Introduction
Breast cancer is the cancer with the highest
incidence worldwide and affects an increasing number of Chinese
women, thus representing an important health problem in China
(1). In 2017, >25,000 women were
newly diagnosed with breast cancer in the United States (2). Systemic therapy has improved the
overall survival (OS) and disease-free survival of patients with
breast cancer in a manner dependent on the molecular subtype and
stage.
Recently, large-scale next-generation sequencing
(NGS) has been introduced to characterize multiple cancer types at
the genomic level, including breast cancer. These efforts have led
to a broad description of the genomic alterations involved in
tumorigenesis or tumor progression in breast cancer. Breast cancer
is characterized by a large number of genomic alterations, such as
ERBB2 amplification (3),
PIK3CA mutation (4),
FGFR1 amplification (5),
CCND1 amplification (6),
AKT1 mutation and GATA3 mutation (7,8).
MAPKs are key mediators of evolutionarily conserved
signaling networks that play an essential role in multiple aspects
of cell physiology. The MAPK pathway is frequently activated in
cancerous cells. MAP3K1 or MEKK1 is a 196-kDa
serine-threonine kinase in the MAP3K family with functions in cell
viability, apoptosis and cell motility/migration in multiple normal
and tumor cell types (9). However,
the role of MAP3K1 mutations in patient survival in
different types of cancer remains poorly understood. In particular,
the relationship between MAP3K1 mutations and the
clinicopathological characteristics of patients with breast cancer
remains an unresolved question.
Thus, the aim of the present study was to
extensively describe the relationship between MAP3K1 genomic
alterations and breast cancer. A total of 412 Chinese patients with
primary breast tumors were enrolled. The prevalence of
MAP3K1 mutations and their association with
clinicopathological characteristics were assessed. MAP3K1
mutations were also analyzed in data from The Cancer Genome Atlas
(TCGA) and from the Molecular Taxonomy of Breast Cancer
International Consortium (METABRIC) cohort (10).
Materials and methods
Patients
The study population consisted of three cohorts of
patients. The Guangdong Provincial People's Hospital (GDPH) cohort
included 412 consecutive patients diagnosed with treatment-naïve
breast cancer at Guangdong Provincial People's Hospital from
October 2016 to December 2017. The following inclusion criteria
were used: i) Patients were diagnosed with invasive breast cancer;
ii) complete clinical characterization was available (including
age, pathological type, axillary lymph node status, TMN staging,
pathological grade, the status of estrogen receptor (ER), the
status of progesterone receptor (PR), the status of hormone
receptor, human epidermal growth factor receptor 2 (HER2),
molecular subtype (11,12); and iii) tumor biopsies were obtained
using an Institutional Review Board-approved protocol and analyzed
using NGS.
The present study was also based on an analysis of
data obtained from TCGA-BRCA and from the METABRIC cohort. Data for
the MAP3K1 mutation profiles and clinical characteristics of
breast cancer in TCGA and METABRIC cohorts were downloaded from
cBioPortal (http://www.cbioportal.org) (11). A total of 1,108 patients with breast
cancer were included in TCGA Pan-Cancer analysis project dataset
(7). In the METABRIC cohort
(10), 2,509 eligible patients with
breast cancer for whom MAP3K1 mutation profiles and clinical
information were included. Patients with breast cancer with
complete MAP3K1 somatic mutation profiles and
clinicopathological characteristics were included in this study.
For the statistical analysis, individuals with missing variables
were excluded. Somatic mutations in the GDPH, TCGA and METABRIC
cohorts were analyzed to compare MAP3K1 mutation
frequencies. In patient survival analysis, patients with missing
MAP3K1 mutation data and missing follow-up were excluded.
OncoPrint data was generated from cBioPortal. The expression
heatmap of genes in the METABRIC cohort are shown as mRNA
expression Z-scores, which were defined as mRNA expression compared
to the expression distribution of each gene tumors, that are
diploid for this gene.
All experiments described in this study were
approved by The Ethics Committee of The Guangdong Provincial
People's Hospital (approval no. GDREC2014122H). All patients
provided written informed consent to participate in this study.
DNA extraction from the tissues
DNA was extracted from 412 tumor samples using a
QIAamp DNA FFPE tissue kit (cat. no. 56404; Qiagen, Inc.) according
to the manufacturer's standard protocol. The concentration and
purity of DNA were measured using a Qubit dsDNA assay kit (Life
Technologies; Thermo Fisher Scientific, Inc.) prior to mutation
analysis. The loading concentration of the final library was 4
nmol.
NGS library preparation and
sequencing
In the current study, sequencing data were obtained
from primary tissue samples. DNA fragmentation was performed using
a M220 (Covaris, Inc.), followed by end repair, phosphorylation and
adaptor ligation. Fragments 200–400 bp in size were selected using
AMPure beads (Agencourt AMPure XP kit; Beckman Coulter, Inc.),
followed by hybridization with capture probe baits, hybrid
selection with magnetic beads and PCR amplification. A total of 1
to 5 µg PCR products was used for hybridization with NimbleGen
SeqCap EZ Hybridization and Wash kits (cat. no. 06953247001; Roche
Diagnostics). The captured DNA fragments from the previous step
were amplified, and the PCR products were purified with the AxyPrep
Mag PCR clean up kit (cat. no. MAG-PCR-CL-250; Corning, Inc.). The
quality of the library was then evaluated using the bioanalyzer
(Agilent Technologies, Inc.) and Real-Time PCR System (ABI
StepOnePlus; 4376600; Thermo Fisher Scientific, Inc.). Indexed
samples were sequenced on a NextSeq500 sequencer (Illumina, Inc.)
with a NextSeq 500 high output kit V2 (cat. no. FC-404-2002;
Illumina, Inc.) with paired-end reads (150 cycles) in a Clinical
Laboratory Improvement Amendments/College of American
Pathologists-certified laboratory using a panel consisting of 520
cancer-related genes, spanning 1.64 megabases of the human genome
(OncoScreen Plus panel; Guangzhou Burning Rock Medical Laboratory
Co., Ltd). The panel was designed to capture whole exons of 312
genes and critical exons, introns and promoter regions of the
remaining 208 genes.
Sequencing data analysis
Sequence data were mapped to the human genome (hg19)
using Burrows-Wheeler Aligner v.0.7.10 (13). Local alignment optimization, variant
calling and annotation were performed using GATK v.3.2 (https://gatk.broadinstitute.org) and VarScan
v.2.4.3 (http://varscan.sourceforge.net) (14). Variants were filtered using the
VarScan fpfilter pipeline, whereby loci with depth <100 were
filtered out. Moreover, ≥5 supporting reads were needed for
insertion-deletions (indels); while 8 supporting reads were needed
for single nucleotide variations to be called. According to the
ExAC (http://http://exac.broadinstitute.org), 1,000 Genomes
(https://www.internationalgenome.org),
dbSNP (http://www.ncbi.nlm.nih.gov/SNP) and ESP6500SI–V2
(http://www.openbioinformatics.org/annovar/download/hg19_1000g2015aug.zip)
databases, variants with population frequency >0.1% were grouped
as single nucleotide polymorphism and excluded from further
analysis. Remaining variants were annotated with ANNOVAR (15) and SnpEff v.3.6 (16). DNA translocation analysis was
performed using Factera v.1.4.3 (17). Data processing methods are also
described in previous reports (18–20). All
sequencing data are available in the NODE repository under the
accession code OEP001295 (http://www.biosino.org/node/project/detail/OEP001295).
Mutation classification
Capture-based targeted sequencing of 412 tumour
tissue samples and paired peripheral blood samples was performed
using a panel consisting of 520 cancer-related genes. The mutations
were classified according to the predicted effect using ANNOVAR
(15) and SnoEff v.3.6 (16) on the protein function as follows: i)
Mis-sense mutations in the DNA-binding motif (DBM); ii) mis-sense
mutations outside of the DBM; and iii) and non-mis-sense mutations
(including splice, in-frame, frameshift and nonsense mutations).
Samples with no somatic gene alteration were classified as
wild-type (WT).
Prognostic analysis of intrinsic
molecular subtypes and DNA promoter methylation
The prognostic analysis of MAP3K1 expression
in different molecular subtypes of breast cancer was performed
using Breast Cancer Gene-Expression Miner v4.4 (http://bcgenex.ico.unicancer.fr) as previously
described (21,22). This prognostic analysis tool based on
intrinsic molecular subtypes is used for the evaluation of the
prognostic factors for a given gene within groups of patients
defined by different subtype clustering models. Molecular subtypes
were classified into four groups based on three subtype clustering
models (SCMOD1, SCMOD2 and SCMGENE). For example, SCMOD models were
produced by analyzing the public database cohorts enrolling
>2,100 patients based on ER and HER2 module scores (23–28).
These groups represented the following intrinsic molecular
subtypes: i) ER−/HER2−; ii)
ER−/HER2+; iii)
ER+/HER2−; and iv)
ER+/HER2+, corresponding roughly to the
intrinsic basal-like, HER2, combined luminal A/B (HER2-), and
luminal B (HER2+) subtypes, respectively (25).
The prognostic effect of MAP3K1 expression
was evaluated using univariate Cox proportional hazards model and
Kaplan-Meier curves. A univariate Cox analysis table is also
provided for robust classification.
The DNA promoter methylation profile from 890
patients in TCGA-BRCA cohort was obtained from UALCAN (http://ualcan.path.uab.edu) (29) to investigate the epigenetic
regulation of MAP3K1 expression in breast cancer. In short,
BRCA DNA methylation array, mRNA expression datasets and
corresponding clinical information from TCGA firehose browser
(http://firebrowse.org) was obtained from UALCAN.
The DNA methylation array data from tumor tissues from 793 patients
with BRCA and 97 adjacent tumor tissues were analyzed.
Statistical analysis
Patient characteristics and sequencing results are
summarized using descriptive statistics, including the mean and
standard deviations for continuous data. Continuous variables were
compared using Student's t-test or Mann-Whitney's U-test.
Categorical data were compared using the χ2 test.
P-values were two-tailed and P<0.05 was considered to indicate a
statistically significant difference. Survival analysis was
conducted using the Kaplan-Meier method, and groups were compared
using the log-rank test. All statistical analyses were conducted
using Statistical Product and Service Solutions 20.0 (IBM
Corp.).
Results
Characterization of MAP3K1
mutations
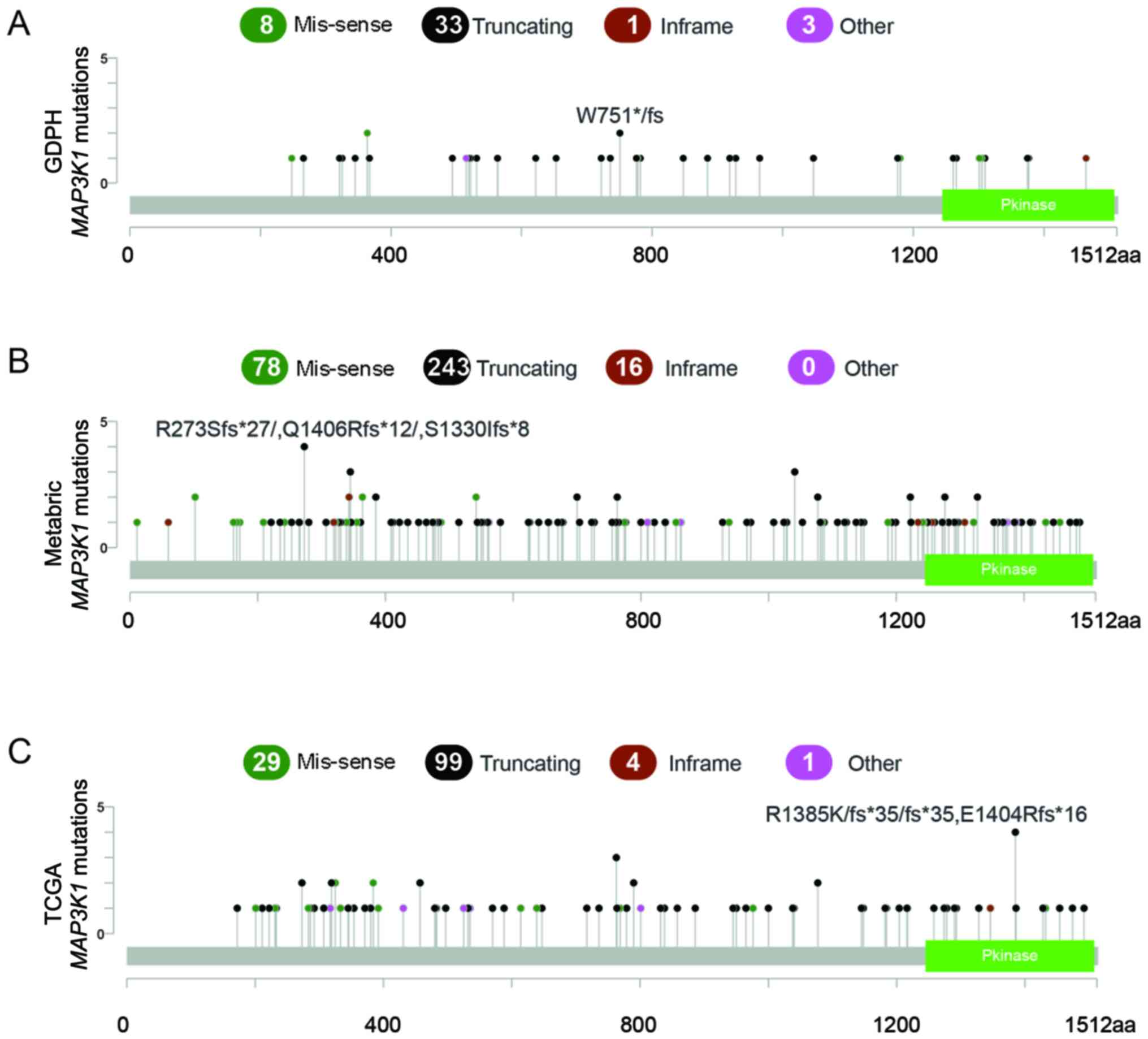
According to the NGS results obtained from the GDPH
cohort, 45 MAP3K1 mutations were observed in 35 patients
(8.5%). Moreover, 8 of the 35 patients (21.6%) concurrently carried
>1 MAP3K1 mutation. Among these eight patients, seven
patients had dual MAP3K1 mutations and one patient had a
triplet MAP3K1 mutation. The majority of the mutations were
truncating mutations (n=33; 74.4%), followed by missense mutations
(n=8; 17.8%) (Fig. 1A).
The MAP3K1 mutation profiles were compared
between GDPH, METABRIC and TCGA cohorts. MAP3K1 mutation
frequencies in the METABRIC and TCGA cohorts were similar to the
mutation frequencies in the GDPH cohort, namely, 9.7% in METABRIC
(number of patients, 244; number of mutations, 337) and 7.9% in
TCGA (number of patients, 88; number of mutations, 133). Similarly,
truncating mutations were the major mutation type, accounting for
72.1% in METABRIC and 74.4% in TCGA (Fig. 1B and C).
The mutated domains in the three cohorts are shown
in Fig. 1. Mutation hotspots were
not uniformly distributed across the three cohorts. No major
hotspot was identified in the mutated domains. For example, the
most highly recurrent hotspot was Arg364Trp in the METABRIC cohort
(n=6), which only occurred in two patients in the GDPH cohort.
Spectrum of MAP3K1 mutations in
patients with different molecular subtypes of breast cancer
The relationship between MAP3K1 mutations and
clinicopathological characteristics of the three cohorts are shown
in Table I. Mutations in
MAP3K1 were predominant in the luminal A breast cancer
subtype, followed by the luminal B breast cancer subtype, both in
TCGA and the METABRIC datasets (P<0.001; Table I). However, there was no significant
association in the GDPH cohort (P=0.179). Only four of 35 patients
(11.4%) had a hormone receptor (HR)− disease and
MAP3K1 mutations were also more dominant in luminal A
subtype, compared with luminal B, HER2+ and
triple-negative breast cancer (TNBC) (23/35; 65.7%). The
non-predominant association with molecular subtype may be due to
the small sample size. In addition, a lower pathological grade was
observed in the MAP3K1 mutant group in the GDPH and the
METABRIC datasets.
 | Table I.Association between MAP3K1
mutations and the clinicopathological characteristics of the three
cohorts. |
Table I.
Association between MAP3K1
mutations and the clinicopathological characteristics of the three
cohorts.
| A, GDPH cohort
(n=412) |
|---|
|
|---|
| Clinicopathological
characteristic | Wild-type | Mutant | P-value |
|---|
| Mean age ± SD,
years | 48.9±10.8 | 51.3±9.7 | 0.207 |
| Type, n
(%) |
|
| 0.863 |
|
Ductal | 331 (87.8) | 30 (85.7) |
|
|
Lobular | - | - | - |
|
Other | 44 (11.7) | 5
(14.3) | - |
|
Missing | 2 (0.5) | 0 (0) |
|
| Node status, n
(%) |
|
| 0.979 |
|
pN0 | 184 (48.8) | 17 (48.6) |
|
|
pN1-pN3 | 193 (51.2) | 18 (51.4) |
|
|
Missing | 0 (0) | 0 (0) |
|
| TNM stage, n
(%) |
|
| 0.304 |
| 1 | 91 (24.1) | 11 (31.4) |
|
| 2 | 175 (46.4) | 16 (45.7) |
|
| 3 | 85 (22.5) | 6
(17.1) |
|
| 4 | 25 (6.6) | 2 (5.7) |
|
|
Missing | 1 (0.3) | 0 (0) |
|
| Pathological grade,
n (%) |
|
| 0.032 |
| 1 | 13 (3.4) | 2 (5.7) |
|
| 2 | 169 (44.8) | 22 (62.9) |
|
| 3 | 189 (50.1) | 9
(25.7) |
|
|
Missing | 6 (1.6) | 2 (5.7) |
|
| Estrogen receptor,
n (%) | | | 0.116 |
|
Positive | 265 (70.3) | 29 (82.9) |
|
|
Negative | 112 (29.7) | 6
(17.1) |
|
|
Missing | 0 (0) | 0 (0) |
|
| Progesterone
receptor, n (%) | | | 0.155 |
|
Positive | 246 (65.3) | 27 (77.1) |
|
|
Negative | 131 (34.7) | 8
(22.9) |
|
|
Missing | 0 (0) | 0 (0) |
|
| Hormone receptor, n
(%) | | | 0.056 |
|
Positive | 279 (74.0) | 31 (88.6) |
|
|
Negative | 98 (26.0) | 4
(11.4) |
|
|
Missing | 0 (0) | 0 (0) |
|
| HER2, n (%) |
| | 0.889 |
|
Positive | 108 (28.7) | 9
(25.7) |
|
|
Negative | 268 (71.1) | 26 (74.3) |
|
|
Missing | 1 (0.3) | 0 (0) |
|
| Subtype, n (%) |
|
| 0.179 |
|
LumA | 76 (20.2) | 11 (31.4) |
|
|
LumB | 203 (53.8) | 20 (57.1) |
|
|
HER2+ | 47 (12.5) | 2 (5.7) |
|
|
TNBC | 51 (13.5) | 2 (5.7) |
|
|
Claudinlow | - | - |
|
|
Basal | - | - |
|
|
Missing | 0 (0) | 0 (0) |
|
| HR/HER2 status, n
(%) |
|
| 0.227 |
|
HR+/HER2− | 214 (56.8) | 23 (65.7) |
|
|
HR+/HER2+ | 65 (17.2) | 8
(22.9) |
|
|
HR−/HER2+ | 47 (12.5) | 2 (5.7) |
|
|
HR−/HER2− | 51 (13.5) | 2
(5.7) |
|
|
Missing | 0 (0) | 0 (0) |
|
|
| B, TCGA
(n=1,108) |
|
|
Clinicopathological
characteristic |
Wild-type | Mutant | P-value |
|
| Mean age ± SD,
years | 58.1±13.3 | 60.9±12.6 | 0.064 |
| Type, n
(%) |
|
| 0.054 |
|
Ductal | 747 (73.2) | 74 (84.1) |
|
|
Lobular | 195 (19.1) | 12 (13.6) | - |
|
Other | 78 (7.6) | 2 (2.3) | - |
|
Missing | 0 (0) | 0 (0) |
|
| Node status, n
(%) |
|
| 0.917 |
|
pN0 | 476 (46.7) | 40 (45.5) |
|
|
pN1-pN3 | 523 (51.3) | 45 (51.1) |
|
|
Missing | 21 (2.0) | 3 (3.4) |
|
| TNM stage, n
(%) |
|
| 0.647 |
| 1 | 165 (16.2) | 18 (20.5) |
|
| 2 | 581 (57.0) | 45 (51.1) |
|
| 3 | 233 (22.8) | 18 (20.5) |
|
| 4 | 18 (17.6) | 2 (2.3) |
|
|
Missing | 23 (2.3) | 5 (5.7) |
|
| Pathological grade,
n (%) |
|
| - |
| 1 | - | - | - |
| 2 | - | - | - |
| 3 | - | - | - |
| ER, n (%) |
|
| 0.043 |
|
Positive | 740 (72.5) | 74 (84.1) |
|
|
Negative | 242 (23.7) | 12 (13.6) |
|
|
Missing | 38 (3.7) | 2 (2.3) |
|
| PR, n (%) | | | 0.017 |
|
Positive | 638 (62.5) | 67 (76.1) |
|
|
Negative | 342 (33.5) | 18 (20.4) |
|
|
Missing | 40 (3.9) | 3 (3.4) |
|
| HR, n (%) | | | 0.01 |
|
Positive | 765 (75.0) | 76 (86.4) |
|
|
Negative | 255 (25.0) | 12 (13.6) |
|
|
Missing | 0 (0) | 0 (0) |
|
| HER2, n (%) | | | 0.53 |
|
Positive | 170 (16.7) | 15 (17.0) |
|
|
Negative | 656 (64.3) | 57 (64.8) |
|
|
Missing | 194 (19.0) | 16 (18.2) |
|
| Subtype, n (%) |
|
| - |
|
LumA | - | - |
|
|
LumB | - | - |
|
|
HER2+ | - | - |
|
|
TNBC | - | - |
|
|
Claudinlow | - | - |
|
|
Basal | - | - |
|
|
Missing | - | - |
|
| HR/HER2 status, n
(%) |
|
| 0.106 |
|
HR+/HER2− | 514 (50.4) | 51 (58.0) |
|
|
HR+/HER2+ | 131 (12.8) | 14 (16.0) |
|
|
HR−/HER2+ | 39 (3.8) | 1 (1.1) |
|
|
HR−/HER2− | 142 (13.9) | 6 (6.8) |
|
|
Missing | 194 (19.0) | 16 (18.2) |
|
|
| C, METABRIC
(n=2,509) |
|
|
Clinicopathological
characteristic |
Wild-type | Mutant | P-value |
|
| Mean age ± SD,
years | 60.2±13.0 | 62.3±12.8 | 0.018 |
| Type, n
(%) |
|
| 0.887 |
|
Ductal | 1,690 (74.6) | 175 (71.7) |
|
|
Lobular | 172
(7.6) | 20 (8.2) | - |
|
Other | 403
(17.8) | 49
(20.1) | - |
|
Missing | 0
(0) | 0
(0) |
|
| Node status, n
(%) |
|
| 0.106 |
|
pN0 | 1,087 (48.0) | 109 (44.7) |
|
|
pN1-pN3 | 930
(41.0) | 117 (48.0) |
|
|
Missing | 248
(10.9) | 18 (7.4) |
|
| TNM stage, n
(%) |
|
| 0.933 |
| 1 | 566
(25.0) | 64
(26.2) |
|
| 2 | 887
(39.2) | 92
(37.7) |
|
| 3 | 131 (5.8) | 13 (5.3) |
|
| 4 | 10
(0.4) | 1
(0.4) |
|
|
Missing | 671
(29.6) | 74
(30.3) |
|
| Pathological grade,
n (%) |
|
| <0.001 |
| 1 | 173 (7.6) | 41
(16.8) |
|
| 2 | 854
(37.7) | 122 (50.0) |
|
| 3 | 1,128 (49.8) | 70
(28.7) |
|
|
Missing | 110 (4.9) | 11 (4.5) |
|
| ER, n (%) |
|
| <0.001 |
|
Positive | 1,610 (71.1) | 215 (88.1) |
|
|
Negative | 617
(27.2) | 27
(11.1) |
|
|
Missing | 38
(1.7) | 2
(0.8) |
|
| PR, n (%) |
|
| <0.001 |
|
Positive | 887 (39.2) | 152 (62.3) |
|
|
Negative | 887 (39.2) | 54
(22.1) |
|
|
Missing | 491 (21.7) | 38
(15.6) |
|
| HR, n (%) | - | - | - |
|
Positive | - | - | - |
|
Negative | - | - | - |
|
Missing | - | - | - |
| HER2, n (%) |
|
| <0.001 |
|
Positive | 1,022 (45.1) | 153 (62.7) |
|
|
Negative | 752
(33.2) | 53
(21.7) |
|
|
Missing | 491
(21.7) | 38
(15.6) |
|
| Subtype, n (%) |
|
| <0.001 |
|
LumA | 589 (26.0) | 111 (45.5) |
|
|
LumB | 433 (19.1) | 42
(17.2) |
|
|
HER2+ | 208 (9.2) | 16 (6.6) |
|
|
TNBC | - | - |
|
|
Claudinlow | 209 (9.2) | 9
(3.7) |
|
|
Basal | 198 (8.7) | 11 (4.5) |
|
|
Missing | 628
(27.7) | 55
(22.5) |
|
| HR/HER2 status, n
(%) |
|
|
|
|
HR+/HER2− | - | - | - |
|
HR+/HER2+ | - | - | - |
|
HR−/HER2+ | - | - | - |
|
HR−/HER2− | - | - | - |
|
Missing |
|
|
|
Prognostic implications of MAP3K1
mutations in breast cancer
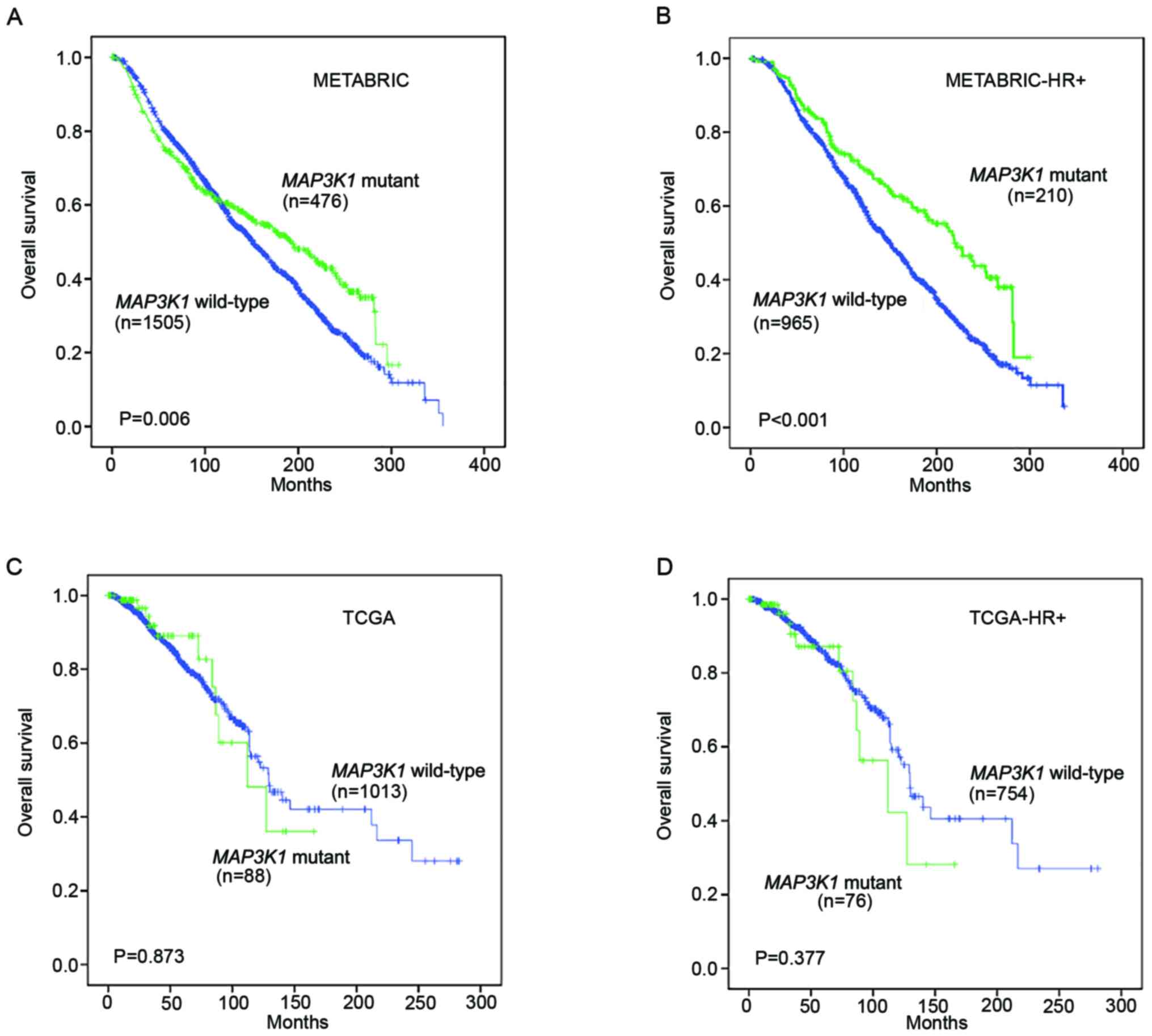
Kaplan-Meier survival analysis was performed on the
METABRIC and TCGA datasets to assess the prognostic value of
MAP3K1 mutations in the follow-up of patients with breast
cancer.
In the METABRIC cohort, patients with MAP3K1
mutations experienced a prolonged OS compared with patients without
MAP3K1 mutations (P=0.006; Fig.
2A). In patients with HR+ breast cancer, a
significantly longer OS was also observed in the MAP3K1
mutation group (P<0.001; Fig.
2B). Unexpectedly, TCGA cohort and TCGA HR+ mutation
subgroup did not display a significant difference in OS (P=0.873
and P=0.377, respectively; Fig. 2C and
D).
Interactions between MAP3K1, MAP2K1,
MAP2K2, MAP2K4 and PIK3CA mutations
OncoPrint data for MAP3K1, MAP2K4 and PI3K
catalytic subunit α (PIK3CA) mutations in the METABRIC
cohort are shown in Fig. 3.
MAP3K1 mutations in breast cancer were mutually exclusive
with respect to mutations in MAP2K4, MAP2K1 and
MAP2K2. Indeed, out of 217 patients who had a MAP3K1
mutation, only 3 also had MAP2K4 mutations, none had a
MAP2K1 mutation and only one had a MAP2K2 mutation.
Nevertheless, 144 PIK3CA mutations were observed in the 217
patients with MAP3K1 mutations, indicating that
MAP3K1 alterations were concurrently accompanied by
PIK3CA mutations (as indicated by the color overlap in the
second and sixth rows; Fig. 3).
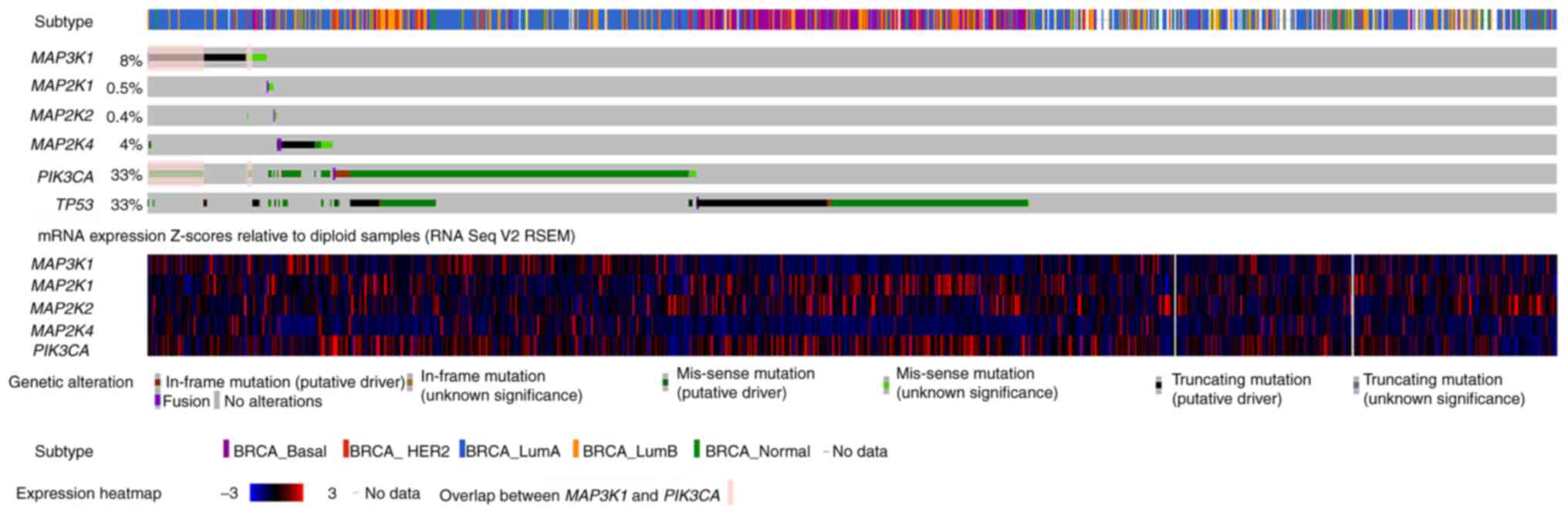 | Figure 3.Characterization of MAP3K1,
MAP2K1, MAP2K2, MAP4K2 and PIK3CA mutations in patients
with breast cancer from the METABRIC cohort. The OncoPrint of
MAP3K1, MAP2K4 and PIK3CA genes is shown, in which
individual samples and genes in the Molecular Taxonomy of Breast
Cancer International Consortium cohort are presented in columns and
rows, respectively. Subtype assignments of the patients are shown
in the first row. mRNA expression is shown in the next 6 rows.
Overlap between MAP3K1 and PIK3CA mutation are shown
in pink. The data were obtained from and visualized using the
cBioPortal database (accession number, EGAS00001001753) with
modifications. RSEM was used for accurate transcript quantification
from RNA Seq data. BRCA, breast cancer; HER2, human epidermal
growth factor receptor 2; PIK3CA, PI3K catalytic subunit α;
TP53, tumor protein 53; RNA Seq, RNA sequencing; RSEM, RNA Seq by
Expectation Maximization. |
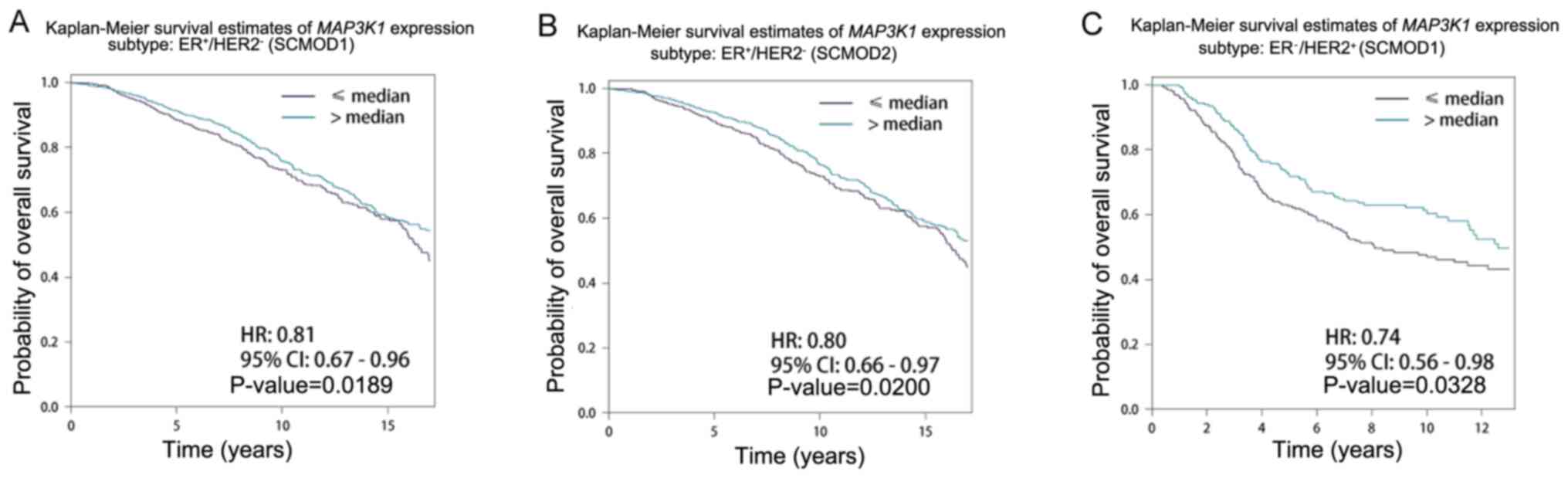
Effect of MAP3K1 expression on the
prognosis of patients with different intrinsic molecular subtypes
of breast cancer
The effect of MAP3K1 expression on patients
with different intrinsic molecular subtypes was further examined.
Patients were grouped into two groups: High (>median) and low
expression (<median) groups. High MAP3K1 expression was
associated with longer survival time in the
ER+/HER2− and ER−/HER2+
subgroups (Table II).
Representative survival curves are shown in Fig. 4. Patients with higher MAP3K1
expression had longer survival time compared with that in patients
with lower MAP3K1 expression in the ER+/HER2-subgroup of
SCMOD1 (P=0.189; Fig. 4A) and SCMOD2
(P=0.020; Fig. 4B), and ER-/HER2+
subgroup of SCMOD1 (P=0.0328; Fig.
4C).
| Table II.Univariate Cox analysis of
MAP3K1 expression level of overall survival in patients with
different breast cancer molecular subtypes using three SCMs. |
Table II.
Univariate Cox analysis of
MAP3K1 expression level of overall survival in patients with
different breast cancer molecular subtypes using three SCMs.
| A, SCMOD1 |
|---|
|
|---|
| Molecular subtype,
high vs. low | HR (95% CI) |
+P-value |
|---|
|
ER−/HER2− | 0.87
(0.68-1.11) | 0.2733 |
|
ER−/HER2+ | 0.74
(0.56-0.98) | 0.0328 |
|
ER+/HER2− | 0.81
(0.67-0.96) | 0.0189 |
|
ER+/HER2+ | 0.88
(0.75-1.03) | 0.1027 |
|
| B,
SCMOD2 |
|
| Molecular
subtype, high vs. low | HR (95%
CI) | P-value |
|
ER−/HER2− | 0.91
(0.72-1.14) | 0.3931 |
|
ER−/HER2+ | 0.78
(0.60-1.00) | 0.0504 |
|
ER+/HER2− | 0.80
(0.66-0.97) | 0.0200 |
|
ER+/HER2+ | 0.87
(0.74-1.03) | 0.0991 |
|
| C,
SCMGENE |
|
| Molecular
subtype, high vs. low | HR (95%
CI) | P-value |
|
|
ER−/HER2− | 0.85
(0.69-1.04) | 0.1192 |
|
ER−/HER2+ | 0.87
(0.68-1.12) | 0.2929 |
|
ER+/HER2− | 0.91
(0.76-1.09) | 0.3063 |
|
ER+/HER2+ | 0.87
(0.74-1.03) | 0.1000 |
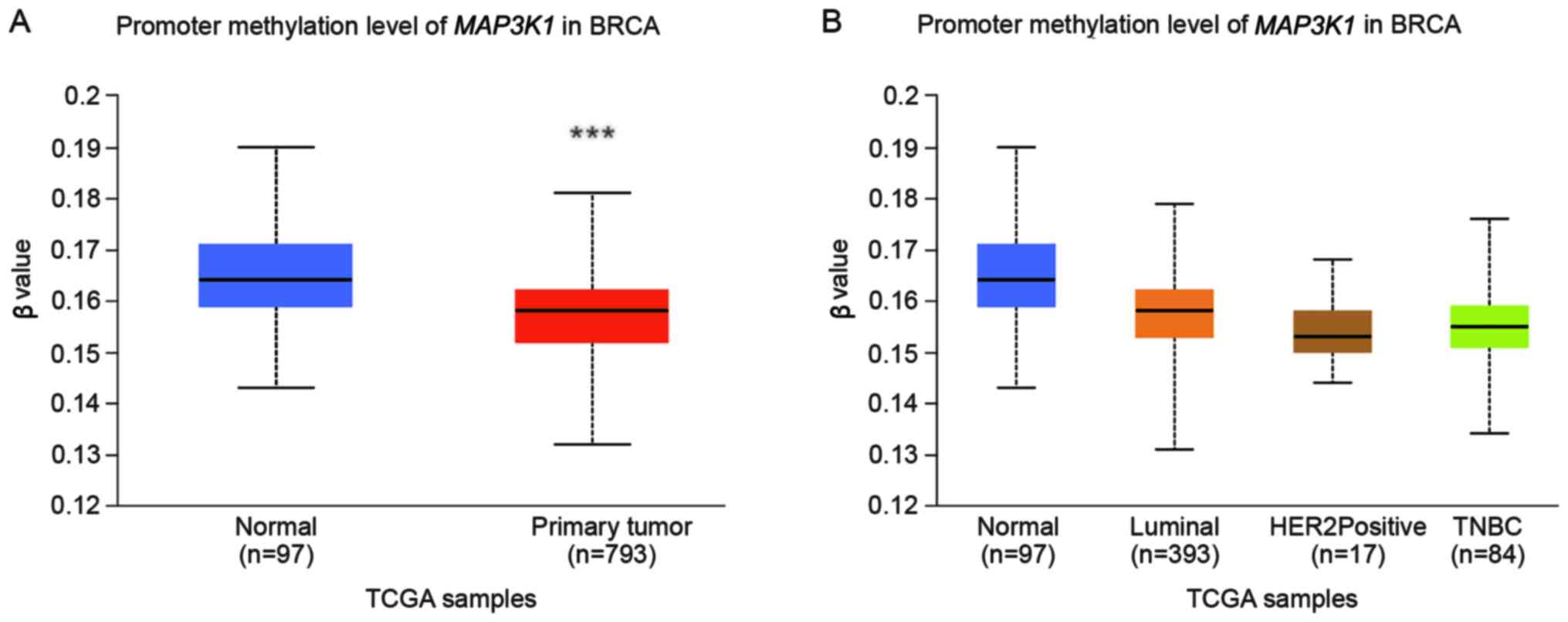
MAP3K1 promoter methylation in breast
cancer
The DNA methylation profile was analysed using
UALCAN to further explore the epigenetic regulation of
MAP3K1 methylation in breast cancer. Significant
hypermethylation of MAP3K1 was observed (Fig. 5A; P<0.0001). However, no
difference in methylation was observed between the normal, luminal
A and B, HER2+ and TNBC subgroups (Fig. 5B).
Discussion
The present findings identified MAP3K1
mutations in Chinese patients with breast cancer, as well as in
public datasets. Given the differences in cancer biology in
patients from different ethnic groups, the MAP3K1 mutation
spectrum in Chinese patients was compared with data available in
public databases. The current retrospective study is the first
comprehensive report to analyse the clinicopathological
characteristics of Chinese patients with MAP3K1 mutations in
a Chinese cohort from a single centre. In the GDPH cohort, 412
breast cancer samples were analysed using NGS and further compared
with data from TCGA and METABRIC cohorts. A similar prevalence in
MAP3K1 mutations was observed in these three cohorts,
whereas the mutated domains and mutation hotspots did not overlap.
A recent study including 122 Chinese patients with breast cancer or
ovarian cancer reported a higher frequency of MAP3K1 mutation
(14.3%) (30). Although
MAP3K1 is among the most frequently mutated genes in cancer,
to the best of our knowledge, no study has reported frequent
hotspot- or domain-specific alterations associated with
MAP3K1 mutations, consistent with the results of the current
study. Notably, 21.6% tumors concurrently carried >1 mutation in
MAP3K1. Among those tumors, seven tumors contained dual
MAP3K1 mutations and one tumor contained triplet mutations.
Although MAP3K1 mutations potentially represent a protective
marker in breast cancer, it was not possible to determine whether
the presence of concurrent dual MAP3K1 mutations had
different clinicopathological characteristics from single mutations
due to the small cohort size.
Breast cancer is a heterogeneous disease.
Distributions of MAP3K1 mutations were analysed. In TCGA and
METABRIC cohorts, MAP3K1 mutations were more frequently
enriched in patients with HR+ breast cancer than in
patients with other subtypes including (HER2+ and TNBC),
consistent with previous studies (9,31). Only
4 of the 35 patients who carried the MAP3K1 mutation in the GDPH
cohort had HR− disease. The insignificance of the
difference in the GDPH cohort may be due to the small cohort
size.
Ellis et al (32) conducted whole-genome sequencing and
pathway analyses to compare patients who were sensitive or
resistant to neoadjuvant treatment with an aromatase inhibitor.
Based on the study, mutations in the MAP3K1 gene were found
to be related to indolent biological features and low proliferation
rates. Therefore, MAP3K1 mutations have been proposed to
correlate with favorable outcomes in patients with breast cancer
(7). Similarly, a two-gene genomic
signature combining with MAP3K1 mutation and FGFR1
amplification was associated with decreased risk of distant
metastasis in patients with HR+/HER2− breast
cancer (33).
MAP3K1-targeting drugs have been developed, including
binimetinib (34), fostamatinib
(35) and AZD-8330 (36). Binimetinib is used to treat
metastatic melanoma and unresectable melanoma, while fostamatinib
is used to treat chronic immune thrombocytopenia. To date, Food and
Drug Administration-approved MAP3K1-targeting drugs are not
available for the treatment of breast cancer.
MAP3K1 activates the JNK pathway by
selectively phosphorylating and activating MAP2K4 (37,38).
Deficiency in the JNK signaling pathway results in defective
apoptosis, leading to unresponsiveness to environmental and
genotoxic stresses (39). Ellis
et al (7) demonstrated that
MAP2K4 mutations were associated with MAP3K1
dysfunction, but mutations in two genes associated with one pathway
were rarely observed in a single patient. Tissues with
MAP3K1 mutation in the METABRIC cohort did not coincide with
mutations in MAP2K4. Considering the differences in samples
with MAP3K1 and MAP2K4 mutations, loss of function of
either gene in the JNK pathway may result in the deregulation of
the other gene and the pathway as a whole.
In addition, MAP3K1 and PIK3CA
mutations co-associated in both the METABRIC and TCGA cohorts. In
the METABRIC cohort, PIK3CA mutations were accompanied with
MAP3K1 mutations in 66.4% of patients. A previous study has
identified MAP3K1 and MAP2K2 as powerful predictors
of responsiveness to MEK inhibition in patient-derived xenograft
models (40). Loss-of-function
mutations in MAP3K1 resulted in higher sensitivity to MEK
inhibition, demonstrating that MEK inhibitors could represent
potential therapeutic targets for tumors with MAP3K1 mutations
(40).
In the present study, MAP3K1 expression was
confirmed to be associated with OS in different subtype clustering
models of breast cancer, supporting the prognostic value of
MAP3K1. However, high MAP3K1 was only a protective
biomarker in patients with ER+/HER2- and ER-/HER2+ disease.
To the best of our knowledge, MAP3K1
methylation has not yet been reported in breast tumors. In the
current study, the methylation of the MAP3K1 gene was
decreased in breast cancer tissue samples from TCGA, compared with
normal tissue. This result may partially explain the epigenetic
regulation of MAP3K1 in breast cancer, although further
studies are needed to confirm this finding.
The current study has a number of limitations. Since
the present findings were obtained from a retrospective,
single-center study, the limited cohort size prevented robust
comparison between the Chinese patient cohort and TCGA and METABRIC
cohorts. As indicated in our previous report (19), the baseline characteristics of the
patients enrolled in the three cohorts, including age, menopausal
status, histological grade, ER status, PR status and HER2 status,
were significantly different. Moreover, 5.5% patients in TCGA
cohort were Asian (60/1084). Ethnicity was not reported for the
METABRIC cohort. All patients in the GDPH cohort were Chinese and
98.8% were Han Chinese (407/412). In addition, due to the absence
of complete follow-up and prognostic data in the GDPH cohort, the
effect of MAP3K1 mutations on the survival of the Chinese
patients could not be established. The methylation data for
MAP3K1 also lacked from the GDPH cohort. Further multicenter
prospective studies with complete follow-up data are needed to
verify the present findings.
Altogether, the present findings identified
MAP3K1 mutations in Chinese patients with breast cancer, as
well as public datasets. MAP3K1 mutations were mainly
detected in HR+/HER2− breast cancer and could
represent possible prognostic factors. MAP3K1 expression was
associated with OS in the HR+ subgroup. Furthermore, a
lower level of MAP3K1 methylation was observed in patients
with breast cancer. A larger sample size is needed to evaluate the
clinical applicability of MAP3K1 mutations in patients with
HR+ breast cancer. The results may provide insight into
the pathophysiological mechanism of breast cancer and the
development of novel therapeutic treatments.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from The Project of Doctoral
Scientific Research of Guangdong Provincial People's Hospital
(grant. no. 2020bq12), The National Natural Science Foundation of
China (grant nos. 81602645, 81702783, 81071851 and 81001189), The
Natural Science Foundation of Guangdong Province (grant nos.
2016A030313768, 2017A030310574 and 2018A030313292) and research
funds from The Guangzhou Municipal Science and Technology Project
(grant nos. 201707010418 and 201804010430).
Availability of data and materials
The datasets generated and/or analyzed in the GDPH
cohort are available in the NODE repository (http://www.biosino.org/node) under accession no.
OEP001295. Data for the MAP3K1 mutation profiles and
clinical characteristics of breast cancer in TCGA and METABRIC
cohorts were extracted from cBioPortal (http://www.cbioportal.org).
Authors' contributions
CL and NL conceived the study. YW, GZ, KL, LC, CR,
XL collected the data. GZ, YW, BC and NL acquired funding. CL, LW,
MJ, HM, JL, WX and XL carried out the experiments. CL, GZ, BC, JL,
XL and NL designed the methodology. YW, BC, KL, LC, CR, LW and WX
oversaw project administration; YW, KL and LC provided software. CL
wrote the original draft; CL, GZ, KL, LC, CR, XL and NL reviewed
and edited the manuscript. CL and NL confirm the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
This study was approved by The Ethics Committee of
The Guangdong Provincial People's Hospital (approval no.
GDREC2014122H). All patients provided written informed consent to
participate in this translational study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Luo CY, Li N, Lu B, Cai J, Lu M, Zhang YH,
Chen HD and Dai M: Global and regional trends in incidence and
mortality of female breast cancer and associated factors at
national level in 2000 to 2019. Chin Med J (Engl). September
28–2021.(Epub ahead of print). doi: 10.1097/CM9.0000000000001814.
View Article : Google Scholar
|
|
2
|
Waks AG and Winer EP: Breast cancer
treatment: A review. JAMA. 321:288–300. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oh DY and Bang YJ: HER2-targeted
therapies-a role beyond breast cancer. Nat Rev Clin Oncol.
17:33–48. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Andre F, Ciruelos E, Rubovszky G, Campone
M, Loibl S, Rugo HS, Iwata H, Conte P, Mayer IA, Kaufman B, et al:
Alpelisib for PIK3CA-mutated, hormone receptor-positive advanced
breast cancer. N Engl J Med. 380:1929–1940. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hortobagyi GN, Chen D, Piccart M, Rugo HS,
Burris HA III, Pritchard KI, Campone M, Noguchi S, Perez AT, Deleu
I, et al: Correlative analysis of genetic alterations and
everolimus benefit in hormone receptor-positive, human epidermal
growth factor receptor 2-negative advanced breast cancer: Results
from BOLERO-2. J Clin Oncol. 34:419–426. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gong X, Litchfield LM, Webster Y, Chio LC,
Wong SS, Stewart TR, Dowless M, Dempsey J, Zeng Y, Torres R, et al:
Genomic aberrations that activate D-type cyclins are associated
with enhanced sensitivity to the CDK4 and CDK6 inhibitor
abemaciclib. Cancer Cell. 32:761–776.e6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ellis MJ, Ding L, Shen D, Luo J, Suman VJ,
Wallis JW, Van Tine BA, Hoog J, Goiffon RJ, Goldstein TC, et al:
Whole-genome analysis informs breast cancer response to aromatase
inhibition. Nature. 486:353–360. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cancer Genome Atlas Network. Comprehensive
molecular portraits of human breast tumours. Nature. 490:61–70.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pham TT, Angus SP and Johnson GL: MAP3K1:
Genomic alterations in cancer and function in promoting cell
survival or apoptosis. Genes Cancer. 4:419–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pereira B, Chin SF, Rueda OM, Vollan HK,
Provenzano E, Bardwell HA, Pugh M, Jones L, Russell R, Sammut SJ,
et al: The somatic mutation profiles of 2,433 breast cancers
refines their genomic and transcriptomic landscapes. Nat Commun.
7:114792016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hammond ME, Hayes DF, Dowsett M, Allred
DC, Hagerty KL, Badve S, Fitzgibbons PL, Francis G, Goldstein NS,
Hayes M, et al: American Society of Clinical Oncology/College Of
American Pathologists guideline recommendations for
immunohistochemical testing of estrogen and progesterone receptors
in breast cancer. J Clin Oncol. 28:2784–2795. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wolff AC, Hammond ME, Hicks DG, Dowsett M,
McShane LM, Allison KH, Allred DC, Bartlett JM, Bilous M,
Fitzgibbons P, et al: Recommendations for human epidermal growth
factor receptor 2 testing in breast cancer: American society of
clinical oncology/college of American pathologists clinical
practice guideline update. J Clin Oncol. 31:3997–4013. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li H and Durbin R: Fast and accurate short
read alignment with Burrows-Wheeler transform. Bioinformatics.
25:1754–1760. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sahraeian SME, Mohiyuddin M, Sebra R,
Tilgner H, Afshar PT, Au KF, Bani Asadi N, Gerstein MB, Wong WH,
Snyder MP, et al: Gaining comprehensive biological insight into the
transcriptome by performing a broad-spectrum RNA-seq analysis. Nat
Commun. 8:592017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang K, Li M and Hakonarson H: ANNOVAR:
Functional annotation of genetic variants from high-throughput
sequencing data. Nucleic Acids Res. 38:e1642010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cingolani P, Platts A, Wang le L, Coon M,
Nguyen T, Wang L, Land SJ, Lu X and Ruden DM: A program for
annotating and predicting the effects of single nucleotide
polymorphisms, SnpEff: SNPs in the genome of Drosophila
melanogaster strain w1118; iso-2; iso-3. Fly (Austin). 6:80–92.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Newman AM, Bratman SV, Stehr H, Lee LJ,
Liu CL, Diehn M and Alizadeh AA: FACTERA: A practical method for
the discovery of genomic rearrangements at breakpoint resolution.
Bioinformatics. 30:3390–3393. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen X, Zhang G, Chen B, Wang Y, Guo L,
Cao L, Ren C, Wen L and Liao N: Association between histone lysine
methyltransferase KMT2C mutation and clinicopathological factors in
breast cancer. Biomed Pharmacother. 116:1089972019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang G, Wang Y, Chen B, Guo L, Cao L, Ren
C, Wen L, Li K, Jia M, Li C, et al: Characterization of frequently
mutated cancer genes in Chinese breast tumors: A comparison of
Chinese and TCGA cohorts. Ann Transl Med. 7:1792019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Stephens PJ, Tarpey PS, Davies H, Van Loo
P, Greenman C, Wedge DC, Nik-Zainal S, Martin S, Varela I, Bignell
GR, et al: The landscape of cancer genes and mutational processes
in breast cancer. Nature. 486:400–404. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jezequel P, Campone M, Gouraud W,
Guerin-Charbonnel C, Leux C, Ricolleau G and Campion L:
bc-GenExMiner: An easy-to-use online platform for gene prognostic
analyses in breast cancer. Breast Cancer Res Treat. 131:765–775.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jezequel P, Frenel JS, Campion L,
Guerin-Charbonnel C, Gouraud W, Ricolleau G and Campone M:
bc-GenExMiner 3.0: New mining module computes breast cancer gene
expression correlation analyses. Database (Oxford).
2013:bas0602013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sorlie T, Tibshirani R, Parker J, Hastie
T, Marron JS, Nobel A, Deng S, Johnsen H, Pesich R, Geisler S, et
al: Repeated observation of breast tumor subtypes in independent
gene expression data sets. Proc Natl Acad Sci USA. 100:8418–8423.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu Z, Fan C, Oh DS, Marron JS, He X,
Qaqish BF, Livasy C, Carey LA, Reynolds E, Dressler L, et al: The
molecular portraits of breast tumors are conserved across
microarray platforms. BMC Genomics. 7:962006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Desmedt C, Haibe-Kains B, Wirapati P,
Buyse M, Larsimont D, Bontempi G, Delorenzi M, Piccart M and
Sotiriou C: Biological processes associated with breast cancer
clinical outcome depend on the molecular subtypes. Clin Cancer Res.
14:5158–5165. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wirapati P, Sotiriou C, Kunkel S, Farmer
P, Pradervand S, Haibe-Kains B, Desmedt C, Ignatiadis M, Sengstag
T, Schütz F, et al: Meta-analysis of gene expression profiles in
breast cancer: Toward a unified understanding of breast cancer
subtyping and prognosis signatures. Breast Cancer Res. 10:R652008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Parker JS, Mullins M, Cheang MC, Leung S,
Voduc D, Vickery T, Davies S, Fauron C, He X, Hu Z, et al:
Supervised risk predictor of breast cancer based on intrinsic
subtypes. J Clin Oncol. 27:1160–1167. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Haibe-Kains B, Desmedt C, Rothe F, Piccart
M, Sotiriou C and Bontempi G: A fuzzy gene expression-based
computational approach improves breast cancer prognostication.
Genome Biol. 11:R182010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kwong A, Cheuk IW, Shin VY, Ho CY, Au CH,
Ho DN, Wong EY, Yu SW, Chen J, Chan KK, et al: Somatic mutation
profiling in BRCA-negative breast and ovarian cancer patients by
multigene panel sequencing. Am J Cancer Res. 10:2919–2932.
2020.PubMed/NCBI
|
|
31
|
Griffith OL, Spies NC, Anurag M, Griffith
M, Luo J, Tu D, Yeo B, Kunisaki J, Miller CA, Krysiak K, et al: The
prognostic effects of somatic mutations in ER-positive breast
cancer. Nat Commun. 9:34762018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ellis MJ, Suman VJ, Hoog J, Goncalves R,
Sanati S, Creighton CJ, DeSchryver K, Crouch E, Brink A, Watson M,
et al: Ki67 proliferation index as a tool for chemotherapy
decisions during and after neoadjuvant aromatase inhibitor
treatment of breast cancer: Results from the American college of
surgeons oncology group Z1031 trial (Alliance). J Clin Oncol.
35:1061–1069. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Carene D, Tran-Dien A, Lemonnier J, Dalenc
F, Levy C, Pierga JY, Jacot W, Canon JL, Richon C, Lacroix L, et
al: Association between FGFR1 copy numbers, MAP3K1 mutations, and
survival in axillary node-positive, hormone receptor-positive, and
HER2-negative early breast cancer in the PACS04 and METABRIC
studies. Breast Cancer Res Treat. 179:387–401. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koelblinger P, Dornbierer J and Dummer R:
A review of binimetinib for the treatment of mutant cutaneous
melanoma. Future Oncol. 13:1755–1766. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sweeny DJ, Li W, Clough J, Bhamidipati S,
Singh R, Park G, Baluom M, Grossbard E and Lau DT: Metabolism of
fostamatinib, the oral methylene phosphate prodrug of the spleen
tyrosine kinase inhibitor R406 in humans: Contribution of hepatic
and gut bacterial processes to the overall biotransformation. Drug
Metab Dispos. 38:1166–1176. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Haasbach E, Hartmayer C and Planz O:
Combination of MEK inhibitors and oseltamivir leads to synergistic
antiviral effects after influenza A virus infection in vitro.
Antiviral Res. 98:319–324. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yan M, Dai T, Deak JC, Kyriakis JM, Zon
LI, Woodgett JR and Templeton DJ: Activation of stress-activated
protein kinase by MEKK1 phosphorylation of its activator SEK1.
Nature. 372:798–800. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fanger GR, Johnson NL and Johnson GL: MEK
kinases are regulated by EGF and selectively interact with
Rac/Cdc42. EMBO J. 16:4961–4972. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wagner EF and Nebreda AR: Signal
integration by JNK and p38 MAPK pathways in cancer development. Nat
Rev Cancer. 9:537–549. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xue Z, Vis DJ, Bruna A, Sustic T, van
Wageningen S, Batra AS, Rueda OM, Bosdriesz E, Caldas C, Wessels
LFA and Bernards R: MAP3K1 and MAP2K4 mutations are associated with
sensitivity to MEK inhibitors in multiple cancer models. Cell Res.
28:719–729. 2018. View Article : Google Scholar : PubMed/NCBI
|