Introduction
Oral cancer has been reported to have an increased
prevalence in males and in older individuals; >90% of patients
with oral cancer are diagnosed with oral squamous cell carcinoma
(SCC) (1–3). The risk factors for oral cancer
include smoking, alcohol intake and betel nut chewing (4–6). In
2017, the Global Burden of Disease Study estimated the oral disease
to affect 3.5 billion population worldwide. Furthermore, oral
cancer has been reported was top 15 most common cancers worldwide
and caused ~180,000 deaths each year, according to the
International Agency for Research (7). It has also been reported that >600
new cases of oral cancer are diagnosed every year in Australia
(6,8). Furthermore, the incidence rate of
oral cavity SCC has been reported to be 146.2 cases per 100,000
person-years for areca/betel quid chewers in Taiwan (3,9). The
five-year survival rate reported was poor in a Taiwanese patient
study (10). In a previous study
of oral cavity SCC, it was reported that tongue SCC was more
prevalent compared with cancer of other oral cavity sites (by ~39%)
in a young population (11). In
order to increase the survival rate, the development of therapeutic
agents for tongue SCC is ongoing.
Quercetin (3,3′,4′,5,7-pentahydroxyflavone) is
categorized as a flavonol and is an important polyphenol of the
flavonoid family, which has been detected in numerous fruits and
vegetables, such as apples, cranberries, blueberries and onions
(12–14). Quercetin is also an active
component in several Chinese herbal medicines, such as Flos
Sophorae Immaturus, Hypericum japonicum Thunb. and
Yang-Yin-Qing-Fei-Tang (15,16).
A number of biological effects of quercetin have been reported,
including antiviral, anti-inflammatory and antioxidant effects
(17,18). Furthermore, quercetin has been
reported to possess anticancer potential (19,20).
However, the underlying molecular mechanisms of quercetin in tongue
SCC have remained to be fully elucidated.
Apoptosis is a well-known anticancer mechanism. The
induction of apoptotic pathways to trigger cancer cell death has
been explored in anticancer drug investigations in which damaged
cells were removed through extrinsic or intrinsic pathways
(21,22). MAPK has been indicated to be
involved in cancer cell apoptosis. Furthermore, numerous studies
have reported that the activation of JNK is a key signaling process
for cancer cell apoptosis via various stimulators (3,23–27).
However, to the best of our knowledge, the molecular mechanisms of
quercetin in tongue SCC cell apoptosis remain to be clarified.
Therefore, the present study investigated the pharmacological
effects and possible mechanisms of quercetin on the apoptosis of
tongue SCC-derived SAS cells. The roles of MAPK and GSK3-α/β
signals and the involvement of mitochondrial dysfunction in
quercetin-induced tongue SCC cell apoptosis and death were examined
and clarified.
Materials and methods
Materials
Quercetin and other chemicals (including SP600125,
PD98059 and LiCl), unless specified otherwise, were purchased from
MilliporeSigma. Quercetin (to prepare the stock solution) was
dissolved and diluted in DMSO. To the control wells, the maximum
volume of DMSO used in the experiments was added, which was <1%
per well and did not induce any cytotoxicity. Laboratory
plasticware was obtained from Falcon (Corning Life Sciences). Mouse
and rabbit monoclonal antibodies specific for caspase-3 (cat. no.
9661), caspase-7 (cat. no. 9491), poly(ADP-ribose) polymerase
(PARP) (cat. no. 9542), phosphorylated (p)-JNK (cat. no. 9255),
p-ERK1/2 (cat. no. 4377), p-p38 (cat. no. 9216), p-AMPKα (cat. no.
4188), p-GSK3-α/β (cat. no. 9331), cytochrome c (cat. no.
11940), Bcl-2 (cat. no. 15071), Bax (cat. no. 89477), Bak (cat. no.
12105), JNK-1 (cat. no. 3708), ERK1/2 (cat. no. 9102), p38 (cat.
no. 8690), AMPKα (cat. no. 2532), GSK3-α/β (cat. no. 5676), β-actin
(cat. no. 8457) and secondary antibodies [horseradish peroxidase
(HRP)-conjugated anti-mouse IgG (cat. no. 7076) or anti-rabbit IgG
(cat. no. 7074)] were purchased from Cell Signaling Technology,
Inc.
Cell culture
The human tongue SCC-derived cell line SAS
(JCRB0260) was purchased from Japanese Collection of Research
Bioresources Cell Bank. SAS cells were cultured in a humidified
chamber containing a 5% CO2−95% air mixture at 37°C. All
cells were maintained in culture medium containing 45% Dulbecco's
modified Eagle's medium, 45% Ham's F12 medium and 10% fetal calf
serum (all from Gibco; Thermo Fisher Scientific, Inc.). Cells were
seeded to 6- or 24-well culture plates for each experiment and
allowed to grow for 12–18 h (the recover overnight), and then
treated with quercetin (10–300 µM) for different time intervals in
the absence or presence of the inhibitors of SP600125 (20 µM),
PD98059 (20 µM) or LiCl (100 µM) for 30 min at 37°C prior to
treatment with quercetin.
Morphological analysis
The changes in cell morphology were detected
according to a previous study (28). The cells were cultured on a glass
slide at a density of 1×106 cells/well at 37°C. After 24
h, a photomicrograph was obtained with a 20× objective lens using a
cooled CCD camera attached to a Zeiss Axiovert 135-TV Inverted
Fluorescence Phase Microscope (Zeiss AG).
Cell viability
An MTT assay was used to determine the effect of
quercetin on the viability of SAS cells. The cells were cultured in
a 24-well plate at a density of 2×105 cells/well. After
recovery overnight, the culture medium was removed and the cells
were washed twice with PBS. Fresh medium with quercetin (10–300 µM)
or cisplatin (10 µg/ml; as a positive control) was then added.
After 24 h at 37°C, the cells were washed twice with PBS and fresh
medium was added with 30 µl MTT (2 mg/ml; MilliporeSigma) at 37°C
for 4 h. The medium was then removed and cells were washed twice
with PBS. DMSO was added to dissolve the blue formazan crystals in
cells and the absorbance of cells was detected at a wavelength of
570 nm to determine cell viability using an ELISA reader (model
550; Bio-Rad Laboratories, Inc.).
Annexin V-FITC/propidium iodide (PI)
staining for apoptosis detection
The externalization of phosphatidylserine residues
on the outer plasma membrane of cells is an early event during
apoptosis, which may be detected by annexin V (29). To assess quercetin-induced
apoptosis and necrosis, flow cytometry was performed using the
Annexin V-FITC-PI assay kit (BioVision, Inc.). SAS cells were
seeded at 2×105 cells/well in a 24-well plate and
incubated with quercetin (10–100 µM) at 37°C for 24 h.
Subsequently, the cells were incubated using 0.05% trypsin/EDTA for
1 min to detach the cells, which were then centrifuged (200 × g at
4°C for 5 min), re-suspended in 100 µl binding buffer, transferred
to a 5-ml fluorescence-activated cell sorting tube and combined
with 5 µl Annexin V-FITC and 10 µl PI (50 µg/ml). After incubation
for 30 min at room temperature in the dark, 400 µl binding buffer
was added to each tube and the samples were immediately analyzed
using flow cytometry (FACScalibur; BD Biosciences) using CellQuest
software (version 5.1; BD Biosciences). Four cell populations were
identified as follows: the viable cell population was in the
lower-left quadrant (low FITC and PI signals), the early apoptotic
population was in the lower-right quadrant (high FITC and low PI
signals), the late apoptotic population was in the upper-right
quadrant (high FITC and PI signals), and the necrotic population
was in the upper-left quadrant (low FITC and high PI signals).
Caspase-3 activity assay
Caspase-3 activity was assessed as previously
described by Lee et al (28) and a Caspase-3 Activity Assay Kit
(cat. no. 5723) was used (Cell Signaling Technology, Inc.). The
cells were cultured in a 24-well plate at a density of
2×105 cells/well. After treatment with quercetin (10, 30
and 50 µM) or cisplatin (10 µg/ml; as a positive control) at 37°C
for 24 h, the cells were lysed and the caspase-3/CPP32 substrate
(Ac-DEVD-AMC; 10 µM) was added for 1 h at 37°C. The fluorescence of
cleaved substrates was determined using a spectrofluorometer
(Spectramax; Molecular Devices, LLC) at an excitation wavelength of
380 nm and an emission wavelength of 460 nm.
Determination of mitochondrial
transmembrane potential (MMP)
The alteration of MMP was monitored by flow
cytometry as previously described by Chen et al (30). The cells were harvested and washed
twice with PBS. All samples were stained with DiOC6 (40
nM) for 30 min at 37°C and the fluorescence of DiOC6 was
analyzed by FACSscan flow cytometry (BD Biosciences), using
CellQuest software (version 5.1; BD Biosciences).
Detection of caspase 3/7 activity
Caspase 3/7 is widely accepted as a reliable
indicator of apoptosis (31). A
FLICA DEVD-FMK Caspase 3/7 Assay Kit (cat. no. 94; ImmunoChemistry
Technologies, LLC) was used to determine apoptosis by flow
cytometry. SAS cells were seeded at 2×105 cells/well in
a 24-well plate and incubated with quercetin (50 µM) in the absence
or presence of SP600125 (20 µM), PD98059 (20 µM), or LiCl (100 µM)
at 37°C for 24 h. Subsequently, the cells were collected in 1.5-ml
Eppendorf tubes, centrifuged at 200 × g for 5 min at 4°C, washed
twice with PBS and stained with fluorescent probes for 10 min in a
dark environment at room temperature. Caspase 3/7 activity was
determined based on the fluorescence intensity in cells using flow
cytometry (FACScalibur; BD Biosciences) using CellQuest software
(version 5.1; BD Biosciences).
Western blot analysis
Western blot analysis was performed according to a
previously described protocol (28). In brief, the cells were lysed using
Protein Extraction Solution (cat. no. 17081; iNtRON Biotechnology,
Inc.) and the protein concentration was determined using a
bicinchoninic acid protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). Protein samples (50 µg) were resolved by
SDS-PAGE (a 13.5% gel for caspase-3, −7, cytochrome c,
Bcl-2, Bax, Bak; a 9% gel for p-JNK, JNK-1. P-ERK1/2, ERK1/2,
p-p38, p38, p-AMPKα, AMPKα, p-GSK3-α/β, GSK3-α/β and β-actin) and
transferred to polyvinylidene difluoride membranes. PBS-0.05%
Tween-20 (PBST) containing 5% nonfat dry milk was used to block the
membranes for 1 h at room temperature. The blots were then probed
with primary antibodies (1:1,000 dilution) for 12–16 h at 4°C. The
blots were washed twice with 0.1% PBST and were then probed with
HRP-conjugated secondary antibodies (1:5,000 dilution) for 1 h at
4°C. The antibody-reactive bands were revealed using an
Immobilon® Western Chemiluminescent HRP substrate kit
(MilliporeSigma) and analyzed using a luminescent image analyzer
(ImageQuant™ LAS-4000; GE Healthcare Bio-Sciences). The bands
underwent densitometric analysis using ImageJ version 1.50d
software (National Institutes of Health). For the detection of
cytosolic cytochrome c expression levels, the cells were
detached, washed twice with PBS, and then homogenized with a pestle
and mortar in the extraction buffer [0.4 M mannitol, 25 mM MOPS (pH
7.8), 1 mM EGTA, 8 mM cysteine, and 0.1% (w/v) bovine serum
albumin]. The cell debris was removed via centrifugation at 6,000 ×
g for 2 min. The supernatant was centrifuged at 12,000 × g for 15
min to pellet the mitochondria. The cytochrome c levels in
the supernatants (cytosolic fraction) were detected using western
blot analysis. For the detection of the phosphorylated proteins and
total proteins, the membrane was first probed with the p-protein
antibody and analyzed. Subsequently, the same membrane was washed
and stripped with western blot stripping buffer (cat. no. ab270550;
Abcam) according to the manufacturer's instructions. After
stripping, the membrane was probed with the total protein antibody
and re-analyzed.
Reverse transcription-quantitative PCT
(RT-qPCR) analysis
The mRNA expression levels of Bcl-2, Bax, Bak,
caspase-3, caspase-7, caspase-9 and PARP were analyzed using
RT-qPCR as described in a previous study (32). Cells were cultured in a
10-cm2 dish and incubated with quercetin (50 µM) in the
absence or presence of SP600125 (20 µM), PD98059 (20 µM) or LiCl
(100 µM) at 37°C for different time intervals (2–16 h).
Subsequently, total intracellular RNA was extracted using an RNeasy
kit (Qiagen GmbH) according to the manufacturer's protocol. To
eliminate genomic DNA (gDNA), the sample was added to DNase I.
Subsequently, the sample was incubated at room temperature for 15
min and heated to 70°C for 10 min to denature DNase I, and then
rapidly placed on ice. To reverse transcribe RNA into cDNA, 5 µg
RNA was added to a reaction buffer: 2.5 mM deoxynucleotide mix, 40
U/µl RNAase inhibitor (Promega Corporation), 100 nmol random
hexamer primers, 1X reverse transcriptase (RTase) buffer (which was
supplied with the RTase enzyme) and 30 units AMV RTase enzyme, to
which nuclease-free water was added to reach a final volume of 20
µl. The temperature steps of the RT reaction consisted of 10 min at
room temperature, 15 min at 42°C and 5 min at 95°C and then the
sample was placed on ice. The real-time SYBR Green primers for
human caspase-3, caspase-7, caspase-9, PARP, Bcl-2, Bax, Bak and
β-actin used in the present study are shown in Table I. Each sample (2 µl) was assessed
using Real-Time SYBR Green PCR reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) and transgene-specific primers in a 25-µl
reaction volume, and amplification was performed using an ABI
StepOnePlus™ Sequence Detection System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The cycling conditions consisted of 2 min
at 50°C, 10 min at 95°C, 40 cycles at 95°C for 30 sec and 60°C for
1 min. Real-time fluorescence detection was performed during the
60°C annealing/extension step of each cycle. Melt-curve analysis
was performed on each primer set to ensure that no primer dimers or
nonspecific amplifications were present under the optimized cycling
conditions. After 40 cycles, data analysis was performed using
StepOne™ software (version 2.1; Applied Biosystems; Thermo Fisher
Scientific, Inc.). The fold differences in mRNA expression between
the treatment and control groups were determined using the relative
quantification method, which utilizes qPCR efficiencies and
normalizes them to a housekeeping gene (β-actin was used in the
present study), thus comparing relative Cq changes (ΔCq) between
the control and experimental samples. The fold change values were
calculated using the expression 2−ΔΔCq, where ΔΔC
represents ΔCqcondition of
interest-ΔCqcontrol (33). Prior to conducting the statistical
analyses, the fold change from the mean of the control group was
calculated for each individual sample (including the individual
control samples to assess the variability in this group).
 | Table I.Primer sequences used for reverse
transcription-quantitative PCR analysis. |
Table I.
Primer sequences used for reverse
transcription-quantitative PCR analysis.
| Gene | Sequence | (Refs.) |
|---|
| Human
caspase-3 | F:
5′-GACTCTAGACGGCATCCAGC-3′ | (57) |
|
| R:
5′-TGACAGCCAGTGAGACTTGG-3′ |
|
| Human
caspase-7 | F:
5-AGTGACAGGTATGGGCGTTC-3 | (57) |
|
| R:
5-CGGCATTTGTATGGTCCTCT-3 |
|
| Human
caspase-9 | F:
5-CTGAGCCAGATGCTGTCCCAT-3 | (58) |
|
| R:
5-CTGAGCCAGATGCTGTCCCAT-3 |
|
| Human PARP | F:
5-GCAGAGTATGCCAAGTCCAACAG-3 | (59) |
|
| R:
5-ATCCACCTCATCGCCTTTTC-3 |
|
| Human Bcl-2 | F:
5-TTAGATCTATGGCGCACGCTGGGAGAAC-3 | (60) |
|
| R:
5-CGAATTCTCACTTGTGGCTCAGATAGG-3 |
|
| Human Bax | F:
5-CTTTTGCTTCAGGGTTTCATCC-3 | (61) |
|
| R:
5-TTGAGACACTCGCTCAGCTTCT-3 |
|
| Human Bak | F:
5-ATGGTCACCTTACCTCTGCAA-3 | (62) |
|
| R:
5-TCATAGCGTCGGTTGATGTCG-3 |
|
| Human β-actin | F:
5-GGCGACGAGGCCCAGA-3 | (63) |
|
| R:
5-CGATTTCCCGCTCGGC-3 |
|
Statistical analysis
Data are presented as the mean ± standard deviation
of at least four independent experiments. All data analyses were
performed using SPSS software version 12.0 (SPSS, Inc.).
Significant differences among the groups were assessed by one-way
analysis of variance followed by Tukey's post-hoc test to identify
group differences. P<0.05 was considered to indicate a
statistically significant difference.
Results
Quercetin induces apoptosis in human
tongue SCC SAS cells
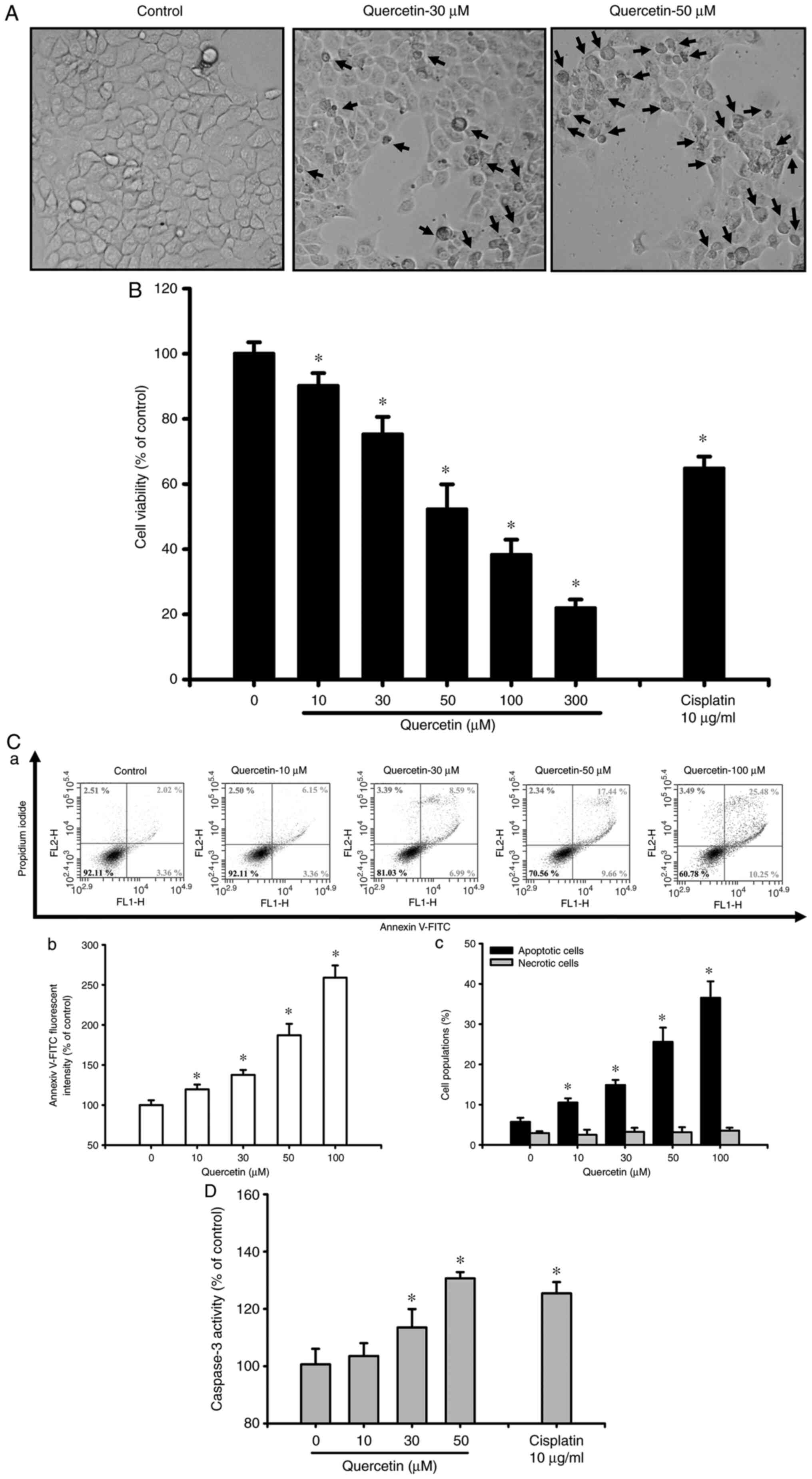
The present study first investigated whether
quercetin had an apoptotic effect on tongue SCC cells. Cells were
treated with quercetin (10–300 µM) for 24 h. As shown in Fig. 1A, quercetin (30 and 50 µM) markedly
induced morphological changes (as indicated by the arrows),
including cell shrinkage, as observed by an inverted phase-contrast
microscope. Furthermore, cell viability was significantly decreased
following treatment with quercetin (10–300 µM; Fig. 1B). The 50% lethal concentration
(LC50) in SAS cells was ~50 µM quercetin (Fig. 1B). As determined by flow cytometry,
the fluorescence intensity of Annexin V-FITC (Fig. 1C-a and b) and the percentage of
apoptotic cells (as indicated by high Annexin V-FITC + low PI plus
high Annexin V-FITC + high PI signals; Fig. 1C-c) were significantly increased in
quercetin-treated cells in a dose-dependent manner. However,
quercetin did not induce cell necrosis in quercetin-treated ASA
cells (Fig. 1C). Furthermore,
caspase-3 activity was also significantly and dose-dependently
increased in cells treated with quercetin (Fig. 1D). Cisplatin (10 µg/ml), as a
positive control, was used to induce cytotoxicity (Fig. 1B) and apoptosis (Fig. 1D) in SAS cells. These results
indicated that quercetin may be capable of inducing SAS cell
apoptosis and death.
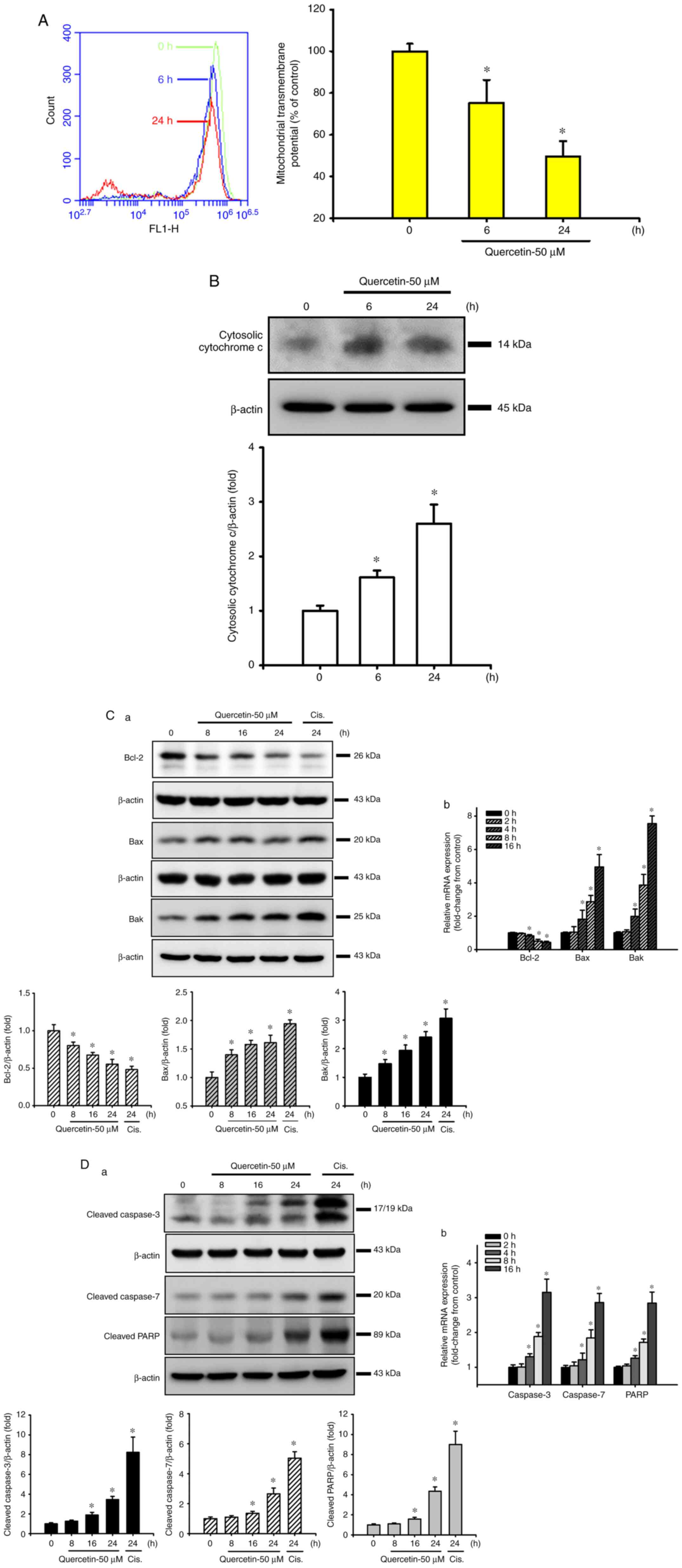
Quercetin induces mitochondrial damage
in human tongue SCC SAS cells
The present study next examined whether the
mitochondrial pathway was involved in quercetin-induced SAS cell
apoptosis. Cells were treated with quercetin (50 µM) for 6 or 24 h.
As shown in Fig. 2A and B, the MMP
was significantly decreased and cytosolic cytochrome c
protein expression was effectively increased in quercetin-treated
cells. Furthermore, the expression levels of mitochondrial
apoptosis-related proteins were also detected. Both Bax and Bak
protein expression levels were time-dependently increased, whereas
Bcl-2 protein expression was time-dependently decreased in response
to quercetin (Fig. 2C-a).
Cisplatin (10 µg/ml), as a positive control, was used to induce
apoptosis-related protein expression in SAS cells (Fig. 2C-a and D-a). In addition, the
protein expression levels of cleaved caspase-3, caspase-7 and PARP
were time-dependently increased in SAS cells following quercetin
exposure (Fig. 2D-a). Moreover,
quercetin decreased Bcl-2, and increased Bax, Bak, caspase-3,
caspase-7 and PARP mRNA expression levels in SAS cells (Fig. 2C-b and D-b). These results
suggested that the mitochondrial pathway may be involved in
quercetin-induced apoptosis in human tongue SCC SAS cell cells.
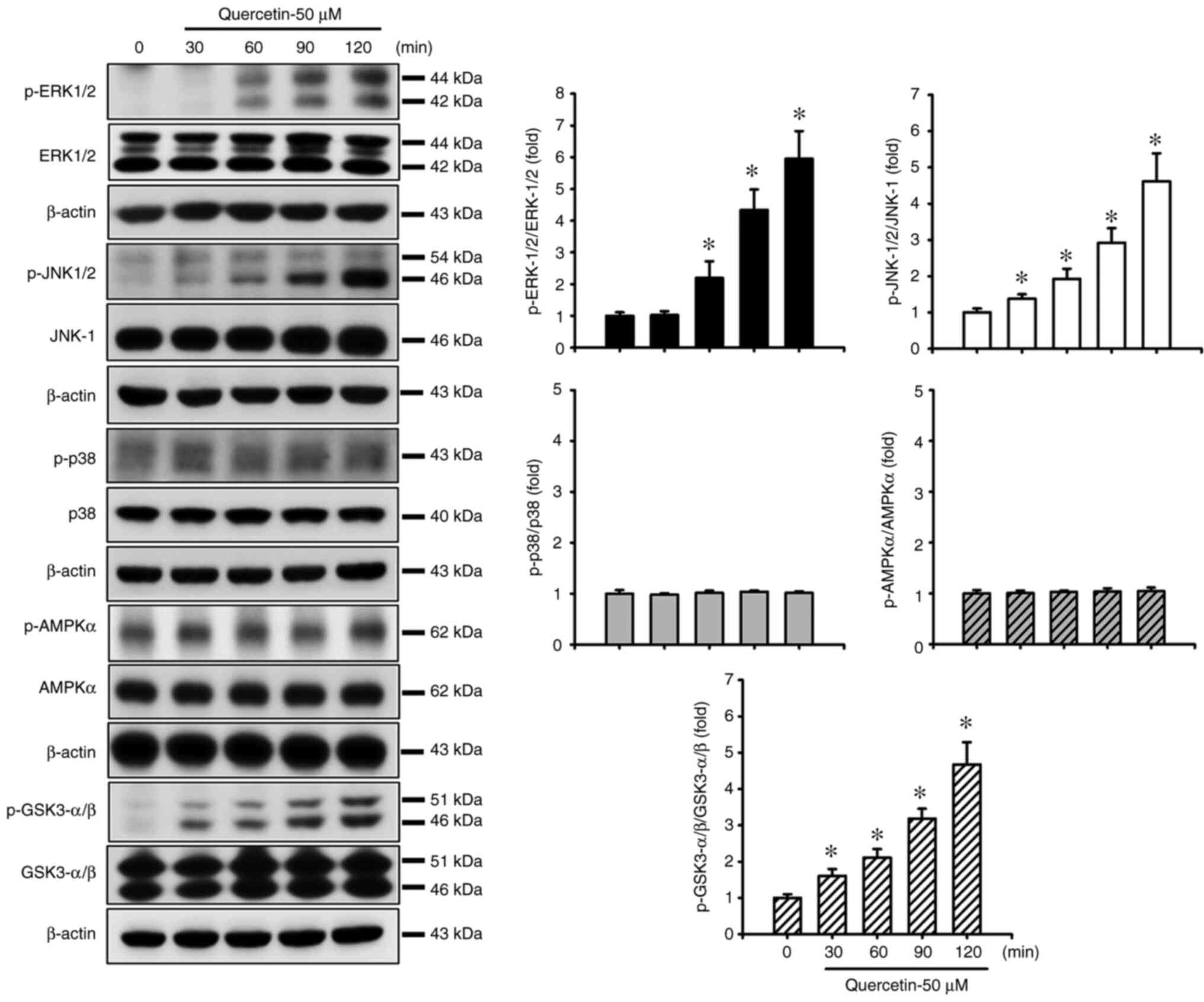
Quercetin induces apoptosis through the JNK, ERK1/2
and GSK3-α/β signaling pathways in human tongue SCC SAS cells. The
present study also investigated the upstream signals for
quercetin-induced mitochondrial apoptosis in SAS cells. Cells were
treated with quercetin (50 µM) for 30–120 min. As shown in Fig. 3, quercetin significantly and
time-dependently increased the protein expression levels of
p-ERK1/2, p-JNK1/2 and p-GSK3-α/β, but not those of p-p38 and
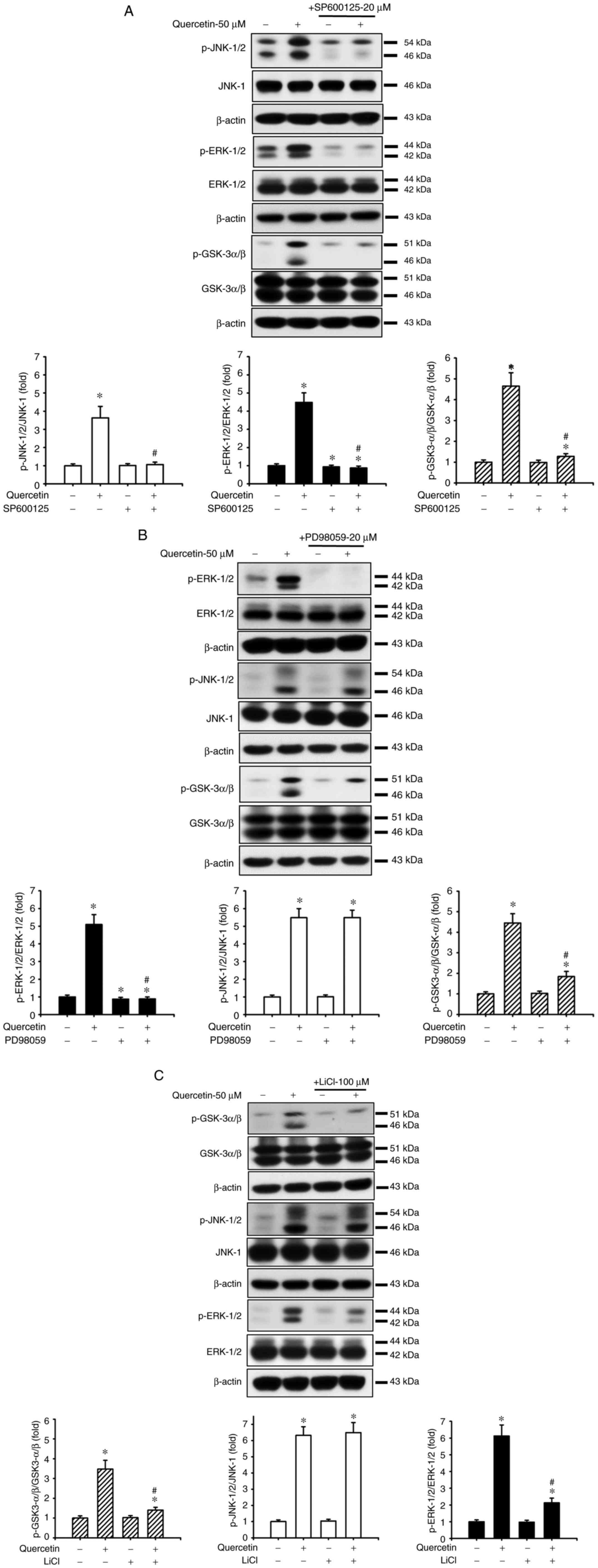
p-AMPKα in SAS cells. Furthermore, the roles of JNK and ERK in
quercetin-induced apoptosis were investigated (Fig. 4). Cells were treated with or
without a JNK inhibitor (SP600125, 20 µM) or an ERK inhibitor
(PD98059, 20 µM) for 30 min, and were then treated with quercetin
(50 µM) for 24 h. As shown in Fig. 4A,
B and D, SP600125 and PD98059 effectively inhibited the
quercetin-induced increase in protein and mRNA expression levels of
caspase-3, caspase-7, caspase-9, PARP and Bax, and reversed the
quercetin-induced decrease in the protein and mRNA expression
levels of Bcl-2 in SAS cells. Furthermore, the role of GSK3 in
quercetin-induced apoptosis was assessed. The cells were treated
with or without the GSK3 inhibitor LiCl (100 µM) for 30 min and
were then treated with quercetin (50 µM) for 16 or 24 h. As shown
in Fig. 4C and D, LiCl effectively
inhibited the increased protein and mRNA expression levels of
caspase-3, caspase-7, caspase-9, PARP and Bax, and decreased the
protein and mRNA expression levels of Bcl-2 in quercetin-treated
cells. These inhibitors (SP600125, PD98059, and LiCl) could also
significantly reverse the decreased MMP (Fig. 4E) and the increased caspase-3/7
activity (Fig. 4F) in
quercetin-treated cells. These results indicated that
quercetin-induced mitochondrial apoptosis may be regulated by
JNK1/2, ERK1/2 and GSK3 signaling pathways in SAS cells.
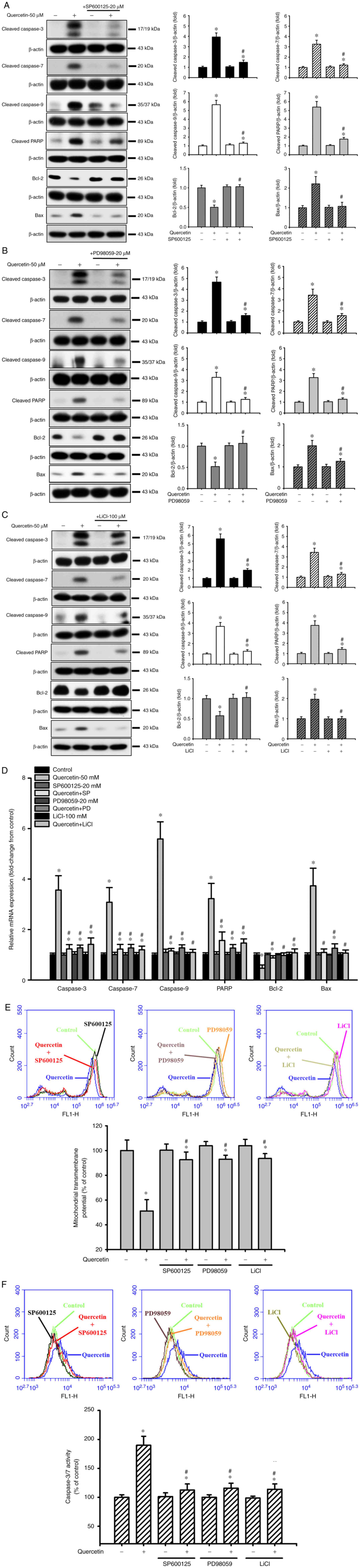 | Figure 4.Effects of selective inhibitors of
JNK, ERK1/2 and GSK3 on mitochondria-dependent apoptotic signals
induced by quercetin in human tongue squamous carcinoma SAS cells.
Cells were treated with or without (A) JNK inhibitor-SP600125 (20
µM), (B) ERK inhibitor-PD98059 (20 µM) or (C) GSK3 inhibitor-LiCl
(100 µM) for 30 min, and were then treated with quercetin (50 µM)
for 24 h. The protein expression levels of cleaved caspase-3,
caspase-7, caspase-9 and PARP, Bcl-2 and Bax were determined by
western blotting. (D) Cells were treated with or without SP600125
(20 µM), PD98059 (20 µM) or LiCl (100 µM) for 30 min, and were then
treated with quercetin (50 µM) for 16 h. The mRNA expression levels
of caspase-3, caspase-7, caspase-9, PARP, Bcl-2 and Bax were
analyzed by reverse transcription-quantitative PCR. (E and F) In
addition, cells were treated with or without SP600125 (20 µM),
PD98059 (20 µM) or LiCl (100 µM) for 30 min and were then treated
with quercetin (50 µM) for 24 h. (E) Mitochondrial transmembrane
potential was determined by flow cytometry. (F) Caspase-3/7
activity was analyzed by measuring fluorescence intensity using
flow cytometry. In A-C, images are representative of at least three
independent experiments. Semi-quantification was determined by
densitometric analysis. Bar graphs are presented as the mean ±
standard deviation of three independent experiments. In D-F, data
are presented as the mean ± standard deviation of four independent
experiments with triplicate determination. *P<0.05 vs. vehicle
control; #P<0.05 vs. quercetin treatment alone. GSK,
glycogen synthase kinase; PARP, poly(ADP-ribose) polymerase. |
The present study examined the relationship between
JNK, ERK and GSK3-α/β signals in SAS cells in the presence or
absence of quercetin. As shown in Fig.
5A, SP600125 effectively reduced the quercetin-increased
protein expression levels of p-JNK1/2, p-ERK1/2 and p-GSK3-α/β.
Moreover, PD98059 treatment effectively reduced the
quercetin-increased protein expression levels of p-ERK/12 and
p-GSK3-α/β but not p-JNK1/2 (Fig.
5B). Similarly, LiCl treatment effectively reduced the
quercetin-increased protein expression levels of p-GSK3-α/β and
p-ERK1/2 but not p-JNK1/2 (Fig.
5C). These results suggested that JNK1/2 activation
downstream-regulated ERK1/2- and GSK3-α/β-mediated apoptotic
signaling pathways may have important roles in quercetin-induced
tongue SCC SAS cell apoptosis.
Discussion
Oral cavity SCC has an increased prevalence in males
and in older individuals, and has had a poor survival rate over the
past two decades (21). The
incidence of tongue SCC is higher than other types of oral SCC.
Among 2,262 cases of oral cavity SCC, in patients aged 20 to 44
years, the incidence of tongue SCC was 39%, whereas the incidence
was 23% in patients ≥45 years of age (11). The main treatment methods of tongue
SCC include radiotherapy, chemotherapy and surgical resection;
however, these clinical therapies can induce adverse effects, such
as mucositis, skin reaction and dysphagia. Moreover, the prognosis
and survival rates for tongue SCC remain poor, and development of
more effective methods is required for prognosis and therapy
(3,21,34–36).
Therefore, the development of therapeutic drugs for improving the
prognosis and survival rates of tongue SCC is required.
Quercetin is a pentahydroxyflavone that occurs in
several natural food products, particularly fruits, vegetables,
black tea and red wine (13,14).
The safe oral dose of quercetin is 1 g/day and absorption is up to
60% (14,37). Furthermore, quercetin has been
determined to be an active component in a number of Chinese herbal
medicines (15,16). Quercetin has been reported to be a
lipophilic compound that can cross the cellular membrane and exert
cytotoxic effects on numerous types of cancer, including ovarian,
lung, breast, nasopharyngeal, kidney, colorectal, prostate and
pancreatic cancer (37–41). A previous study revealed that
quercetin, at a concentration of 300 µM, could induce cytotoxicity
in human lung embryonic fibroblasts, and could induce cell death in
umbilical vein endothelial cells at concentrations between 61 and
303 µM (42). The present study
demonstrated that quercetin, at concentrations of 30 and 50 µM,
markedly induced morphological changes, including cell shrinkage,
in SAS cells. The LC50 of quercetin in SAS cells was ~50
µM; notably, this concentration did not induce cytotoxicity to
human normal cells according to the findings of Matsuo et al
(42).
During apoptosis, cascade protease activities are
induced to cleave apoptosis-related proteins, leading to rapid cell
death (43). It has been reported
that most of the stimuli that induce apoptosis occur through the
mitochondria-dependent pathway, which is affected by the
mitochondrial outer membrane permeabilization (44). Following MMP disruption and
cytochrome c release from the mitochondria to the cytosol,
cytosolic cytochrome c binds to the adaptor molecule
apoptotic protease-activating factor 1 to form the apoptosome and
to activate caspases, including caspase-3, caspase-7 and caspase-9
(44,45). A JNK-regulated
mitochondria-dependent signaling pathway has been reported to
participate in cantharidin-induced oral SCC apoptosis and death
(3). Furthermore, quercetin has
been revealed to induce cytotoxicity and oxidative stress, leading
to mitochondria-related apoptosis in tumor cells (37,46,47).
Notably, quercetin has been shown to induce reactive oxygen species
generation, to decrease the MMP, to increase the expression of
proapoptotic proteins Bad, Bax and Bid, and to decrease the
expression of antiapoptotic proteins Mcl-1, Bcl-2 and Bcl-x, in
gastric cancer cells (48). A
previous study has also shown that quercetin may increase the
cytosolic calcium concentration, protein expression of Bax, release
of apoptosis-inducing factors, and activation of caspase-3,
caspase-8 and caspase-9, and decrease the MMP and protein
expression of Bcl-2 in human breast cancer cells (49). An in vitro study has shown
that quercetin alters the ratio of anti/proapoptotic proteins,
decreases the MMP, and increases the release of cytochrome
c, apoptosis-inducing factor and Endo G from the
mitochondria, leading to apoptosis-triggered cell destruction in
oral SCC SAS cells (50). The
present study also demonstrated that quercetin induced apoptosis
and triggered apoptosis-related signals, including the activation
of caspase-3/7, cleavage of caspase-3/7 and PARP, decrease in MMP,
increase in cytosolic cytochrome c, increase in Bax and Bak
protein and mRNA expression, and decrease in Bcl-2 protein and mRNA
expression in SAS cells. These previous findings and the present
results indicated the pharmacological efficiency of quercetin in
inducing apoptosis of SAS cells. Nevertheless, a further
understanding of the molecular mechanisms underlying the effects of
quercetin on SAS cell apoptosis is required.
MAPKs, including ERK1/2, JNK and p38 kinase, have
been reported to be involved in apoptosis signaling pathways.
Several stimuli, including DNA damage, hydrogen peroxide, cell
surface receptor Fas, UV irradiation, heat, osmotic shock and
chemotherapeutic drugs, can induce apoptosis-related cell death
(51–53). Both JNK and p38 kinase activation
have been shown to induce mitochondria-related apoptosis in HepG2
cells (53). It has also been
reported that ERK1/2 activation is involved in stress-induced
neuronal cell apoptosis (54). In
the present study, quercetin increased the phosphorylation of both
JNK and ERK, but not p38, in SAS cells. Thus, the activation of
both JNK and ERK signals may be involved in the quercetin-induced
apoptosis of SAS cells.
GSK3 is a serine/threonine kinase and has two
isoforms, GSK3-α and GSK3-β (55).
Phorbol esters, the tumor-promoting reagent, have been shown to
inhibit GSK3 signaling through the classical MAPK cascade (56). However, the distinct relationship
between GSK3 and MAPK signals involved in the inhibitory effects of
quercetin on SAS cell survival still requires further
clarification. In the present study, quercetin significantly
increased GSK3-α/β phosphorylation in SAS cells. Pretreatment of
the SAS cells with the specific inhibitors SP600125 (JNK
inhibitor), PD98059 (ERK inhibitor) and LiCl (GSK3 inhibitor)
effectively abrogated the quercetin-induced apoptosis. Moreover, it
was revealed that SP600125 effectively inhibited the
phosphorylation of JNK, ERK and GSK3-α/β, and PD98059 and LiCl
effectively inhibited both ERK and GSK3-α/β phosphorylation, but
not JNK phosphorylation, in quercetin-treated SAS cells. These
results suggested that JNK activation-mediated ERK/GSK3-α/β
signaling may serve a critical role in the downstream regulation of
mitochondria-dependent apoptosis in quercetin-induced tongue SCC
cell death.
In conclusion, the present study demonstrated that
quercetin induced cell death in tongue SCC cells via the JNK
activation-regulated ERK1/2 and GSK3-α/β downstream-mediated
mitochondria-dependent apoptotic signaling pathway. Future in
vivo research is still required to clarify the therapeutic
potential of quercetin in tongue SCC.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the Changhua Christian
Hospital, Changhua, Taiwan (grant no. 110-CCH-IRP-057), the
Taichung Tzu chi Hospital, The Buddhist Tzuchi Medical Foundation
(grant no. TTCRD 109-13), the Far Eastern Memorial Hospital, New
Taipei City, Taiwan (grant no. FEMH-2013-C-028) and the China
Medical University, Taichung, Taiwan (grant no. CMU109-S-64).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
Study conception and design: CFH and CCS.
Implementation of the experiments: CFH, SHL, KMF, JML and CCW.
Acquisition of data and original draft preparation: CFH, SHL, TJH,
KIL and KMF. Analysis and interpretation of data: CFH, KIL, KMF and
WCL. CFH and CCS confirm the authenticity of all the raw data.
Writing, reviewing and editing the manuscript: SHL and CCS. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
SCC
|
squamous cell carcinoma
|
|
MMP
|
mitochondrial transmembrane
potential
|
|
PARP
|
poly(ADP-ribose) polymerase
|
References
|
1
|
Johnson NW, Jayasekara P and Amarasinghe
AA: Squamous cell carcinoma and precursor lesions of the oral
cavity: Epidemiology and aetiology. Periodontol 2000. 57:19–37.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hung CM, Chang CC, Lin CW, Ko SY and Hsu
YC: Cucurbitacin E as inducer of cell death and apoptosis in human
oral squamous cell carcinoma cell line SAS. Int J Mol Sci.
14:17147–17156. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Su CC, Lee KI, Chen MK, Kuo CY, Tang CH
and Liu SH: Cantharidin induced oral squamous cell carcinoma cell
apoptosis via the JNK-regulated mitochondria and endoplasmic
reticulum stress-related signaling pathways. PLoS One.
11:e01680952016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Blot WJ, McLaughlin JK, Winn DM, Austin
DF, Greenberg RS, Preston-Martin S, Bernstein L, Schoenberg JB,
Stemhagen A and Fraumeni JF Jr: Smoking and drinking in relation to
oral and pharyngeal cancer. Cancer Res. 48:3282–3287.
1988.PubMed/NCBI
|
|
5
|
Koo K, Barrowman R, McCullough M, Iseli T
and Wiesenfeld D: Non-smoking non-drinking elderly females: A
clinically distinct subgroup of oral squamous cell carcinoma
patients. Int J Oral Maxillofac Surg. 42:929–933. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong T and Wiesenfeld D: Oral cancer. Aust
Dent J. 63 (Suppl 1):S91–S99. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
World Health Organization, (WHO), . Oral
health. Available from. https://www.who.int/news-room/fact-sheets/detail/oral-healthMarch
25–2020
|
|
8
|
Australian Institute of Health, Welfare
(AIHW), . Australian Cancer Incidence and Mortality (ACIM) books:
Head and neck including lip. Available from. http://www.aihw.gov.au/acim-booksJuly;2017.
|
|
9
|
Yang YH, Chen CH, Chang JS, Lin CC, Cheng
TC and Shieh TY: Incidence rates of oral cancer and oral
pre-cancerous lesions in a 6-year follow-up study of a Taiwanese
aboriginal community. J Oral Pathol Med. 34:596–601. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin NC, Hsien SI, Hsu JT and Chen MY:
Impact on patients with oral squamous cell carcinoma in different
anatomical subsites: A single-center study in Taiwan. Sci Rep.
11:154462021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shiboski CH, Schmidt BL and Jordan RC:
Tongue and tonsil carcinoma: Increasing trends in the U.S.
population ages 20–44 years. Cancer. 103:1843–1849. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Day AJ and Williamson G: Human metabolism
of dietary quercetin glycosides. Plant Polyphenols 2: Chemistry,
Biology, Pharmacology, Ecology. Gross GG, Hemingway RW, Yoshida T.
Basic Life Sciences. Vol 66. Kluwer Academic/Plenum Publishers; New
York, NY: pp. 415–434. 1999, View Article : Google Scholar
|
|
13
|
Harnly JM, Doherty RF, Beecher GR, Holden
JM, Haytowitz DB, Bhagwat S and Gebhardt S: Flavonoid content of US
fruits, vegetables, and nuts. J Agric Food Chem. 54:9966–9977.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Harwood M, Danielewska-Nikiel B,
Borzelleca JF, Flamm GW, Williams GM and Lines TC: A critical
review of the data related to the safety of quercetin and lack of
evidence of in vivo toxicity, including lack of
genotoxic/carcinogenic properties. Food Chem Toxicol. 45:2179–2205.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li H, Tan L, Zhang JW, Chen H, Liang B,
Qiu T, Li QS, Cai M and Zhang QH: Quercetin is the active component
of Yang-Yin-Qing-Fei-Tang to induce apoptosis in non-small cell
lung cancer. Am J Chin Med. 47:879–893. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu H, Chen M, Fan Y, Elsebaei F and Zhu Y:
Determination of rutin and quercetin in Chinese herbal medicine by
ionic liquid-based pressurized liquid extraction-liquid
chromatography-chemiluminescence detection. Talanta. 88:222–229.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee S, Lee HH, Shin YS, Kang H and Cho H:
The anti-HSV-1 effect of quercetin is dependent on the suppression
of TLR-3 in Raw 264.7 cells. Arch Pharm Res. 40:623–630. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu D, Hu MJ, Wang YQ and Cui YL:
Antioxidant activities of quercetin and its complexes for medicinal
application. Molecules. 24:11232019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Khan F, Niaz K, Maqbool F, Ismail Hassan
F, Abdollahi M, Nagulapalli Venkata KC, Nabavi SM and Bishayee A:
Molecular targets underlying the anticancer effects of quercetin:
An update. Nutrients. 8:5292016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Murakami A, Ashida H and Terao J:
Multitargeted cancer prevention by quercetin. Cancer Lett.
269:315–325. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kim JY, An JM, Chung WY, Park KK, Hwang
JK, Kim du S, Seo SR and Seo JT: Xanthorrhizol induces apoptosis
through ROS-mediated MAPK activation in human oral squamous cell
carcinoma cells and inhibits DMBA-induced oral carcinogenesis in
hamsters. Phytother Res. 27:493–498. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lorenzo PI and Saatcioglu F: Inhibition of
apoptosis in prostate cancer cells by androgens is mediated through
downregulation of c-Jun N-terminal kinase activation. Neoplasia.
10:418–428. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim DM, Won M, Chung CS, Kim S, Yim HJ,
Jung SH and Jeong S: JNK-mediated transcriptional upregulation of
RhoB is critical for apoptosis of HCT-116 colon cancer cells by a
novel diarylsulfonylurea derivative. Apoptosis. 15:1540–1548. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wei G, Wang M and Carr BI: Sorafenib
combined vitamin K induces apoptosis in human pancreatic cancer
cell lines through RAF/MEK/ERK and c-Jun NH2-terminal kinase
pathways. J Cell Physiol. 224:112–119. 2010.PubMed/NCBI
|
|
26
|
Gopalan A, Yu W, Sanders BG and Kline K:
Simvastatin inhibition of mevalonate pathway induces apoptosis in
human breast cancer cells via activation of JNK/CHOP/DR5 signaling
pathway. Cancer Lett. 329:9–16. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ryu MJ and Chung HS: [10]-Gingerol induces
mitochondrial apoptosis through activation of MAPK pathway in
HCT116 human colon cancer cells. In Vitro Cell Dev Biol Anim.
51:92–101. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee KI, Lin JW, Su CC, Fang KM, Yang CY,
Kuo CY, Wu CC, Wu CT and Chen YW: Silica nanoparticles induce
caspase-dependent apoptosis through reactive oxygen
species-activated endoplasmic reticulum stress pathway in neuronal
cells. Toxicol In Vitro. 63:1047392020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee SH, Meng XW, Flatten KS, Loegering DA
and Kaufmann SH: Phosphatidylserine exposure during apoptosis
reflects bidirectional trafficking between plasma membrane and
cytoplasm. Cell Death Differ. 20:64–76. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen YW, Yang YT, Hung DZ, Su CC and Chen
KL: Paraquat induces lung alveolar epithelial cell apoptosis via
Nrf-2-regulated mitochondrial dysfunction and ER stress. Arch
Toxicol. 86:1547–1558. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu TH, Su CC, Tang FC, Chen CH, Yen CC,
Fang KM, Lee KI, Hung DZ and Chen YW: Chloroacetic acid triggers
apoptosis in neuronal cells via a reactive oxygen species-induced
endoplasmic reticulum stress signaling pathway. Chem Biol Interact.
225:1–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chung YP, Yen CC, Tang FC, Lee KI, Liu SH,
Wu CC, Hsieh SS, Su CC, Kuo CY and Chen YW: Methylmercury exposure
induces ROS/Akt inactivation-triggered endoplasmic reticulum
stress-regulated neuronal cell apoptosis. Toxicology.
425:1522452019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Simon C and Plinkert PK: Combined modality
approaches in the treatment of head and neck cancer patients. HNO.
56:575–584. 2008.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu JY, Yi C, Chung HR, Wang DJ, Chang WC,
Lee SY, Lin CT, Yang YC and Yang WC: Potential biomarkers in saliva
for oral squamous cell carcinoma. Oral Oncol. 46:226–231. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Pérez-Sayáns M, Suárez-Peñaranda JM,
Gayoso-Diz P, Barros-Angueira F, Gándara-Rey JM and García-García
A: Tissue inhibitor of metalloproteinases in oral squamous cell
carcinomas-a therapeutic target? Cancer Lett. 323:11–19. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shafabakhsh R and Asemi Z: Quercetin: A
natural compound for ovarian cancer treatment. J Ovarian Res.
12:552019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Choi JA, Kim JY, Lee JY, Kang CM, Kwon HJ,
Yoo YD, Kim TW, Lee YS and Lee SJ: Induction of cell cycle arrest
and apoptosis in human breast cancer cells by quercetin. Int J
Oncol. 19:837–844. 2001.PubMed/NCBI
|
|
39
|
Kuo PC, Liu HF and Chao JI: Survivin and
p53 modulate quercetin-induced cell growth inhibition and apoptosis
in human lung carcinoma cells. J Biol Chem. 279:55875–55885. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ong CS, Tran E, Nguyen TT, Ong CK, Lee SK,
Lee JJ, Ng CP, Leong C and Huynh H: Quercetin-induced growth
inhibition and cell death in nasopharyngeal carcinoma cells are
associated with increase in Bad and hypophosphorylated
retinoblastoma expressions. Oncol Rep. 11:727–733. 2004.PubMed/NCBI
|
|
41
|
Sharmila G, Bhat FA, Arunkumar R, Elumalai
P, Raja Singh P, Senthilkumar K and Arunakaran J: Chemopreventive
effect of quercetin, a natural dietary flavonoid on prostate cancer
in in vivo model. Clin Nutr. 33:718–726. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Matsuo M, Sasaki N, Saga K and Kaneko T:
Cytotoxicity of flavonoids toward cultured normal human cells. Biol
Pharm Bull. 28:253–259. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Taylor RC, Cullen SP and Martin SJ:
Apoptosis: Controlled demolition at the cellular level. Nat Rev Mol
Cell Biol. 9:231–241. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lopez J and Tait SW: Mitochondrial
apoptosis: Killing cancer using the enemy within. Br J Cancer.
112:957–962. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Tait SW, Parsons MJ, Llambi F,
Bouchier-Hayes L, Connell S, Muñoz-Pinedo C and Green DR:
Resistance to caspase-independent cell death requires persistence
of intact mitochondria. Dev Cell. 18:802–813. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Metodiewa D, Jaiswal AK, Cenas N,
Dickancaité E and Segura-Aguilar J: Quercetin may act as a
cytotoxic prooxidant after its metabolic activation to semiquinone
and quinoidal product. Free Radic Biol Med. 26:107–116. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Awad HM, Boersma MG, Vervoort J and
Rietjens IM: Peroxidase-catalyzed formation of quercetin quinone
methide-glutathione adducts. Arch Biochem Biophys. 378:224–233.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Shang HS, Lu HF, Lee CH, Chiang HS, Chu
YL, Chen A, Lin YF and Chung JG: Quercetin induced cell apoptosis
and altered gene expression in AGS human gastric cancer cells.
Environ Toxicol. 33:1168–1181. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Chien SY, Wu YC, Chung JG, Yang JS, Lu HF,
Tsou MF, Wood WG, Kuo SJ and Chen DR: Quercetin-induced apoptosis
acts through mitochondrial- and caspase-3-dependent pathways in
human breast cancer MDA-MB-231 cells. Hum. Exp Toxicol. 28:493–503.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Ma YS, Yao CN, Liu HC, Yu FS, Lin JJ, Lu
KW, Liao CL, Chueh FS and Chung JG: Quercetin induced apoptosis of
human oral cancer SAS cells through mitochondria and endoplasmic
reticulum mediated signaling pathways. Oncol Lett. 15:9663–9672.
2018.PubMed/NCBI
|
|
51
|
Ichijo H: From receptors to
stress-activated MAP kinases. Oncogene. 18:6087–6093. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mansouri A, Ridgway LD, Korapati AL, Zhang
Q, Tian L, Wang Y, Siddik ZH, Mills GB and Claret FX: Sustained
activation of JNK/p38 MAPK pathways in response to cisplatin leads
to Fas ligand induction and cell death in ovarian carcinoma cells.
J Biol Chem. 278:19245–19256. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yuan L, Wang J, Xiao H, Wu W, Wang Y and
Liu X: MAPK signaling pathways regulate mitochondrial-mediated
apoptosis induced by isoorientin in human hepatoblastoma cancer
cells. Food Chem Toxicol. 53:62–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Song H, Kim W, Choi JH, Kim SH, Lee D,
Park CH, Kim S, Kim DY and Kim KT: Stress-induced nuclear
translocation of CDK5 suppresses neuronal death by downregulating
ERK activation via VRK3 phosphorylation. Sci Rep. 6:286342016.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mancinelli R, Carpino G, Petrungaro S,
Mammola CL, Tomaipitinca L, Filippini A, Facchiano A, Ziparo E and
Giampietri C: Multifaceted roles of GSK-3 in cancer and
autophagy-related diseases. Oxid Med Cell Longev. 2017:46294952017.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Frame S and Cohen P: GSK3 takes centre
stage more than 20 years after its discovery. Biochem J. 359:1–16.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Oh BY, Kim KH, Chung SS and Lee RA:
Silencing the livin gene enhances the cytotoxic effects of
anticancer drugs on colon cancer cells. Ann Surg Treat Res.
91:273–277. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yu JH, Zheng GB, Liu CY, Zhang LY, Gao HM,
Zhang YH, Dai CY, Huang L, Meng XY, Zhang WY and Yu XF: Dracorhodin
perchlorate induced human breast cancer MCF-7 apoptosis through
mitochondrial pathways. Int J Med Sci. 10:1149–1156. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Uchiumi F, Watanabe T, Ohta R, Abe H and
Tanuma S: PARP1 gene expression is downregulated by knockdown of
PARG gene. Oncol Rep. 29:1683–1688. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Nencioni L, De Chiara G, Sgarbanti R,
Amatore D, Aquilano K, Marcocci ME, Serafino A, Torcia M, Cozzolino
F, Ciriolo MR, et al: Bcl-2 expression and p38MAPK activity in
cells infected with influenza A virus: impact on virally induced
apoptosis and viral replication. J Biol Chem. 284:16004–16015.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kotsafti A, Farinati F, Cardin R, Cillo U,
Nitti D and Bortolami M: Autophagy and apoptosis-related genes in
chronic liver disease and hepatocellular carcinoma. BMC
Gastroenterol. 12:1182012. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hasan Z, Ashraf M, Tayyebi A and Hussain
R: M. leprae inhibits apoptosis in THP-1 cells by downregulation of
Bad and Bak and upregulation of Mcl-1 gene expression. BMC
Microbiol. 6:782006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Goidin D, Mamessier A, Staquet MJ, Schmitt
D and Berthier-Vergnes O: Ribosomal 18S RNA prevails over
glyceraldehyde-3-phosphate dehydrogenase and beta-actin genes as
internal standard for quantitative comparison of mRNA levels in
invasive and noninvasive human melanoma cell subpopulations. Anal
Biochem. 295:17–21. 2001. View Article : Google Scholar : PubMed/NCBI
|