Introduction
Lung cancer is one of the leading causes of
cancer-related death worldwide. Despite great advances in novel
treatments over the past few decades, the survival rate is still
low compared to that of other cancers (1,2).
Non-small cell lung cancer (NSCLC) is the most common form,
accounting for 85% of new diagnoses of lung cancer (3,4).
p53 is closely linked with a variety of
transcriptional and non-transcriptional activities that regulate
cell proliferation, DNA repair, cell senescence, and cell death
(5,6). Mouse double minute 2 homolog (MDM2)
is an E3 ligase that negatively regulates p53. When TP53 is
abnormally activated, MDM2 promotes poly-ubiquitination and causes
proteasomal degradation to downregulate p53, thereby maintaining
p53 at a normal level (7,8). Therefore, interference in the
interaction between MDM2 and p53 is an important part of the
anti-cancer treatment method targeting p53 activation (9).
HS-1793 (4-(6-hydroxy-2-naphthyl)-1.3-benzenediol),
a synthetic resveratrol analog, substituted the unstable double
bond present in resveratrol with a naphthalene ring (10,11).
HS-1793 improved photosensitivity and metabolic instability and is
more potent than resveratrol (12,13).
It is a well-known drug that causes cell death by inducing
apoptosis in several types of cancer, including breast, prostate,
colon, and renal cancers and multiple myeloma (14–17).
HS-1793 causes mitochondrial-mediated apoptosis in breast cancer
and also induces cell cycle arrest by reducing Akt phosphorylation
in colon cancer and multiple myeloma (14,17,18).
In prostate cancer cells, it inhibits migration by inhibiting the
expression of hypoxia-inducible factor-1α and vascular endothelial
growth factor (19). However, the
effect of HS-1793 on lung cancer has not yet been elucidated.
In the present study, we report the effect of
HS-1793 on A549 and H460 cell lines harboring p53 wild type.
HS-1793 inhibits the binding of p53 and MDM2, resulting in
increased p53 protein stability and expression of target genes and
subsequently inhibiting the proliferation of NSCLC cells.
Therefore, HS-1793 acts as a binding inhibitor of p53 and MDM2 and
is a potential drug target for lung cancer.
Materials and methods
Chemicals and antibodies
4-(6-Hydroxy-2-naphthyl)-1,3-benzenediol (HS-1793)
was purchased from AOBIOUS Inc. Antibodies against p53 (cat. no.
sc-126), GAPDH (cat. no. sc-47724), p21 waf1/Cip1 (cat. no.
sc-6246), and ubiquitin (cat. no. sc-8017) were purchased from
Santa Cruz Biotechnology. MDM2 (cat. no. op46) was purchased from
Calbiochem®. Anti-mouse IgG and anti-rabbit IgG were
purchased from Sigma-Aldrich; Merck KGaA.
Cell culture
A549 and H460 cell lines were obtained from American
Type Culture Collection (ATCC). The cells were cultured in
Dulbecco's modified Eagle medium (DMEM; HyClone, GE Healthcare) or
RPMI-1640 medium (HyClone) supplemented with 10% fetal bovine serum
(FBS; Corning Inc.) and 1% penicillin-streptomycin (Gibco;
Invitrogen; Thermo Fisher Scientific, Inc) and then incubated in a
5% CO2 incubator at 37°C.
Proliferation assay and cell
cycle
Cell viability was determined using a cell count kit
(CCK)-8 assay kit (Dojindo), according to the manufacturer's
instructions. Briefly, lung cancer cells were seeded into a 96-well
microplate, and CCK-8 solution (10/100 µl medium) was added to each
well of the plate. After incubation of the plate for 1–4 h in a
CO2 incubator at 37°C, the absorbance of each well was
measured at 450 nm using a microplate reader (Molecular Devices)
with a reference wavelength of 650 nm. Lung cancer cells were
treated with HS-1793 at the dose indicated in the figures. Cell
cycle distribution was determined by DNA staining with PI. Cells
were harvested and fixed in 70% ethanol. Cell pellets were
suspended in PI and simultaneously treated with RNase at 37°C for
30 min. The percentage of cells in different phases of the cell
cycle was measured using a FACScanto II flow cytometer (BD
biosciences).
Clonogenic assay
Cells were plated at equal densities in 6-well
plates (1×103 cells/well) for 24 h and then treated with
HS-1793 at the doses and times indicated in Fig. 1. After treatment, the cells were
trypsinized, serially diluted, and re-plated. The cells were grown
at 37°C with 5% CO2 for 7 days, and the colonies were
fixed and stained with 0.01% crystal violet. Subsequently, the
stained dyes were extracted, and the optical density (OD) at 560 nm
was measured using a 96-well plate reader (Molecular Devices).
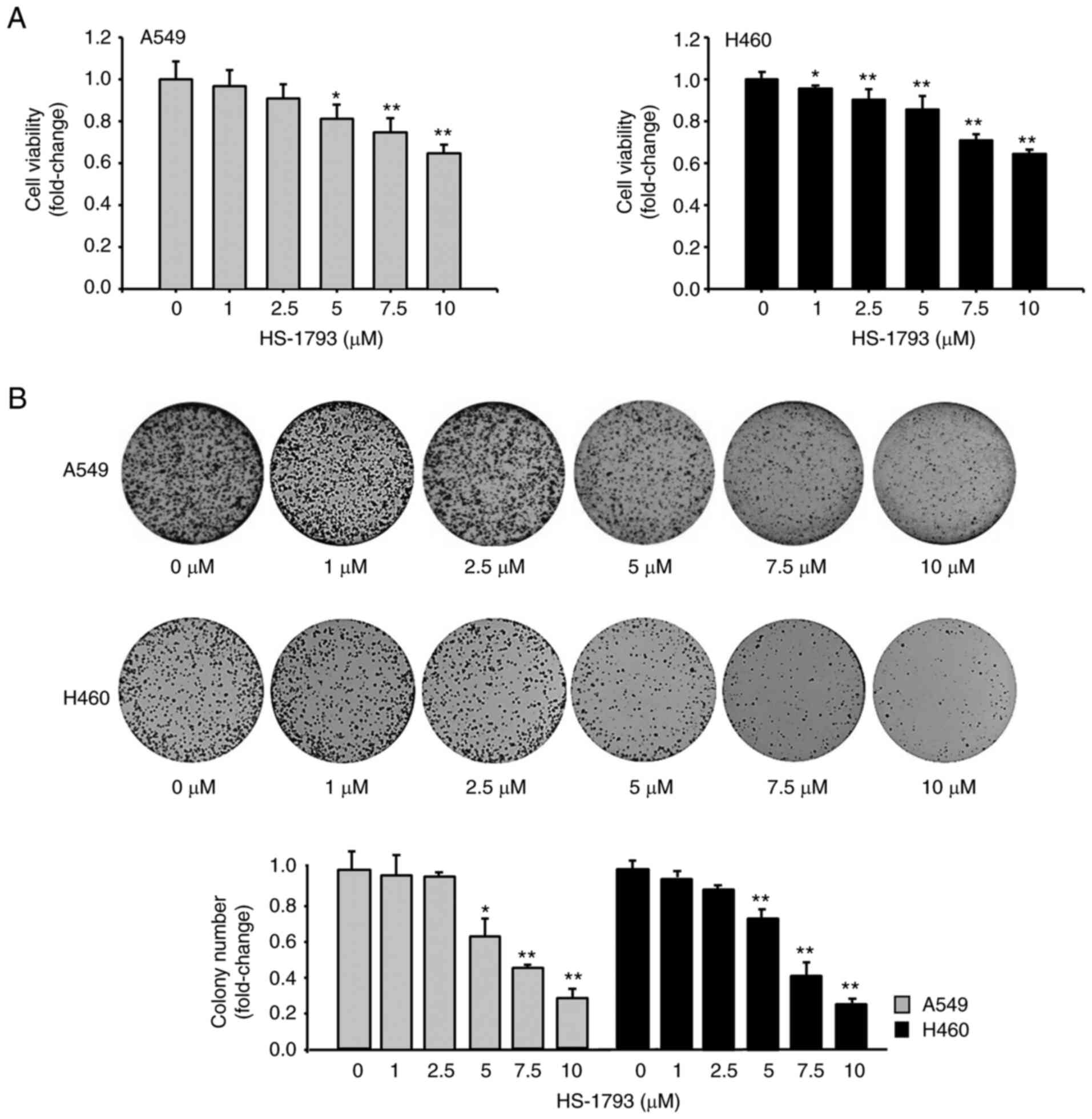 | Figure 1.HS-1793 diminishes the viability of
A549 and H460 cells. (A) A549 and H460 cells were treated with
HS-1793 at different concentrations (0, 1, 2.5, 5, 7.5, and 10 µM)
for 24 h, and absorbance was measured at 450 nm after 1 h
incubation, using a CCK-8 reagent to confirm cell viability.
Results are expressed as fold change. Data indicate the mean ±
standard deviation (SD) of three independent experiments.
*P<0.05, **P<0.01 vs. control. (B) The clonogenic assay was
performed using the same cells as above. After treatment with
HS-1793 in a dose-dependent manner, the media was replaced after 24
h. The media was then changed once every 2–3 days; the cells were
fixed after 7 days of observation and counted by staining with
crystal violet. Results are expressed as fold change. Data indicate
the mean ± SD of three independent experiments. *P<0.05,
**P<0.01 vs. control. CCK-8, cell counting kit-8. |
Protein isolation and western blot
analysis
Cells were lysed in lysis buffer A [20 mM HEPES (pH
7.5), 150 mM NaCl, 1 mM EDTA, 2 mM EGTA, 1% Triton X-100, 10%
glycerol, and protease inhibitor cocktail Set II (Sigma-Aldrich;
Merck KGaA)], and the cellular debris was removed by centrifugation
at 10,000 × g for 10 min. Proteins were separated using sodium
dodecyl sulfate-polyacrylamide gel electrophoresis, transferred
onto nitrocellulose membranes, blocked with 5% skim milk in 0.01 M
TBS (pH 7.5) containing 0.5% Tween-20, and labelled with the
appropriate primary antibodies. Antigen-antibody complexes were
detected based on chemiluminescence detection (Abclone). Protein
band intensity was measured using the ImageJ software.
Cycloheximide chase assay
Cycloheximide (CHX) chase assay was performed to
evaluate the regulation of p53 protein stability by HS-1793. Cells
were pre-incubated with or without 5 µM HS-1793 for 2 h.
Subsequently, 20 µg/ml CHX was added to inhibit protein synthesis.
The cells were collected at 0, 1, 3, 6, and 9 h following treatment
with CHX. Total cell lysates were analyzed using western blot with
anti-p53 and anti-GAPDH. The results from western blot analysis
were quantified using ImageJ, and the stable p53 protein level was
quantified.
RNA isolation and quantitative reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
RNA was extracted from A549 and H460 cells using an
Accuprep® Universal RNA extraction kit (Bioneer).
Briefly, 1 µg RNA was reverse-transcribed with oligo (dT) 12–18
primers using the First-Strand cDNA Synthesis Kit (Fermentas;
Thermo Fisher Scientific, Inc.). All reactions were performed in
triplicate, and the β-2-microglobulin (B2M) gene was used as
control. Using the comparative threshold cycle (Ct) or standard
method, relative quantification of p53 gene expression was
calculated after normalization against B2M for each sample. The
primers for qPCR were designed as follows: p53 primer, forward,
5′-CCTCACCATCATCACACTGG-3′ and reverse, 5′-CCTCATTCAGCTCTCGGAAC-3′;
B2M primer, forward, 5′-CTCGCTCCGTGGCCTTAG-3′ and reverse,
5′-CAAATGCGGCATCTTCAA-3′.
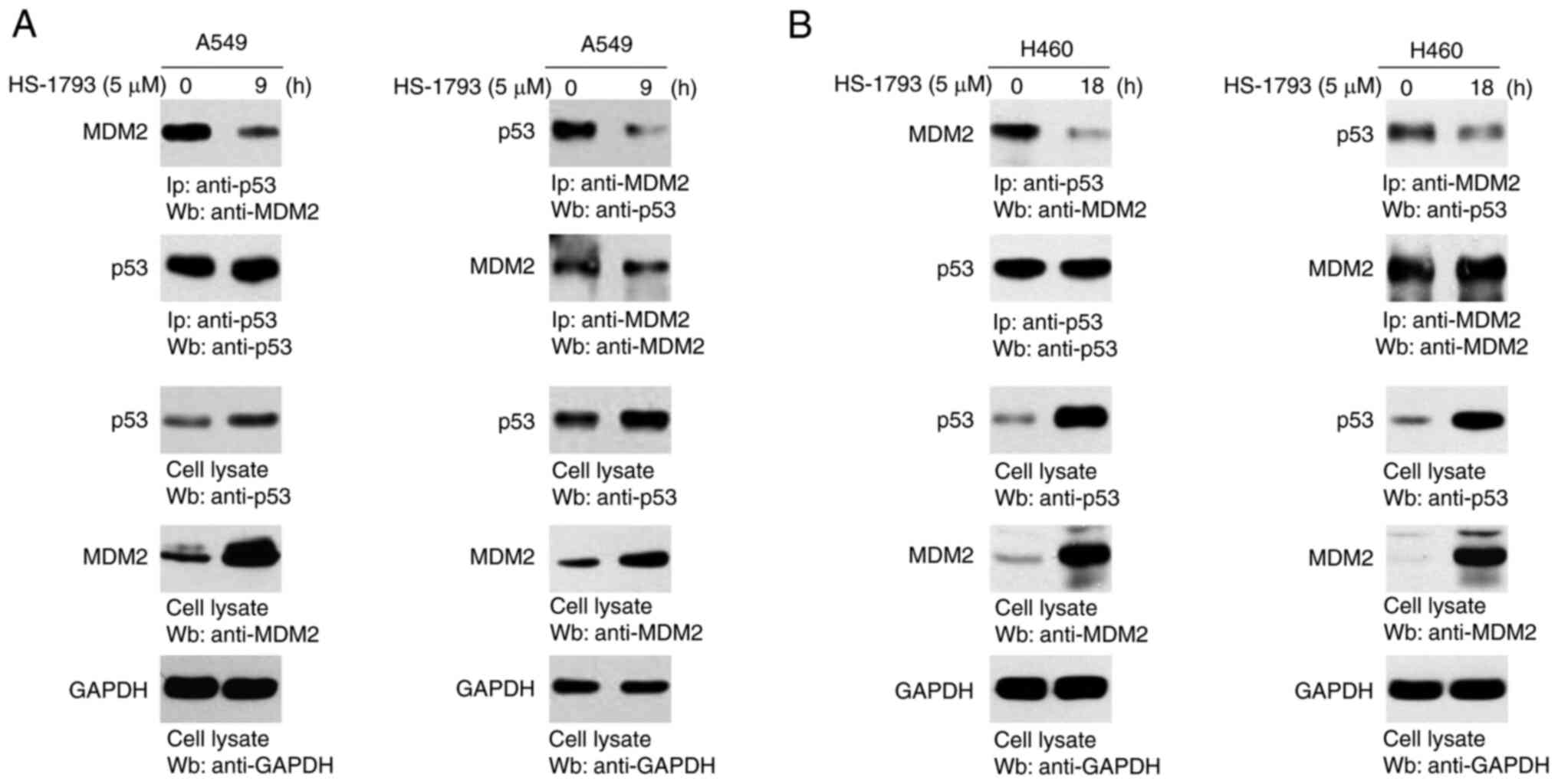
Immunoprecipitation (IP)
For co-IP, A549 and H460 cells were treated with
HS-1793 (5 µM) or DMSO for 9 h or 18 h. Cells for endogenous IP
were lysed in lysis buffer A. The lysate was immunoprecipitated
with antibodies to detect p53, MDM2, or normal immunoglobulin G.
Protein A/G sepharose beads (Santa Cruz Biotechnology) were then
applied, and the final precipitated proteins were quantified using
western blot analysis with the corresponding antibodies.
Statistical analysis
All data are obtained from three replicate
experiments and expressed as mean ± standard deviation of three
independent experiments. Statistical difference between multiple
groups were analyzed using one-way analysis of variance (ANOVA)
followed by Tukey's post hoc test, using SigmaPlot 12.0 software
(2013; Systat Software Inc.). P<0.05 was considered to indicate
a statistically significant difference.
Results
HS-1793 diminishes cell viability in
A549 and H460 cells
HS-1793 affects cell viability in various carcinomas
(14–17). The anti-cancer effect was
extensively examined in H1299, A549 and H460 lung cancer cells by
CCK-8 assay. Results showed that HS-1793 significantly inhibited
cell viability in A549 and H460 cells (high dose; 7.5 and 10 µM)
but not in H1299 cells, suggesting that A549 and H460 cells were
HS-1793 sensitive (Fig. S1).
Therefore, we decided to proceed with our studies in A549 and H460
cells. As shown in Fig. 1A, cell
viability was determined after treatment of A549 and H460 cells
with 0 to 10 µM HS-1793 for 24 h. HS-1793 has a slightly weaker
effect on cell viability at concentrations up to 5 µM at the 24 h
time point. However, at concentrations of 7.5 and 10 µM, HS-1793
induced a significant loss of viability (Fig. 1A). We made similar observation at
the different time points (Fig.
S2) We also determined G0-G1 arrest after exposure to low doses
of HS-1793 (<7.5 µM); however, the proportion of cells in the
G2-M was significantly increased after exposure to 10 µM (S3). In
addition, the clonogenic assay demonstrated that HS-1793 diminished
colony production in a dose-dependent manner (Fig. 1B). Based on these results, we
focused on the effects of HS-1793 at a concentration of 5 µM and a
24 h treatment in subsequent experiments.
HS-1793 increases p53 protein
expression and its target genes
Based on the findings of previous breast cancer
studies, to confirm the relationship between HS-1793 and p53 in
lung cancer, p53 protein level was confirmed using western
blotting. We first compared the basal level of p53 protein between
A549 and H460 cells. As shown in Fig.
S3, the basal level of p53 protein in A549 cells is
approximately 50% higher than that in H460 cells (Fig. S4). Next, to evaluate whether
HS-1793 regulated p53 protein level, we treated A549 and H460 cells
with concentration-dependent manner. HS-1793 increased the p53
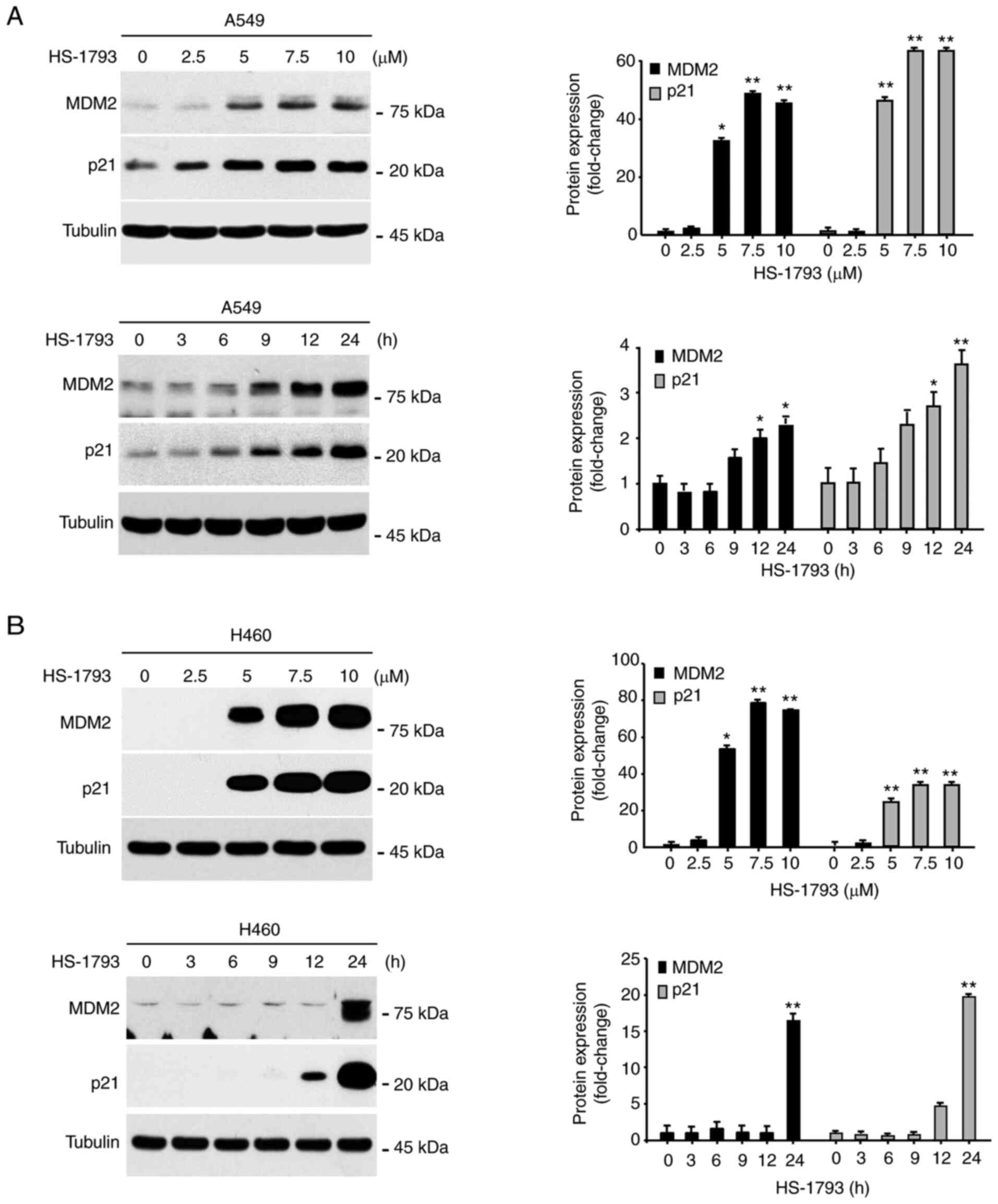
protein level in a concentration-dependent manner (Fig. 2A and B). As shown in Fig. 2C and D, HS-1793 also increased the
p53 protein level in a time-dependent manner. Next, we confirmed
whether p53 regulates its target genes when upregulated by HS-1793.
A549 and H460 cells were treated with HS-1793 at different doses
and time durations, cell lysates were prepared, and western blot
analysis was performed. As shown in Fig. 3A and B, HS-1793 significantly
increased the protein expression of p21 and MDM2 in A549 and H460
cells in a dose- and time-dependent manner. We also evaluated the
mRNA levels of p53 target gene (p21) in a time-dependent manner and
made a similar observation in A549 and H460 cells in response to
HS-1793 treatment (Fig. S5).
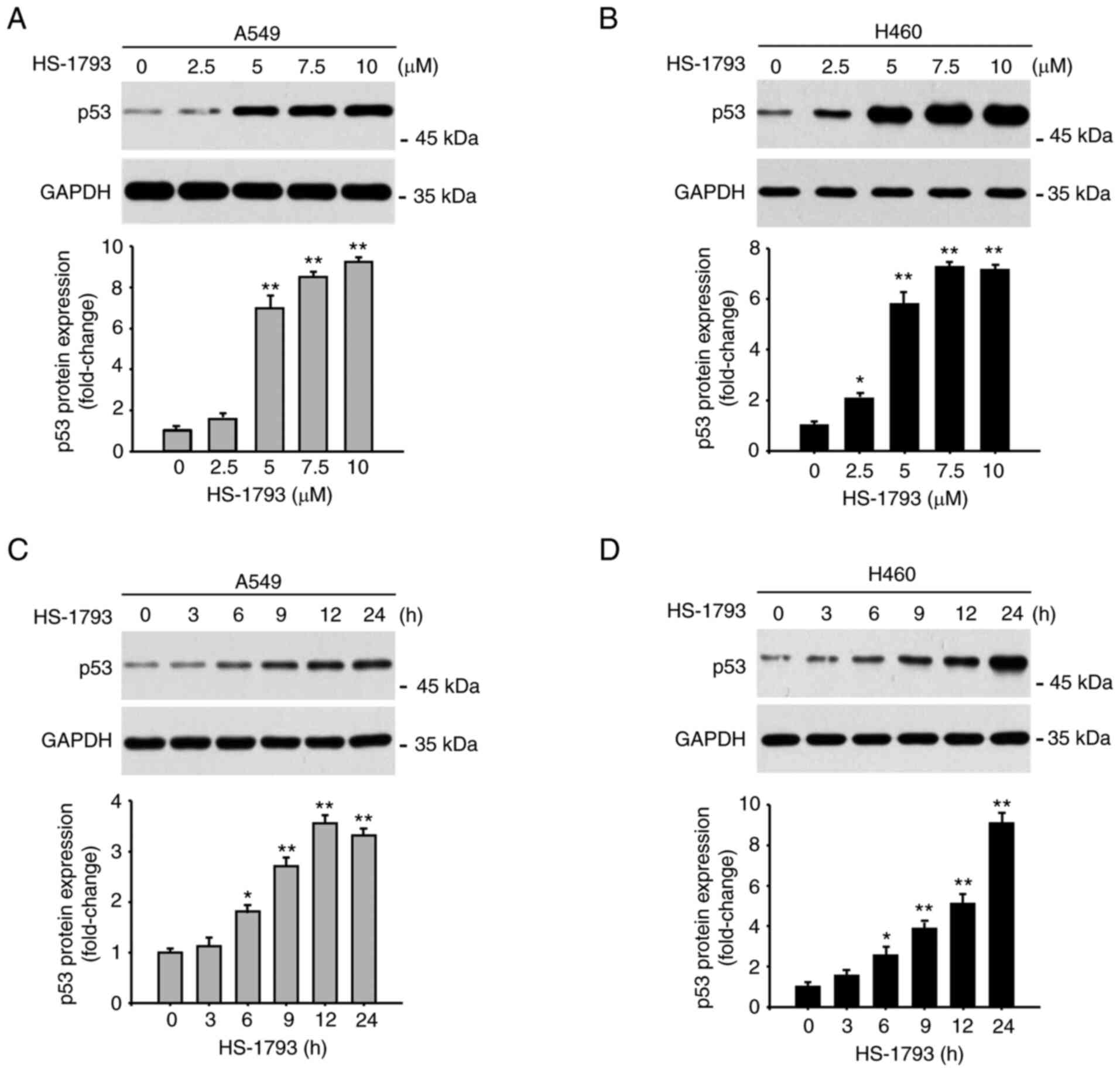 | Figure 2.HS-1793 increases p53 protein
expression. The protein expression of p53 in (A) A549 and (B) H460
cells was measured by western blot analysis. GAPDH was used as a
loading control at each concentration (0, 2.5, 5, 7.5, and 10 µM)
for 24 h. The experiment was independently performed twice,
quantified through the ImageJ program, normalized with GAPDH, and
displayed as a graph. The expression of p53 in (C) A549 and (D)
H460 cells was confirmed in a time-dependent manner (0, 3, 6, 9,
12, and 24 h) by selecting 5 µM concentration of HS-1793, which
showed a significant increase in the data above. Results are
expressed as fold change. Data indicate the mean ± standard
deviation of two independent experiments. *P<0.05, **P<0.01
vs. control. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. |
HS-1793-induced p53 protein expression
is mediated by a post-translational mechanism
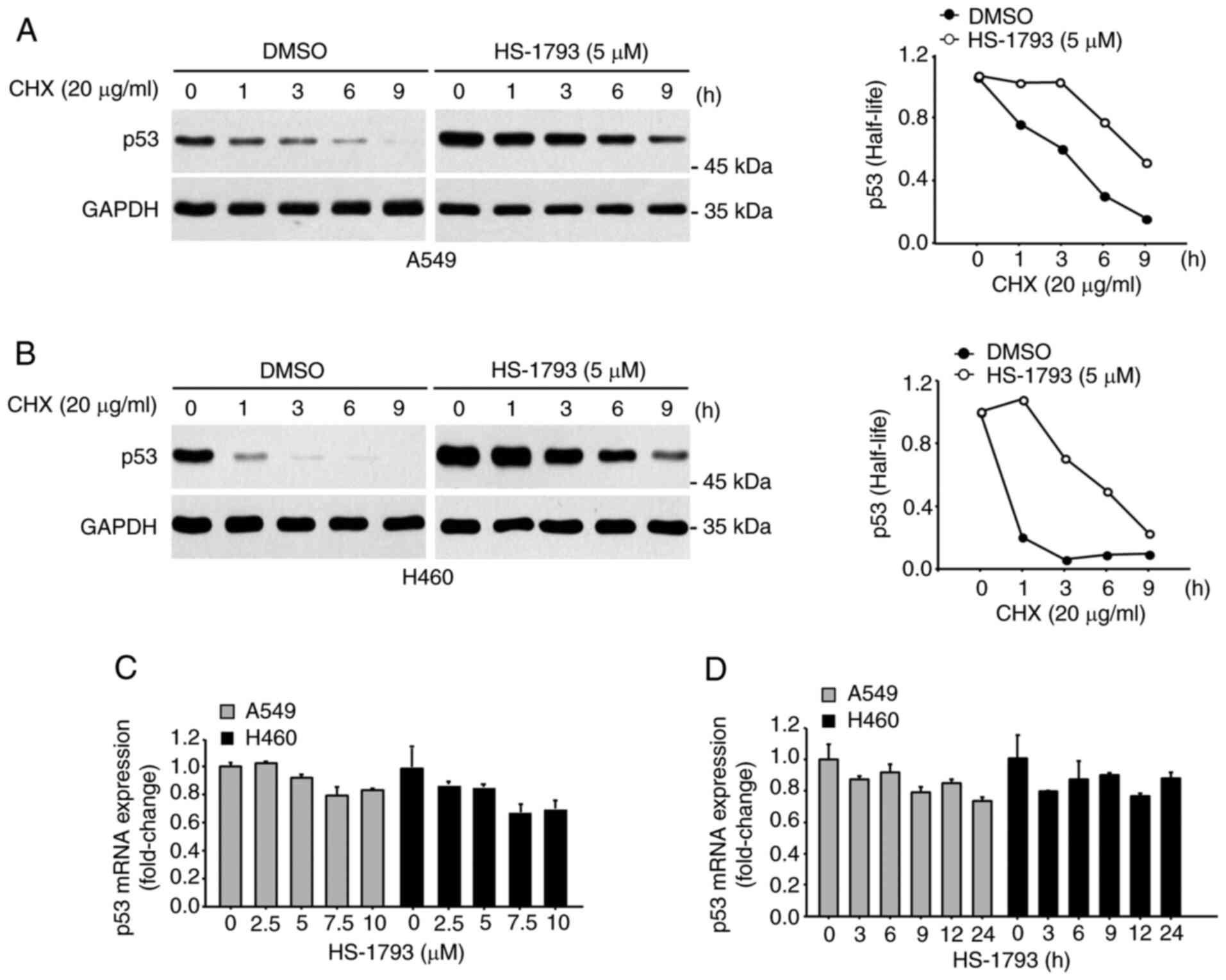
A CHX chase assay was performed to determine whether
the upregulation of p53 expression by HS-1793 was due to
transcription upregulation or increased protein stability. A549
(Fig. 4A) and H460 cells (Fig. 4B) were first treated with HS-1793
and then with CHX for the indicated durations. As shown in Fig. 4A and B, p53 half-life increased in
the presence of HS-1793. In contrast, the mRNA expression of p53
was not significantly changed in the presence of HS-1793 (Fig. 4C and D).
HS-1793 blocks the interaction between
p53 and MDM2
To identify the molecular mechanism underlying p53
stability, we performed an IP assay using antibodies specific for
p53 and MDM2. As shown in Fig. 5A and
B, p53 and MDM2 were strongly bound to each other when
antibodies for p53 or MDM2 were used. In contrast, HS-1793-treated
cells decreased to precipitate either protein, emphasizing that
HS-1793 regulates the interaction between p53 and MDM2.
Discussion
Lung cancer is one of the most common carcinomas
worldwide and is the primary cause of all cancer-related deaths in
both male and female cancer patients. There are four major
histological types of lung cancer, including small cell lung cancer
(SCLC) and three types of NSCLC (20,21).
As NSCLC accounts for over 85% of new lung cancer diagnoses, its
treatment is of great importance (22–24).
Early detection and multimodal care have selectively shown high
patient survival rates (25,26).
However, the overall treatment and survival rates for NSCLC are
low, and further studies are needed.
Resveratrol has demonstrated potential anti-cancer
activity against several types of human cancer, including prostate
(27,28), breast (29,30),
colorectal (31,32), and skin cancers (33), and has also been used as a
chemotherapeutic drug to treat various lung cancer cells (34–36).
Its anti-lung cancer effect has been widely studied, with results
suggesting that it could exert potent anti-tumor effects by
upregulating microtubule-associated protein 1 light chain 3
(37) and p53 (38), downregulating smad activator 2 and
4 (39), and inhibiting the
phosphatidylinositol-3-kinase pathway (40). Although resveratrol has potential
therapeutic properties, it has poor pharmacokinetic properties,
such as increased photosensitivity and rapid metabolization
(41,42). We, therefore, tested the
anti-cancer effect of resveratrol in lung cancer using its
synthetic analog HS-1793.
HS-1793 has enhanced pharmacokinetic properties,
such as reduced photosensitivity and metabolic instability, and is
more potent than resveratrol (12,13).
HS-1793 exhibits anti-cancer effects such as cell cycle arrest and
cell death in various carcinomas (14–18,43),
but its effects have not been demonstrated in lung cancer. We have
determined the effect of HS-193 on the growth of human lung cancer
cells. HS-1793 significantly inhibited the proliferation of both
A549 and H460 cells harboring wild-type p53. However, H1299 cells
containing p53 null were less affected (Fig. S3), suggesting that the p53
signaling pathway may play a pivotal role in HS-1793 treated cells.
Therefore, we decided to proceed with our studies in A549 and H460
cells harboring wild-type p53 allele.
In a previous paper, Kim et al identified a
relationship between HS-1793 and p53 in breast cancer (18), but the exact mechanism was unclear.
In this study, we showed that HS-1793 induced a dose-dependent
increase in p53 expression. In addition, we revealed that HS-1793
could significantly increase p53 stability in both A549 and H460
cells. Although a similar effects of HS-1793 observed for both
cells, unlike A549 cells, expression of the target genes of p53,
including p21 and MDM2, was delayed in H460 cells after HS-1793.
While wild-type p53 expression is observed in A549 and H460 cells,
the relative protein level is not equal between the cell lines. As
shown in Fig. S4, the protein
level in A549 cells is approximately 50% higher than that in H460
cells. The differential expression of p53 between these two cancer
cell lines may be attributed to the response of the HS-1793-treated
cells.
Several reports demonstrated that Akt represses p53
protein stability via Akt-mediated phosphorylation of MDM2
(44–47). Phosphorylation of MDM2 by Akt
enhances the nuclear accumulation of MDM2 and consequently augments
the destabilization of p53 by MDM2. Therefore, the Akt/MDM2 and
MDM2/p53 signaling pathways serve an important role in the
regulation of apoptosis and proliferation. We have previously shown
that HS-1793 regulates p-Akt activity as an inhibitor of Akt-HSP90
binding (17). Notably, in
multiple myeloma, Akt inhibition by HS-1793 results in NF-κB
pathway inhibition. We also have analyzed the dose-dependent effect
of HS-1793 on the p-Akt level in both A549 and H460 cells.
Interestingly, western blot analysis revealed that the p-Akt level
was markedly downregulated in response to HS-1793 in A549 cells,
whereas a gradual reduction in the p-Akt level was observed in H460
cells (data not shown). Therefore, the subtle difference between
the two cell lines for HS-1793 is probably due to p-Akt levels.
However, it is also unclear how HS-1793 regulates p53 expression
and target genes in lung cancer cells. Therefore, we consider the
detailed mechanisms for HS-1793-mediated regulation of p53 target
genes by defining a subset of lung tumors for further study.
p53 is a tumor suppressor gene that inhibits tumor
development by activating transcription factors related to the cell
cycle and apoptosis (48,49). Conversely, MDM2 is a
tumor-promoting gene that acts as a negative regulator of p53
(50,51). In various tumors, overexpression of
MDM2 inhibits p53 and promotes cell growth (9). Reactivation strategies of wild-type
p53, which do not function properly, such as inhibition of E3
ligase activity of MDM2 and using drugs that bind to the p53
pockets of MDM2, are known to be effective chemotherapy methods
(52). Thus, we assessed the
association between p53 and MDM2 in HS-1793-treated A549 and H460
cells. Our findings revealed that HS-1793 markedly reduced p53-MDM2
interactions. However, further studies are needed to determine
whether HS-1793 interferes with the binding of p53 to MDM2 directly
or indirectly.
We demonstrated that HS-1793 decreased cell
proliferation in lung cancer and increased p53 stability to elevate
the expression of its target genes p21 and MDM2. Under normal
conditions, MDM2, a negative regulator of p53, binds to and
degrades p53 by acting as an E3 ligase. However, this function is
blocked by HS-1793, confirming that p53 protein stability is
increased. In summary, the anti-cancer effect of HS-1793 in lung
cancer is due to p53 upregulation via blocking the p53-MDM2
interaction.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was supported by the Basic Research Program
through the National Research Foundation of Korea (NRF) and funded
by the MSIT (grant no. NRF-2020R1A4A1016029).
Availability of data and materials
The datasets used and/or analyzed during this study
are available from the corresponding author upon reasonable
request.
Authors' contributions
CL and SWJ designed the research. CL, PCWL and SS
performed the research. SS and SJ analyzed the data. SS and SWJ
wrote the paper. CL, PCWL, SS and SWJ confirm the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mangiameli G, Cioffi U, Alloisio M and
Testori A: Lung metastases: Current surgical indications and new
perspectives. Front Surg. 9:8849152022. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
J Saller J and Boyle TA: Molecular
pathology of lung cancer. Cold Spring Harb Perspect Med.
12:a0378122022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Isaacs J and Stinchcombe TE: Neoadjuvant
and adjuvant systemic therapy for early-stage non-small-cell lung
cancer. Drugs. 82:855–863. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tabbò F, DE Filippis M, Jacobs F and
Novello S: Strengths and pitfalls of brigatinib in non-small cell
lung cancer patients' management. Minerva Med. 113:315–332. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Thomas AF, Kelly GL and Strasser A: Of the
many cellular responses activated by TP53, which ones are critical
for tumour suppression? Cell Death Differ. 29:961–971. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koo N, Sharma AK and Narayan S:
Therapeutics targeting p53-MDM2 interaction to induce cancer cell
death. Int J Mol Sci. 23:50052022. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang S and Chen FE: Small-molecule MDM2
inhibitors in clinical trials for cancer therapy. Eur J Med Chem.
236:1143342022. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang H, Luo J and Lei H: The roles of
mouse double minute 2 (MDM2) oncoprotein in ocular diseases: A
review. Exp Eye Res. 217:1089102022. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chène P: Inhibiting the p53-MDM2
interaction: An important target for cancer therapy. Nat Rev
Cancer. 3:102–109. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jeong MH, Yang K, Lee CG, Jeong DH, Park
YS, Choi YJ, Kim JS, Oh SJ, Jeong SK and Jo WS: In vitro
genotoxicity assessment of a novel resveratrol analogue, HS-1793.
Toxicol Res. 30:211–220. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saiko P, Szakmary A, Jaeger W and Szekeres
T: Resveratrol and its analogs: Defense against cancer, coronary
disease and neurodegenerative maladies or just a fad? Mutat Res.
658:68–94. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Jeong SH, Jo WS, Song S, Suh H, Seol SY,
Leem SH, Kwon TK and Yoo YH: A novel resveratrol derivative,
HS1793, overcomes the resistance conferred by Bcl-2 in human
leukemic U937 cells. Biochem Pharmacol. 77:1337–1347. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jeong SH, Lee JS, Jeong NY, Kim TH, Yoo
KS, Song S, Suh H, Kwon TK, Park BS and Yoo YH: A novel resveratrol
analogue HS-1793 treatment overcomes the resistance conferred by
Bcl-2 and is associated with the formation of mature PML nuclear
bodies in renal clear cell carcinoma Caki-1 cells. Int J Oncol.
35:1353–1360. 2009.PubMed/NCBI
|
|
14
|
Jeong SH, Song IS, Kim HK, Lee SR, Song S,
Suh H, Yoon YG, Yoo YH, Kim N, Rhee BD, et al: An analogue of
resveratrol HS-1793 exhibits anticancer activity against MCF-7
cells via inhibition of mitochondrial biogenesis gene expression.
Mol Cells. 34:357–365. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jeong NY, Yoon YG, Rho JH, Lee JS, Lee SY,
Yoo KS, Song S, Suh H, Choi YH and Yoo YH: The novel resveratrol
analog HS-1793-induced polyploid LNCaP prostate cancer cells are
vulnerable to downregulation of Bcl-xL. Int J Oncol. 38:1597–1604.
2011.PubMed/NCBI
|
|
16
|
Kim DH, Kim MJ, Sung B, Suh H, Jung JH,
Chung HY and Kim ND: Resveratrol analogue, HS-1793, induces
apoptotic cell death and cell cycle arrest through downregulation
of AKT in human colon cancer cells. Oncol Rep. 37:281–288. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Song IS, Jeong YJ, Jeong SH, Kim HK, Ha
NC, Shin M, Ko KS, Rhee BD, Shim S, Jang SW and Han J:
Pharmacologic inhibition of AKT leads to cell death in relapsed
multiple myeloma. Cancer Lett. 432:205–215. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim JA, Kim DH, Hossain MA, Kim MY, Sung
B, Yoon JH, Suh H, Jeong TC, Chung HY and Kim ND: HS-1793, a
resveratrol analogue, induces cell cycle arrest and apoptotic cell
death in human breast cancer cells. Int J Oncol. 44:473–480. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim DH, Hossain MA, Kim MY, Kim JA, Yoon
JH, Suh HS, Kim GY, Choi YH, Chung HY and Kim ND: A novel
resveratrol analogue, HS-1793, inhibits hypoxia-induced HIF-1α and
VEGF expression, and migration in human prostate cancer cells. Int
J Oncol. 43:1915–1924. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Minna JD, Roth JA and Gazdar AF: Focus on
lung cancer. Cancer Cell. 1:49–52. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lemjabbar-Alaoui H, Hassan OU, Yang YW and
Buchanan P: Lung cancer: Biology and treatment options. Biochim
Biophys Acta. 1856:189–210. 2015.PubMed/NCBI
|
|
22
|
Gridelli C, Rossi A, Carbone DP, Guarize
J, Karachaliou N, Mok T, Petrella F, Spaggiari L and Rosell R:
Non-small-cell lung cancer. Nat Rev Dis Primers. 1:150092015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zappa C and Mousa SA: Non-small cell lung
cancer: Current treatment and future advances. Transl Lung Cancer
Res. 5:288–300. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hoffman PC, Mauer AM and Vokes EE: Lung
cancer. Lancet. 355:479–485. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yang K, Wang YJ, Chen XR and Chen HN:
Effectiveness and safety of bevacizumab for unresectable
non-small-cell lung cancer: A meta-analysis. Clin Drug Investig.
30:229–241. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Silk N, Reich J, Sinha R, Chawla S, Geary
K and Zhang D: The effects of resveratrol on prostate cancer
through targeting the tumor microenvironment. J Xenobiot. 11:16–32.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zaffaroni N and Beretta GL: Resveratrol
and prostate cancer: The power of phytochemicals. Curr Med Chem.
28:4845–4862. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Medina-Aguilar R, Marchat LA, Arechaga
Ocampo E, Gariglio P, García Mena J, Villegas Sepúlveda N, Martínez
Castillo M and López-Camarillo C: Resveratrol inhibits cell cycle
progression by targeting Aurora kinase A and Polo-like kinase 1 in
breast cancer cells. Oncol Rep. 35:3696–3704. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alamolhodaei NS, Tsatsakis AM, Ramezani M,
Hayes AW and Karimi G: Resveratrol as MDR reversion molecule in
breast cancer: An overview. Food Chem Toxicol. 103:223–232. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schneider Y, Vincent F, Duranton B, Badolo
L, Gossé F, Bergmann C, Seiler N and Raul F: Anti-proliferative
effect of resveratrol, a natural component of grapes and wine, on
human colonic cancer cells. Cancer Lett. 158:85–91. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tessitore L, Davit A, Sarotto I and
Caderni G: Resveratrol depresses the growth of colorectal aberrant
crypt foci by affecting bax and p21(CIP) expression.
Carcinogenesis. 21:1619–1622. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fabbrocini G, Kisslinger A, Iannelli P,
Vitale N, Procaccini C, Sparaneo G, Chieffi P, Ayala F, Mancini FP
and Tramontano D: Resveratrol regulates p66Shc activation in HaCaT
cells. Exp Dermatol. 19:895–903. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Feng Y, Zhou J and Jiang Y: Resveratrol in
lung cancer-a systematic review. J BUON. 21:950–953.
2016.PubMed/NCBI
|
|
35
|
Rasheduzzaman M, Jeong JK and Park SY:
Resveratrol sensitizes lung cancer cell to TRAIL by p53 independent
and suppression of Akt/NF-κB signaling. Life Sci. 208:208–220.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kong F, Zhang R, Zhao X, Zheng G, Wang Z
and Wang P: Resveratrol raises in vitro anticancer effects of
paclitaxel in NSCLC cell line A549 through COX-2 expression. Korean
J Physiol Pharmacol. 21:465–474. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ohshiro K, Rayala SK, Kondo S, Gaur A,
Vadlamudi RK, El-Naggar AK and Kumar R: Identifying the estrogen
receptor coactivator PELP1 in autophagosomes. Cancer Res.
67:8164–8171. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luo H, Yang A, Schulte BA, Wargovich MJ
and Wang GY: Resveratrol induces premature senescence in lung
cancer cells via ROS-mediated DNA damage. PLoS One. 8:e600652013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Whyte L, Huang YY, Torres K and Mehta RG:
Molecular mechanisms of resveratrol action in lung cancer cells
using dual protein and microarray analyses. Cancer Res.
67:12007–12017. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ebi H, Tomida S, Takeuchi T, Arima C, Sato
T, Mitsudomi T, Yatabe Y, Osada H and Takahashi T: Relationship of
deregulated signaling converging onto mTOR with prognosis and
classification of lung adenocarcinoma shown by two independent in
silico analyses. Cancer Res. 69:4027–4035. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Shaito A, Posadino AM, Younes N, Hasan H,
Halabi S, Alhababi D, Al-Mohannadi A, Abdel-Rahman WM, Eid AH,
Nasrallah GK and Pintus G: Potential adverse effects of
resveratrol: A literature review. Int J Mol Sci. 21:20842020.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Neves AR, Lucio M, Lima JL and Reis S:
Resveratrol in medicinal chemistry: A critical review of its
pharmacokinetics, drug-delivery, and membrane interactions. Curr
Med Chem. 19:1663–1681. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kim DH, Hossain MA, Kim MY, Kim JA, Yoon
JH, Suh HS, Kim GY, Choi YH, Chung HY and Kim ND: A novel
resveratrol analogue, HS-1793, inhibits hypoxia-induced HIF-1α and
VEGF expression, and migration in human prostate cancer cells. Int
J Oncol. 43:1915–1924. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chen Y, Li H, Zhang W, Qi W, Lu C, Huang
H, Yang Z, Liu B and Zhang L: Sesamin suppresses NSCLC cell
proliferation and induces apoptosis via Akt/p53 pathway. Toxicol
Appl Pharmacol. 387:1148482020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chibaya L, Karim B, Zhang H and Jones SN:
Mdm2 phosphorylation by Akt regulates the p53 response to oxidative
stress to promote cell proliferation and tumorigenesis. Proc Natl
Acad Sci USA. Jan 26–2021.(Epub aheac of print). View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhao Y, Cai J, Shi K, Li H, Du J, Hu D,
Liu Z and Wang W: Germacrone induces lung cancer cell apoptosis and
cell cycle arrest via the Akt/MDM2/p53 signaling pathway. Mol Med
Rep. 23:4522021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu L, Xu Z, Yu B, Tao L and Cao Y:
Berbamine inhibits cell proliferation and migration and induces
cell death of lung cancer cells via regulating c-Maf, PI3K/Akt, and
MDM2-P53 pathways. Evid Based Complement Alternat Med.
2021:55171432021.PubMed/NCBI
|
|
48
|
Aubrey BJ, Kelly GL, Janic A, Herold MJ
and Strasser A: How does p53 induce apoptosis and how does this
relate to p53-mediated tumour suppression? Cell Death Differ.
25:104–113. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Ozaki T and Nakagawara A: Role of p53 in
cell death and human cancers. Cancers (Basel). 3:994–1013. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Shi D and Gu W: Dual roles of MDM2 in the
regulation of p53: Ubiquitination dependent and ubiquitination
independent mechanisms of MDM2 repression of p53 activity. Genes
Cancer. 3:240–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Klein AM, de Queiroz RM, Venkatesh D and
Prives C: The roles and regulation of MDM2 and MDMX: It is not just
about p53. Genes Dev. 35:575–601. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Athar M, Elmets CA and Kopelovich L:
Pharmacological activation of p53 in cancer cells. Curr Pharm Des.
17:631–639. 2011. View Article : Google Scholar : PubMed/NCBI
|