Introduction
Breast cancer (BC) has now surpassed lung cancer as
the most common cancer worldwide, with ~2.3 million new cases in
2020 (1), which makes the disease a
major public health burden (2).
Despite recent advances in medical technology, the molecular basis
of breast cancer progression remains unclear. Clarifying the
mechanisms of breast cancer biology can help identify new
therapeutic targets, thereby providing a basis for the development
of more effective treatments.
The incidence of postmenopausal breast cancer is
increasing with the aging world population (3). Follicle-stimulating hormone (FSH)
levels remain high after menopause owing to the loss of estrogen
negative feedback. However, the relationship between FSH levels and
breast cancer onset and progression remains unclear. FSH is a
glycoprotein hormone related to luteinizing hormone (LH),
thyroid-stimulating hormone (TSH) and human chorionic gonadotropin
(hCG). These hormones are secreted by the pituitary gland and
consist of a heterodimer of two noncovalently linked α- and
β-subunits. The former is identical in all these hormones (4) and is encoded by a single gene, the
glycoprotein hormone α-subunit (CGA), whereas β-subunits in
each hormone are encoded by a different gene (5). CGA has been implicated in the
occurrence and progression of a number of solid tumors, including
lung (6), prostate (7) and gastric (8) cancer. In addition, CGA is a candidate
marker for predicting the response to tamoxifen treatment in breast
cancer (9,10). However, the precise role and
mechanisms of action of CGA in breast cancer are unclear.
The main function of the gonadotropin-releasing
hormone (GnRH) is to control the release of FSH and LH. GnRHa is a
synthetic GnRH derivative that inhibits the synthesis and release
of LH and FSH, and reduces CGA levels in the body by continuous
administration. There is still a debate about the clinical use of
GnRHa in premenopausal women with breast cancer. Those who
supported the use of GnRHa believed that it protected patient
ovarian function during chemotherapy (11), objections were made over the use of
GnRHa, as no apparent benefit and ovarian protection were observed
in another study (12). Whether the
benefits of GnRHa treatment are related to the decrease in CGA
levels in the body remains to be understood. Therefore, it is
necessary to study the mechanism of action of CGA in breast cancer,
which has practical significance for the clinical treatment of this
disease.
The present study aimed to investigate the role and
mechanism of action of CGA in breast cancer by comparing its
expression between breast cancer and normal breast tissues, and the
overexpression and deletion of CGA in different cancer cell
lines.
Materials and methods
Tissue microarrays and
immunohistochemistry
Tissue microarrays (TMAs) containing 160 breast
cancer tissues were obtained from Shanghai Outdo Biotech, Co., Ltd.
Immunohistochemistry (IHC) was performed on the TMA sections. The
sections were first deparaffinized with xylene, rehydrated using a
graded series of ethanol, and washed with water. Antigen retrieval
was performed by heating the sections in citrate buffer for 20 min,
followed by cooling to room temperature. The sections were
incubated in 3% hydrogen peroxide for 10 min at room temperature to
quench endogenous peroxidase. Subsequently, a sodium citrate buffer
solution (0.01 M; pH 6.0) was added to incubate the sections for 3
min under boiling conditions. Bovine serum albumin (5%) (Beijing
Solarbio Science & Technology Co., Ltd.) was used to block the
sections for 30 min at 37°C. The sections were incubated overnight
at 4°C with a mouse anti-human monoclonal antibody against CGA
(1:500; ab11232; Abcam), followed by incubation with a HRP-polymer
conjugated anti-mouse IgG secondary antibody (Fuzhou Maixin Biotech
Co., Ltd.) for 30 min at room temperature. After washing in
phosphate-buffered saline (PBS), the sections were stained with
diaminobenzidine at room temperature for 2 min and counterstained
with hematoxylin at room temperature for 10 sec. Immunoreactivity
was scored by experienced pathologists (who were blinded to patient
identity) according to the immunoreactivity scoring system (IRS),
which used the percentage of immunopositive cells (0%, 0; 1–10%, 1;
11–50%, 2; 51–80%, 3; and 81–100%, 4) and staining intensity
(negative, 0; weak, 1; moderate, 2; and strong, 3). IRS score
ranging from 0 to 12 was calculated by multiplication of the two
scores. Patient consent was obtained and the use of TMAs containing
human tissues was approved by the Ethics Committee of Shanghai
Outdo Biotech, Co., Ltd. (approval no. SHYJS-CP-1804004). The study
was approved by the Ethics Committee of the Women's Hospital,
Zhejiang University School of Medicine (Hangzhou, China) and was in
accordance with the Declaration of Helsinki.
Cell lines and cell culture maintenance. SK-BR-3,
MDA-MB-231, MCF-7 and T-47D human breast cancer cell lines and
MCF-10A normal mammary epithelial cells were obtained from the Cell
Bank of the Shanghai Institute of Cell Biology. SK-BR-3 and
MDA-MB-231 cells were cultured in McCoy's 5a, RPMI-1640 and L-15
media supplemented with 10% fetal bovine serum (FBS) (all Gibco;
Thermo Fisher Scientific, Inc.) and 1% antibiotics
(penicillin-streptomycin, ST488; Shanghai Biyuntian Biotechnology
Co., Ltd.). MCF-7 and T-47D cells were maintained in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.) containing 10% FBS and 1% antibiotics. MCF-10A cells were
grown in DMEM/F12 containing 5% horse serum (Thermo Fisher
Scientific, Inc.), 0.5 µg/ml hydrocortisone, 100 ng/ml cholera
toxin, 20 ng/ml EGF, 10 µg/ml insulin and 1% antibiotics. All cells
were grown in a humidified atmosphere of 5% CO2 at
37°C.
RNA isolation and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from cultured cells
(MCF-10A, MCF-7, T-47D, MDA-MB-231 and SK-BR-3) using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and reverse-transcribed into cDNA using the PrimeScript
RT-PCR kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. qPCR was performed using SYBR Premix Ex
Taq (Takara Biotechnology Co., Ltd.) according to the
manufacturer's instructions on a StepOnePlus Real-Time PCR System
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
thermocycling conditions were as follows: 95°C for 10 min, followed
by 38 cycles of 95°C for 15 sec and 60°C for 40 sec. The results
are expressed as the fold-change in relative mRNA expression
levels, calculated using the 2−ΔΔCq method (13). Each reaction was performed in
triplicate. The following primers were used: CGA forward,
5′-TGCCCAGGCTGCTCTCAAACT-3′ and reverse,
5′-GCAAGTGGACTCTGAGGTGACG-3′; and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH; internal control) forward,
5′-GGTATCGTGGAAGGACTC-3′ and reverse,
5′-GGGATGATGTTCTGGAGAG-3′.
Protein extraction and western blot
analysis
Breast cells (MCF-10A, MCF-7, T-47D, MDA-MB-231 and
SK-BR-3) were washed twice with ice-cold PBS and lysed in a lysis
buffer (cat. no. 87787; Pierce; Thermo Fisher Scientific, Inc.)
containing a protease inhibitor cocktail (1:100 dilution;
MilliporeSigma) for 30 min at 4°C. Protein concentrations were
determined by the bicinchoninic acid assay (cat. no. 23227; Thermo
Fisher Scientific, Inc.). Protein samples (20–40 µg) were separated
on a 10% gel using sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred to a polyvinylidene difluoride
membrane. After being blocked with 5% skimmed milk for 2 h at room
temperature, the membranes were incubated overnight at 4°C with the
following primary antibodies: GAPDH antibody (1:2,500; cat. no.
ab9485; Abcam), CGA antibody (1:1,000; cat. no. ab11232; Abcam),
His-tag antibody (1:1,000; cat. no. 9991s; Cell Signaling
Technology, Inc.), Phospho-EGF Receptor Pathway Antibody Sampler
Kit (1:1,000; cat. no. 9789; Cell Signaling Technology, Inc.),
Erk1/2 antibody (1:1,000; cat. no. 4695; Cell Signaling Technology,
Inc.), Akt antibody (1:1,000; cat. no. 4685; Cell Signaling
Technology, Inc.) and EGFR antibody (1:1,000; cat. no. 4267; Cell
Signaling Technology, Inc.). Next, the membranes were incubated
with goat anti-mouse secondary antibodies (1:2,000; cat. no. A0216;
Shanghai Biyuntian Biotechnology Co., Ltd.) for 1 h at room
temperature. The protein bands were visualized with ECL detection
reagents (cat. no. P0018FM; Shanghai Biyuntian Biotechnology Co.,
Ltd.) and analyzed with ImageJ software (version 1.47; National
Institute of Health).
Small interfering (si)RNA and plasmid
transfection
Using standard plasmid construction procedures, CGA
cDNA (Shenggong Biotechnology) was cloned into the pcDNA3.1-His
plasmid. CGA siRNAs and control siRNAs were designed by Genepharma
Co., Ltd. For siRNA transfection, MDA-MB-231 cells were seeded at
70–80% confluence in 6-well plates 1 day before transfection.
Transfections of siRNA (at 100 pmol) were performed using the
Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at room temperature for 20 min, according to the
manufacturer's protocols. Controls included nonspecific siRNA
(negative control) and GAPDH siRNA (positive control). Similarly,
pcDNA3.1-His-CGA was transfected into T-47D cells according to the
standard plasmid transfection procedure, with the transfection of
empty vector as the negative control. Breast cancer cell lines
without any treatment were used as normal controls. The cells were
evaluated for CGA expression and biological effects 72 h after
transfection. The following forward and reverse primers were used
to generate the siRNAs for the knockdown experiment: GAPDH
(positive control), 5′-UGACCUCAACUACAUGGUUTT-3′ and
5′-AACCAUGUAGUUGAGGUCATT-3′; negative control scrambled siRNA,
5′-UUCUCCGAACGUGUCACGUTT-3′ and 5′-ACGUGACACGUUCGGAGAATT-3′;
CGA-554 (no. 1), 5′-CUGCAGUACUUGUUAUUAUTT-3′ and
5′-AUAAUAACAAGUACUGCAGTT-3′; CGA-490 (no. 2),
5′-CUAAAUCAUAUAACAGGGUTT-3′ and 5′-ACCCUGUUAUAUGAUUUAGTT-3′; and
CGA-404 (no. 3), 5′-CUCUAGAGCAUAUCCCACUTT-3′ and
5′-AGUGGGAUAUGCUCUAGAGTT-3′.
Cell proliferation assay
Breast cancer cells were cultured in 96-well plates
seeded at a concentration of 5×103 cells/ml for 72, 120
and 168 h. When the cells were in the exponential growth phase, 20
µl of 5 mg/ml stock solution of
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide
(Sigma-Aldrich; Merck KGaA) was added to each well and incubated
for 4 h at 37°C. The medium was replaced with 150 µl dimethyl
sulfoxide and the plate was incubated with gentle agitation for 10
min. The absorbance was measured at 490 nm using a microplate
reader.
In vivo xenograft tumor model
A total of 20 female BALB/c nude mice (4–5 weeks
old; weight, 16–20 g) were obtained from Shanghai SLAC Laboratory
Animal Co., Ltd. The mice were housed in Makrolon cages (5 per
cage) in an airflow cabinet under pathogen-free conditions at 23°C
on a 12/12 h day/night cycle, with water and food ad
libitum. Animal health was regularly monitored by professional
laboratory managers, including dietary intake, respiration, body
weight, activity and tumor burden. Different groups of breast
cancer cells (T-47D, T-47D/CGA+, MDA-MB-231 and
MDA-MB-231/CGA−) in the exponential growth phase were
collected, washed and resuspended in a sterile DMEM. The nude mice
were subcutaneously inoculated with 5×106 cells into the
dorsal side of the right forelimb. Five mice were used per
experiment and sample. After the tumor was formed, its maximum
diameter and minimum diameter were measured regularly until the end
of the experiment. Tumor volume was calculated using the formula: L
× S2 × 0.5, where L and S represent the maximum and
minimum diameter of the tumor, respectively. The criterion for
stopping the experiment was whether the largest tumor in each group
had reached 1.5 cm in diameter or the tumor had ruptured. The mice
were sacrificed by cervical dislocation under anesthesia.
Anesthesia was performed by inhalation of isoflurane at a dose
range of 2–6% for induction and 1–3% for maintenance. Next, tumors
were dissected and weighed, and images were captured. All animal
experiments were performed in compliance with the guidelines of the
Institute of Experimental Animal Sciences (Zhejiang University
Laboratory Animal Center, Hangzhou, China). The protocol was
approved by the Animal Care and Use Committee of Zhejiang
University (approval no. ZJU2015-339-01).
Bioinformatics analysis
CGA expression and associated proteins were analyzed
using published databases. First, the cbioportal platform
(http://www.cbioportal.org), an open data
analysis platform was used, entering the query genes in the webpage
according to the requirements, selecting different tumor databases
and obtaining the CGA expression levels in various tumors. Second,
CGA expression and survival data was obtained for patients with CGA
upregulation in different breast cancer databases. By using
GENEMANIA (https://genemania.org), the interaction
of the analyzed proteins was predicted.
Statistical analysis
GraphPad Prism version 6.0 (GraphPad Software, Inc.)
was used to perform all statistical analyses. Differences between
groups were evaluated using either a two-tailed unpaired Student's
t-test or a one-way analysis of variance and Tukey's post hoc test.
The association between CGA expression and patient prognosis was
determined using the Kaplan-Meier method and Mantel-Cox test. A
statistically significant difference was indicated by P<0.05.
Each experiment was performed independently three times.
Results
High expression level of CGA in breast
cancer is associated with a poor prognosis
CGA expression was examined in TMAs obtained from
160 patients with ductal carcinoma using IHC. The baseline
characteristics of the patients are presented in Table I. Some cores in the TMAs were lost
during sample processing; therefore, 141 of the 160 breast cancer
specimens were available for the IHC analysis of CGA expression.
Representative images of CGA expression in breast cancer tissues
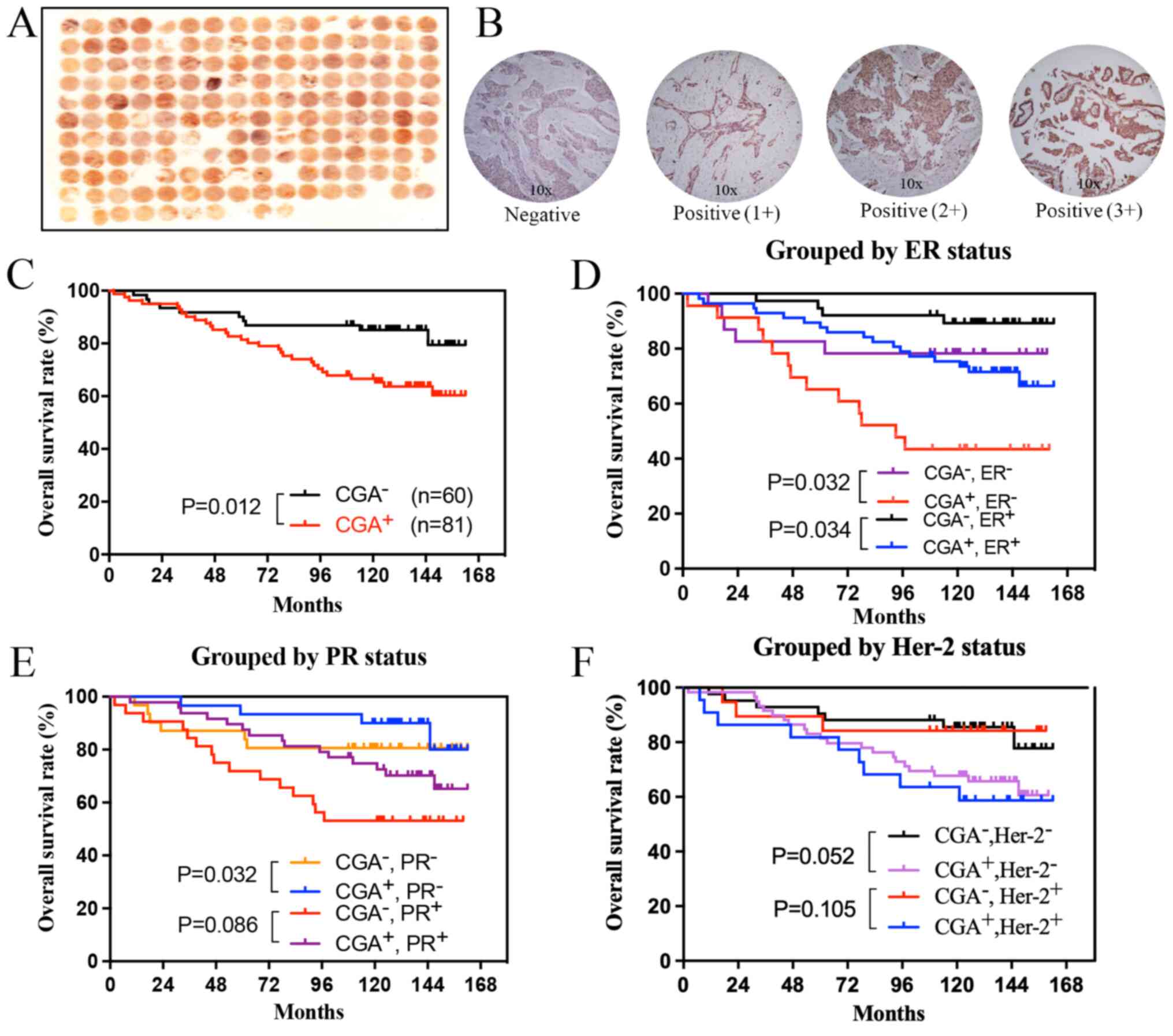
and staining intensity scores are shown in Fig. 1A and B. CGA expression was mainly
cytoplasmic in the tumor cells and negative in the normal ductal
epithelial tissue.
 | Table I.Characteristics of ductal carcinoma
specimens in the tissue microarray analyzed using
immunohistochemistry (n=141). |
Table I.
Characteristics of ductal carcinoma
specimens in the tissue microarray analyzed using
immunohistochemistry (n=141).
| Characteristic | Value |
|---|
| Mean age ± SD
(range), years | 53.9±13.3
(29–83) |
| Tumor size in cm, n
(%) |
|
| ≤2 | 21 (14.9) |
|
>2 | 120 (85.1) |
| Tumor
stagea, n (%) |
|
| 1 | 12 (8.5) |
| 2 | 83 (58.9) |
| 3 | 46 (32.6) |
| 4 | 0 (0.0) |
| Histological grade,
n (%) |
|
| 1 | 41 (29.1) |
| 2 | 60 (42.6) |
| 3 | 40 (28.4) |
| Lymph node
metastasis, n (%) |
|
|
Positive | 86 (61.0) |
|
Negative | 55 (39.0) |
Patients with breast cancer who were positive for
CGA expression had a shorter overall survival time compared with
those who were negative for CGA expression (Fig. 1C). Subgroup analysis according to
estrogen receptor (ER) status confirmed that CGA expression was
associated with a shorter overall survival time (Fig. 1D). In the progesterone receptor
(PR)-negative group, the CGA-positive patients had longer survival
times (Fig. 1E). No differences in
survival based on CGA levels were found in the human epidermal
growth factor receptor (Her)-2 subgroup (Fig. 1F).
CGA is highly expressed in breast
cancer cell lines
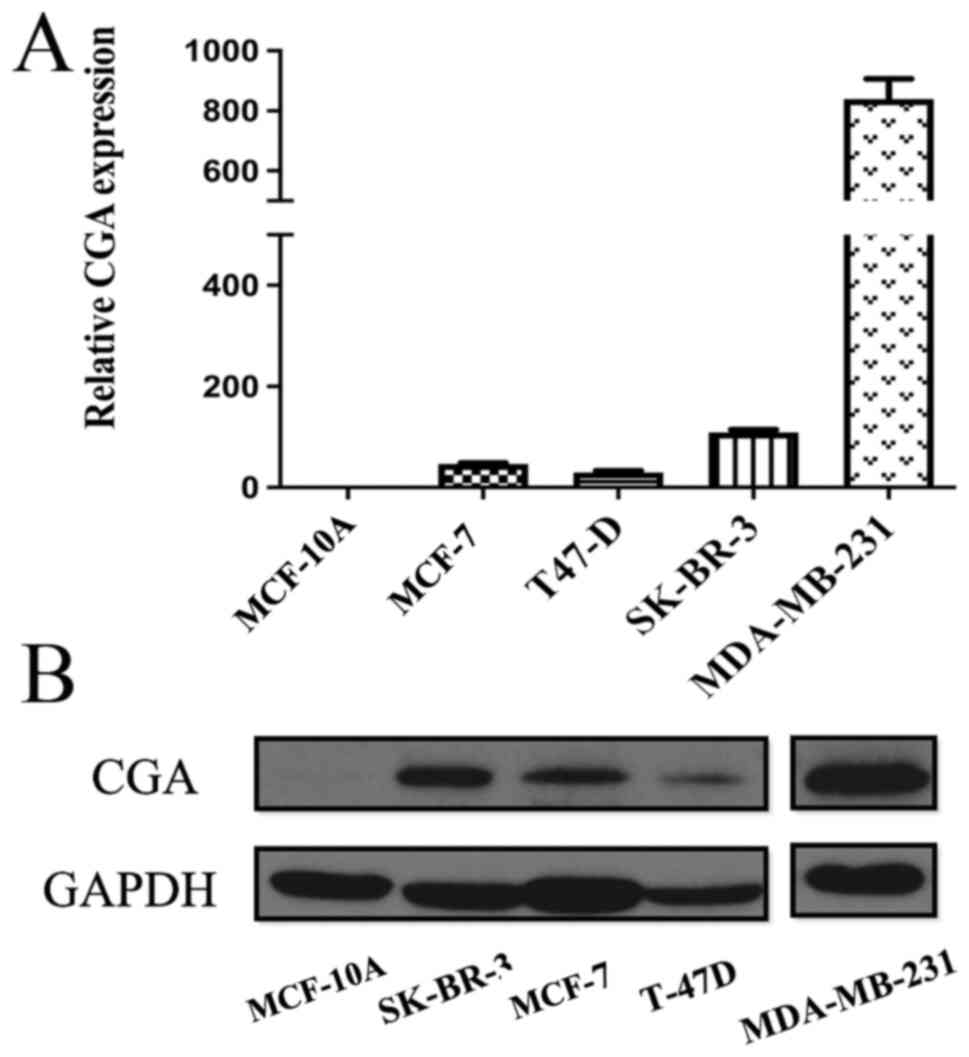
CGA mRNA and protein expression levels were compared
between a normal mammary epithelial cell line and breast cancer
cell lines, and it was found that MCF-10A cells expressed very low
levels of CGA transcripts and CGA protein (Fig. 2A and B). By contrast, breast cancer
cell lines showed variable expression of CGA: MDA-MB-231, a
triple-negative (ER−/PR−/Her-2−)
cell line, had the highest levels of CGA, whereas T-47D
(ER+/PR+/Her-2−), a moderately
malignant cell line, showed relatively low CGA expression.
CGA overexpression enhances breast
cancer cell proliferation
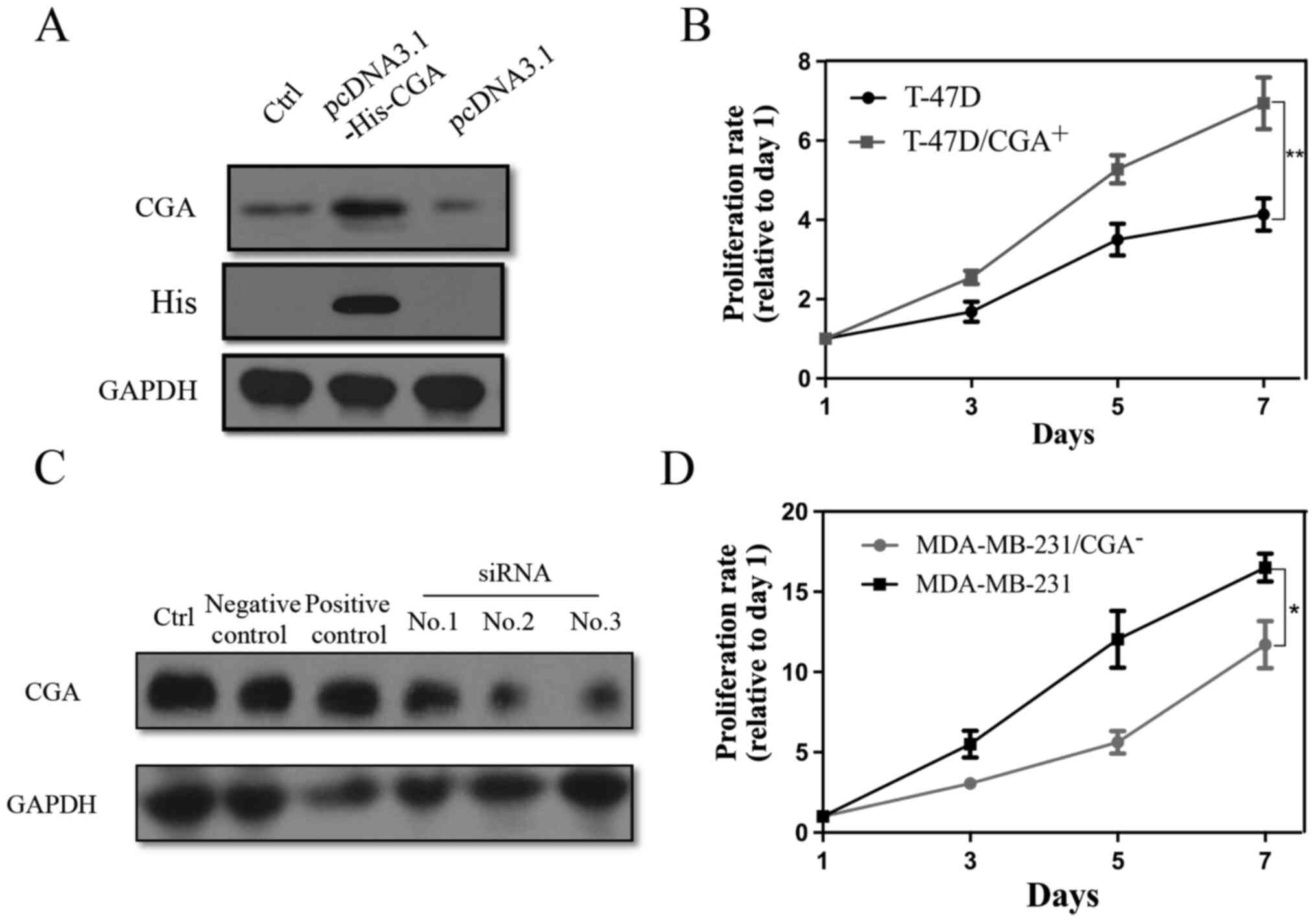
To investigate the role of CGA in breast cancer, a
plasmid containing CGA coding sequence (Gene ID, 1081) fused with a
His tag was first constructed for CGA overexpression
(T-47D/CGA+), and knockdown MDA-MB-231/CGA−
cell lines with siRNA were established. Successful transfection of
pcDNA3.1-His-CGA into T-47D cells was confirmed (Fig. 3A). T-47D/CGA+ cells showed
a higher rate of cell proliferation than the control cells
(Fig. 3B). Of the three siRNAs
tested, no. 2 showed the highest knockdown efficiency (Fig. 3C). MDA-MB-231/CGA− cells
showed decreased proliferative capacity compared with control cells
(Fig. 3D).
CGA increases EGFR, ERK1/2 and Akt
phosphorylation
Signaling pathways such as those of EFGR,
mitogen-activated protein kinase (MAPK) and Akt play important
roles in the proliferation of breast cancer cells and are mainly
activated by phosphorylation (14).
To clarify the mechanism by which CGA stimulates breast cancer cell
proliferation, the total and phosphorylated protein levels of EGFR,
ERK1/2 and Akt in CGA-overexpressing and CGA-depleted cells were
evaluated. The phosphorylation of EGFR (Y1173), ERK1/2 and Akt
(Y473) was increased in T-47D/CGA+ cells, whereas total
protein levels were largely unchanged compared with those in
control cells (Fig. 4). Conversely,
MDA-MB-231/CGA− cells exhibited decreased levels of
phosphorylated (p-)EGFR (Y1173), p-ERK1/2 and p-Akt (Y473). These
results suggest that CGA enhances breast cancer cell proliferation
via the activation of EGFR, ERK1/2 and Akt signaling cascades.
CGA induces breast cancer growth in
vivo
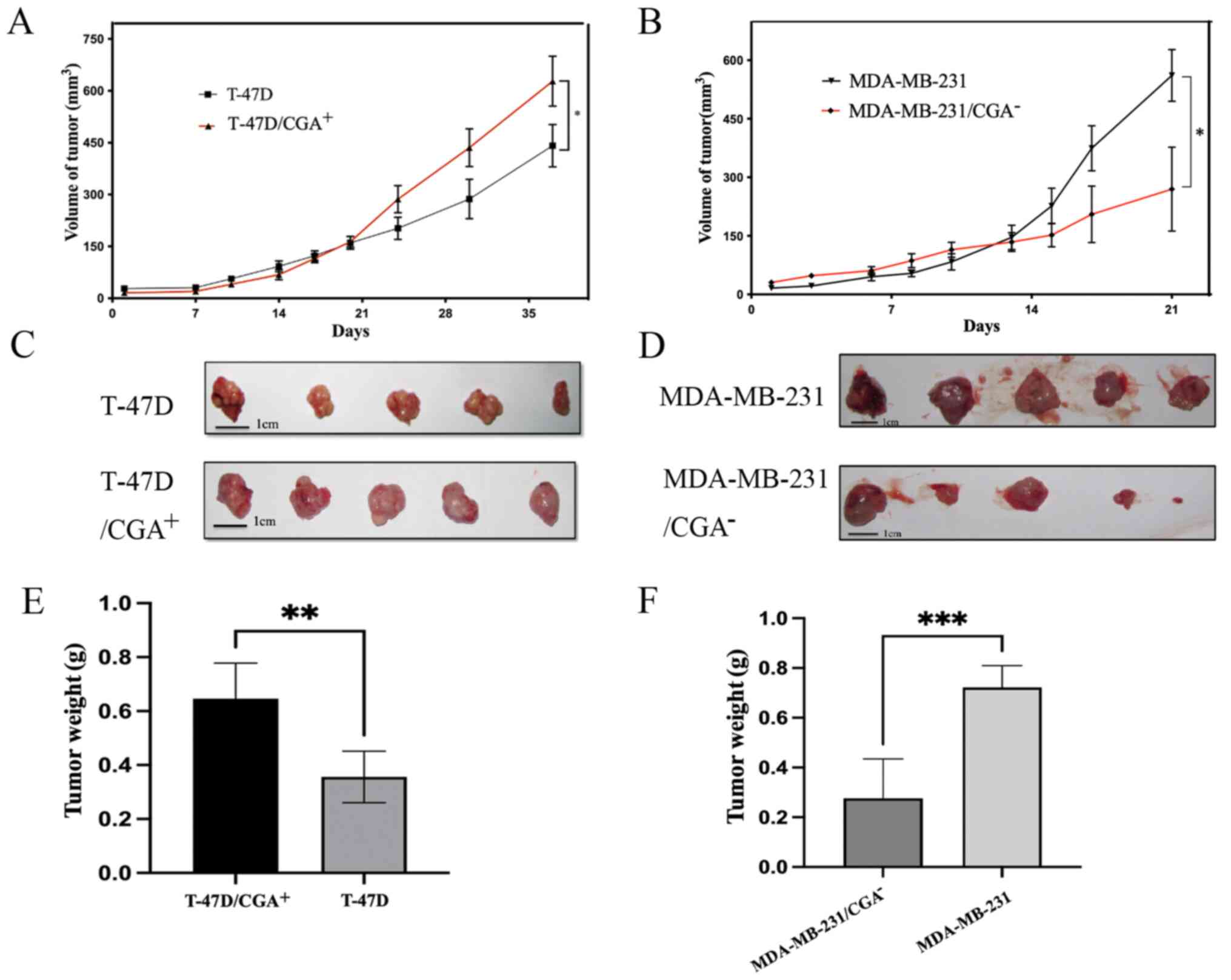
To evaluate the role of CGA in tumor growth in
vivo, a xenograft mouse model was established by subcutaneous
injection of T-47D/CGA+, T-47D,
MDA-MB-231/CGA− or MDA-MB-231 cells into BALB/c nude
mice. Tumors in the T-47D/CGA+ and MDA-MB-231 groups
grew more rapidly than those in the control groups, as assessed by
measuring the diameter of the tumor (Fig. 5A and B). As MDA-MB-231 cells grew
faster and T-47D cells grew relatively slowly, the cutoff times for
observations were therefore different. At the end of observation,
the largest tumor diameter in the T-47D cell group was 1.1 cm and
the largest volume was 617 mm3, while those values in
the T-47D/CGA+ group were 1.3 cm and 750 mm3,
respectively. The maximum tumor diameter and volume of the
MDA-MB-231 and MDA-MB-231/CGA− groups were 1.5 cm and
772 mm3, and 1.3 cm and 534 mm3,
respectively. The xenografts of the T-47D/CGA+ group
were relatively larger and heavier, while the low CGA expression in
the MDA-MB-231 group led to relatively smaller and lighter
xenografts (Fig. 5C-F).
Discussion
The present study found that increased CGA
expression was significantly associated with a poor prognosis in
patients with breast cancer. Consistent with the aforementioned
findings, CGA was undetectable in a normal mammary epithelial cell
line but was upregulated in breast cancer cell lines at both the
mRNA and protein levels. The upregulated expression of CGA promotes
the proliferation of breast cancer cells both in vitro and
in vivo. Although further research is needed to understand
the exact mechanism of CGA action, the present results indicated
that it promotes breast cancer cell proliferation through the
activation of the EGFR, ERK1/2 and Akt signaling cascades.
CGA is the α-subunit of glycoprotein hormones,
including FSH, LH and TSH, and is widely present in all parts of
the body. In recent years, it has been found to be closely
associated with some malignant tumors, such as those of prostate
(7), gastric (8) and breast cancer (15). Additionally, lung cancer cells lose
their tumor phenotypes and show decreased proliferative capacity
and tumorigenicity in mice upon depletion of α-hCG (6). In breast cancer, expression of the
α-subunit of hCG has been linked to lymph node metastasis and a
worse prognosis (15). In our
previous study, it was found that FSH levels were associated with
Her-2 and Ki67 expression in postmenopausal women with breast
cancer (16). In the present study,
it was further confirmed that a subunit of FSH was overexpressed in
breast cancer cells and promoted their proliferation. This suggests
that glycoprotein hormones may play a role in the tumorigenesis of
breast cancer, and are worthy of further study.
EGFR is a receptor tyrosine kinase that is usually
upregulated in cancer and functions as a proto-oncogene by
promoting proliferation and suppressing the apoptosis in cancer
cells (17). Activation of EGFR
induces downstream signaling molecules, including Akt and ERK1/2,
which are the components of two major pathways regulating cell
proliferation and survival (18).
The present study demonstrated that the increased proliferation of
breast cancer cells induced by CGA overexpression was associated
with the upregulation of p-EGFR (Y1173), p-ERK1/2 and p-Akt,
implying that CGA acts via these pathways to promote cell
proliferation. However, the direct target of CGA is not clear at
present, and this mechanism requires continued study.
To investigate the molecular basis for the
activation of intracellular signaling cascades by CGA in greater
detail, a gene association analysis of breast cancer samples from
TCGA database was performed using the CBioPortal platform
(http://www.cbioportal.org) (19,20). Big
data analysis also confirmed the expression variation of CGA in
various tumors, such as downregulated expression in some blood
tumors and high expression in lung, ovarian and breast cancer
(Fig. S1A). The upregulated
expression rate of CGA in breast cancer is nearly 1% according to
research data from five different databases (Fig. S1B). Patients with CGA-positive
breast cancer exhibited a lower overall survival rate, but the
difference was not statistically significant due to the small
amount of samples (Fig. S1C).
Pathway Mapper shows that CGA is associated with a number of breast
cancer proliferation-related signaling pathways, such as those of
WNT, TP53, PI3K and NOTCH. According to the results of GENEMANIA
analysis, one of the direct interaction partner proteins of CGA is
protein tyrosine phosphatase non-receptor type 12 (PTPN12)
(Fig. S1D) (21). PTPN12 is a member of the PTP family
and was recently identified as a tumor suppressor (22); it is downregulated in a variety of
human malignancies, including colon (23), breast (24) and ovarian (25). Inactivation of PTPN12 resulted in
HER-2/EGFR hyperactivation and stimulation of downstream MAPK
signaling in human mammary epithelial cells (24). In ovarian cancer cells, PTPN
silencing activated phosphatidylinositide 3-kinase/AKT signaling
(26). In the present study, T-47D
cells with high CGA expression were also evaluated with regard to
PTPN12, and it was found that PTPN12 was downregulated at the mRNA
level (Fig. S1E). According to the
experimental results, CGA can activate MAPK and AKT signaling
pathways and regulate the expression of PTPN12, indicating that CGA
may act through PTPN12, which is worthy of further study.
Based on the aforementioned observations and the
present study findings, it was indicated that CGA promoted breast
cancer progression via EGFR, ERK1/2 and Akt signaling. Although
additional studies are required to elucidate the underlying
mechanisms, these results indicate that therapeutic strategies
targeting CGA may be an effective treatment for breast cancer.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to acknowledge Dr Jiaping
Peng (The Key Laboratory of Cancer Prevention and Intervention,
China National Ministry of Education, Hangzhou, China), for
providing guidance on IHC.
Funding
This study was supported by the funding of the Key Program of
the Natural Science Foundation of Zhejiang Province (grant no.
LZ16H160002) and the National Science Foundation for Young
Scientists of China (grant no. 81502270).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ and XZ participated in the design and data
acquisition of the study, and wrote the original manuscript. YC,
HC, and SW participated in the data analysis and interpretation of
the study. All authors have read and approved the final manuscript.
JZ and XZ confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
the Women's Hospital, Zhejiang University School of Medicine
(Hangzhou, China) and was in accordance with the Declaration of
Helsinki. The protocol was approved by the Animal Care and Use
Committee of Zhejiang University (approval no. ZJU2015-339-01).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ji P, Gong Y, Jin ML, Hu X, Di GH and Shao
ZM: The burden and trends of breast cancer from 1990 to 2017 at the
global, regional, and national levels: Results from the global
burden of disease study 2017. Front Oncol. 10:6502020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Heer E, Harper A, Escandor N, Sung H,
McCormack V and Fidler-Benaoudia MM: Global burden and trends in
premenopausal and postmenopausal breast cancer: A population-based
study. Lancet Glob Health. 8:e1027–e1037. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fiddes JC and Goodman HM: The gene
encoding the common alpha subunit of the four human glycoprotein
hormones. J Mol Appl Genet. 1:3–18. 1981.PubMed/NCBI
|
|
5
|
Watkins PC, Eddy R, Beck AK, Vellucci V,
Leverone B, Tanzi RE, Gusella JF and Shows TB: DNA sequence and
regional assignment of the human follicle-stimulating hormone
beta-subunit gene to the short arm of human chromosome 11. DNA.
6:205–212. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rivera RT, Pasion SG, Wong DT, Fei YB and
Biswas DK: Loss of tumorigenic potential by human lung tumor cells
in the presence of antisense RNA specific to the ectopically
synthesized alpha subunit of human chorionic gonadotropin. J Cell
Biol. 108:2423–2434. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bièche I, Latil A, Parfait B, Vidaud D,
Laurendeau I, Lidereau R, Cussenot O and Vidaud M: CGA gene (coding
for the alpha subunit of glycoprotein hormones) overexpression in
ER alpha-positive prostate tumors. Eur Urol. 41:335–341. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li YL, Yang TS, Ruan WM, Cui W, Jin Y and
Zou XM: Effect of trichostatin a on SGC-7901 gastric cancer cells.
Int J Clin Exp Med. 7:1958–1966. 2014.PubMed/NCBI
|
|
9
|
Bieche I, Parfait B, Le Doussal V, Olivi
M, Rio MC, Lidereau R and Vidaud M: Identification of CGA as a
novel estrogen receptor-responsive gene in breast cancer: An
outstanding candidate marker to predict the response to endocrine
therapy. Cancer Res. 61:1652–1658. 2001.PubMed/NCBI
|
|
10
|
Bièche I, Parfait B, Noguès C, Andrieu C,
Vidaud D, Spyratos F, Lidereau R and Vidaud M: The CGA gene as new
predictor of the response to endocrine therapy in ER alpha-positive
postmenopausal breast cancer patients. Oncogene. 20:6955–6959.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Del Mastro L, Ceppi M, Poggio F, Bighin C,
Peccatori F, Demeestere I, Levaggi A, Giraudi S, Lambertini M,
D'Alonzo A, et al: Gonadotropin-releasing hormone analogues for the
prevention of chemotherapy-induced premature ovarian failure in
cancer women: Systematic review and meta-analysis of randomized
trials. Cancer Treat Rev. 40:675–683. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Blumenfeld Z: Fertility preservation and
GnRHa for chemotherapy: Debate. Cancer Manag Res. 6:313–315. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Day EK, Sosale NG and Lazzara MJ: Cell
signaling regulation by protein phosphorylation: A multivariate,
heterogeneous, and context-dependent process. Curr Opin Biotechnol.
40:185–192. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Walker RA: Significance of alpha-subunit
HCG demonstrated in breast carcinomas by the immunoperoxidase
technique. J Clin Pathol. 31:245–249. 1978. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou J, Chen Y, Huang Y, Long J, Wan F and
Zhang S: Serum follicle-stimulating hormone level is associated
with human epidermal growth factor receptor type 2 and Ki67
expression in post-menopausal females with breast cancer. Oncol
Lett. 6:1128–1132. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wee P and Wang Z: Epidermal growth factor
receptor cell proliferation signaling pathways. Cancers (Basel).
9:522017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lawlor MA and Alessi DR: PKB/Akt: A key
mediator of cell proliferation, survival and insulin responses? J
Cell Sci. 114:2903–2910. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Warde-Farley D, Donaldson SL, Comes O,
Zuberi K, Badrawi R, Chao P, Franz M, Grouios C, Kazi F, Lopes CT,
et al: The GeneMANIA prediction server: biological network
integration for gene prioritization and predicting gene function.
Nucleic Acids Res. 38:W214–W220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xunyi Y, Zhentao Y, Dandan J and Funian L:
Clinicopathological significance of PTPN12 expression in human
breast cancer. Braz J Med Biol Res. 45:1334–1340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Espejo R, Rengifo-Cam W, Schaller MD,
Evers BM and Sastry SK: PTP-PEST controls motility, adherens
junction assembly, and Rho GTPase activity in colon cancer cells.
Am J Physiol Cell Physiol. 299:C454–C463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun T, Aceto N, Meerbrey KL, Kessler JD,
Zhou C, Migliaccio I, Nguyen DX, Pavlova NN, Botero M, Huang J, et
al: Activation of multiple proto-oncogenic tyrosine kinases in
breast cancer via loss of the PTPN12 phosphatase. Cell.
144:703–718. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Villa-Moruzzi E: Tyrosine phosphatases in
the HER2-directed motility of ovarian cancer cells: Involvement of
PTPN12, ERK5 and FAK. Anal Cell Pathol (Amst). 34:101–112. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Villa-Moruzzi E: PTPN12 controls PTEN and
the AKT signalling to FAK and HER2 in migrating ovarian cancer
cells. Mol Cell Biochem. 375:151–157. 2013.PubMed/NCBI
|
|
27
|
Singletary SE and Greene FL; Breast Task
Force, : Revision of breast cancer staging: The 6th edition of the
TNM Classification. Semin Surg Oncol. 21:53–59. 2003. View Article : Google Scholar : PubMed/NCBI
|