survival, while
the Wnt, Notch, and Sonic hedgehog (Shh) signalling pathways
regulate stem cell renewal and differentiation (54–57). A
study also proposed two novel mechanisms that help to maintain the
stemness of MSCs via the scrapie responsive gene 1 (SCRG1)/bone
marrow stromal cell anti-gene 1 (BST1) ligand-receptor combination
and cell-cell adhesion through N-cadherin (52). An improved understanding of the
underlying mechanism involved in stem cell renewal and
differentiation is important because the original abilities are
lost at a high rate during long-term in vitro culture
(58,59). Therefore, current work should develop
novel techniques to ensure that MSCs maintain their multipotency
despite long-term in vitro culture. This would, in turn,
maintain the potential of MSCs to be used in regenerative medicine
and cell therapy.
Epigenetic factors influence the differential gene
expression in MSCs that causes cell differentiation. Hence, the DNA
sequences of MSCs and their specialized cell types are similar,
with almost no difference. Commonly studied epigenetic
modifications include DNA methylation and histone modification,
e.g., methylation, acetylation, ubiquitylation, and microRNAs. Once
epigenetic modifications occur, gene expression can be influenced
by changing the availability of gene promoters, thus affecting the
recruitment of supplementary chromatin-modifying enzymes or
transcriptional regulators that drive stem cell differentiation
(60). For example, runt-related
transcription factor 2 (Runx2) regulates most osteoblast-specific
genes by working together with numerous coactivators and
corepressors that alter the binding of Runx2 to the osteocalcin
promoter. This binding modification occurs through DNA methylation
and acetylation of histones H3 and H4 (61). Additionally, Runx2 changes the
expression of its target in response to other signals, e.g.,
transforming growth factor-beta (TGF-β), bone morphogenetic protein
(BMP) and Wnt signalling pathways (60), is responsible for the osteogenic
lineage. MSCs can also undergo adipogenic differentiation, whereby
hypomethylation of the genes encoding peroxisome
proliferator-activated receptors gamma-2 (PPARγ2), fatty
acid-binding protein 4 (FABP4), leptin (lep) and lipoprotein lipase
(lpl) was reported to be responsible for these mechanisms (61,62).
In addition to secreted factors, the cyclic tensile
strain that can alter cell behaviour should be considered another
microenvironmental factor. MSCs have been observed to lose
multipotency and spontaneously differentiate after prolonged
passaging in vitro (25,63).
Therefore, in vitro culture conditions must be optimized to
maintain the multipotency of MSCs for their therapeutic potential
in clinical settings. A study found that low actomyosin
contractility induced by restricting the cells to small islands
during initial culture is necessary to ensure the stemness of MSCs
(64). A disparity in differential
gene expression when MSCs are cultured in 2D and 3D culture systems
is likely due to the interaction between the cells in an intricate
3D structure compared to that in a monolayer 2D culture (65). Recent studies have also found that
cyclic tensile strain promotes bone marrow-derived MSCs (BMSCs) to
differentiate into cardiomyocyte-like cells (66) and adipose stem cells to differentiate
into the osteogenic lineage (67).
However, the regulatory pathways and epigenetic factors that might
be involved seem to depend on the source of MSCs and the desired
cell lineage.
MSCs have been the subject of clinical trials for
the past decade, but the outcomes have fallen short of expectations
despite promising data in animal models. Studies continue to
emerge, as there is no denying the potential of MSCs to treat a
wide variety of human afflictions, e.g., neurodegeneration, ageing,
blindness, diabetes, and cancers (1). It is crucial to realistically assess
the time and effort required to establish new clinical settings for
numerous therapeutic applications. The same concern regarding the
efficacy and safety of treatment must also always be at the
forefront when considering the usage of MSCs, as there are crucial
biological and pharmacological discrepancies in preclinical and
clinical studies. The first clinical trial using MSCs as a
therapeutic agent was in 1995 (68).
Since then, MSCs have become the most widely clinically studied
cell-based therapy worldwide (69).
MSCs are currently classified as advanced therapy medicinal
products (ATMPs), which follow the Good Manufacturing Practices
(GMP) guidelines of the Food and Drug Administration (FDA) and the
European Medicines Agency (EMA) to authenticate and ensure the
quality of cells before their administration to patients (70). This compliance with GMP includes the
sources of MSCs, reagents, equipment, packaging materials,
procedures, laboratory staff, environment, and final cellular
medicine (71).
It is of the utmost importance that GMP conditions
are maintained according to the international and national
medicinal governing framework. This act ensures the quality of the
administered MSCs and prevents possible contamination issues that
may cause adverse reactions in patients and even death. However,
there is currently a lack of unified and standard criteria for
manufacturing MSCs as a therapeutic agent due to some differences
over specific issues depending on the USA, Europe, Canada,
Singapore, Japan and so forth. Despite this challenge, consistent
physical and microbiological testing of the MSC production
laboratory and cleanrooms to ensure the sterility of the production
process is also warranted (72).
This act fulfils the requirement of International Standard
Organization (ISO) standard 14644.
Currently, 1,088 studies registered as clinical
trials list MSCs as a clinical intervention. The majority of these
trials, whether ongoing or completed, are phase 1 or 2 studies that
evaluate the safety and efficacy of MSCs in humans. Despite the
most promising results, MSC-based therapies still have significant
limitations due to the nature of the stem cells, e.g., MSCs
markedly differ in gene expression profile, cell differentiation
ability, growth rate, and therapeutic capacity, depending on their
tissue source (63). Therefore, it
may be vital to isolate and culture homogenous populations of MSCs
to improve the efficacy and safety of the treatment. The method of
transplanting MSCs isolated and grown in large batches from
unrelated donor tissues is known as allogeneic transplantation; in
contrast to autologous therapy, MSCs are extracted and grown from
treated patients. The benefits of allogeneic transplantation
include:
1. Efficiency, such as
the isolation, expansion, and validation of MSCs from the patient,
is not required.
2. The therapeutic
functions of allogenic MSCs remain the same, unlike autologous
MSCs, which have been reported to have impaired functions when
isolated from elderly individuals (73,74).
3. A well-established
stock of MSCs following strict GMP requirements reduces the
variability of donors and improves the success rate of the
treatment.
In contrast, autologous transplantation, which
triggers less risk of immunogenic response, is an alternative.
Autologous MSCs are easily available without identifying a suitable
donor (81). Autologous MSCs also
overcome the limitation of long-term in vitro culture for
allogeneic MSCs, leading to loss of multipotency, morphological
changes, and an increased risk of malignancy (25,74).
Nonetheless, the challenge and reliance on autologous MSC
transplantation mean that a well-optimized and established protocol
for the isolation and ex vivo preparation of MSCs will be
required. Such precise standardization may be difficult, as several
exogenous factors greatly affect the biological properties of MSCs
(70). Autologous MSCs may not be
suitable for treating certain genetic diseases due to the mutations
present in stem cells. Flaws in the genetic sequence hinder both
the immunomodulatory function and regenerative traits of MSCs. For
example, MSCs isolated from patients suffering from systemic lupus
erythaematosus have a senescent phenotype with diminished
capabilities to differentiate, migrate and regulate the immune
system (79,82–84).
Therefore, more preclinical and clinical studies are required to
obtain more information related to the utility of MSCs as a
therapeutic approach. Supplementary studies on the basic biology of
MSC maintenance and the regulators of MSC differentiation would
also provide a clearer picture of how to better administer MSCs as
therapeutic agents in the future.
Most of the published clinical studies employing
MSCs for diseases have specific treatments with positive outcomes.
In neurology, ischaemic stroke patients treated with MSCs yielded
positive results, whereby the patients showed significantly
improved neurological and motor functions (85–88).
Among all of the studies conducted, serious adverse events that
were reported included transient ischaemic attack, seizure,
asymptomatic subdural haematoma/hygroma, urinary tract infection,
sepsis, pneumonia, hyperglycaemia, neutrophilia, shingles,
ischaemic stroke, cellulitis, muscle cramps, fracture neck femur,
and peripheral vascular disease (89). However, these side effects were
attributed to the procedure rather than cell therapy. The study
also reported promising results in the field of cardiology. Studies
have shown that diseases, e.g., dilated cardiomyopathy and
ischaemic or nonischaemic heart failure, have had clinical and
pathophysiological improvements; no serious adverse effects were
reported, demonstrating the treatment's safety profile (19,90–92).
Patients suffering from cartilage lesions and/or osteoarthritis,
especially in the knee, were reported to have a clinical
improvement in pain, stiffness, and functionality when treated with
MSCs. These results show the broad potential of MSCs for clinical
usage with no serious adverse effects linked to cell therapy.
Interest in developing MSCs as therapeutic agents
has not waned in the slightest, despite the obstacles faced,
largely due to their immense therapeutic potential. In addition to
being multipotent with self-renewing capabilities, MSCs also have
the added benefits of migrating to the injury site and promoting
tissue regeneration (26). This
phenomenon means that MSCs can be a form of personalized therapy
(when opting for autologous therapy) that is site-directed,
promotes tissue restoration, and replaces damaged cells through
differentiation. It is, therefore, unsurprising that scientists are
so invested in advancing this field of research since the
therapeutic agent reaches the targeted tissue for effective disease
treatment. As MSCs have a natural tendency to be attracted towards
damaged sites and the tumour microenvironment, the cells are a
prime candidate for further investigation, as MSCs seem to be
independent of the type of tumour, immunocompetence and delivery
route (93).
Insight into the mechanism underlying the
mobilization of MSCs to the injury site is still limited, but
CXCL12-mediated CXCR4 signalling is most likely involved as a
pathway that mediates cell migration (94). Secreted chemokines can mediate
inflammation in the tumour microenvironment, and wounds are
responsible for attracting MSCs (95). As the chemotactic properties of MSCs
seem to be similar to those of other immune cells, the established
model of leukocyte migration can be used as a template to study the
factors involved in MSC migration (95). Other chemokine receptors that react
to signals from the injury site or tumour microenvironment induce
CCR1-2, CXCR1-2, CCR4, CXCR4-6, CCR7-10, and CX3R1 expression in
MSCs (95). In addition, cell
adhesion molecules expressed by MSCs, e.g., CD44, CD49d, CD54,
CD102 and CD106, are thought to be involved in MSC migration to
injury sites (26,96).
A wide variety of trophic mediators and growth
factors are secreted to initiate tissue regeneration once MSCs
arrive at the injury site. The pleiotropic effects conferred by
MSCs towards damaged tissues include anti-inflammation,
immunomodulation, and enhanced cell survival and angiogenesis
(97,98). Among these therapeutic effects,
anti-inflammation and immunomodulation are key elements that make
MSCs an attractive target to study because the immune system plays
an integral role in regulating tissue repair and regeneration
through healing, scarring and fibrosis (99). The immunomodulatory process of MSCs
occurs through the secretion of several soluble factors that
interfere with the immune system, and the inflammation process
takes place through cell-cell interactions (100,101).
The immunosuppressive effect of MSCs was enhanced by increasing the
binding between MSCs and T-cells through intercellular adhesion
molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1)
(102). A similar phenomenon was
reported when MSCs were shown to heighten the suppressive
regulation of T-cells and macrophages regarding proinflammatory
macrophages (103).
The flexibility of multipotent MSCs to differentiate
into a wide variety of cells would then allow the cells to replace
damaged or dead cells. However, reports on this mechanism are
inconclusive, as the engraftment of MSCs is transient, and instead,
MSCs secrete specific factors that grow and differentiate into
local precursor cells (26). The
potential of MSCs in tissue repair and regeneration is undeniable,
regardless of the exact mechanisms.
Over the years, multiple reports have been published
that strongly suggest the mechanism of action of MSCs. These
actions are mainly attributed to the ability to migrate to the
injury site (104–106), the paracrine effect of the
secretome (107,108), and the immunomodulatory ability
(109,110). The benefits of MSCs are enticing,
and it is important to consider the potential side effects and
major risk factors that are often associated with stem cell
transplantation. There have been contradictory results in
describing the anti- and pro-tumour effects of MSCs. As mentioned
above, the therapeutic role of MSCs in cancer therapy is similar to
that in other diseases; tumours secrete similar chemoattractants to
damaged tissues, which initiate the migration of MSCs to the target
site through the CXCL12-CXCR4 signalling pathway (111–114).
MSCs have also been reported to interact with cancer cells,
directly and indirectly, affecting tumour development (26). Moreover, MSCs secrete various
cytokines and growth factors, which alter cellular activities,
e.g., cell proliferation (cell cycle), angiogenesis, cell survival,
and immunomodulation, to indirectly influence tumour growth. For
example, BMSCs were described to enhance the proliferation of
B16-LacZ cells and increase tumour size when both cell lines were
coinjected into syngeneic mice via enhanced angiogenesis (115). In contrast, BMSCs were also
reported to inhibit proliferation, migration, and invasion and
induce cell cycle arrest, which led to apoptosis of human glioma
U251 cells by downregulating the PI3K/Akt pathway (116).
Indeed, such paradoxical results are not uncommon,
as divergent effects on cell growth, invasion, and migration have
been reported when MSCs sourced from the human umbilical cord were
cocultured with glioblastoma cancer stem cells, e.g., direct
contact between both cell lines caused an inhibitory response
(117). At the same time, the
release of soluble factors triggered a stimulatory reaction
(117). Similar opposing effects
were observed during an in vivo study investigating whether
coinjection and distant injection of MSCs with breast tumour 4T1
cells exerted different effects on tumour growth (118). Coinjection supported tumour growth,
while in the distant injection model, it inhibited tumour growth by
promoting host antitumour immunity (118). Likewise, MSCs derived from
umbilical cord blood and adipose tissue also had divergent effects
on the proliferation of glioblastoma multiforme. The former
inhibited and promoted the proliferation process (119).
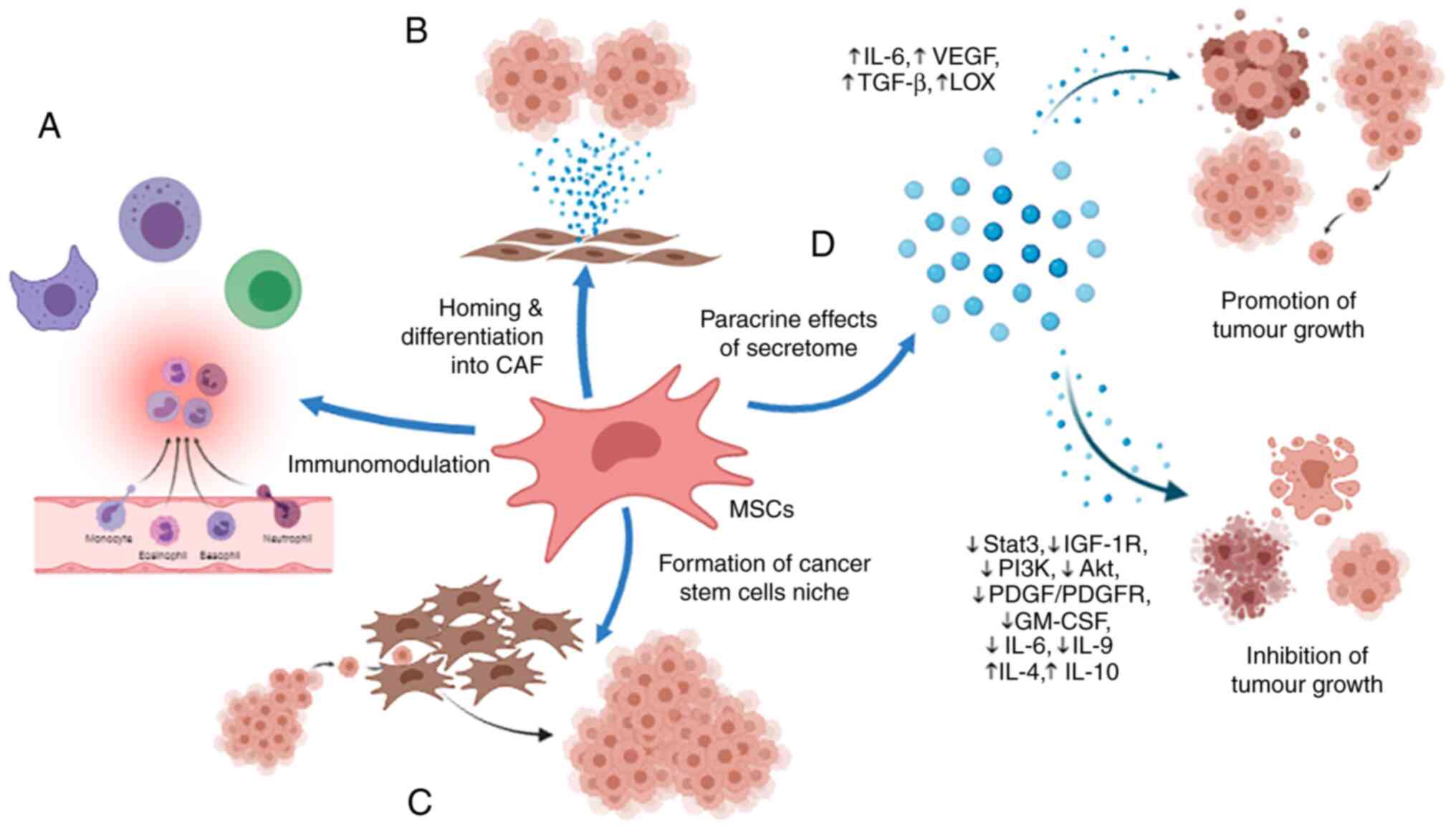
Several studies have found that upon being recruited
to tumour sites, the multipotency of MSCs enables their
self-differentiation into carcinoma-associated fibroblasts, which
directly contribute to cancer progression (120–122).
In addition, MSCs were reported to promote tumour growth and
angiogenesis through the secretion of proangiogenic cytokines,
e.g., interleukin (IL)-6, vascular endothelial growth factor
(VEGF), and transforming growth factor-β (TGF-β) (123–125)
(Fig. 1). MSCs also enhanced the
metastasis of human breast cancer cells by promoting de novo
production of lysyl oxidase (LOX) by cancer cells (126). In addition, MSCs are able to
modulate the production of regulatory T-cells and inhibit the
activity of natural killer (NK) cells and cytotoxic T lymphocytes
(CTLs), protecting breast cancer cells from the immune system
(127). Similar immunosuppressive
effects were observed when MSCs were reported to promote lung
cancer metastasis (128). It was
suggested that MSCs have the ability to form a cancer stem cell
niche in vivo where tumour cells can preserve the potential
to proliferate, thus sustaining the malignant process (129).
In contrast, MSCs increased the sensitivity of
breast cancer cells to radiotherapy and impeded tumour progression
by downregulating the signal transducer and activator of
transcription 3 (Stat3) signalling pathway (130). Another study found that MSCs
hampered hepatic cancer growth through the secretion of paracrine
factors that lowered the insulin-like growth factor 1 receptor
(IGF-1R), phosphatidylinositol 3-kinase (PI3K) and Akt signalling
pathways (131). In addition,
microRNA-4461 isolated from BMSCs was reported to inhibit tumour
pathogenesis in colorectal cell lines and tissues by downregulating
the expression of COPB2 (132).
MSCs also inhibited vascular growth in glioma cells by
downregulating the platelet-derived growth factor (PDGF)/PDGFR axis
(133). Antiproliferative effects
and apoptosis were observed when ovarian cancer cell lines were
cocultured with conditioned media of MSCs derived from human bone
marrow, adipose tissue, and umbilical cord (134). The study found that the conditioned
media of MSCs showed an increase in IL-4 and IL-10 but a decrease
in granulocyte/macrophage colony-stimulating factor (GM-CSF), IL-6,
and IL-9. It is undeniable that anti-inflammatory cytokines play an
important role in cancers (135–137).
However, controversial findings have been reported regarding
whether cytokines support or hinder tumour progression (138–141).
Regardless, MSCs have been shown to modulate the immune response
through the balanced secretion of proinflammatory and
anti-inflammatory cytokines (142).
Therefore, this duality of function found in the secretome of MSCs
and the complex cell-to-cell interaction between MSCs and cancer
cells might be the reason for the conflicting reports regarding the
role of MSCs in cancers.
Although the underlying mechanisms are not yet fully
understood, there is a consensus that the differences in
experimental design, e.g., tumour models used, route of cell
administration, control group, tissue source, dosage use, and
timing of the treatment that may affect the final results, should
be considered (37,117–119,143,144).
Research should not make conclusions about the utility of MSCs in
cancer therapy based on a single study. Instead, standardized
protocols should be established to ensure that the data obtained
are more comparable to understand the interaction of MSCs with
cancer cells. Additionally, precautions should be taken before the
clinical introduction of MSCs for treating cancers since the
heterogeneous characteristics of MSCs are easily susceptible to
different pathological conditions present in patients, which can
hinder the therapeutic mechanisms.
Numerous studies have been conducted to explore the
possibility of enhancing the inherent therapeutic properties of
MSCs using genetic engineering. These studies mainly focused on
four crucial points: improving migration, adhesion, and
survivability while reducing the cell senescence of transplanted
MSCs (160–162). This phenomenon is accomplished by
inserting a vector loaded with a constructed genetic cassette into
MSCs; the cassette expresses certain genes constantly or can be
controlled with a gene switch (163). For example, adipose-derived MSCs
(AdMSCs) were transduced with a retroviral vector to upregulate the
expression of CXCR4. The study reported that the transduced MSCs
showed increased motility, invasion, and placement in the bone
marrow when injected into nonobese diabetic/severe combined
immunodeficiency (NOD/SCID) mice (164). In addition to CXCR4, other genes
involved in MSC migration, e.g., aquaporin-1, can be modified. It
was reported that the overexpression of aquaporin-1 and CXCR4
promoted the migratory ability of MSCs via the Akt and Erk pathways
(165). MSCs have also been
genetically engineered to overexpress integrin-linked kinase (ILK).
The study found that genetically modified MSCs had 1.5-fold higher
survivability and a 32.3% higher adhesion rate when engrafted into
an ischaemic myocardium model, with a higher retention rate of
~4-fold (166). In addition, BMSCs
and AdMSCs were reported to have increased proliferation and
differentiation potential when engineered to overexpress Oct4 and
Sox2 (167,168). Genetic engineering has the
potential to circumvent the current problems that limit the
application of MSCs in clinical settings and improve their
potential therapeutic properties. Despite the immense benefits,
this technique also has potential drawbacks, e.g., the risk of
insertional oncogenesis due to viral vectors to introduce plasmid
DNA, adverse immune reactions, and high production costs (169). Great precautions should be taken
when considering the use of genetically modified MSCs for cancer
therapy.
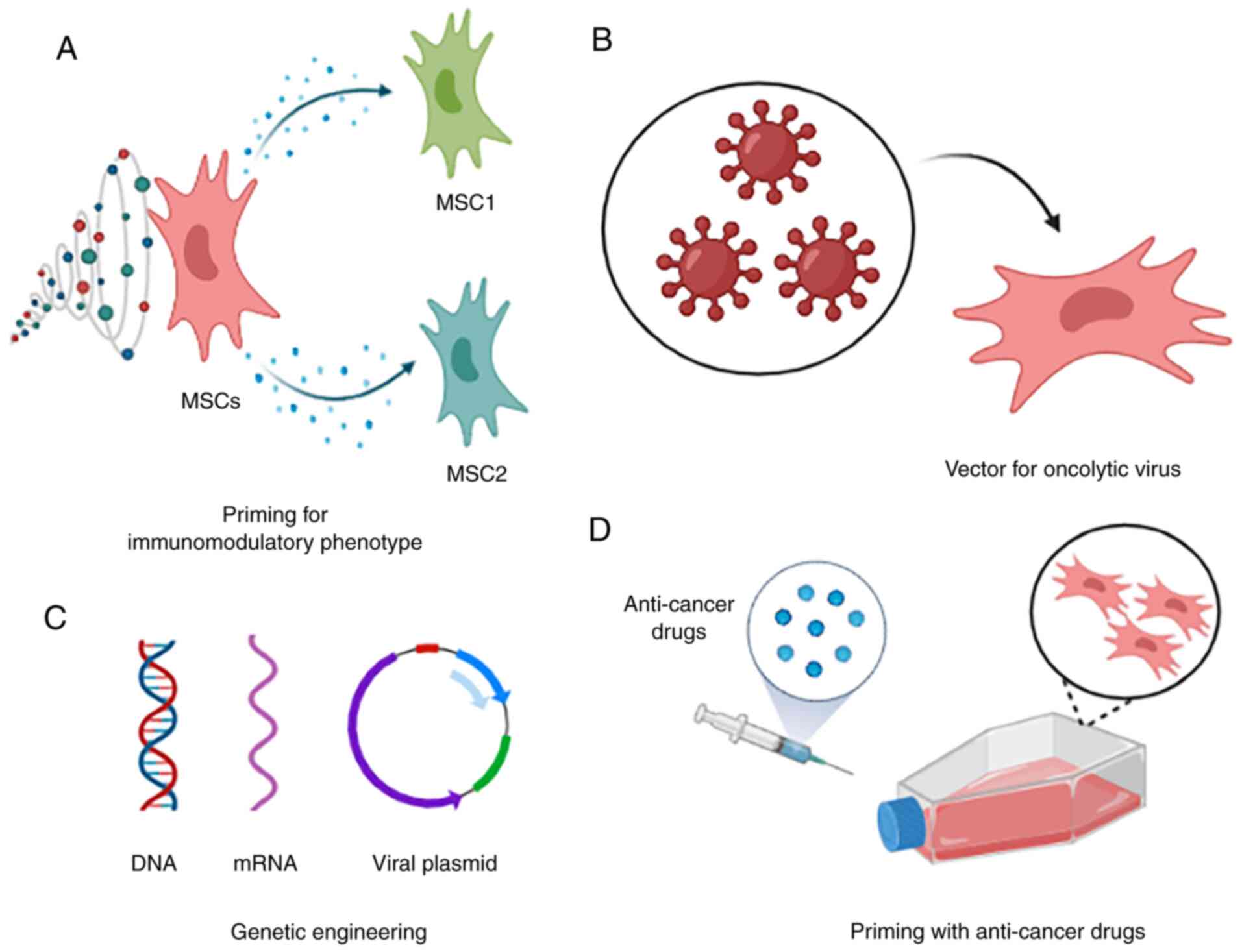
In addition, previous studies have established a
connection between specific Toll-like receptors (TLRs) and the
immunomodulatory properties of MSCs (170–172).
Interestingly, a study reported that TLR-4-primed MSCs (MSC1)
exhibited a proinflammatory phenotype, while TLR-3-primed MSCs
(MSC2) secreted immunosuppressive mediators (173). Indeed, the polarization of MSCs
into specific immunomodulatory phenotypes is a promising strategy
as well. For example, macrophages cocultured with MSCs showed
evidence of alternatively activated macrophages with high levels of
CD206 and IL-10 but low levels of IL-12, which displayed a higher
level of phagocytic activity (174). Studies have also reported that
TL-3- and TL-4-primed MSCs preserved and enhanced the function of
neutrophils through the combined action of IL-6, IFN-β, and GM-CSF
(175,176). Furthermore, MSC1 was observed to
recruit lymphocytes by activating T-cells and secreting macrophage
inflammatory protein-1 (MIP-1), CCL5, CXCL9, and CXCL9 (177). In contrast, MSCs can change
macrophages from a TNFα-secreting MSC1 phenotype to an
immunosuppressive IL-10-expressing phenotype through a
prostaglandin-(PGE-)2-based mechanism (178). MSCs have also been reported to
inhibit IL-2-induced NK cell proliferation and prevent the
initiation of effector functions, e.g., cytotoxic activity and
cytokine production, with the production of the soluble factors
indoleamine 2,3-dioxygenase (IDO) and prostaglandin E2 (PGE2)
(179). MSCs influence tumour
growth through immunomodulation, and as discussed earlier, the
polarization of MSCs for cancer treatment warrants further
investigation. After all, it is widely accepted that chronic
inflammation is a critical hallmark of cancer that elevates the
risk of malignancy (180). The
anti-inflammatory cytokines secreted by MSCs can circumvent these
effects. On the other hand, tumour cells evade the immune system by
avoiding immune recognition and developing an immunosuppressive
microenvironment (181), which can
be overcome with the help of MSCs boosting the innate immune
system. Therefore, careful and purposeful polarization will benefit
the field of cancer therapy and facilitate manipulation of the
immunomodulatory capacity of MSCs.
Studies have also investigated the potential of MSCs
to act as vectors for oncolytic viruses. For example, MSCs were
used as vectors to deliver oncolytic herpes simplex virus to human
brain melanoma metastasis models grown in immunodeficient and
immunocompetent mice. This study reported that the intervention
significantly prolonged the life of the mice through
immunomodulatory actions compared to the control group (182). A recent in vivo study also
explored the possibility of using MSCs derived from menstrual blood
as a vector for CRAd5/F11 chimaeric oncolytic adenovirus to treat
colorectal cancer. It was reported that the chimaeric oncolytic
adenovirus was successfully delivered and accumulated at the tumour
site, and it inhibited tumour growth (183). A mathematical model to
quantitatively predict the efficacy of MSCs acting as vectors for
virotherapeutic agents in vivo has been developed,
indicating that MSCs are a promising strategy that improves the
efficacy and safety profile of the treatment (184).
MSCs can also be primed with anticancer drugs for
targeted delivery due to their preferential migration towards the
tumour site and relative resistance to cytostatic and cytotoxic
drugs (185–187). For example, MSCs acquire strong
antitumour activity after packaging and delivering paclitaxel (PTX)
through extracellular vesicles (188). The same study also demonstrated
that it is possible to produce drugs with higher cell-target
specificity by utilizing MSCs as a factory to package the drugs.
Similar studies reported that MSCs isolated from different sources
were primed with PTX and tested against different cancer cell lines
(187,189–191).
Other drugs were also tested for priming MSCs, e.g., doxorubicin
and gemcitabine. A study reported similar results whereby MSCs
effectively incorporated the active form of the drugs and released
sufficient quantities to produce a significant inhibition of
squamous cell carcinoma growth in vitro (192). Researchers have explored the
possibility of using nanoparticles to improve the payload and
delivery capacity of MSCs (193,194).
All of these studies indicate that MSCs are able to take up and
subsequently release drugs in a targeted and gradual manner, which
improves the efficacy of anticancer drugs.
Due to the short half-life of most anticancer drugs
in the body and their high toxicity to healthy cells, direct
administration of these drugs is often associated with unwanted
side effects. For example, nausea and vomiting, tiredness, changes
in taste, dry mouth, loss of appetite, constipation, and hair loss
are common side effects faced by chemotherapy patients (195). Thus, using MSCs as vectors to
deliver therapeutic proteins or anticancer drugs can help to solve
this issue advantageously. MSCs can exert therapeutic effects
locally due to selective migration and accumulation in tumour
sites, increasing treatment efficacy and reducing systemic
toxicity. Currently, divergent drugs are being investigated for
different cancer therapeutic purposes. For example, MSCs were
reported to enhance the therapeutic capabilities of tendon repair
when pretreated with pioglitazone (196). Other studies using pioglitazone as
the priming agent also found similar results, where pretreated MSCs
had greater therapeutic effects on lung regeneration in an
emphysema mouse model (197,198).
Pioglitazone has been administered indirectly to breast cancer
cells via stem-and-cancer cell interaction (199). Through this process, modified and
viable pretreated stem cells are subsequently administered to
patients, and pretreated stem cells are allowed to interact with
cancer cells in the patients' bodies. Considering that pioglitazone
has been reported to possess anticancer effects (200–202),
it may be beneficial to examine the possibility of priming MSCs
with pioglitazone for cancer therapy. After all, using MSCs
pretreated with pioglitazone as a strategy to improve the overall
therapeutic effects, as reported in our study (199), remains rare. Despite the study on
cardiomyogenic transdifferentiation and cardiac function (203), as mentioned above, MSCs pretreated
with pioglitazone for cancer therapy remain to be characterized. A
similar strategy was conducted using AdMSCs pretreated with a
peroxisome proliferator-activated receptor gamma (PPARγ) agonist to
improve the regeneration effects in an elastase-induced emphysema
mouse model (197). Indeed, human
umbilical cord-derived mesenchymal stem cells pretreated with IL-6
were also found to abolish the stem cell growth-promoting effect on
gastric cancer cells (204). The
potential therapeutic strategies of MSCs in cancer therapy are
summarized in Fig. 2.
Although the potential benefit is undeniable, there
are potential risks in using MSCs for cancer treatments. These
risks can be categorized as acute issues, e.g., inflammatory
reaction or embolic phenomenon, intermediate issues, e.g.,
graft-versus-host disease (GVHD) or secondary infection, or
long-term issues, e.g., risk of tumour growth (142). It was reported in a clinical study
that patients treated with MSCs commonly died due to infection
(205). This phenomenon, coupled
with the fact that MSCs can potentially promote tumour growth
instead of inhibiting it, as previously discussed, makes it a risky
treatment option. However, more studies must be conducted to
provide future evidence and improve the therapeutic effects of
modified MSCs in cancer treatments. These cells hold great
potential to revolutionize the current cancer therapies that are
available.
It is undeniable that stem cells are promising
therapeutic alternatives for numerous human diseases. While the
motivation to benefit human health is noble, researchers should
take precautions in this field to prevent the potential
exploitation of vulnerable groups. Efforts should also be directed
towards using MSCs in autologous and allogeneic transplantation, as
they do not raise the same ethical concerns as ESCs. In addition,
MSCs benefit from their ability to carry anticancer payloads
through genetic manipulation or pretreatment of the cells, leading
to use in regenerative medicine and potentially oncology.
Therefore, it is important to obtain as much information as
possible to ensure that stem cell-based therapy is reliable,
effective, efficient, safe, and affordable. It should be developed
with the physiological condition of the patients in mind to truly
benefit humanity.
Not applicable.
The present project was funded by the Exploratory
Research Grant Scheme Fasa 1/2013 (grant no.
203/CIPPM/6730098).
Not applicable.
SKL and BYK contributed to the conception and design
of the study. SKL drafted the manuscript and BYK revised the
manuscript. Both authors have read and approved the final
manuscript. Data sharing is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Watt FM and Driskell RR: The therapeutic
potential of stem cells. Philos Trans R Soc B Biol Sci.
365:155–163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alvarez CV, Garcia-Lavandeira M,
Garcia-Rendueles MER, Diaz-Rodriguez E, Garcia-Rendueles AR,
Perez-Romero S, Vila TV, Rodrigues JS, Lear PV and Bravo SB:
Defining stem cell types: Understanding the therapeutic potential
of ESCs, ASCs, and iPS cells. J Mol Endocrinol. 49:R89–R111. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zakrzewski W, Dobrzyński M, Szymonowicz M
and Rybak Z: Stem cells: Past, present, and future. Stem Cell Res
Ther. 10:682019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh VK, Saini A, Kalsan M, Kumar N and
Chandra R: Describing the stem cell potency: The various methods of
functional assessment and in silico diagnostics. Front Cell Dev
Biol. 4:1342016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pittenger MF, Discher DE, Péault BM,
Phinney DG, Hare JM and Caplan AI: Mesenchymal stem cell
perspective: Cell biology to clinical progress. NPJ Regen Med.
4:222019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Martin GR: Isolation of a pluripotent cell
line from early mouse embryos cultured in medium conditioned by
teratocarcinoma stem cells. Proc Natl Acad Sci USA. 78:7634–7638.
1981. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Friedenstein AJ, Chailakhjan RK and
Lalykina KS: The development of fibroblast colonies in monolayer
cultures of guinea-pig bone marrow and spleen cells. Cell Tissue
Kinet. 3:393–403. 1970.PubMed/NCBI
|
|
8
|
Thomson JA, Itskovitz-Eldor J, Shapiro SS,
Waknitz MA, Swiergiel JJ, Marshall VS and Jones JM: Embryonic stem
cell lines derived from human blastocysts. Science. 282:1145–1147.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Haynesworth SE, Goshima J, Goldberg VM and
Caplan AI: Characterization of cells with osteogenic potential from
human marrow. Bone. 13:81–88. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
McLeod C and Baylis F: Feminists on the
inalienability of human embryos. Hypatia. 21:1–14. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Caulfield T and Ogbogu U: Stem cell
research, scientific freedom and the commodification concern. EMBO
Rep. 13:12–16. 2012. View Article : Google Scholar
|
|
12
|
Marway H, Johnson SL and Widdows H:
Commodification of human tissue. Handbook of Global Bioethics. ten
Have H.A.M.J and Gordijn B: Springer Netherlands; Dordrecht,
Netherlands: pp. 581–598. 2014, View Article : Google Scholar
|
|
13
|
Lee JS, Hong JM, Moon GJ, Lee PH, Ahn YH
and Bang OY; STARTING collaborators, : A long-term follow-up study
of intravenous autologous mesenchymal stem cell transplantation in
patients with ischemic stroke. Stem Cells. 28:1099–1106. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bhasin A, Srivastava MVP, Kumaran SS,
Mohanty S, Bhatia R, Bose S, Gaikwad S, Garg A and Airan B:
Autologous mesenchymal stem cells in chronic stroke. Cerebrovasc
Dis Extra. 1:93–104. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Honmou O, Houkin K, Matsunaga T, Niitsu Y,
Ishiai S, Onodera R, Waxman SG and Kocsis JD: Intravenous
administration of auto serum-expanded autologous mesenchymal stem
cells in stroke. Brain. 134((Pt 6)): 1790–1807. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Connick P, Kolappan M, Crawley C, Webber
DJ, Patani R, Michell AW, Du MQ, Luan SL, Altmann DR, Thompson AJ,
et al: Autologous mesenchymal stem cells for the treatment of
secondary progressive multiple sclerosis: An open-label phase 2a
proof-of-concept study. Lancet Neurol. 11:150–156. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weiss DJ, Casaburi R, Flannery R,
LeRoux-Williams M and Tashkin DP: A placebo-controlled, randomized
trial of mesenchymal stem cells in COPD. Chest. 143:1590–1598.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Götherström C, Westgren M, Shaw SWS,
Aström E, Biswas A, Byers PH, Mattar CNZ, Graham GE, Taslimi J,
Ewald U, et al: Pre- and postnatal transplantation of fetal
mesenchymal stem cells in osteogenesis imperfecta: A two-center
experience. Stem Cells Transl Med. 3:255–264. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Heldman AW, DiFede DL, Fishman JE,
Zambrano JP, Trachtenberg BH, Karantalis V, Mushtaq M, Williams AR,
Suncion VY, McNiece IK, et al: Transendocardial mesenchymal stem
cells and mononuclear bone marrow cells for ischemic
cardiomyopathy: The TAC-HFT randomized trial. JAMA. 311:62–73.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Karantalis V, DiFede DL, Gerstenblith G,
Pham S, Symes J, Zambrano JP, Fishman J, Pattany P, McNiece I,
Conte J, et al: Autologous mesenchymal stem cells produce
concordant improvements in regional function, tissue perfusion, and
fibrotic burden when administered to patients undergoing coronary
artery bypass grafting: The prospective randomized study of
mesenchymal stem cell therapy in patients undergoing cardiac
surgery (PROMETHEUS) trial. Circ Res. 114:1302–1310. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rushkevich YN, Kosmacheva SM, Zabrodets
GV, Ignatenko SI, Goncharova NV, Severin IN, Likhachev SA and
Potapnev MP: The use of autologous mesenchymal stem cells for cell
therapy of patients with amyotrophic lateral sclerosis in Belarus.
Bull Exp Biol Med. 159:576–581. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Thakkar UG, Trivedi HL, Vanikar AV and
Dave SD: Insulin-secreting adipose-derived mesenchymal stromal
cells with bone marrow-derived hematopoietic stem cells from
autologous and allogenic sources for type 1 diabetes mellitus.
Cytotherapy. 17:940–947. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Vega A, Martín-Ferrero MA, Del Canto F,
Alberca M, García V, Munar A, Orozco L, Soler R, Fuertes JJ, Huguet
M, et al: Treatment of knee osteoarthritis with allogeneic bone
marrow mesenchymal stem cells: A randomized controlled trial.
Transplantation. 99:1681–1690. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fernández O, Izquierdo G, Fernández V,
Leyva L, Reyes V, Guerrero M, León A, Arnaiz C, Navarro G, Páramo
MD, et al: Adipose-derived mesenchymal stem cells (AdMSC) for the
treatment of secondary-progressive multiple sclerosis: A triple
blinded, placebo controlled, randomized phase I/II safety and
feasibility study. PLoS One. 13:e01958912018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Musiał-Wysocka A, Kot M and Majka M: The
pros and cons of mesenchymal stem cell-based therapies. Cell
Transplant. 28:801–812. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hmadcha A, Martin-Montalvo A, Gauthier BR,
Soria B and Capilla-Gonzalez V: Therapeutic potential of
mesenchymal stem cells for cancer therapy. Front Bioeng Biotechnol.
8:432020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
da Silva Meirelles L, Chagastelles PC and
Nardi NB: Mesenchymal stem cells reside in virtually all post-natal
organs and tissues. J Cell Sci. 119((Pt 11)): 2204–2213. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Secunda R, Vennila R, Mohanashankar AM,
Rajasundari M, Jeswanth S and Surendran R: Isolation, expansion and
characterisation of mesenchymal stem cells from human bone marrow,
adipose tissue, umbilical cord blood and matrix: A comparative
study. Cytotechnology. 67:793–807. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dominici M, Le Blanc K, Mueller I,
Slaper-Cortenbach I, Marini F, Krause D, Deans R, Keating A,
Prockop D and Horwitz E: Minimal criteria for defining multipotent
mesenchymal stromal cells. The international society for cellular
therapy position statement. Cytotherapy. 8:315–317. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Viswanathan S, Shi Y, Galipeau J, Krampera
M, Leblanc K, Martin I, Nolta J, Phinney DG and Sensebe L:
Mesenchymal stem versus stromal cells: International society for
cell & gene therapy (ISCT®) mesenchymal stromal cell
committee position statement on nomenclature. Cytotherapy.
21:1019–1024. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Păunescu V, Deak E, Herman D, Siska IR,
Tănasie G, Bunu C, Anghel S, Tatu CA, Oprea TI, Henschler R, et al:
In vitro differentiation of human mesenchymal stem cells to
epithelial lineage. J Cell Mol Med. 11:502–508. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Quevedo HC, Hatzistergos KE, Oskouei BN,
Feigenbaum GS, Rodriguez JE, Valdes D, Pattany PM, Zambrano JP, Hu
Q, McNiece I, et al: Allogeneic mesenchymal stem cells restore
cardiac function in chronic ischemic cardiomyopathy via trilineage
differentiating capacity. Proc Natl Acad Sci USA. 106:14022–14027.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gervois P, Struys T, Hilkens P, Bronckaers
A, Ratajczak J, Politis C, Brône B, Lambrichts I and Martens W:
Neurogenic maturation of human dental pulp stem cells following
neurosphere generation induces morphological and
electrophysiological characteristics of functional neurons. Stem
Cells Dev. 24:296–311. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mishra PJ, Mishra PJ, Humeniuk R, Medina
DJ, Alexe G, Mesirov JP, Ganesan S, Glod JW and Banerjee D:
Carcinoma-associated fibroblast-like differentiation of human
mesenchymal stem cells. Cancer Res. 68:4331–4339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jotzu C, Alt E, Welte G, Li J, Hennessy
BT, Devarajan E, Krishnappa S, Pinilla S, Droll L and Song YH:
Adipose tissue-derived stem cells differentiate into
carcinoma-associated fibroblast-like cells under the influence of
tumor-derived factors. Anal Cell Pathol (Amst). 33:61–79. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Miyazaki Y, Oda T, Inagaki Y, Kushige H,
Saito Y, Mori N, Takayama Y, Kumagai Y, Mitsuyama T and Kida YS:
Adipose-derived mesenchymal stem cells differentiate into
heterogeneous cancer-associated fibroblasts in a stroma-rich
xenograft model. Sci Rep. 11:46902021. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lee MW, Ryu S, Kim DS, Lee JW, Sung KW,
Koo HH and Yoo KH: Mesenchymal stem cells in suppression or
progression of hematologic malignancy: Current status and
challenges. Leukemia. 33:597–611. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liang W and Chen X, Zhang S, Fang J, Chen
M, Xu Y and Chen X: Mesenchymal stem cells as a double-edged sword
in tumor growth: Focusing on MSC-derived cytokines. Cell Mol Biol
Lett. 26:32021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bellagamba BC, de Abreu BRR, Grivicich I,
Markarian CF, Chem E, Camassola M, Nardi NB and Dihl RR: Human
mesenchymal stem cells are resistant to cytotoxic and genotoxic
effects of cisplatin in vitro. Genet Mol Biol. 39:129–134. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Honczarenko M, Le Y, Swierkowski M, Ghiran
I, Glodek AM and Silberstein LE: Human bone marrow stromal cells
express a distinct set of biologically functional chemokine
receptors. Stem Cells. 24:1030–1041. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lee RH, Seo MJ, Pulin AA, Gregory CA,
Ylostalo J and Prockop DJ: The CD34-like protein PODXL and
alpha6-integrin (CD49f) identify early progenitor MSCs with
increased clonogenicity and migration to infarcted heart in mice.
Blood. 113:816–826. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Williams SA, Maecker HL, French DM, Liu J,
Gregg A, Silverstein LB, Cao TC, Carano RAD and Dixit VM: USP1
deubiquitinates ID proteins to preserve a mesenchymal stem cell
program in osteosarcoma. Cell. 146:918–930. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhuo L, Gong J, Yang R, Sheng Y, Zhou L,
Kong X and Cao K: Inhibition of proliferation and differentiation
and promotion of apoptosis by cyclin L2 in mouse embryonic
carcinoma P19 cells. Biochem Biophys Res Commun. 390:451–457. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Puchert M and Engele J: The peculiarities
of the SDF-1/CXCL12 system: In some cells, CXCR4 and CXCR7 sing
solos, in others, they sing duets. Cell Tissue Res. 355:239–253.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Janssens R, Struyf S and Proost P: The
unique structural and functional features of CXCL12. Cell Mol
Immunol. 15:299–311. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
De La Luz Sierra M, Yang F, Narazaki M,
Salvucci O, Davis D, Yarchoan R, Zhang HH, Fales H and Tosato G:
Differential processing of stromal-derived factor-1alpha and
stromal-derived factor-1beta explains functional diversity. Blood.
103:2452–2459. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gomes AC, Hara T, Lim VY,
Herndler-Brandstetter D, Nevius E, Sugiyama T, Tani-ichi S,
Schlenner S, Richie E, Rodewald HR, et al: Hematopoietic stem cell
niches produce lineage-instructive signals to control multipotent
progenitor differentiation. Immunity. 45:1219–1231. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Reid JC, Tanasijevic B, Golubeva D, Boyd
AL, Porras DP, Collins TJ and Bhatia M: CXCL12/CXCR4 signaling
enhances human PSC-derived hematopoietic progenitor function and
overcomes early in vivo transplantation failure. Stem Cell Reports.
10:1625–1641. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Binder ZA, Siu IM, Eberhart CG, Rhys CA,
Bai RY, Staedtke V, Zhang H, Smoll NR, Piantadosi S, Piccirillo SG,
et al: Podocalyxin-like protein is expressed in glioblastoma
multiforme stem-like cells and is associated with poor outcome.
PLoS One. 8:e759452013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Chang J, Liu F, Lee M, Wu B, Ting K, Zara
JN, Soo C, Hezaimi KA, Zou W, Chen X, et al: NF-κB inhibits
osteogenic differentiation of mesenchymal stem cells by promoting
β-catenin degradation. Proc Natl Acad Sci. 110:9469–9474. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kolosova IA, Angelini D, Fan C, Skinner J,
Cheadle C and Johns RA: Resistin-like molecule α stimulates
proliferation of mesenchymal stem cells while maintaining their
multipotency. Stem Cells Dev. 22:239–247. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Chosa N and Ishisaki A: Two novel
mechanisms for maintenance of stemness in mesenchymal stem cells:
SCRG1/BST1 axis and cell-cell adhesion through N-cadherin. Jpn Dent
Sci Rev. 54:37–44. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Sugiyama T, Kohara H, Noda M and Nagasawa
T: Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4
chemokine signaling in bone marrow stromal cell niches. Immunity.
25:977–988. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gharibi B, Ghuman MS and Hughes FJ: Akt-
and Erk-mediated regulation of proliferation and differentiation
during PDGFRβ-induced MSC self-renewal. J Cell Mol Med.
16:2789–2801. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Takebe N, Miele L, Harris PJ, Jeong W,
Bando H, Kahn M, Yang SX and Ivy SP: Targeting Notch, hedgehog, and
wnt pathways in cancer stem cells: Clinical update. Nat Rev Clin
Oncol. 12:445–464. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Batsali AK, Pontikoglou C, Koutroulakis D,
Pavlaki KI, Damianaki A, Mavroudi I, Alpantaki K, Kouvidi E,
Kontakis G and Papadaki HA: Differential expression of cell cycle
and WNT pathway-related genes accounts for differences in the
growth and differentiation potential of Wharton's jelly and bone
marrow-derived mesenchymal stem cells. Stem Cell Res Ther.
8:1022017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Pelullo M, Zema S, Nardozza F, Checquolo
S, Screpanti I and Bellavia D: Wnt, Notch, and TGF-β pathways
impinge on Hedgehog signaling complexity: An open window on cancer.
Front Genet. 10:7112019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Wagner W, Horn P, Castoldi M, Diehlmann A,
Bork S, Saffrich R, Benes V, Blake J, Pfister S, Eckstein V and Ho
AD: Replicative senescence of mesenchymal stem cells: A continuous
and organized process. PLoS One. 3:e22132008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Halfon S, Abramov N, Grinblat B and Ginis
I: Markers distinguishing mesenchymal stem cells from fibroblasts
are downregulated with passaging. Stem Cells Dev. 20:53–66. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Pérez-Campo FM and Riancho JA: Epigenetic
mechanisms regulating mesenchymal stem cell differentiation. Curr
Genomics. 16:368–383. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Teven CM, Liu X, Hu N, Tang N, Kim SH,
Huang E, Yang K, Li M, Gao JL, Liu H, et al: Epigenetic regulation
of mesenchymal stem cells: A focus on osteogenic and adipogenic
differentiation. Stem Cells Int. 2011:2013712011. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Srinageshwar B, Maiti P, Dunbar GL and
Rossignol J: Role of epigenetics in stem cell proliferation and
differentiation: Implications for treating neurodegenerative
diseases. Int J Mol Sci. 17:1992016. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Escacena N, Quesada-Hernández E,
Capilla-Gonzalez V, Soria B and Hmadcha A: Bottlenecks in the
efficient use of advanced therapy medicinal products based on
mesenchymal stromal cells. Stem Cells Int. 2015:8957142015.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Zhang D and Kilian KA: The effect of
mesenchymal stem cell shape on the maintenance of multipotency.
Biomaterials. 34:3962–3969. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Rathbone SR, Glossop JR, Gough JE and
Cartmell SH: Cyclic tensile strain upon human mesenchymal stem
cells in 2D and 3D culture differentially influences CCNL2, WDR61
and BAHCC1 gene expression levels. J Mech Behav Biomed Mater.
11:82–91. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Cao C, Li L, Li H, He X, Wu G and Yu X:
Cyclic biaxial tensile strain promotes bone marrow-derived
mesenchymal stem cells to differentiate into cardiomyocyte-like
cells by miRNA-27a. Int J Biochem Cell Biol. 99:125–132. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhang L, Wang Y, Zhou N, Feng Y and Yang
X: Cyclic tensile stress promotes osteogenic differentiation of
adipose stem cells via ERK and p38 pathways. Stem Cell Res.
37:1014332019. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Lazarus HM, Haynesworth SE, Gerson SL,
Rosenthal NS and Caplan AI: Ex vivo expansion and subsequent
infusion of human bone marrow-derived stromal progenitor cells
(mesenchymal progenitor cells): Implications for therapeutic use.
Bone Marrow Transplant. 16:557–564. 1995.PubMed/NCBI
|
|
69
|
Galipeau J and Sensébé L: Mesenchymal
stromal cells: Clinical challenges and therapeutic opportunities.
Cell Stem Cell. 22:824–833. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Lukomska B, Stanaszek L, Zuba-Surma E,
Legosz P, Sarzynska S and Drela K: Challenges and controversies in
human mesenchymal stem cell therapy. Stem Cells Int.
2019:96285362019. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Gálvez P, Clares B, Bermejo M, Hmadcha A
and Soria B: Standard requirement of a microbiological quality
control program for the manufacture of human mesenchymal stem cells
for clinical use. Stem Cells Dev. 23:1074–1083. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Galvez-Martin P, Sabata R, Verges J,
Zugaza JL, Ruiz A and Clares B: Mesenchymal stem cells as
therapeutics agents: Quality and environmental regulatory aspects.
Stem Cells Int. 2016:97834082016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Yang YHK: Aging of mesenchymal stem cells:
Implication in regenerative medicine. Regen Ther. 9:120–122. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Fafián-Labora JA, Morente-López M and
Arufe MC: Effect of aging on behaviour of mesenchymal stem cells.
World J Stem Cells. 11:337–346. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Huang XP, Sun Z, Miyagi Y, McDonald KH,
Zhang L, Weisel RD and Li RK: Differentiation of allogeneic
mesenchymal stem cells induces immunogenicity and limits their
long-term benefits for myocardial repair. Circulation.
122:2419–2429. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Cho PS, Messina DJ, Hirsh EL, Chi N,
Goldman SN, Lo DP, Harris IR, Popma SH, Sachs DH and Huang CA:
Immunogenicity of umbilical cord tissue-derived cells. Blood.
111:430–438. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Faiella W and Atoui R: Immunotolerant
properties of mesenchymal stem cells: Updated review. Stem Cells
Int. 2016:18595672016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Dhingra S, Li P, Huang XP, Guo J, Wu J,
Mihic A, Li SH, Zang WF, Shen D, Weisel RD, et al: Preserving
prostaglandin E2 level prevents rejection of implanted allogeneic
mesenchymal stem cells and restores postinfarction ventricular
function. Circulation. 128((11 Suppl 1)): S69–S78. 2013.PubMed/NCBI
|
|
79
|
Gu Z, Tan W, Ji J, Feng G, Meng Y, Da Z,
Guo G, Xia Y, Zhu X, Shi G and Cheng C: Rapamycin reverses the
senescent phenotype and improves immunoregulation of mesenchymal
stem cells from MRL/lpr mice and systemic lupus erythematosus
patients through inhibition of the mTOR signaling pathway. Aging
(Albany NY). 8:1102–1114. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Ankrum JA, Ong JF and Karp JM: Mesenchymal
stem cells: Immune evasive, not immune privileged. Nat Biotechnol.
32:252–260. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zazzeroni L, Lanzoni G, Pasquinelli G and
Ricordi C: Considerations on the harvesting site and donor
derivation for mesenchymal stem cells-based strategies for
diabetes. CellR4 Repair Replace Regen Reprogram.
5:e24352017.PubMed/NCBI
|
|
82
|
Gao L, Bird AK, Meednu N, Dauenhauer K,
Liesveld J, Anolik J and Looney RJ: Bone marrow-derived mesenchymal
stem cells from patients with systemic lupus erythematosus have a
senescence-associated secretory phenotype mediated by a
mitochondrial antiviral signaling protein-interferon-β feedback
loop. Arthritis Rheumatol. 69:1623–1635. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Ji J, Wu Y, Meng Y, Zhang L, Feng G, Xia
Y, Xue W, Zhao S, Gu Z and Shao X: JAK-STAT signaling mediates the
senescence of bone marrow-mesenchymal stem cells from systemic
lupus erythematosus patients. Acta Biochim Biophys Sin (Shanghai).
49:208–215. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhu Y and Feng X: Genetic contribution to
mesenchymal stem cell dysfunction in systemic lupus erythematosus.
Stem Cell Res Ther. 9:1492018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Chen DC, Lin SZ, Fan JR, Lin CH, Lee W,
Lin CC, Liu YJ, Tsai CH, Chen JC, Cho DY, et al: Intracerebral
implantation of autologous peripheral blood stem cells in stroke
patients: A randomized phase II study. Cell Transplant.
23:1599–1612. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Taguchi A, Sakai C, Soma T, Kasahara Y,
Stern DM, Kajimoto K, Ihara M, Daimon T, Yamahara K, Doi K, et al:
Intravenous autologous bone marrow mononuclear cell transplantation
for stroke: Phase1/2a clinical trial in a homogeneous group of
stroke patients. Stem Cells Dev. 24:2207–2218. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Hess DC, Wechsler LR, Clark WM, Savitz SI,
Ford GA, Chiu D, Yavagal DR, Uchino K, Liebeskind DS, Auchus AP, et
al: Safety and efficacy of multipotent adult progenitor cells in
acute ischaemic stroke (MASTERS): A randomised, double-blind,
placebo-controlled, phase 2 trial. Lancet Neurol. 16:360–368. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Bhatia V, Gupta V, Khurana D, Sharma RR
and Khandelwal N: Randomized assessment of the safety and efficacy
of intra-arterial infusion of autologous wtem cells in wubacute
ischemic stroke. AJNR Am J Neuroradiol. 39:899–904. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Gautam J, Alaref A, Hassan A, Kandel RS,
Mishra R and Jahan N: Safety and efficacy of stem cell therapy in
patients with ischemic stroke. Cureus. 12:e99172020.PubMed/NCBI
|
|
90
|
Trachtenberg B, Velazquez DL, Williams AR,
McNiece I, Fishman J, Nguyen K, Rouy D, Altman P, Schwarz R,
Mendizabal A, et al: Rationale and design of the transendocardial
injection of autologous human cells (bone marrow or mesenchymal) in
chronic ischemic left ventricular dysfunction and heart failure
secondary to myocardial infarction (TAC-HFT) trial: A randomized,
double-blind, placebo-controlled study of safety and efficacy. Am
Heart J. 161:487–493. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Mushtaq M, DiFede DL, Golpanian S, Khan A,
Gomes SA, Mendizabal A, Heldman AW and Hare JM: Rationale and
design of the percutaneous stem cell injection delivery effects on
neomyogenesis in dilated cardiomyopathy (The POSEIDON-DCM Study). J
Cardiovasc Transl Res. 7:769–780. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Hare JM, DiFede DL, Rieger AC, Florea V,
Landin AM, El-Khorazaty J, Khan A, Mushtaq M, Lowery MH, Byrnes JJ,
et al: Randomized comparison of allogeneic versus autologous
mesenchymal stem cells for nonischemic dilated cardiomyopathy:
POSEIDON-DCM Trial. J Am Coll Cardiol. 69:526–537. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kidd S, Spaeth E, Dembinski JL, Dietrich
M, Watson K, Klopp A, Battula L, Weil M, Andreeff M and Marini FC:
Direct evidence of mesenchymal stem cell tropism for tumor and
wounding microenvironments using in vivo bioluminescence imaging.
Stem Cells. 27:2614–2623. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Sun X, Cheng G, Hao M, Zheng J, Zhou X,
Zhang J, Taichman RS, Pienta KJ and Wang J: CXCL12/CXCR4/CXCR7
chemokine axis and cancer progression. Cancer Metastasis Rev.
29:709–722. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Spaeth E, Klopp A, Dembinski J, Andreeff M
and Marini F: Inflammation and tumor microenvironments: Defining
the migratory itinerary of mesenchymal stem cells. Gene Ther.
15:730–738. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Kolaczkowska E and Kubes P: Neutrophil
recruitment and function in health and inflammation. Nat Rev
Immunol. 13:159–175. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Dimarino AM, Caplan AI and Bonfield TL:
Mesenchymal stem cells in tissue repair. Front Immunol. 4:2012013.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Ayala-Cuellar AP, Kang JH, Jeung EB and
Choi KC: Roles of mesenchymal stem cells in tissue regeneration and
immunomodulation. Biomol Ther (Seoul). 27:25–33. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Julier Z, Park AJ, Briquez PS and Martino
MM: Promoting tissue regeneration by modulating the immune system.
Acta Biomater. 53:13–28. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Prockop DJ and Oh JY: Mesenchymal
stem/stromal cells (MSCs): Role as guardians of inflammation. Mol
Ther. 20:14–20. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Song J, Kang HJ, Ju HM, Park A, Park H,
Hong JS, Kim CJ, Shim JY, Yu J and Choi J: Umbilical cord-derived
mesenchymal stem cell extracts ameliorate atopic dermatitis in mice
by reducing the T cell responses. Sci Rep. 9:66232019. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Ren G, Zhao X, Zhang L, Zhang J,
L'Huillier A, Ling W, Roberts AI, Le AD, Shi S, Shao C and Shi Y:
Inflammatory cytokine-induced intercellular adhesion molecule-1 and
vascular cell adhesion molecule-1 in mesenchymal stem cells are
critical for immunosuppression. J Immunol. 184:2321–2328. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Li Y, Zhang D, Xu L, Dong L, Zheng J, Lin
Y, Huang J, Zhang Y, Tao Y, Zang X, et al: Cell-cell contact with
proinflammatory macrophages enhances the immunotherapeutic effect
of mesenchymal stem cells in two abortion models. Cell Mol Immunol.
16:908–920. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Nitzsche F, Müller C, Lukomska B,
Jolkkonen J, Deten A and Boltze J: Concise review: MSC adhesion
cascade-insights into homing and transendothelial migration. Stem
Cells. 35:1446–1460. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Caplan H, Olson SD, Kumar A, George M,
Prabhakara KS, Wenzel P, Bedi S, Toledano-Furman NE, Triolo F,
Kamhieh-Milz J, et al: Mesenchymal stromal cell therapeutic
delivery: Translational challenges to clinical application. Front
Immunol. 10:16452019. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Ullah M, Liu DD and Thakor AS: Mesenchymal
stromal cell homing: Mechanisms and strategies for improvement.
iScience. 15:421–438. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Fiore EJ, Domínguez LM, Bayo J, García MG
and Mazzolini GD: Taking advantage of the potential of mesenchymal
stromal cells in liver regeneration: Cells and extracellular
vesicles as therapeutic strategies. World J Gastroenterol.
24:2427–2440. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Li H, Rong P, Ma X, Nie W, Chen C, Yang C,
Zhang J, Dong Q and Wang W: Paracrine effect of mesenchymal stem
cell as a novel therapeutic strategy for diabetic nephropathy. Life
Sci. 215:113–118. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Zheng G, Huang R, Qiu G, Ge M, Wang J, Shu
Q and Xu J: Mesenchymal stromal cell-derived extracellular
vesicles: Regenerative and immunomodulatory effects and potential
applications in sepsis. Cell Tissue Res. 374:1–15. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Weiss ARR and Dahlke MH: Immunomodulation
by mesenchymal stem cells (MSCs): Mechanisms of action of living,
apoptotic, and dead MSCs. Front Immunol. 10:11912019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Coussens LM and Werb Z: Inflammation and
cancer. Nature. 420:860–867. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Wobus M, List C, Dittrich T, Dhawan A,
Duryagina R, Arabanian LS, Kast K, Wimberger P, Stiehler M,
Hofbauer LC, et al: Breast carcinoma cells modulate the
chemoattractive activity of human bone marrow-derived mesenchymal
stromal cells by interfering with CXCL12. Int J Cancer. 136:44–54.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Kalimuthu S, Oh JM, Gangadaran P, Zhu L,
Lee HW, Rajendran RL, Baek SH, Jeon YH, Jeong SY, Lee SW, et al: In
vivo tracking of chemokine receptor CXCR4-engineered mesenchymal
stem cell migration by optical molecular imaging. Stem Cells Int.
2017:80856372017. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Ratajczak MZ, Bujko K, Mack A, Kucia M and
Ratajczak J: Cancer from the perspective of stem cells and
misappropriated tissue regeneration mechanisms. Leukemia.
32:2519–2526. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Suzuki K, Sun R, Origuchi M, Kanehira M,
Takahata T, Itoh J, Umezawa A, Kijima H, Fukuda S and Saijo Y:
Mesenchymal stromal cells promote tumor growth through the
enhancement of neovascularization. Mol Med. 17:579–587. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Lu L, Chen G, Yang J, Ma Z, Yang Y, Hu Y,
Lu Y, Cao Z, Wang Y and Wang X: Bone marrow mesenchymal stem cells
suppress growth and promote the apoptosis of glioma U251 cells
through downregulation of the PI3K/AKT signaling pathway. Biomed
Pharmacother. 112:1086252019. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Bajetto A, Pattarozzi A, Corsaro A,
Barbieri F, Daga A, Bosio A, Gatti M, Pisaturo V, Sirito R and
Florio T: Different effects of human umbilical cord mesenchymal
stem cells on glioblastoma stem cells by direct cell interaction or
via released soluble factors. Front Cell Neurosci. 11:3122017.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Zheng H, Zou W, Shen J, Xu L, Wang S, Fu
YX and Fan W: Opposite effects of coinjection and distant injection
of mesenchymal stem cells on breast tumor cell growth. Stem Cells
Transl Med. 5:1216–1228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Akimoto K, Kimura K, Nagano M, Takano S,
To'a Salazar G, Yamashita T and Ohneda O: Umbilical cord
blood-derived mesenchymal stem cells inhibit, but adipose
tissue-derived mesenchymal stem cells promote, glioblastoma
multiforme proliferation. Stem Cells Dev. 22:1370–1386. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Barcellos-de-Souza P, Comito G,
Pons-Segura C, Taddei ML, Gori V, Becherucci V, Bambi F, Margheri
F, Laurenzana A, Del Rosso M and Chiarugi P: Mesenchymal stem cells
are recruited and activated into carcinoma-associated fibroblasts
by prostate cancer microenvironment-derived TGF-β1. Stem Cells.
34:2536–2547. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Hill BS, Pelagalli A, Passaro N and
Zannetti A: Tumor-educated mesenchymal stem cells promote
pro-metastatic phenotype. Oncotarget. 8:73296–73311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Tan HX, Xiao ZG, Huang T, Fang ZX, Liu Y
and Huang ZC: CXCR4/TGF-β1 mediated self-differentiation of human
mesenchymal stem cells to carcinoma-associated fibroblasts and
promoted colorectal carcinoma development. Cancer Biol Ther.
21:248–257. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Walter M, Liang S, Ghosh S, Hornsby PJ and
Li R: Interleukin 6 secreted from adipose stromal cells promotes
migration and invasion of breast cancer cells. Oncogene.
28:2745–2755. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Tsai KS, Yang SH, Lei YP, Tsai CC, Chen
HW, Hsu CY, Chen LL, Wang HW, Miller SA, Chiou SH, et al:
Mesenchymal stem cells promote formation of colorectal tumors in
mice. Gastroenterology. 141:1046–1056. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Zhang T, Lee YW, Rui YF, Cheng TY, Jiang
XH and Li G: Bone marrow-derived mesenchymal stem cells promote
growth and angiogenesis of breast and prostate tumors. Stem Cell
Res Ther. 4:702013. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
El-Haibi CP, Bell GW, Zhang J, Collmann
AY, Wood D, Scherber CM, Csizmadia E, Mariani O, Zhu C, Campagne A,
et al: Critical role for lysyl oxidase in mesenchymal stem
cell-driven breast cancer malignancy. Proc Natl Acad Sci USA.
109:17460–17465. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Patel SA, Meyer JR, Greco SJ, Corcoran KE,
Bryan M and Rameshwar P: Mesenchymal stem cells protect breast
cancer cells through regulatory T cells: Role of mesenchymal stem
cell-derived TGF-beta. J Immunol. 184:5885–5894. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Gazdic M, Markovic BS, Jovicic N,
Misirkic-Marjanovic M, Djonov V, Jakovljevic V, Arsenijevic N,
Lukic ML and Volarevic V: Mesenchymal stem cells promote metastasis
of lung cancer cells by downregulating systemic antitumor immune
response. Stem Cells Int. 2017:62947172017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Ramasamy R, Lam EWF, Soeiro I, Tisato V,
Bonnet D and Dazzi F: Mesenchymal stem cells inhibit proliferation
and apoptosis of tumor cells: Impact on in vivo tumor growth.
Leukemia. 21:304–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
He N, Kong Y, Lei X, Liu Y, Wang J, Xu C,
Wang Y, Du L, Ji K, Wang Q, et al: MSCs inhibit tumor progression
and enhance radiosensitivity of breast cancer cells by
down-regulating Stat3 signaling pathway. Cell Death Dis.
9:10262018. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Yulyana Y, Ho IAW, Sia KC, Newman JP, Toh
XY, Endaya BB, Chan JKY, Gnecchi M, Huynh H, Chung AY, et al:
Paracrine factors of human fetal MSCs inhibit liver cancer growth
through reduced activation of IGF-1R/PI3K/Akt signaling. Mol Ther.
23:746–756. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Chen HL, Li JJ, Jiang F, Shi WJ and Chang
GY: MicroRNA-4461 derived from bone marrow mesenchymal stem cell
exosomes inhibits tumorigenesis by downregulating COPB2 expression
in colorectal cancer. Biosci Biotechnol Biochem. 84:338–346. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Ho IAW, Toh HC, Ng WH, Teo YL, Guo CM, Hui
KM and Lam PYP: Human bone marrow-derived mesenchymal stem cells
suppress human glioma growth through inhibition of angiogenesis.
Stem Cells. 31:146–155. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Khalil C, Moussa M, Azar A, Tawk J,
Habbouche J, Salameh R, Ibrahim A and Alaaeddine N:
Anti-proliferative effects of mesenchymal stem cells (MSCs) derived
from multiple sources on ovarian cancer cell lines: An in-vitro
experimental study. J Ovarian Res. 12:702019. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
DiDonato JA, Mercurio F and Karin M: NF-κB
and the link between inflammation and cancer. Immunol Rev.
246:379–400. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Li M, Kouzmina E, McCusker M, Rodin D,
Boutros PC, Paige CJ and Rodin G: Pro- and anti-inflammatory
cytokine associations with major depression in cancer patients.
Psychooncology. 26:2149–2156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Ahechu P, Zozaya G, Martí P,
Hernández-Lizoáin JL, Baixauli J, Unamuno X, Frühbeck G and Catalán
V: NLRP3 inflammasome: A possible link between obesity-associated
low-grade chronic inflammation and colorectal cancer development.
Front Immunol. 9:29182018. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Mocellin S, Panelli MC, Wang E, Nagorsen D
and Marincola FM: The dual role of IL-10. Trends Immunol. 24:36–43.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Tanikawa T, Wilke CM, Kryczek I, Chen GY,
Kao J, Núñez G and Zou W: Interleukin-10 ablation promotes tumor
development, growth, and metastasis. Cancer Res. 72:420–429. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Nappo G, Handle F, Santer FR, McNeill RV,
Seed RI, Collins AT, Morrone G, Culig Z, Maitland NJ and Erb HHH:
The immunosuppressive cytokine interleukin-4 increases the
clonogenic potential of prostate stem-like cells by activation of
STAT6 signalling. Oncogenesis. 6:e3422017. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Setrerrahmane S and Xu H: Tumor-related
iCnterleukins: Old validated targets for new anti-cancer drug
development. Mol Cancer. 16:1532017. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Saeedi P, Halabian R and Fooladi AA: A
revealing review of mesenchymal stem cells therapy, clinical
perspectives and modification strategies. Stem Cell Investig.
6:342019. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Bortolotti F, Ukovich L, Razban V,
Martinelli V, Ruozi G, Pelos B, Dore F, Giacca M and Zacchigna S:
In vivo therapeutic potential of mesenchymal stromal cells depends
on the source and the isolation procedure. Stem Cell Rep.
4:332–339. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Fathi E, Sanaat Z and Farahzadi R:
Mesenchymal stem cells in acute myeloid leukemia: A focus on
mechanisms involved and therapeutic concepts. Blood Res.
54:165–174. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Chen Y, He Y, Wang X, Lu F and Gao J:
Adipose-derived mesenchymal stem cells exhibit tumor tropism and
promote tumorsphere formation of breast cancer cells. Oncol Rep.
41:2126–2136. 2019.PubMed/NCBI
|
|
146
|
Dührsen L, Hartfuß S, Hirsch D, Geiger S,
Maire CL, Sedlacik J, Guenther C, Westphal M, Lamszus K, Hermann FG
and Schmidt NO: Preclinical analysis of human mesenchymal stem
cells: Tumor tropism and therapeutic efficiency of local HSV-TK
suicide gene therapy in glioblastoma. Oncotarget. 10:6049–6061.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Dissanayake S, Denny WA, Gamage S and
Sarojini V: Recent developments in anticancer drug delivery using
cell penetrating and tumor targeting peptides. J Control Release.
250:62–76. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Lagoa R, Silva J, Rodrigues JR and
Bishayee A: Advances in phytochemical delivery systems for improved
anticancer activity. Biotechnol Adv. 38:1073822020. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Studeny M, Marini FC, Champlin RE,
Zompetta C, Fidler IJ and Andreeff M: Bone marrow-derived
mesenchymal stem cells as vehicles for interferon-beta delivery
into tumors. Cancer Res. 62:3603–3608. 2002.PubMed/NCBI
|
|
150
|
Ahn JO, Lee HW, Seo KW, Kang SK, Ra JC and
Youn HY: Anti-tumor effect of adipose tissue derived-mesenchymal
stem cells expressing interferon-β and treatment with cisplatin in
a xenograft mouse model for canine melanoma. PLoS One.
8:e748972013. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Shen CJ, Chan TF, Chen CC, Hsu YC, Long CY
and Lai CS: Human umbilical cord matrix-derived stem cells
expressing interferon-β gene inhibit breast cancer cells via
apoptosis. Oncotarget. 7:34172–34179. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Yuan ZQ, Kolluri KK, Sage EK, Gowers KHC
and Janes SM: Mesenchymal stromal cell delivery of full-length
tumor necrosis factor-related apoptosis-inducing ligand is superior
to soluble type for cancer therapy. Cytotherapy. 17:885–896. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Marini I, Siegemund M, Hutt M, Kontermann
RE and Pfizenmaier K: Antitumor activity of a mesenchymal stem cell
line stably secreting a tumor-targeted TNF-related
apoptosis-inducing ligand fusion protein. Front Immunol. 8:5362017.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Guiho R, Biteau K, Grisendi G, Chatelais
M, Brion R, Taurelle J, Renault S, Heymann D, Dominici M and Redini
F: In vitro and in vivo discrepancy in inducing apoptosis by
mesenchymal stromal cells delivering membrane-bound tumor necrosis
factor-related apoptosis inducing ligand in osteosarcoma
pre-clinical models. Cytotherapy. 20:1037–1045. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Shamili FH, Bayegi HR, Salmasi Z, Sadri K,
Mahmoudi M, Kalantari M, Ramezani M and Abnous K: Exosomes derived
from TRAIL-engineered mesenchymal stem cells with effective
anti-tumor activity in a mouse melanoma model. Int J Pharm.
549:218–229. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Yang X, Du J, Xu X, Xu C and Song W:
IFN-γ-secreting-mesenchymal stem cells exert an antitumor effect in
vivo via the TRAIL pathway. J Immunol Res. 2014:e3180982014.
View Article : Google Scholar
|
|
157
|
You Q, Yao Y, Zhang Y, Fu S, Du M and
Zhang G: Effect of targeted ovarian cancer therapy using amniotic
fluid mesenchymal stem cells transfected with enhanced green
fluorescent protein-human interleukin-2 in vivo. Mol Med
Rep. 12:4859–4866. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Zhao W, Cheng J, Shi P and Huang J: Human
umbilical cord mesenchymal stem cells with adenovirus-mediated
interleukin 12 gene transduction inhibits the growth of ovarian
carcinoma cells both in vitro and in vivo. Nan Fang Yi Ke Da Xue
Xue Bao. 31:903–907. 2011.(In Chinese). PubMed/NCBI
|
|
159
|
Zhang X, Zhang L, Xu W, Qian H, Ye S, Zhu
W, Cao H, Yan Y, Li W, Wang M, et al: Experimental therapy for lung
cancer: Umbilical cord-derived mesenchymal stem cell-mediated
interleukin-24 delivery. Curr Cancer Drug Targets. 13:92–102. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Nowakowski A, Walczak P, Lukomska B and
Janowski M: Genetic engineering of mesenchymal stem cells to induce
their migration and survival. Stem Cells Int. 2016:e49560632016.
View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Wei W, Huang Y, Li D, Gou HF and Wang W:
Improved therapeutic potential of MSCs by genetic modification.
Gene Ther. 25:538–547. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Ocansey DKW, Pei B, Yan Y, Qian H, Zhang
X, Xu W and Mao F: Improved therapeutics of modified mesenchymal
stem cells: An update. J Transl Med. 18:422020. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Phillips MI and Tang YL: Genetic
modification of stem cells for transplantation. Adv Drug Deliv Rev.
60:160–172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Bobis-Wozowicz S, Miekus K, Wybieralska E,
Jarocha D, Zawisz A, Madeja Z and Majka M: Genetically modified
adipose tissue-derived mesenchymal stem cells overexpressing CXCR4
display increased motility, invasiveness, and homing to bone marrow
of NOD/SCID mice. Exp Hematol. 39:686–696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Pelagalli A, Nardelli A, Lucarelli E,
Zannetti A and Brunetti A: Autocrine signals increase ovine
mesenchymal stem cells migration through Aquaporin-1 and CXCR4
overexpression. J Cell Physiol. 233:6241–6249. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Song SW, Chang W, Song BW, Song H, Lim S,
Kim HJ, Cha MJ, Choi E, Im SH, Chang BC, et al: Integrin-linked
kinase is required in hypoxic mesenchymal stem cells for
strengthening cell adhesion to ischemic myocardium. Stem Cells.
27:1358–1365. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Fan YX, Gu CH, Zhang YL, Zhong BS, Wang
LZ, Zhou ZR, Wang ZY, Jia RX and Wang F: Oct4 and Sox2
overexpression improves the proliferation and differentiation of
bone mesenchymal stem cells in Xiaomeishan porcine. Genet Mol Res.
12:6067–6079. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Han SM, Han SH, Coh YR, Jang G, Ra JC,
Kang SK, Lee HW and Youn HY: Enhanced proliferation and
differentiation of Oct4- and Sox2-overexpressing human adipose
tissue mesenchymal stem cells. Exp Mol Med. 46:e1012014. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Becker AD and Riet IV: Homing and
migration of mesenchymal stromal cells: How to improve the efficacy
of cell therapy? World J Stem Cells. 8:73–87. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
DelaRosa O, Dalemans W and Lombardo E:
Toll-like receptors as modulators of mesenchymal stem cells. Front
Immunol. 3:1822012. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Najar M, Krayem M, Meuleman N, Bron D and
Lagneaux L: Mesenchymal stromal cells and toll-like receptor
priming: A critical review. Immune Netw. 17:89–102. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Mekhemar MK, Dörfer CE and El-Sayed KMF:
Toll-like receptors: The key of immunotherapy in MSCs.
Immunoregulatory aspects of immunotherapy. IntechOpen; pp.
1732018
|
|
173
|
Waterman RS, Tomchuck SL, Henkle SL and
Betancourt AM: A new mesenchymal stem cell (MSC) paradigm:
Polarization into a pro-inflammatory MSC1 or an immunosuppressive
MSC2 phenotype. PLoS One. 5:e100882010. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Kim J and Hematti P: Mesenchymal stem
cell-educated macrophages: A novel type of alternatively activated
macrophages. Exp Hematol. 37:1445–1453. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Cassatella MA, Mosna F, Micheletti A, Lisi
V, Tamassia N, Cont C, Calzetti F, Pelletier M, Pizzolo G and
Krampera M: Toll-like receptor-3-activated human mesenchymal
stromal cells significantly prolong the survival and function of
neutrophils. Stem Cells. 29:1001–1011. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Hall SRR, Tsoyi K, Ith B, Padera RF Jr,
Lederer JA, Wang Z, Liu X and Perrella MA: Mesenchymal stromal
cells improve survival during sepsis in the absence of heme
oxygenase-1: The importance of neutrophils. Stem Cells. 31:397–407.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Jiang W and Xu J: Immune modulation by
mesenchymal stem cells. Cell Prolif. 53:e127122019.PubMed/NCBI
|
|
178
|
Kudlik G, Hegyi B, Czibula Á, Monostori É,
Buday L and Uher F: Mesenchymal stem cells promote macrophage
polarization toward M2b-like cells. Exp Cell Res. 348:36–45. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Spaggiari GM, Capobianco A, Abdelrazik H,
Becchetti F, Mingari MC and Moretta L: Mesenchymal stem cells
inhibit natural killer-cell proliferation, cytotoxicity, and
cytokine production: Role of indoleamine 2,3-dioxygenase and
prostaglandin E2. Blood. 111:1327–1333. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Gonzalez H, Hagerling C and Werb Z: Roles
of the immune system in cancer: From tumor initiation to metastatic
progression. Genes Dev. 32:1267–1284. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Vinay DS, Ryan EP, Pawelec G, Talib WH,
Stagg J, Elkord E, Lichtor T, Decker WK, Whelan RL, Kumara HM, et
al: Immune evasion in cancer: Mechanistic basis and therapeutic
strategies. Semin Cancer Biol. 35 (Suppl):S185–S198. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Du W, Seah I, Bougazzoul O, Choi G, Meeth
K, Bosenberg MW, Wakimoto H, Fisher D and Shah K: Stem
cell-released oncolytic herpes simplex virus has therapeutic
efficacy in brain metastatic melanomas. Proc Natl Acad Sci USA.
114:E6157–E6165. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Guo Y, Zhang Z, Xu X, Xu Z, Wang S, Huang
D, Li Y, Mou X, Liu F and Xiang C: Menstrual blood-derived stem
cells as delivery vehicles for oncolytic adenovirus virotherapy for
colorectal cancer. Stem Cells Dev. 28:882–896. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Mahasa KJ, Pillis Ld, Ouifki R, Eladdadi
A, Maini P, Yoon AR and Yun CO: Mesenchymal stem cells used as
carrier cells of oncolytic adenovirus results in enhanced oncolytic
virotherapy. Sci Rep. 10:4252020. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Pessina A, Coccè V, Pascucci L, Bonomi A,
Cavicchini L, Sisto F, Ferrari M, Ciusani E, Crovace A, Falchetti
ML, et al: Mesenchymal stromal cells primed with paclitaxel attract
and kill leukaemia cells, inhibit angiogenesis and improve survival
of leukaemia-bearing mice. Br J Haematol. 160:766–778. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Gilazieva Z, Tazetdinova L, Arkhipova S,
Solovyeva V and Rizvanov A: Effect of cisplatin on ultrastructure
and viability of adipose-derived mesenchymal stem cells.
BioNanoScience. 6:534–539. 2016. View Article : Google Scholar
|
|
187
|
Nicolay NH, Perez RL, Rühle A, Trinh T,
Sisombath S, Weber KJ, Ho AD, Debus J, Saffrich R and Huber PE:
Mesenchymal stem cells maintain their defining stem cell
characteristics after treatment with cisplatin. Sci Rep.
6:200352016. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Pascucci L, Coccè V, Bonomi A, Ami D,
Ceccarelli P, Ciusani E, Viganò L, Locatelli A, Sisto F, Doglia SM,
et al: Paclitaxel is incorporated by mesenchymal stromal cells and
released in exosomes that inhibit in vitro tumor growth: A new
approach for drug delivery. J Control Release. 192:262–270. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Pessina A, Bonomi A, Coccè V, Invernici G,
Navone S, Cavicchini L, Sisto F, Ferrari M, Viganò L, Locatelli A,
et al: Mesenchymal stromal cells primed with paclitaxel provide a
new approach for cancer therapy. PLoS One. 6:e283212011. View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Bonomi A, Coccè V, Cavicchini L, Sisto F,
Dossena M, Balzarini P, Portolani N, Ciusani E, Parati E,
Alessandri G and Pessina A: Adipose tissue-derived stromal cells
primed in vitro with paclitaxel acquire anti-tumor activity. Int J
Immunopathol Pharmacol. 26 (Suppl 1):S33–S41. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Coccè V, Franzè S, Brini AT, Giannì AB,
Pascucci L, Ciusani E, Alessandri G, Farronato G, Cavicchini L,
Sordi V, et al: In vitro anticancer activity of extracellular
vesicles (EVs) secreted by gingival mesenchymal stromal cells
primed with paclitaxel. Pharmaceutics. 11:612019. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Coccè V, Farronato D, Brini AT, Masia C,
Giannì AB, Piovani G, Sisto F, Alessandri G, Angiero F and Pessina
A: Drug loaded gingival mesenchymal stromal cells (GinPa-MSCs)
inhibit in vitro proliferation of oral squamous cell carcinoma. Sci
Rep. 7:93762017. View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Layek B, Sadhukha T, Panyam J and Prabha
S: Nano-engineered mesenchymal stem cells increase therapeutic
efficacy of anticancer drug through true active tumor targeting.
Mol Cancer Ther. 17:1196–1206. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Moku G, Layek B, Trautman L, Putnam S,
Panyam J and Prabha S: Improving payload capacity and anti-tumor
efficacy of mesenchymal stem cells using TAT peptide functionalized
polymeric nanoparticles. Cancers (Basel). 11:4912019. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Altun İ and Sonkaya A: The most common
side effects experienced by patients were receiving first cycle of
chemotherapy. Iran J Public Health. 47:1218–1219. 2018.PubMed/NCBI
|
|
196
|
Kim W, Lee SK, Kwon YW, Chung SG and Kim
S: Pioglitazone-primed mesenchymal stem cells stimulate cell
proliferation, collagen synthesis and matrix gene expression in
tenocytes. Int J Mol Sci. 20:4722019. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Hong Y, Kim YS, Hong SH and Oh YM:
Therapeutic effects of adipose-derived stem cells pretreated with
pioglitazone in an emphysema mouse model. Exp Mol Med. 48:e2662016.
View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Park JS, Kim HK, Kang EY, Cho R and Oh YM:
Potential therapeutic strategy in chronic obstructive pulmonary
disease using pioglitazone-augmented Wharton's jelly-derived
mesenchymal stem cells. Tuberc Respir Dis (Seoul). 82:158–165.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Khoo BY, Nadarajan K, Shim SY, Miswan N,
Zang CB, Possinger K and Elstner E: Pretreatment of BMSCs with TZD
solution decreases the proliferation rate of MCF-7 cells by
reducing FGF4 protein expression. Mol Med Rep. 13:3406–3414. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Tsubaki M, Takeda T, Tomonari Y, Kawashima
K, Itoh T, Imano M, Satou T and Nishida S: Pioglitazone inhibits
cancer cell growth through STAT3 inhibition and enhanced AIF
expression via a PPARγ-independent pathway. J Cell Physiol.
233:3638–3647. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
201
|
Moghareabed R, Hemati S, Akhavan A, Emami
H, Farghadani M, Roayaei M, Tavajoh S and Feizi A: Randomized phase
II clinical trial of pioglitazone plus chemotherapy versus
chemotherapy alone in patients with metastatic breast cancer. J
Glob Oncol. 5:832019. View Article : Google Scholar
|
|
202
|
Esmaeili S, Safaroghli-Azar A,
Pourbagheri-Sigaroodi A, Salari S, Gharehbaghian A, Hamidpour M and
Bashash D: Activation of PPARγ intensified the effects of arsenic
trioxide in acute promyelocytic leukemia through the suppression of
PI3K/Akt pathway: Proposing a novel anticancer effect for
pioglitazone. Int J Biochem Cell Biol. 122:1057392020. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Shinmura D, Togashi I, Miyoshi S,
Nishiyama N, Hida N, Tsuji H, Tsuruta H, Segawa K, Tsukada Y, Ogawa
S and Umezawa A: Pretreatment of human mesenchymal stem cells with
pioglitazone improved efficiency of cardiomyogenic
transdifferentiation and cardiac function. Stem Cells. 29:357–366.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Wang M, Cai J, Huang F, Zhu M, Zhang Q,
Yang T, Zhang X, Qian H and Xu W: Pre-treatment of human umbilical
cord-derived mesenchymal stem cells with interleukin-6 abolishes
their growth-promoting effect on gastric cancer cells. Int J Mol
Med. 35:367–375. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
von Bahr L, Sundberg B, Lönnies L, Sander
B, Karbach H, Hägglund H, Ljungman P, Gustafsson B, Karlsson H,
Blanc KL and Ringdén O: Long-term complications, immunologic
effects, and role of passage for outcome in mesenchymal stromal
cell therapy. Biol Blood Marrow Transplant. 18:557–564. 2012.
View Article : Google Scholar : PubMed/NCBI
|