Introduction
Epigenetic mechanisms are essential for the normal
development and maintenance of cell and tissue-specific gene
expression patterns in mammals (1). DNA methylation, histone modification,
nucleosome remodeling, and RNA-mediated targeting regulate numerous
biological processes that are fundamental to the genesis of cancer.
Disruption of epigenetic processes can lead to altered gene
function that can induce malignant cell transformation (2). Histone methylation plays an important
role in the regulation of genes expression, its dysregulation has
been observed in various cancers (3–7).
Regulation of methylation is mediated by two types of
enzymes-histone methyltransferases, which add methyl groups to
arginine and lysine residues, and histone demethylases (lysine
demethylase-KDM), which remove methyl groups (3). The KDM5 family of lysine demethylases
known also as Jumonji C (JmjC) or JARID1 that have four members
(KDM5A-D), demethylate di- and tri-methylated H3K4 (8). These enzymes are
2-oxoglutarate-dependent dioxygenases that require for their
function Fe2+ and oxygen in order to undergo the
hydroxylation necessary to remove the methyl groups (8). KDM5B, also known as JARID1B, has been
found to associate with transcription factors PAX9, FOXC2 and
FOXG1. It can also repress or promote activation of target genes by
interacting with nuclear hormonal receptors (9). The levels of enzyme modifying
histones KDM5B determine the hyperactivation of PI3K/AKT signaling
in prostate cancer (10).
Dysregulation of KDM5B has been identified in numerous
cancers e.g. laryngeal squamous cell carcinoma, bladder, breast
cancer, and is closely correlated with tumorigenesis, metastasis,
and worse survival in humans (5–7).
Therefore, this enzyme might be a potential promising target for
novel cancer diagnostic and/ or treatment.
KDM5B has been described as important for the
formation and maintenance of cancer stem cells in neuroblastoma
cell lines (NBL) (11). In
addition, its overexpression was a marker of shorter relapse-free
survival in patients with NBL (11). NBL is a malignant embryonal tumor
in children, emerging from the peripheral nervous system. The
biology of NBL is heterogeneous; small groups of NBL regress
spontaneously, while numerous cases have aggressive behavior. For
high-risk neuroblastoma (HR-NBL) is characteristic development of
chemoresistance (12). Patients
suffering from HR-NBL have a 5-year overall survival rate of ~40%
despite all intensive multimodal therapies. To date, there are no
salvage treatment regimens known to be curative (12,13).
Knowledge of MYCN properties is limited because its expression is
in physiological conditions limited to the early stages of
embryonic development (14). N-myc
protein interacts with Max and its high levels, which occurs in
MYCN amplified NBL, lead to a large number of
transcription-activating complexes (15). MYCN overexpression induces
proliferation and suppresses apoptosis and differentiation in NBL
cells (16). Several studies
proved that MYCN silencing in MYCN amplified NBL cells suppressed
growth and induced apoptosis and differentiation e.g. (14,16,17).
Its expression was higher in MYCN amplified NBL cell lines than in
MYCN-non-amplified NBL cells (11).
MYCN amplification correlates with poor outcome of
NBL patients. Examination of MYCN amplification is part of
diagnostic scheme in NBL. MYCN is MYC family of transcription
factors member. Those transcription factors are regulators of
cellular proliferation, differentiation and survival (12,13).
The aim of this study is to investigate the
importance of KDM5 expression for the growth of NBL cells and their
chemoresistance to cisplatin [CDDP abbreviation of
cis-diaminedichloroplatinum (II)].
Materials and methods
Cell culture and chemicals
Human HR-NBL cell lines UKF-NB-4,
UKF-NB-4CDDP, SK-N-AS, SK-N-ASCDDP,UKF-NB-3
and UKF-NB-3CDDP were donated by prof. J. Cinatl, Dr.
Sc. From Goethe University in Frankfurt am Main. Cells were grown
in Iscove's Modified Dulbecco's medium (IMDM) supplemented with 10%
(v/v) fetal bovine serum (both Thermo Fisher Scientific) and
incubated at 37°C in 5% CO2. For experiments,
8×105 cells were seeded in 22,1 cm2 dishes
and after 24 h treated with cisplatin (Ebewe) in final
concentration 20 µM for 48 h. Both cell lines amplified MYCN gene
as we proved by FISH (data not shown).
Assessment of cisplatin
cytotoxicity
To evaluate CDDP cytotoxicity, MTT
(3-(4,5-dimethylthiazol-2-yl)-2.5 diphenyltetrazolium bromide)
assay was performed. 104 cells/well were seeded in
96-well cell culture plate and cells were treated with CDDP at
final concentration 0.6–300 µM for 48 h. Subsequently, MTT solution
(2 mg/ml in PBS) (Fluka) was added and the plate was placed in an
incubator for 2 h. Cells were then lysed in solution of 20% of SDS
(Invitrogen) containing 50% N,N-dimethylformamide
(Sigma-Aldrich), pH 4.5, and the absorbance at 570 nm was measured
by multiwell ELISA reader Versamax (Molecular Devices). The optical
density of the medium was read as background and the optical
density value of the live control cells was taken as 100%. The
values of IC50 were determined using at least 3
independent measurements by SOFTmaxPro software.
Transfection
NB cells were transfected with a smart pool siRNA to
KDM5B ON-TARGETplus Human KDM5B siRNA, cat. No. L-009899-00-0020
(https://horizondiscovery.com/en/search?searchterm=L-009899-00-0020)
and Lincode non-targeting siRNA Lincode Non-targeting siRNA #1,
cat. No. D-001320-01-20 (https://horizondiscovery.com/en/search?searchterm=D-001320-01-20+)
using Dharmafect transfection reagent (all purchased from
Dharmacon) according to the manufacturer's instructions. The siRNA
concentration was 25 nM.
RNA isolation and quiantitative
RT-PCR
RNA was isolated using PureLink RNA Mini Kit (Thermo
Fisher Scientific) according to the manufacturer's protocol.
Quantity and quality were verified using the NanoDrop One
spectrophotometer (Thermo Fisher Scientific). Reverse transcription
was performed using gb Reverse Transcription Kit (Generi Biotech)
and 1,000 ng of RNA was used for complementary DNA synthesis.
Primers and probes hKDM5B_Q1 and POLR2A that was used as an
endogenous control (18), were
designed and produced by Generi Biotech. Custom oligo synthesis,
cat. No. 1000-020 for gene: KDM5B; Gene ID: 10765 POLR2A; Gene ID:
5430 (https://www.generi-biotech.com/products/custom-oligo-synthesis/).
We used POLR2A as an internal standard because it is homogeneously
and uniformly expressed in NBL cells (18). It is also used by other groups
studying NBL (19,20). The quantification of gene
expression was performed using QuantStudio 3 Real-Time PCR System
(Thermo Fisher Scientific) in triplicate. The temperature profile
was: 95°C for 3 min, 50 cycles of 95°C for 10 sec, 60°C 20 sec.
Fold change values were determined using REST 2009 software.
Western blot analysis
Proteins were extracted in RIPA Buffer supplemented
with Complete protease inhibitor cocktail (Roche) and their
concentration was measured by DC protein assay (Bio-Rad
Laboratories). Samples (40 µg) were resolved on SDS polyacrylamide
gels and blotted on nitrocellulose membranes (Bio-Rad). Primary
antibody JARID1B Rabbit mAb (Cell Signaling Technology) was diluted
1:1,000, β-actin Mouse mAb (Sigma-Aldrich) diluted 1:3,000 was used
as a loading control. Secondary antibodies Europium conjugated
anti-IgG (Molecular Devices) were diluted 1:5,000. Membranes were
visualized by SpectraMax i3× Multi-Mode Microplate Reader
(Molecular Devices). ImageJ 1.52a software was employed for the
analysis.
Cell viability assay
Cells were seeded in 24-well cell culture plate at a
density of 4×104 cells/well and incubated with
PrestoBlue Cell Viability Reagent (Thermo Fisher Scientific) for 30
min at 37°C. The fluorescence was measured using an excitation
wavelength of 560 nm and emission of 590 nm by SpectraMax i3×
Multi-Mode Microplate Reader (Molecular Devices). Each sample was
analyzed in triplicate.
Cell proliferation
Cells after transfection were seeded into 16-well
E-plates for impedance-based detection (ACEA Bioscience Inc) at
density of 104 cells per well. The xCELLigence RTCA DP
Instrument (ACEA Bioscience Inc) placed in a humidified incubator
at 37°C and 5% CO2 was used for real-time monitoring of
cell proliferation. The cell index was monitored every 30 min for
85 h and data were recorded by the supplied RTCA software. Each
sample was analyzed in triplicate.
Determination of histone H3K4
methylation status and KDM5B level
Flow cytometry was used for the detection of H3K4
trimethylation and expression of KDM5B on protein level. Cells
after treatment and/or transfection were washed with cold PBS
(Thermo Fisher Scientific), trypsinized (trypsin-Thermo Fisher
Scientific) and collected by centrifugation. Pellets of cells were
washed with PBS and fixed in 3.6% paraformaldehyde for 15 min at
room temperature. Cell pellets were then washed with PBS and
permeabilized by 90% methanol for 1 h at −20°C. Pellets were
subsequently washed 3 times with 0.5% bovine serum albumin
(BSA-Roth) in PBS and were resuspended in primary antibody JARID1B
Rabbit mAB diluted 1:1,000 (Cell Signaling Technology) or
Anti-trimethyl-Histone H3 (Lys4) Rabbit (EMD Millipore Corp.) at
dilution 1:400 and incubated for 1 h at laboratory temperature.
Cells were then washed with 0.5% BSA, resuspended in
fluorochrome-conjugated secondary antibody Anti-Rabbit IgG (H+L)
Alexa Fluor® 647 Conjugate (Thermo Fisher Scientific)
diluted 1:500 and incubated for 30 min at room temperature in dark.
Washed and re-suspended cells were measured using a BD FACSCelesta
(BD Bioscience), and data were analyzed by Flowlogic software
(Inivai Technologies).
Cell cycle analysis
Cells after treatment and/or transfection were
washed with cold PBS, trypsinized and collected by centrifugation.
Pellets of cells were washed with PBS and fixed in 3.6%
paraformaldehyde for 10 min at room temperature. Cell pellets were
then washed with PBS and permeabilized by 90% methanol for 1 h at
−20°C. Pellets were washed with PBS, resuspended in 500 µl PBS and
one drop of FxCycle™ Violet Ready Flow™ Reagent (Thermo Fisher
Scientific) was added and after 30 min incubation were cells
measured using a BD FACSCelesta (BD Bioscience), and data were
analyzed by Flowlogic software (Inivai Technologies).
Wound healing assay
Neuroblastoma cells were seeded in 9.2
cm2 dish in number 1.6×105 cells/ml of
sensitive cells and 2.2×105 cells/ml of resistant cells,
that allowed to reach 70% confluence for 24 h at 37°C in 5%
CO2 and then transfected with siRNA to KDM5B and Lincode
non-targeting siRNA. 48 h after transfection was drawn the line
across the dish's surface using a 1,000 µl sterile plastic tip, at
that time the confluence was more than 80%. After wounding, cells
were grown in Iscove's Modified Dulbecco's medium (IMDM) with 5%
(v/v) FBS (both Thermo Fisher Scientific) and incubated at 37°C in
5% CO2. For scratch assay, 80–90% confluence is
recommended so that the cells do not overgrow (21,22).
Pictures were captured at the same field immediately, 24 and 48 h
after the wounding by microscope Olympus IX71 (Olympus) and ImageJ
1.52a software was employed for the analysis.
Statistical analysis
All experiments were independently repeated at least
three times and data are shown as averages ± standard error.
One-way Anova with post-hoc Tukey HSD and two-way ANOVA followed by
Bonferroni test (https://astatsa.com/OneWay_Anova_with_TukeyHSD/) were
utilized when comparing the situations. Results from RT-qPCR were
statistically compared using REST 2009 software (23). Significances (P<0.05 was
considered as significant) of the statistical analyses are shown in
individual Figures and described in their legends.
Results
KDM5B is downregulated in resistant
neuroblastoma cell line
UKF-NB-4CDDP resulting from long-term
cultivation with an increasing dose of CDDP was used as a model of
drug resistance (24,25). We used this wide concentration
range only to determine IC50 using MTT test to
demonstrate lower sensitivity in the cell line with experimentally
induced chemoresistance (UKF-NB-4CDDP) compared to
sensitive cells (UKF-NB-4). We used only one concentration (20
mikroM) in further experiments. This cell line has ~4 times higher
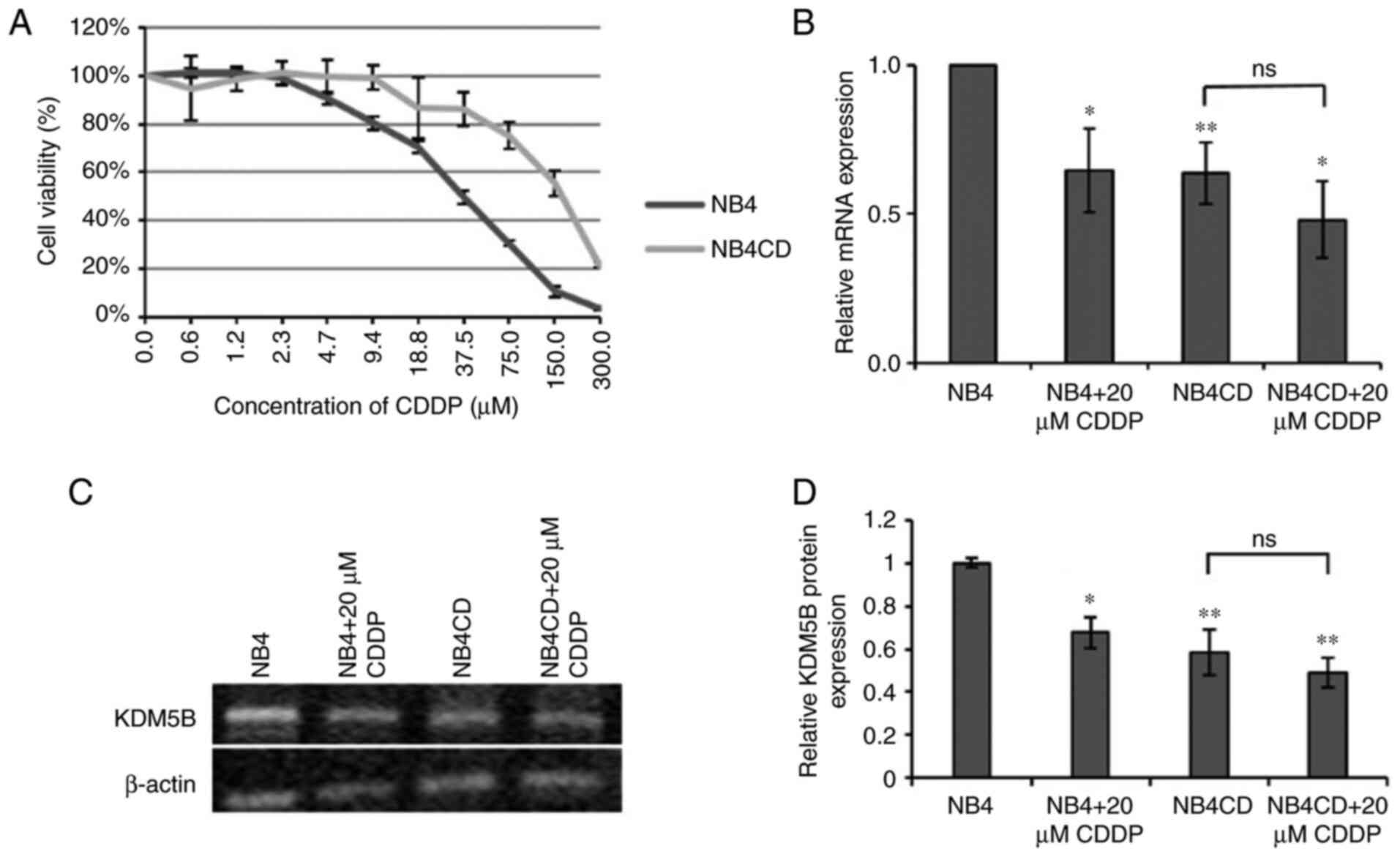
IC50 compared to the parental line UKF-NB-4 (Fig. 1A). We examined the level of KDM5B
mRNA and protein in both cell lines and the expression of this gene
in UKF-NB-4CDDP and in both lines after incubation with
cisplatin was related to the expression in UKF-NB-4 control.
QRT-PCR results showed that KDM5B expression was noticeably lower
in resistant cell line (P<0.01). The same result was observed
after incubation of these cells with CDDP; however, this compound
did not further alter expression in resistant cell line (Fig. 1B). A decrease in the level of KDM5B
expression was also observed in UKF-NB-4CDDP at the protein level
(Fig. 1C, D). Furthermore, we
observed the same results in another NBL cell line UKF-NB-3, where
UKF-NB-3CDDP had lower levels of KDM5B mRNA (P<0.05)
and protein. CDDP did not modulate KDM5B expression (Fig. S1). In SK-N-AS KDM5B level was
decreased by 48 h incubation with CDDP. In SK-N-ASCDDP,
KDM5B was not modulated by cisplatin and there was no significant
difference between sensitive SK-N-AS and resistant
SK-N-ACDDP (Fig. S2).
UKF-NB-4 and UKF-NB-3 cell lines have MYCN amplification, while
SK-N-AS has no MYCN amplification. For further experiments, we
selected UKF-NB-4 and UKF-NB-4CDDP cell lines.
KDM5B knockdown reduced KDM5B
expression and upregulated histone H3K4 trimethylation in
neuroblastoma cells
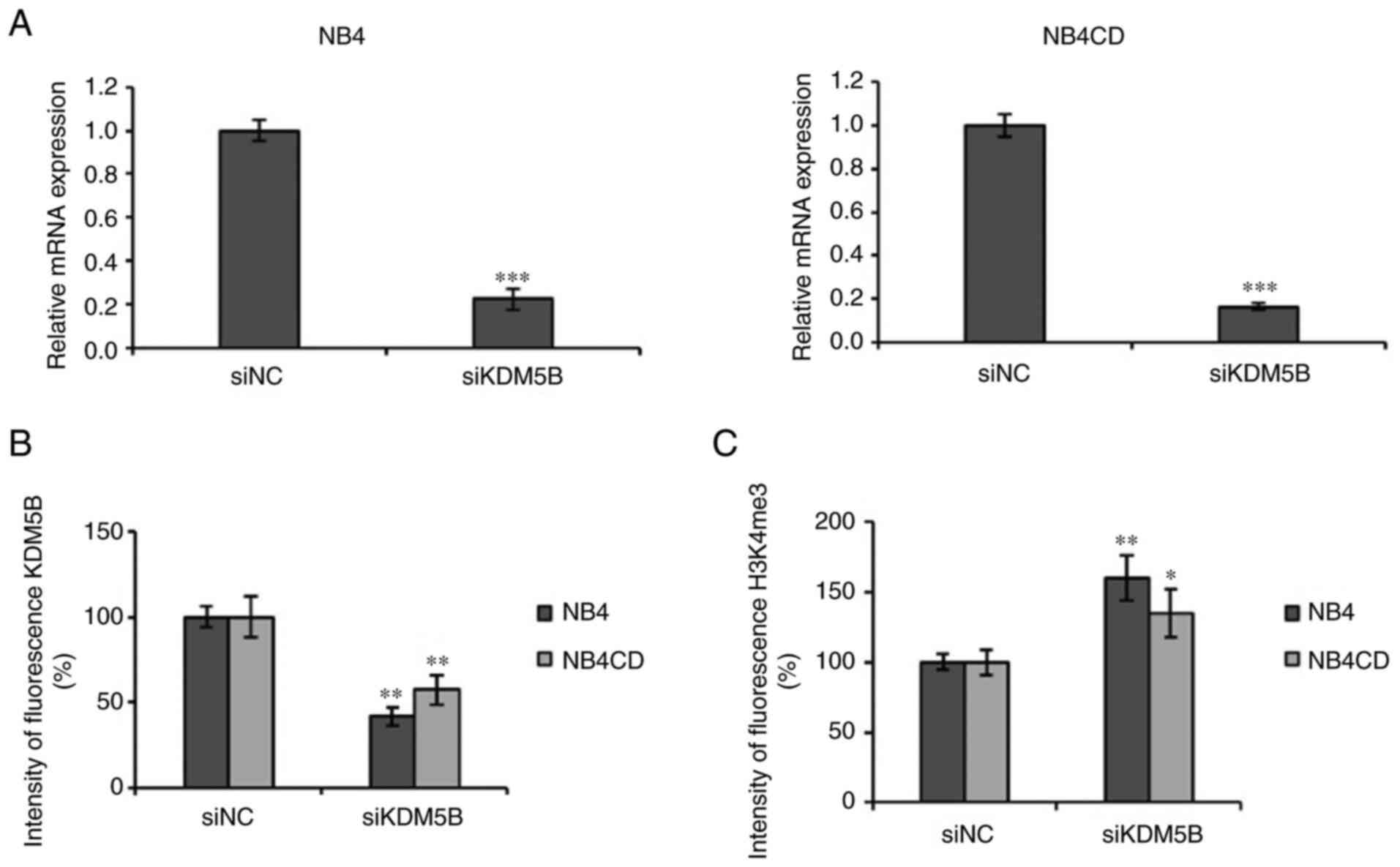
UKF-NB-4 and UKF-NB-4CDDP cells were
transfected with KDM5B siRNA for 48 h and transfection resulted in
a significant suppression of KDM5B level compared to cells
transfected with non-coding siRNA transfected cells (P<0.001)
(Fig. 2A). Flow cytometry was
performed to determine the level of KDM5B protein, which decreased
in both siRNA transfected cell lines (P<0.01), while the
trimethylation of histone H3K4me3 was significantly increased
compared to the control group in UKF-NB-4 (P<0.01) and
UKF-NB-4CDDP (P<0.05) (Fig. 2B,
C). The results demonstrated that KDM5B siRNA reduced KDM5B
mRNA and protein expression and elevated protein H3K4me3 increased
the trimethylation of histone H3K4 in UKF-NB-4 and
UKF-NB-4CDDP cell lines.
KDM5B knockdown promoted cell
proliferation and migration in resistant cell line
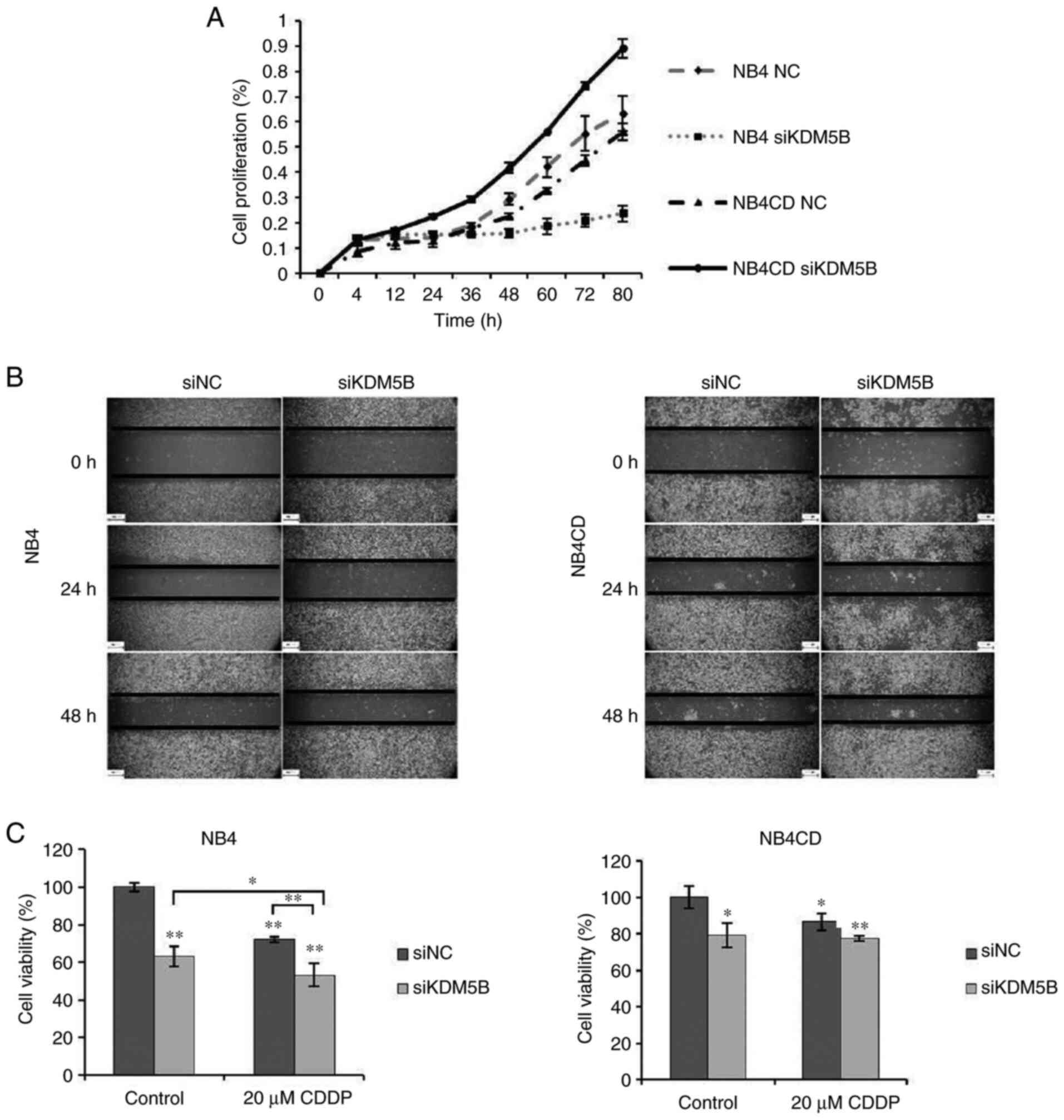
Proliferation of neuroblastoma cells after KDM5B
siRNA transfection was evaluated by xCELLigence system. We found
that KDM5B knockdown inhibited cell proliferation in sensitive cell
line; however, silencing of KDM5B in resistant cells led to
increased proliferation (Fig. 3A).
The wound healing assay showed that down-regulation of KDM5B
promoted the migration of UKF-NB-4CDDP cells compared to
UKF-NB-4 (Fig. 3B). We also
performed a cell viability assay, to see the impact of transfection
on neuroblastoma cell lines. Results show, that KDM5B siRNA reduced
the number of viable cells compared with non-coding siRNA
transfected cells in sensitive cell line more significantly
(P<0.01), than in resistant cell line (P<0.05). Increased
sensitivity to CDDP (48 h treatment of these cells with CDDP) after
silencing of KDM5B in sensitive cell line was observed (P<0.05).
Interestingly, KDM5B knockdown affected neither viability nor
response to CDDP in resistant cells (Fig. 3C).
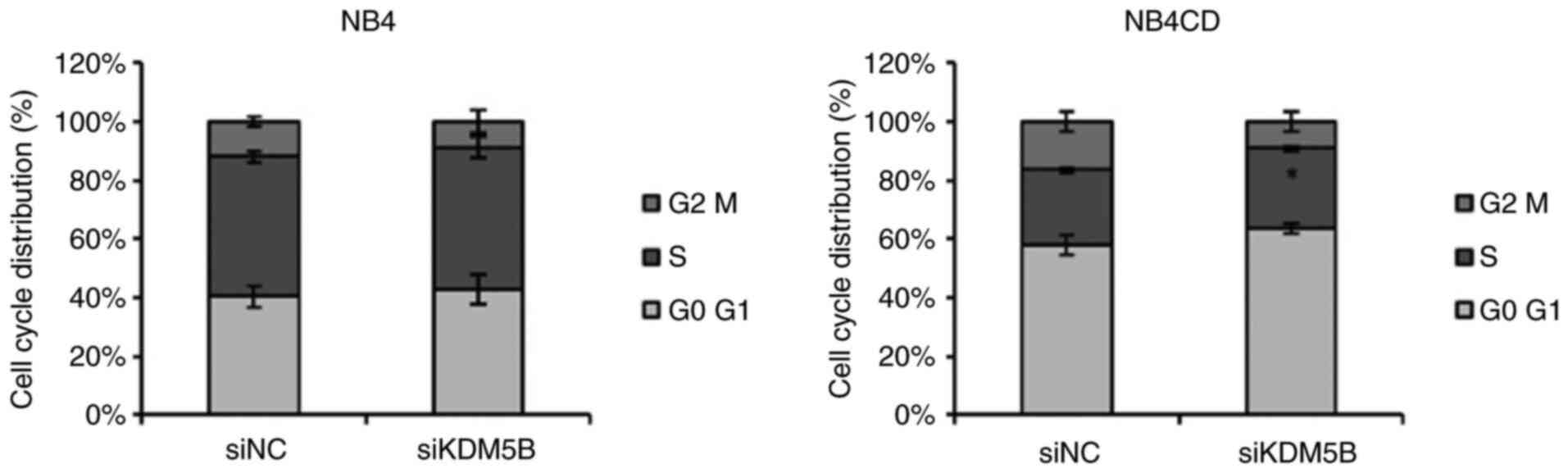
KDM5B knockdown increases cell cycle S
phase in resistant cell line
As shown above, KDM5B downregulation promotes cell
proliferation and migration in resistant NBL cells (Fig. 3). Thus, we explored the role of
KDM5B in cell cycle, using flow cytometry. Consistent with
proliferation and migration data, we found that KDM5B knockdown
resulted in a significant increase in the S phase in UKF-NB-4CDDP
resistant cell line (P<0.05). In the sensitive cell line
UKF-NB-4, silencing did not lead to any significant change in cell
cycle (Fig. 4).
Discussion
Aberrant epigenetic modifications, such as histone
methylation, are widely described as essential players in cancer
development and progression (3,26).
KDM5B, is a histone lysine demethylase, whose dysregulation has
been observed in numerous types of cancers and also has a role in
the appearance of a drug-tolerant population (11,27,28).
Growing evidence indicates that KDM5B act as an oncogene in
numerous types of cancer, such as bladder, breast, lung, prostate,
and ovarian cancer, and also in NBL (29–35).
Since the development of chemoresistance in high-risk NBL is a
negative prognostic marker, we decided to investigate the
importance of KDM5B expression for NBL cell growth and its
chemoresistance to CDDP that is used in high-risk NBL therapy.
In this study, we demonstrated that KDM5B
expression is markedly reduced in NBL cisplatin chemoresistant
cells, compared to parental sensitive cells (Fig. 1) which is associated with enhanced
cell migration and invasion, as well as it may be possibly involved
in drug resistance (Fig. 3).
However, KDM5B silencing did not changed the sensitivity of
resistant cells to CDDP. Several different mechanisms in
chemoresistance that were described in several studies may be
involved and KDM5B expression is only one of those mechanisms. For
example in (25) we described
several different mechanisms of chemoresistance in one cell line
with experimentally induced chemoresistance to ellipticine. These
findings are consistent with the reported data, showing that KDM5B
can play a dual role in cancer (36). Roesch et al found that
KDM5B expression in malignant melanoma, especially in
advanced and metastatic melanoma tissues, was significantly
downregulated and this lysine demethylase has been shown to have
immediate antiproliferative effects, but later has a role in
continuous tumor growth and maintenance (37). Furthermore, the elimination of
KDM5B leads to an initial acceleration of melanoma growth
(28). The MYCN plays a
crucial role in the malignant behavior of NBL and is associated
with a poor prognosis (13,38).
We detected a decrease in the level of KDM5B in
UKF-NB-4CDDP and UKF-NB-3CDDP cisplatin
resistant cell lines that are MYCN amplified while in
SK-N-ASCDDP cell line without MYCN amplification KDM5B
level has not been changed compared to sensitive SK-N-AS cells. We
suppose that this may be caused by increased expression of MYCN in
lines with amplification of this gene (39). Zhang et al suggested that
n-Myc represses KDM5B gene transcription by direct binding
to the Sp1-binding site-enriched region of the KDM5B gene
promoter, most likely through the recruitment of histone
deacetylases (40). This work
showed that the suppression of KDM5B expression reduces NBL
cell proliferation. However, n-Myc induces the proliferation of NBL
cells and represses KDM5B expression, suggesting that
n-Myc-mediated transcriptional repression of KDM5B
counterintuitively reduces tumor cell proliferation (30).
In conclusion, the results of this study show that
KDM5B knockdown leads to increased levels of H3K4me in both
cisplatin sensitive and resistant cell lines (Fig. 2) Based on this finding, it can be
concluded that the function of lysine demethylase KDM5B i.e.
demethylation of di- and tri-methylated histone H3K4 cannot be
fully replaced by the other KDM5 family members in NBL cells. We
proved increased H3K4 me 3 also after silencing of KDM5D (Podhorska
N. unpublished results) and KDM5A and C we did not tested. We
supposed that all KDM5 isoforms are necessary to ensure the
adequate level of H3K4 me3. It can be concluded that the function
of this histone lysine demethylase cannot be fully replaced by the
others KDM5. Also, KDM5B silencing led to an increase of
proliferation, and wound healing assay showed an increase in
migration in resistant cell line. Moreover, in chemoresistant
cells, it only minimally decreased viability after cisplatin
treatment compared to sensitive cells (Fig. 3). Compliant with these results in
resistant cells, siKDM5B transfection resulted in an increase in
cell cycle S phase (Fig. 4). The
effect of KDM5B on cell proliferation and the cell cycle of tumor
cells varies in different tumors. The mechanism of the relationship
between KDM5B and the cell cycle is not yet known, PI3K-AKT pathway
activation (41), BRCA1 (42) and transcription factors E2F1 and
E2F2 (43) are expected to be
affected, but other mechanisms are also possible. In a series of
tumors, its silence inhibits and reduces the percentage of cells in
S phase, for example in prostate cancer (44), hepatocellular carcinoma (43), bladder cancer and small cell lung
carcinoma (33) or acute
lymphoblastic leukemia (42). On
the other hand, in melanoma it has the opposite effect, i.e. the
silencing of KDM5B accelerates growth and increases the proportion
of cells in the S phase (37).
KDM5B transfection induced cell differentiation in hypopharyngeal
squamous cell carcinoma and, on the contrary, it's silencing
accelerated growth of cells (41).
The explanation of different response of sensitive and resistant
NBL cells to KDM5B silencing is not clear and will be subject of
further studies. However, we hypothesized that it is related to the
different expression of this gene in sensitive and resistant NBL
cell lines.
There is emerging evidence for the deregulation of
KDM5B and the important phenotypic consequences in various types of
cancer, making this enzyme a promising factor for the prediction of
sensitivity to CDDP. It will be necessary to study the relationship
between cisplatin sensitivity and histone methylation to understand
resistance to this drug.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This research was funded by the Grant Agency of Charles
University, Czech Republic (grant no. 812217).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TE designed and led this study. MB performed
RT-qPCR, western blotting, flow cytometry and cell proliferation
assays. NP performed siRNA transfection, RT-qPCR, western blotting
and flow cytometry. MB and NP analyzed the data and performed the
statistical analysis. MB and NP wrote the manuscript. AV designed
experiments. MB, NP, AV and TE confirm the authenticity of all the
raw data. All authors have reviewed the manuscript and read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sharma S, Kelly TK and Jones PA:
Epigenetics in cancer. Carcinogenesis. 31:27–36. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dawson MA and Kouzarides T: Cancer
epigenetics: From mechanism to therapy. Cell. 150:12–27. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Greer EL and Shi Y: Histone methylation: A
dynamic mark in health, disease and inheritance. Nat Rev Genet.
13:343–357. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hoffmann I, Roatsch M, Schmitt ML, Carlino
L, Pippel M, Sippl W and Jung M: The role of histone demethylases
in cancer therapy. Mol Oncol. 6:683–703. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li X, Su Y, Pan J, Zhou Z, Song B, Xiong E
and Chen Z: Connexin 26 is down-regulated by KDM5B in the
progression of bladder cancer. Int J Mol Sci. 14:7866–7879. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sun H, Jiang W, Hu J and Ma Z: Prognostic
value of elevated KDM5B expression in patients with laryngeal
squamous cell carcinoma. Int J Clin Exp Pathol. 12:3500–3506.
2019.PubMed/NCBI
|
|
7
|
Bamodu OA, Huang WC, Lee WH, Wu A, Wang
LS, Hsiao M, Yeh CT and Chao TY: Aberrant KDM5B expression promotes
aggressive breast cancer through MALAT1 overexpression and
downregulation of hsa-miR-448. BMC Cancer. 16:1602016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Plch J, Hrabeta J and Eckschlager T: KDM5
demethylases and their role in cancer cell chemoresistance. Int J
Cancer. 144:221–231. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cloos PAC, Christensen J, Agger K and
Helin K: Erasing the methyl mark: Histone demethylases at the
center of cellular differentiation and disease. Genes Dev.
22:1115–1140. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li G, Kanagasabai T, Lu W, Zou MR, Zhang
SM, Celada SI, Izban MG, Liu Q, Lu T, Ballard BR, et al: KDM5B is
essential for the hyperactivation of PI3K/AKT signaling in prostate
tumorigenesis. Cancer Res. 80:4633–4643. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuo YT, Liu YL, Adebayo BO, Shih PH, Lee
WH, Wang LS, Liao YF, Hsu WM, Yeh CT and Lin CM: JARID1B Expression
plays a critical role in chemoresistance and stem cell-like
phenotype of neuroblastoma cells. PLoS One. 10:e01253432015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Maris JM: Recent advances in
neuroblastoma. N Engl J Med. 362:2202–2211. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cheung NK and Dyer MA: Neuroblastoma:
Developmental biology, cancer genomics, and immunotherapy. Nat Rev
Cancer. 13:397–411. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Nara K, Kusafuka T, Yoneda A, Oue T,
Sangkhathat S and Fukuzawa M: Silencing of MYCN by RNA interference
induces growth inhibition, apoptotic activity and cell
differentiation in a neuroblastoma cell line with MYCN
amplification. Int J Oncol. 30:1189–1196. 2007.PubMed/NCBI
|
|
15
|
M. S. MYCN Amplification in Neuroblastoma,
. Brodeur GM, Sawada T and Tsuchida YVP: Neuroblastoma. Elsevier
Science B.V; pp. 75–84. 2000
|
|
16
|
Kang JH, Rychahou PG, Ishola TA, Qiao J,
Evers BM and Chung DH: MYCN silencing induces differentiation and
apoptosis in human neuroblastoma cells. Biochem Biophys Res Commun.
351:192–197. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Maeshima R, Moulding D, Stoker AW and Hart
SL: MYCN Silencing by RNAi induces neurogenesis and suppresses
proliferation in models of neuroblastoma with resistance to
retinoic acid. Nucleic Acid Ther. 30:237–248. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bachetti T, Di Paolo D, Di Lascio S,
Mirisola V, Brignole C, Bellotti M, Caffa I, Ferraris C, Fiore M,
Fornasari D, et al: PHOX2B-mediated regulation of ALK expression:
In vitro identification of a functional relationship between two
genes involved in neuroblastoma. PLoS One. 5:e131082010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Di Paolo D, Pastorino F, Brignole C,
Corrias MV, Emionite L, Cilli M, Tamma R, Priddy L, Amaro A,
Ferrari D, et al: Combined replenishment of miR-34a and let-7b by
targeted nanoparticles inhibits tumor growth in neuroblastoma
preclinical models. Small. 16:e19064262020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Olechnowicz SW, Fedele AO and Peet DJ:
Hypoxic Induction of the Regulator of G-protein signalling 4 gene
is mediated by the hypoxia-inducible factor pathway. PLoS One.
7:e445642012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zaatiti H, Abdallah J, Nasr Z, Khazen G,
Sandler A and Abou-Antoun TJ: Tumorigenic proteins upregulated in
the MYCN-amplified IMR-32 human neuroblastoma cells promote
proliferation and migration. Int J Oncol. 52:787–803.
2018.PubMed/NCBI
|
|
22
|
Cormier N, Yeo A, Fiorentino E and Paxson
J: Optimization of the wound scratch assay to detect changes in
murine mesenchymal stromal cell migration after damage by soluble
cigarette smoke extract. J Vis Exp. e534142015.PubMed/NCBI
|
|
23
|
Pfaffl MW, Horgan GW and Dempfle L:
Relative expression software tool (REST) for group-wise comparison
and statistical analysis of relative expression results in
real-time PCR. Nucleic Acids Res. 30:e362002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bedrnicek J, Vicha A, Jarosova M,
Holzerova M, Cinatl J Jr, Michaelis M, Cinatl J and Eckschlager T:
Characterization of drug-resistant neuroblastoma cell lines by
comparative genomic hybridization. Neoplasma. 52:415–419.
2005.PubMed/NCBI
|
|
25
|
Procházka P, Libra A, Zemanová Z,
Hřebačková J, Poljaková J, Hraběta J, Bunček M, Stiborová M and
Eckschlager T: Mechanisms of ellipticine-mediated resistance in
UKF-NB-4 neuroblastoma cells. Cancer Sci. 103:334–341. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kanwal R and Gupta S: Epigenetic
modifications in cancer. Clin Genet. 81:303–311. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pippa S, Mannironi C, Licursi V, Bombardi
L, Colotti G, Cundari E, Mollica A, Coluccia A, Naccarato V, La
Regina G, et al: Small molecule inhibitors of KDM5 histone
demethylases increase the radiosensitivity of breast cancer cells
overexpressing JARID1B. Molecules. 24:17392019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu W, Zhou B, Zhao X, Zhu L, Xu J, Jiang
Z, Chen D, Wei Q, Han M, Feng L, et al: KDM5B demethylates H3K4 to
recruit XRCC1 and promote chemoresistance. Int J Biol Sci.
14:1122–1132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xiang Y, Zhu Z, Han G, Ye X, Xu B, Peng Z,
Ma Y, Yu Y, Lin H, Chen AP and Chen CD: JARID1B is a histone H3
lysine 4 demethylase up-regulated in prostate cancer. Proc Natl
Acad Sci USA. 104:19226–19231. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Klein BJ, Piao L, Xi Y, Rincon-Arano H,
Rothbart SB, Peng D, Wen H, Larson C, Zhang X, Zheng X, et al: The
histone-H3K4-specific demethylase KDM5B Binds to its substrate and
product through distinct PHD fingers. Cell Rep. 6:325–335. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lu PJ, Sundquist K, Baeckstrom D, Poulsom
R, Hanby A, Meier-Ewert S, Jones T, Mitchell M, Pitha-Rowe P,
Freemont P and Taylor-Papadimitriou J: A novel gene (PLU-1)
containing highly conserved putative DNA/chromatin binding motifs
is specifically up-regulated in breast cancer. J Biol Chem.
274:15633–15645. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yamane K, Tateishi K, Klose RJ, Fang J,
Fabrizio LA, Erdjument-Bromage H, Taylor-Papadimitriou J, Tempst P
and Zhang Y: PLU-1 Is an H3K4 demethylase involved in
transcriptional repression and breast cancer cell proliferation.
Mol Cell. 25:801–812. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hayami S, Yoshimatsu M, Veerakumarasivam
A, Unoki M, Iwai Y, Tsunoda T, Field HI, Kelly JD, Neal DE, Yamaue
H, et al: Overexpression of the JmjC histone demethylase KDM5B in
human carcinogenesis: Involvement in the proliferation of cancer
cells through the E2F/RB pathway. Mol Cancer. 9:592010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yamamoto S, Wu Z, Russnes HG, Takagi S,
Peluffo G, Vaske C, Zhao X, Moen Vollan HK, Maruyama R, Ekram MB,
et al: JARID1B is a luminal lineage-driving oncogene in breast
cancer. Cancer Cell. 25:762–777. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang L, Mao Y, Du G, He C and Han S:
Overexpression of JARID1B is associated with poor prognosis and
chemotherapy resistance in epithelial ovarian cancer. Tumour Biol.
36:2465–2472. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Roesch A, Becker B, Meyer S, Wild P,
Hafner C, Landthaler M and Vogt T: Retinoblastoma-binding protein
2-homolog 1: A retinoblastoma-binding protein downregulated in
malignant melanomas. Mod Pathol. 18:1249–1257. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Roesch A, Fukunaga-Kalabis M, Schmidt EC,
Zabierowski SE, Brafford PA, Vultur A, Basu D, Gimotty P, Vogt T
and Herlyn M: A Temporarily distinct subpopulation of slow-cycling
melanoma cells is required for continuous tumor growth. Cell.
141:583–594. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Smith V and Foster J: High-risk
neuroblastoma treatment review. Children (Basel).
5:1142018.PubMed/NCBI
|
|
39
|
Feriancikova B, Feglarova T, Krskova L,
Eckschlager T, Vicha A and Hrabeta J: Miat is an upstream regulator
of NMYC and the disruption of the MIAT/NMYC axis induces cell death
in NMYC amplified neuroblastoma cell lines. Int J Mol Sci.
22:33932021. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang L, Sokolowski N, Atmadibrata B and
Tao L: Histone demethylase JARID1B promotes cell proliferation but
is downregulated by N-Myc oncoprotein. Oncol Rep. 31:1935–1939.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang J, An X, Han Y, Ma R, Yang K, Zhang
L, Chi J, Li W, Llobet-Navas D, Xu Y and Jiang Y: Overexpression of
JARID1B promotes differentiation via SHIP1/AKT signaling in human
hypopharyngeal squamous cell carcinoma. Cell Death Dis.
7:e23582016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
YQ H, Y Z, RJ Z and XD M: Down-regulation
of JARID1B expression inhibits cell proliferation, induces
apoptosis and blocks cell cycle in human acute lymphoblastic
leukemia cells. Eur Rev Med Pharmacol Sci. 22:1366–1373.
2018.PubMed/NCBI
|
|
43
|
Shigekawa Y, Hayami S, Ueno M, Miyamoto A,
Suzaki N, Kawai M, Hirono S, Okada KI, Hamamoto R and Yamaue H:
Overexpression of KDM5B/JARID1B is associated with poor prognosis
in hepatocellular carcinoma. Oncotarget. 9:34320–34335. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yang Z, Xu JX, Fang DP and Ke J: Analysis
of key genes reveal lysine demethylase 5B promotes prostate cancer
progression. Oncol Lett. 20:622020.PubMed/NCBI
|