Introduction
Thyroid cancer is the most commonly diagnosed
endocrine cancer and mainly originates from follicular epithelial
cells (1). The incidence of
thyroid cancer increased by 3.6% per year on average during
1974–2013 (from 4.56 per 100,000 person-years in 1974–1977 to 14.42
per 100,000 person-years in 2010–2013 (2). According to histological
classification, papillary thyroid carcinoma, which accounts for
~80% of cases, and follicular thyroid carcinoma, which accounts for
~10% of cases, are the two major types of thyroid cancer (3). Treated with specific methods such as
surgery (4), radioiodine (5), and thyroid-stimulating hormone
suppression (6), numerous patients
with solid cancers exerted a relatively optimal improvement in
their conditions. However, there is also recurrence and metastasis
of tumor, which lead to death in certain cases (7). Thus, it is of great importance to
conduct an in-depth exploration into the molecular mechanisms of
thyroid cancer, to provide significant insights into the
improvements of therapy.
MicroRNAs (miRNAs or miRs), which locate in the
non-coding region of the genome and are highly conserved in
evolution, are a family of non-coding single-stranded small RNAs
with the length of 19–23 nucleotides (nt) and regulate gene
expression at translation level (8). miRNAs participate in multiple
physiological metabolic processes, including development, cell
differentiation and cell apoptosis (9). With the advancement in researches on
molecular biology and molecular genetics, miRNAs have been
indicated to be related with the development, and metastasis of
tumors, including thyroid cancer (10).
Long non-coding RNAs (lncRNAs) are defined as
molecules consisting of >200 nt with no protein-coding potential
(11). Evidence indicates that
lncRNAs have a critical regulatory function in eukaryotic cells
(12,13). Moreover, it has been reported that
lncRNAs interact with key proteins and sponge miRNAs to regulate
cell proliferation, differentiation, motility and metabolism
(14–16). The dysregulation of lncRNAs is
involved in the pathogenesis of numerous types of cancer. For
example, mini-chromosome maintenance complex component 3 associated
protein (MCM3AP)-antisense RNA 1 (AS1) is upregulated in thyroid
cancer. This lncRNA promotes thyroid cancer cell proliferation and
invasion via acting as an miR-211-5p sponge and elevating secreted
protein acidic and cysteine rich (SPARC) protein expression levels
(17). Moreover, miR-592 inhibits
the progression of thyroid cancer via the regulation of lncRNA
nuclear paraspeckle assembly transcript 1 and the downregulation of
NOVA alternative splicing regulator 1 (18). Furthermore, the hypoxia-induced
lncRNA-AC020978 induces cell proliferation of non-small cell lung
cancer via the regulation of the pyruvate kinase
M2/hypoxia-inducible factor-1α axis (19). Although the function of several
abnormally expressed lncRNAs has been reported, the roles of
numerous lncRNAs in cancer remain elusive. In 2018, a novel lncRNA,
that is, EGF like and EMI domain containing 1 (EGFEM1P) was found
to be associated with thyroid cancer in the study by You et
al (20); however, since its
role in thyroid cancer development has not been discovered, the
present study aimed to explore it.
Materials and methods
Tissue samples
From September 2017 to June 2020, thyroid tumor
tissues (n=35) and normal adjacent tissues (n=24) were collected
from 35 patients (aged from 41 to 72 years old) diagnosed with
papillary thyroid cancer during surgery in The Central Hospital of
Enshi Tujia and Miao Autonomous Prefecture (Enshi, China). The
clinicopathological characteristics of the patients were recorded
in Table I. Patients did not
receive radiotherapy or other treatments before enrollment in the
present study. Written informed consent was obtained from all
patients. The present study was approved (approval no.
CHESTJMAP-2017-08) by the Ethics Committee of The Central Hospital
of Enshi Tujia and Miao Autonomous Prefecture (Enshi, China). The
tissues were stored at −80°C before being used in the following
experiments.
 | Table I.The clinicopathological
characteristics of the patients with papillary thyroid cancer. |
Table I.
The clinicopathological
characteristics of the patients with papillary thyroid cancer.
| Clinicopathological
factors | Number of patients
(%) |
|---|
| Age, years |
|
|
≤55 | 17 (48.57) |
|
>55 | 18 (51.43) |
| Sex |
|
|
Male | 15 (42.86) |
|
Female | 20 (57.14) |
| TNM staging |
|
|
I–II | 25 (71.43) |
|
III–IV | 10 (28.57) |
| Tumor size, cm |
|
| ≤2 | 22 (62.86) |
|
>2 | 13 (37.14) |
| Lymph node
metastasis |
|
|
Yes | 21 (60.00) |
| No | 14 (40.00) |
| Distant
metastasis |
|
|
Yes | 11 (31.43) |
| No | 24 (68.57) |
| Extrathyroidal
extension |
|
|
Yes | 19 (54.29) |
| No | 16 (45.71) |
Cell culture
The human normal thyroid epithelial Nthy-ori 3–1
cell line was purchased from the European Collection of
Authenticated Cell Cultures. The two papillary thyroid carcinoma
cell lines (TPC-1 and KTC-1) and one follicular thyroid carcinoma
cell line (FTC-133) were purchased from the American Type Culture
Collection. Cells were maintained in RPMI-1640 medium (Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) and 1% penicillin-streptomycin solution
(Invitrogen; Thermo Fisher Scientific, Inc.). Cells were cultured
in a humid incubator with 5% CO2 at 37°C.
Bioinformatic analysis
To screen thyroid cancer-related lncRNAs, the gene
expression data of papillary thyroid tumor tissues and normal
adjacent tissues from the GSE66783 dataset (normal adjacent thyroid
tissues, 5; papillary thyroid carcinoma tissues, 5) (21) and The Cancer Genome Atlas (TCGA;
solid tumor normal tissues, 54; primary tumors, 478; metastatic
tumors, 8. Tissues were not derived from the same patients)
(22) were used. The
differentially expressed genes from these two datasets were
overlapped and EGF-like and EMI domain-containing protein 1
(EGFEM1P) was identified as one of the most significantly
upregulated lncRNAs. The patients were divided into two groups
according to high and low EGFEM1P expression levels. Furthermore,
Kaplan-Meier analysis was performed to investigate the association
between EGFEM1P expression levels and the prognosis of patients.
The target miRNAs of EGFEM1P were predicted using miRDB (http://mirdb.org/) (23). The target mRNAs of miR-369-3p were
also predicted using miRDB (23),
TargetScan (http://www.targetscan.org/vert_72/) (24) and miRWalk (http://mirwalk.umm.uni-heidelberg.de/) (25).
Transfection
Control small interfering (si)RNA
(5′-UUCUCCGAACGUGUCACGUU-3′), si-EGFEM1P-1
(5′-GCGGCGAGCGCGUUUCCAUGG-3′) and siEGFEM1P-2
(5′-GGAGGGCGGCGAGCGCGUUUC-3′) were designed and synthesized by
Suzhou GenePharma Co., Ltd. miR-negative control (NC) mimic
(5′-CUGAACUGCUAGGACGCGUA-3′), miR-NC inhibitor
(5′-CUCGAUAGCGCAUGGUCCGAGCUA-3′), miR-369-3p mimic
(5′-AAUAAUACAUGGUUGAUCUUU-3′) and miR-369-3p inhibitor
(5′-AAAGAUCAACCAUGUAUUAUU-3′) were synthesized by Guangzhou RiboBio
Co., Ltd. siRNA (100 nM), miR-369-3p mimic (40 nM) or miR-369-3p
inhibitor (40 nM) or the corresponding controls was transfected
into KTC-1 cells using Lipofectamine® RNAiMAX
Transfection Reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
at 37°C for 48 h according to the manufacturer's protocol. The
transfection efficiencies of the siRNA, miR-369-3p mimic and
miR-369-3p inhibitor constructs were determined using reverse
transcription-quantitative PCR (RT-qPCR) 48 h after
transfection.
Cell proliferation assay
Cell proliferation of KTC-1 cells
(1×105/well) was analyzed using the Cell Counting Kit-8
(CCK-8; Dojindo Molecular Technologies, Inc.) assay. In brief, the
cell medium was replaced with medium containing 10% CCK-8 solution
and cells were incubated for 2 h at 37°C. The absorbance was
detected at 450 nm using a Microplate Reader (Bio-Rad Laboratories,
Inc.).
Cell apoptosis assay
The percentage of apoptotic KTC-1 cells was
determined using the Dead Cell Apoptosis Kit with an Annexin V FITC
and PI kit (Invitrogen; Thermo Fisher Scientific, Inc.). In brief,
cells were collected and suspended in Annexin V binding buffer
containing PI. Annexin V-FITC was then added and cells were
incubated for 15 min at room temperature. Subsequently the cells
were subjected to flow cytometric analysis. The apoptotic cells
were estimated by a FACSCalibur flow cytometer (BD Biosciences).
The data were analyzed using FlowJo software 10.7 (BD Biosciences).
Apoptotic cells were identified as being positive for Annexin V
and/or PI.
RT-qPCR
Total RNA was extracted from cells and tissues using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). Complementary DNA was synthesized using the PrimeScript™ RT
Reagent kit (Takara Bio, Inc.) according to the manufacturer's
instructions. qPCR was performed using TB Green Advantage qPCR
Premix (Takara Bio, Inc.). The thermocycling conditions were as
listed: Initial denaturation at 95°C for 1 min; followed with 42
cycles of denaturation at 95°C for 15 sec, annealing at 60°C for 31
sec and elongation at 72°C for 30 sec, and final extension at 72°C
for 5 min. Expression levels were normalized to GAPDH and U6 for
mRNA and miRNA, respectively. Expression levels were analyzed using
the 2−ΔΔCq method (26). The primer sequences were listed in
Table II.
| Table II.Primer sequences. |
Table II.
Primer sequences.
| Primer name | Primer sequence
(5′-3′) |
|---|
| EGFEM1P | F:
ATATTGTATTATAATTAGTT |
|
| R:
AACTAATTATAATACAATAT |
| miR-30d-3p | F:
CTTTCAGTCAGATGTTTGCTGC |
|
| R:
GCAGCAAACATCTGACTGAAAG |
| miR-30e-3p | F:
CTTTCAGTCGGATGTTTACAGC |
|
| R:
GCTGTAAACATCCGACTGAAAG |
| miR-369-3p | F:
AATAATACATGGTTGATCTTT |
|
| R:
AAAGATCAACCATGTATTATT |
| TCF4 | F:
CTTAACAGCTGTATTATCTTAAACCCA |
|
| R:
TGGGTTTAAGATAATACAGCTGTTAAG |
| GAPDH | F:
GCACCGTCAAGGCTGAGAAC |
|
| R:
TGGTGAAGACGCCAGTGGA |
| U6 | F:
GTGCTCGCTTCGGCAGCACAT |
|
| R:
AATATGGAACGCTTCACGAAT |
Western blot analysis
KTC-1 cells were lysed by radioimmunoprecipitation
assay buffer (Beyotime Institute of Biotechnology), then the
bicinchoninic acid assay (Beyotime Institute of Biotechnology) was
conducted to assess protein concentration. Protein lysates (20
µg/lane) were separated by 10% SDS-PAGE, then electrophoretically
transferred onto PVDF membranes, which were blocked by 5% BSA
(Beijing Solarbio Science & Technology Co., Ltd.) in
Tris-buffered saline (TBS)-0.5% Tween-20 at room temperature for 1
h. Subsequently, proteins were incubated sequentially with
antibodies from Cell Signaling Technology, Inc. against GAPDH (cat.
no. 5174; dilution 1:1,000), cleaved caspase-3 (cat. no. 9664;
dilution 1:1,000), caspase-3 (cat. no. sc-7272; dilution 1:1,000),
cleaved caspase-8 (cat. no. 9496; dilution 1:1,000), caspase-8
(cat. no. sc-56070; dilution 1:1,000), cleaved PARP (cat. no. 5625;
dilution 1:1,000), PARP (cat. no. sc-136208; dilution 1:1,000), BAX
(cat. no. 2772; dilution 1:1,000), and TCF4 (cat. no. 2569;
dilution 1:1,000) for 24 h at 4°C, followed by anti-rabbit (cat.
no. 7074; dilution 1:2,000) horseradish peroxidase (HRP)-conjugated
secondary antibody for 2 h at room temperature. Enhanced
chemiluminescence detection system for HRP (EMD Millipore) was
used. Proteins were analyzed with a LAS-4000 luminescence image
analyzer (FUJIFILM Wako Pure Chemical Corporation). The band
intensities were analyzed using Image Lab™ (version 4.0; Bio-Rad
Laboratories, Inc.).
Dual-luciferase reporter assay
EGFEM1P wild-type (WT), EGFEM1P truncate (1–3,000
nt), EGFEM1P mutant (Mut), TCF4 3′untranslated region (UTR) WT and
TCF4 3′UTR Mut were inserted into the pmirGLO plasmid (Promega
Corporation). pmirGLO-EGFEM1P, the truncated form, or Mut form were
co-transfected with miR-NC mimic or miR-369-3p mimic into KTC-1
cells using Lipofectamine 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). To confirm the interaction between TCF4 and
miR-369-3p, TCF4 3′UTR WT or TCF4 3′UTR Mut were co-transfected
with miR-NC mimic or miR-369-3p mimic into TPC-1 and KTC-1 cells
using Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific,
Inc.). After incubation at 37°C for 48 h, the luciferase activity
of each group was determined using the Dual-Luciferase Reporter
Assay System (Promega Corporation) according to the manufacturer's
protocol. The method of normalization was comparison with
Renilla luciferase activity.
Statistical analysis
All data were analyzed using GraphPad Prism 5.0
software (GraphPad Software, Inc.) and are presented as the mean ±
SD. All experiments were repeated three times. Student's t-test
(paired for tissues, unpaired for cell lines) was used for
statistical comparisons between two groups. More than two groups
(including differentially expressed genes from TCGA) were
statistically compared using one-way ANOVA followed by Tukey's post
hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
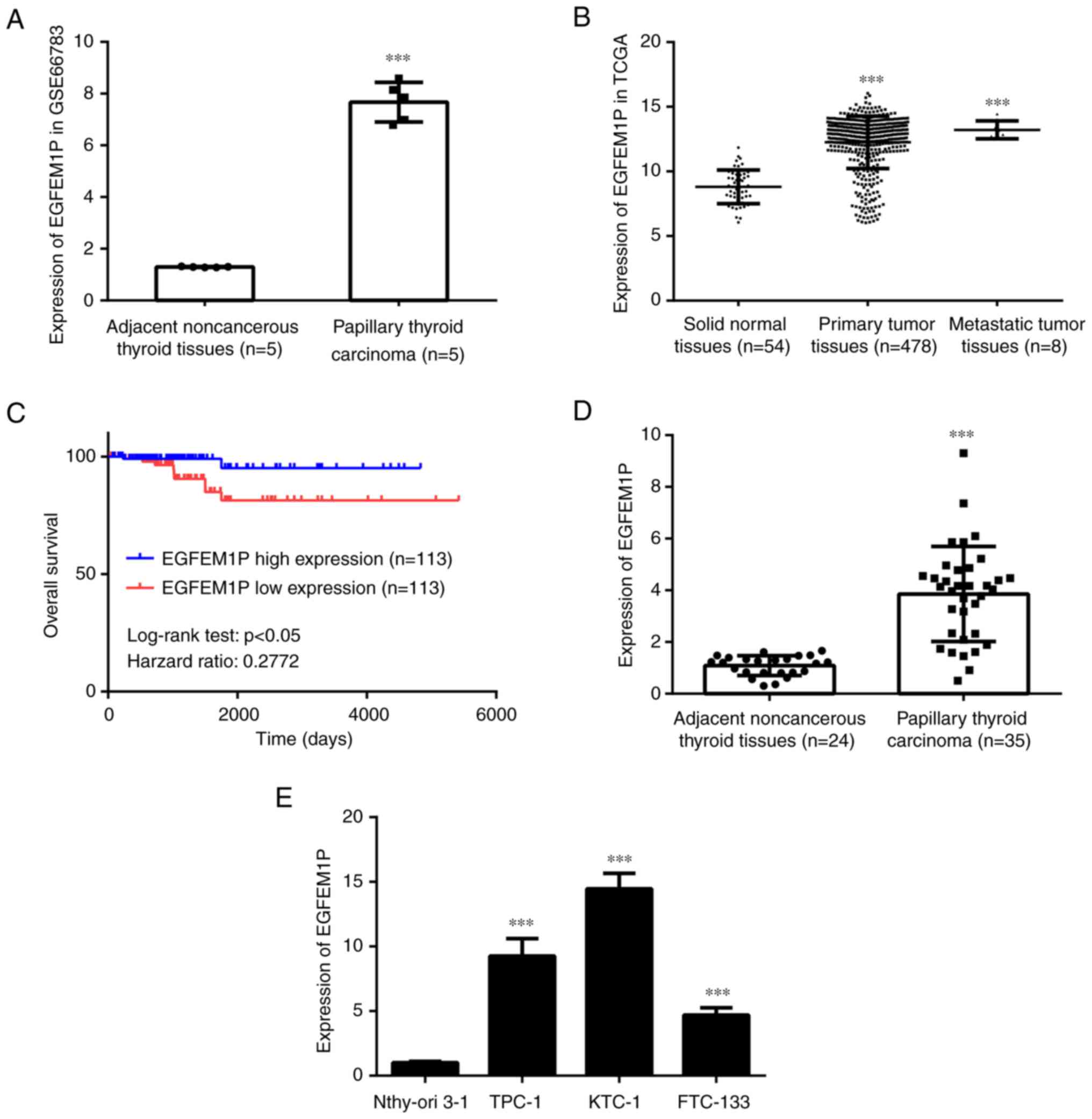
EGFEM1P is upregulated in thyroid
cancer tissues and cell lines
According to the data from GSE66783, EGFEM1P was
significantly increased in papillary thyroid cancer tissues
compared with normal adjacent thyroid tissues (Fig. 1A). Furthermore, the data from the
TCGA database demonstrated that EGFEM1P was also significantly
increased in primary tumor tissues and metastatic tumor tissues
compared with normal adjacent tissues (Fig. 1B). Kaplan-Meier analysis
demonstrated a positive association between EGFEM1P overexpression
and the poor prognosis of patients with thyroid cancer (Fig. 1C). In the collected samples from
the present study, EGFEM1P expression levels were significantly
increased in papillary thyroid cancer tissues compared with the
normal adjacent thyroid tissues (Fig.
1D). Moreover, compared with Nthy-ori 3–1 cells, EGFEM1P
expression levels were significantly increased in TPC-1, KTC-1 and
FTC-133 cells (Fig. 1E). As KTC-1
cells exhibited relatively higher EGFEM1P expression levels, this
cell line was used in the subsequent experiments.
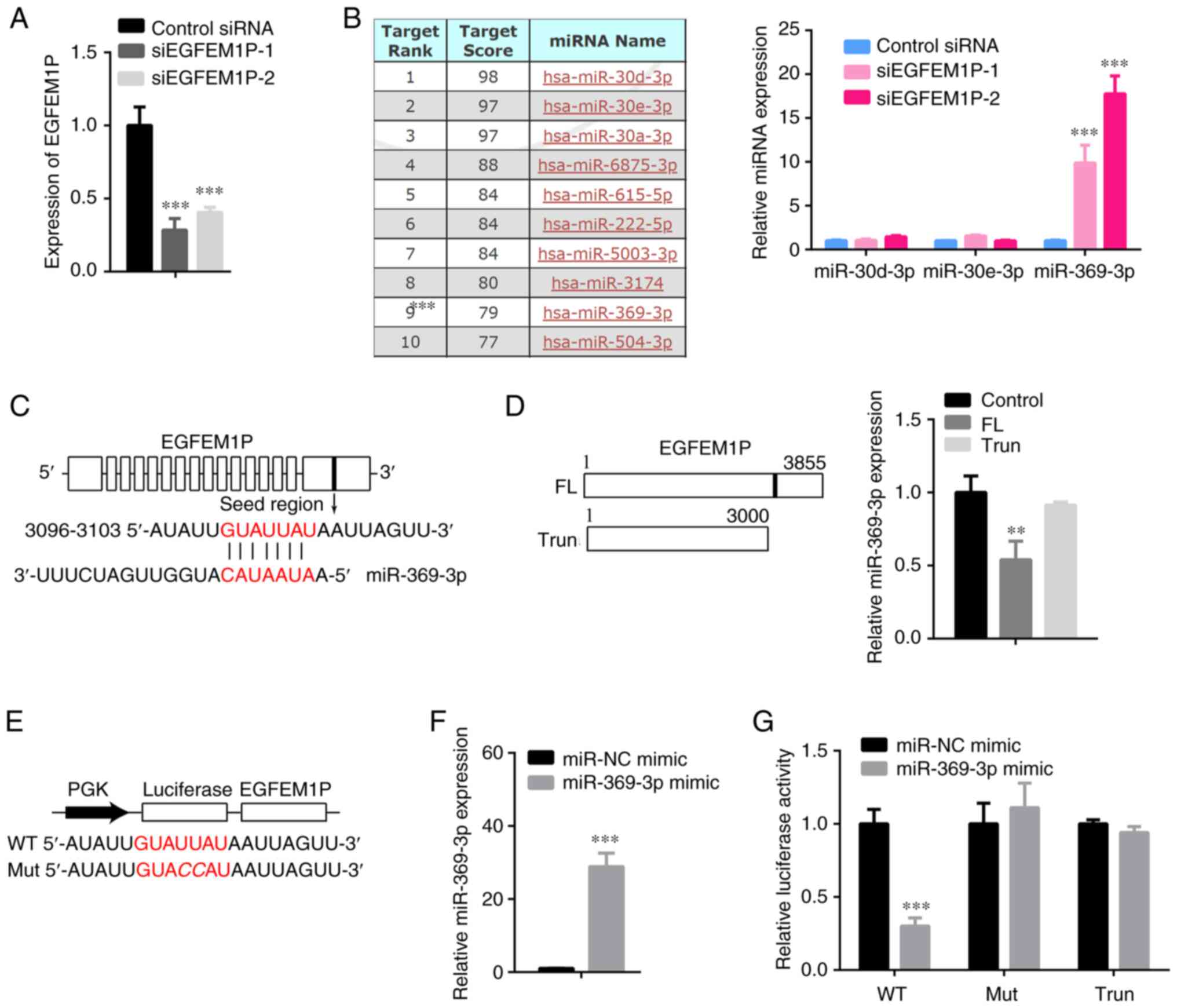
miR-369-3p is sponged by EGFEM1P
Subsequently, the miRNA targets of EGFEM1P were
explored. Prior to this investigation siRNAs were used to knockdown
EGFEM1P expression levels. Both siEGFEM1P-1 and siEGFEM1P-2 were
determined to effectively reduce EGFEM1P expression levels
(Fig. 2A). However, siEGFEM1P-1
was demonstrated to be more efficient and was therefore used for
the subsequent experiments.
Several target miRNAs of EGFEM1P were identified,
including miR-30d-3p, miR-30e-3p, miR-30a-3p, miR-6875-3p,
miR-615-5p, miR-222-5p, miR-5003-3p, miR-3174, miR-369-3p and
miR-504-3p. Moreover, a number of miRNAs were randomly selected to
examine their interaction with EGFEM1P. The results demonstrated
that compared with control siRNA, siEGFEM1P-1 and siEGFEM1P-2
significantly increased miR-369-3p expression levels but not those
of other miRNAs in KTC-1 cells (Fig.
2B). The complementary sites between EGFEM1P and miR-369-3p are
presented in Fig. 2C. Furthermore,
EGFEM1P significantly decreased the expression levels of miR-369-3p
in KTC-1 cells (Fig. 2D). The WT
and Mut sequences of EGFEM1P complementary sites for miR-369-3p are
presented in Fig. 2E. Compared
with miR-NC mimic, miR-369-3p was demonstrated to be increased by
miR-369-3p mimic transfection in KTC-1 cells (Fig. 2F). Subsequently, miR-369-3p mimic
was demonstrated to reduce luciferase activity in KTC-1 cells
transfected with WT-EGFEM1P compared with cells transfected with
the miR-NC mimic (Fig. 2G).
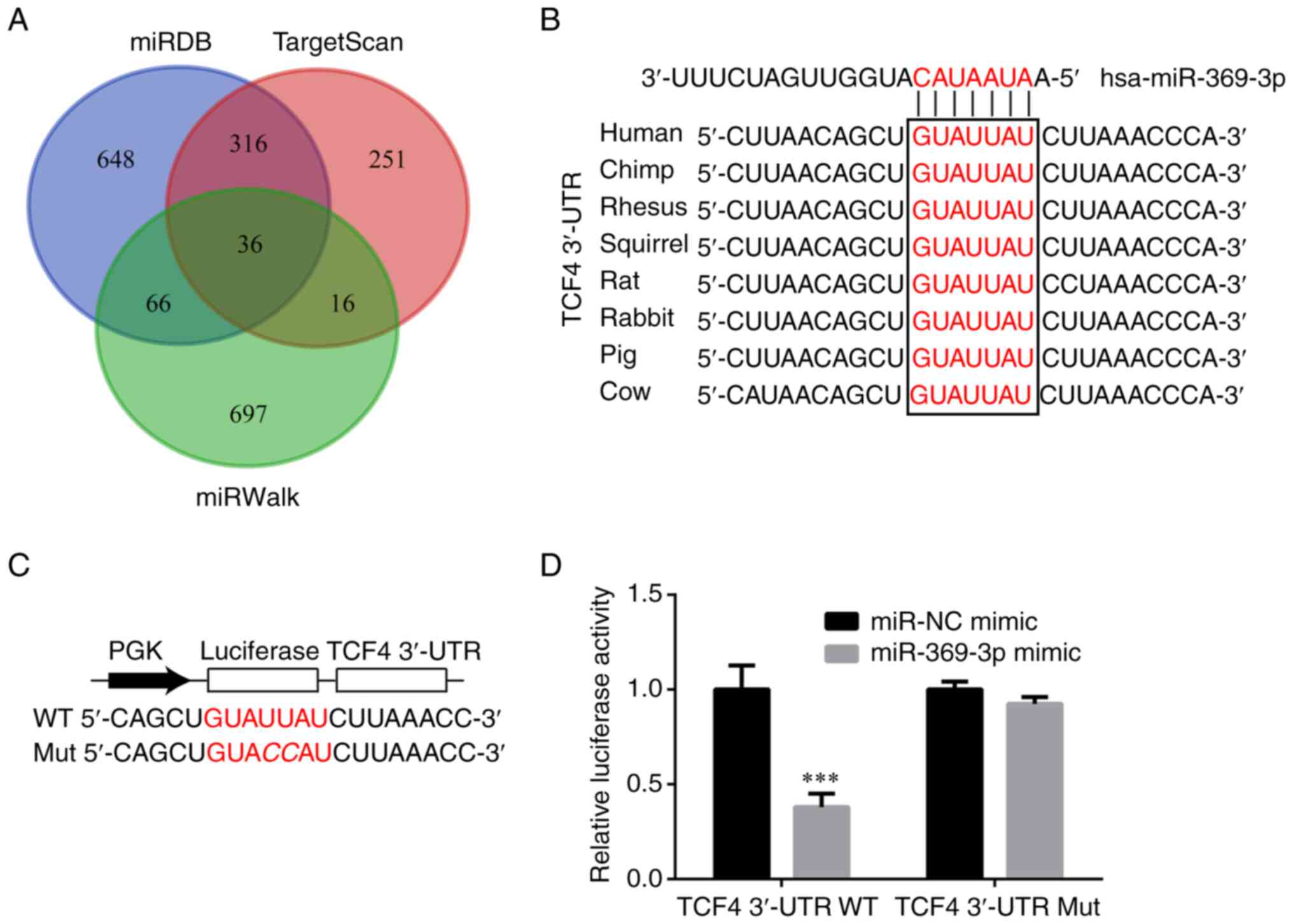
TCF4 is targeted by miR-369-3p
The target genes of miR-369-3p were predicted using
miRDB (23), TargetScan (24) and miRWalk (25) and are presented in Fig. 3A. These results demonstrated that
among the potential target genes identified 36 mRNAs were
overlapped. Of these overlapping mRNAs, which were complementary to
miR-369-3p, the TCF4 3′UTR was conserved among numerous species
(Fig. 3B) and therefore became a
focus of the present study. Subsequently, luciferase vectors were
constructed containing WT- and Mut-TCF4 3′UTR (Fig. 3C). The results demonstrated that
compared with miR-NC mimic, miR-369-3p mimic reduced luciferase
activity in KTC-1 cells, which were transfected with WT-TCF4
(Fig. 3D).
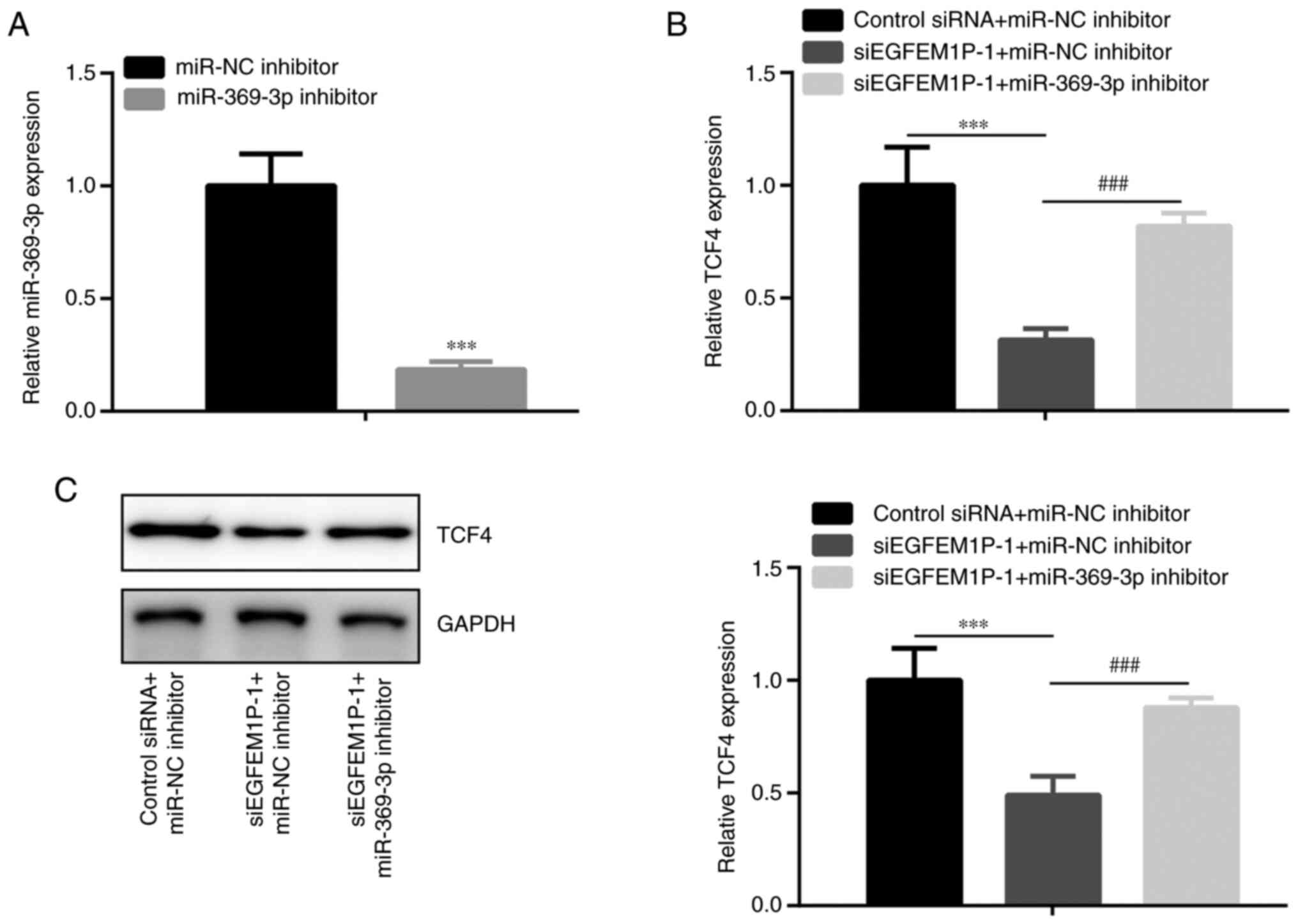
EGFEM1P induces TCF4 expression via
sponging miR-369-3p
Subsequently the effects of EGFEM1P on TCF4
expression levels were assessed. The results demonstrated that in
KTC-1 cells, miR-369-3p was decreased by miR-369-3p inhibitor
compared with miR-NC inhibitor (Fig.
4A). Moreover, in KTC-1 cells, TCF4 mRNA and protein expression
levels were observed to be decreased by siEGFEM1P-1 compared with
the control siRNA + miR-NC inhibitor group. This effect was rescued
via co-transfection with siEGFEM1P-1 + miR-369-3p inhibitor
(Fig. 4B and C).
EGFEM1P induces cell proliferation and
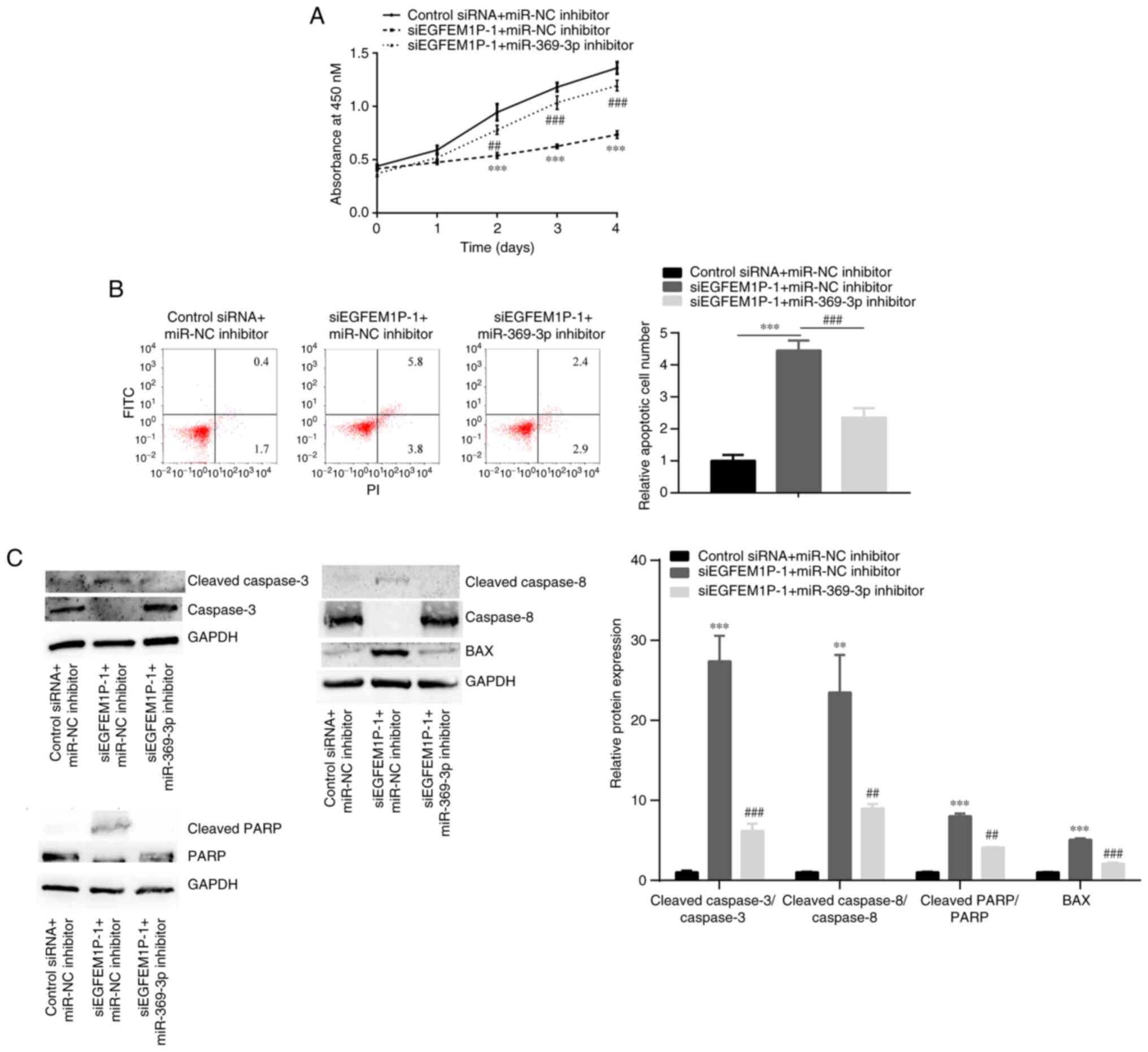
reduces cell apoptosis via sponging miR-369-3p
The relationship between EGFEM1P and miR-369-3p in
cell proliferation and cell apoptosis was investigated. In KTC-1
cells, compared with the control siRNA + miR-NC inhibitor group,
siEGFEM1P-1 significantly suppressed cell proliferation (Fig. 5A); however, siEGFEM1P-1
significantly promoted apoptosis and BAX, cleaved
caspase-3/caspase-3, cleaved caspase-8/ caspase-8, cleaved
caspase-PARP/PARP ratio (Fig. 5B and
C). These aforementioned effects were abolished by the
co-transfection of siEGFEM1P-1 + miR-369-3p inhibitor.
Discussion
lncRNAs serve an important regulatory role in
eukaryotic cells (12,13). Dysregulation of lncRNAs is involved
in the pathogenesis of thyroid cancer. For example, MCM3AP-AS1
promotes cell proliferation and invasion of PTC cells by sponging
the miR-211-5p/SPARC axis (17).
Furthermore, lncRNA solute carrier family 26 member 4-AS1 inhibits
metastasis in thyroid cancer by destabilizing DEAD-box helicase 5
(4). Moreover, lncRNA
metastasis-associated lung adenocarcinoma transcript 1 promotes the
cell proliferation, migration and invasion of thyroid cancer cells
via the miR-204/insulin-like growth factor 2 mRNA binding protein
2/m6A-MYC signaling pathway (27).
However, even though the functions of several abnormally expressed
lncRNAs have been reported, the roles of numerous abnormally
expressed lncRNAs in cancer remain to be elucidated.
In the present study the comprehensive analysis of
microarray and RNA sequencing data of thyroid tumors identified
EGFEM1P as one of the most significantly upregulated lncRNAs in
papillary thyroid tumors. Kaplan-Meier analysis demonstrated that
there was a positive association between EGFEM1P overexpression and
a poor prognosis in patients with thyroid cancer. RT-qPCR was
performed, which confirmed the upregulation of EGFEM1P in papillary
thyroid tumors as well as papillary thyroid carcinoma and
follicular thyroid carcinoma cell lines. The findings were in
consistent with a previous study in 2018, which identified EGFEM1P
as an upregulated lncRNA in papillary thyroid cancer tumors, and
was positively associated with the TNM staging and lymphatic
metastasis of thyroid cancer (20). However, EGFEM1P was previously
observed to be a downregulated in and act as a tumor suppressor in
human lung adenocarcinoma. This difference between this previous
study and the present study could be attributed to the different
characteristics of various types of cancer (28). Subsequently, in the present study
the miRNA targets for EGFEM1P in thyroid cancer were explored.
Current study further investigated the role of EGFEM1P in the
progression of thyroid cancer.
Using miRDB (23),
several target miRNAs of EGFEM1P were predicted, including
miR-30d-3p, miR-30e-3p, miR-30a-3p, miR-6875-3p, miR-615-5p,
miR-222-5p, miR-5003-3p, miR-3174, miR-369-3p and miR-504-3p. The
results demonstrated that EGFEM1P interacted with miR-369-3p and
decreased miR-369-3p expression levels, which demonstrated the
potential tumor suppressive role of miR-369-3p in thyroid cancer.
This result was consistent with previous studies, which also
reported the tumor suppressive role of miR-369-3p in several types
of cancer. For example, miR-369-3p inhibits the cell viability and
motility of hepatocellular carcinoma by binding to paired box 6
(29), miR-369-3p overexpression
inhibits cell proliferation and migration of endometrioid
adenocarcinoma (30), miR-369-3p
was reduced in the inflamed intestinal regions of patients with
inflammatory bowel disease, overexpression of miR-369-3p alleviated
LPS-induced inflammation (31),
miR-369-3p serves as a tumor suppressor in gastric cancer (32). Moreover, the overexpression of
miR-369-3p inhibits cell proliferation and induces cell apoptosis
of papillary thyroid cancer cells (33).
MiRDB (23),
TargetScan (24) and miRWalk
(25) were used to predict the
target genes of miR-369-3p, demonstrating that 36 mRNAs were
overlapped, of which, the TCF4 3′UTR was conserved among numerous
species and therefore became a focus of the present study.
Subsequently it was verified in the present study that TCF4 acted
as a target gene of miR-369-3p. The results also demonstrated that
EGFEM1P supported TCF4 expression via the regulation of miR-369-3p
expression levels. TCF4 is a crucial member of the TCF-4/lymphoid
enhancer factor gene family (34)
and acts as an oncogene in several types of cancer. For example,
TCF4 inhibition represses cell proliferation and invasion but
promotes cell cycle arrest and cell apoptosis in glioma (35). Moreover, in thyroid cancer, TCF4
binds to the promoter of lncRNA HLA complex P5 (HCP5) and activates
HCP5 expression to facilitate cancer cell proliferation (36). Although TCF4 overexpression has
previously been reported in thyroid cancer (34), the underlying mechanism that
regulates its expression is unknown. In the present study it was
identified that this regulation may be potentially associated with
the EGFEM1P/miR-369-3p/TCF4 axis in thyroid cancer.
Apoptosis is involved in the physiological
maintenance of normal cell homeostasis, and pathological lesions
which accompany numerous diseases (37). The caspase activation is a major
leading cause to apoptosis (38),
including effector (caspase-3) and initiator (caspase-8) (39,40).
poly ADP-ribose polymerase (PARP) is a DNA-binding enzyme involved
in apoptosis (41,42). Bax is a direct target in
p53-mediated apoptosis (43).
Furthermore, the present study demonstrated that EGFEM1P served a
critical role in promoting rapid cell proliferation, and inhibiting
apoptosis. In addition, detection of apoptosis-related proteins,
BAX and cleaved caspase-3/8/PARP was consistent with the apoptotic
rate.
In conclusion, the results of the present study
indicated that EGFEM1P may be important in promoting thyroid cancer
progression via acting as a miR-369-3p sponge.
However, there are several limitations to the
present study: i) Immunohistochemistry for TCF4 in thyroid cancer
patient samples and normal tissues was not conducted, which is
performed in the related ongoing study; ii) The animal study was
not conducted, which is performed in the related ongoing study; and
iii) The expression profiles of the remaining miRNAs (eg
miR-30a-3p) which were predicted to be targeted by EGFEM1P were not
detected, which will be performed in the future study.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SY and LL conducted the experiments and analyzed the
data. ZC conceived the study, analyzed the data and drafted the
manuscript. All authors read and approved the final manuscript. SY
and ZC confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
The study was approved (approval no.
CHESTJMAP-2017-08) by the Ethics Committee of The Central Hospital
of Enshi Tujia and Miao Autonomous Prefecture (Enshi, China).
Signed written informed consent was obtained from all patients.
Patient consent for publication
Signed written informed consent was obtained from
all patients.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Choi D, Ramu S, Park E, Jung E, Yang S,
Jung W, Choi I, Lee S, Kim KE, Seong YJ, et al: Aberrant activation
of Notch signaling inhibits PROX1 activity to enhance the malignant
behavior of thyroid cancer cells. Cancer Res. 76:582–593. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lim H, Devesa SS, Sosa JA, Check D and
Kitahara CM: Trends in thyroid cancer incidence and mortality in
the united states, 1974–2013. JAMA. 317:1338–1348. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chmielik E, Rusinek D, Oczko-Wojciechowska
M, Jarzab M, Krajewska J, Czarniecka A and Jarzab B: Heterogeneity
of thyroid cancer. Pathobiology. 85:117–129. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang TS and Sosa JA: Thyroid surgery for
differentiated thyroid cancer-recent advances and future
directions. Nat Rev Endocrinol. 14:670–683. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schlumberger M and Leboulleux S: Current
practice in patients with differentiated thyroid cancer. Nat Rev
Endocrinol. 17:176–188. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kwak D, Ha J, Won Y, Kwon Y and Park S:
Effects of thyroid-stimulating hormone suppression after
thyroidectomy for thyroid cancer on bone mineral density in
postmenopausal women: A systematic review and meta-analysis. BMJ
Open. 11:e0430072021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan J, Song Y, Pan W, Li Y, Xu Y, Xie M,
Shen Y, Zhang N, Liu J, Hua H, et al: LncRNA SLC26A4-AS1 suppresses
the MRN complex-mediated DNA repair signaling and thyroid cancer
metastasis by destabilizing DDX5. Oncogene. 39:6664–6676. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Landgraf P, Rusu M, Sheridan R, Sewer A,
Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M,
et al: A mammalian microRNA expression atlas based on small RNA
library sequencing. Cell. 129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Syeda ZA, Langden SSS, Munkhzul C, Lee M
and Song SJ: Regulatory mechanism of microRNA expression in cancer.
Int J Mol Sci. 21:17232020. View Article : Google Scholar
|
|
10
|
Pan D, Lin P, Wen D, Wei Y, Mo Q, Liang L,
Chen G, He Y, Chen J and Yang H: Identification of down-regulated
microRNAs in thyroid cancer and their potential functions. Am J
Transl Res. 10:2264–2276. 2018.PubMed/NCBI
|
|
11
|
Goodall GJ and Wickramasinghe VO: RNA in
cancer. Nat Rev Cancer. 21:22–36. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang J, Su Z, Lu S, Fu W, Liu Z, Jiang X
and Tai S: LncRNA HOXA-AS2 and its molecular mechanisms in human
cancer. Clin Chim Acta. 485:229–233. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Han X, Zhang J, Liu Y, Fan X, Ai S, Luo Y,
Li X, Jin H, Luo S, Zheng H, et al: The lncRNA Hand2os1/Uph locus
orchestrates heart development through regulation of precise
expression of Hand2. Development. 146:dev1761982019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo H, Xu C, Le W, Ge B and Wang T: lncRNA
CASC11 promotes cancer cell proliferation in bladder cancer through
miRNA-150. J Cell Biochem. 120:13487–13493. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shui X, Chen S, Lin J, Kong J, Zhou C and
Wu J: Knockdown of lncRNA NEAT1 inhibits Th17/CD4+ T cell
differentiation through reducing the STAT3 protein level. J Cell
Physiol. 234:22477–22484. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yu T, Yu J, Lu L, Zhang Y, Zhou Y, Zhou Y,
Huang F, Sun L, Guo Z, Hou G, et al: MT1JP-mediated
miR-24-3p/BCL2L2 axis promotes Lenvatinib resistance in
hepatocellular carcinoma cells by inhibiting apoptosis. Cell Oncol
(Dordr). 44:821–834. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liang M, Jia J, Chen L, Wei B, Guan Q,
Ding Z, Yu J, Pang R and He G: LncRNA MCM3AP-AS1 promotes
proliferation and invasion through regulating miR-211-5p/SPARC axis
in papillary thyroid cancer. Endocrine. 65:318–326. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Luo Y, Hao T, Zhang J, Zhang M, Sun P and
Wu L: MicroRNA-592 suppresses the malignant phenotypes of thyroid
cancer by regulating lncRNA NEAT1 and downregulating NOVA1. Int J
Mol Med. 44:1172–1182. 2019.PubMed/NCBI
|
|
19
|
Hua Q, Mi B, Xu F, Wen J, Zhao L, Liu J
and Huang G: Hypoxia-induced lncRNA-AC020978 promotes proliferation
and glycolytic metabolism of non-small cell lung cancer by
regulating PKM2/HIF-1α axis. Theranostics. 10:4762–4778. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
You X, Zhao Y, Sui J, Shi X, Sun Y, Xu J,
Liang G, Xu Q and Yao Y: Integrated analysis of long noncoding RNA
interactions reveals the potential role in progression of human
papillary thyroid cancer. Cancer Med. 7:5394–5410. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lan X, Zhang H, Wang Z, Dong W, Sun W,
Shao L, Zhang T and Zhang D: Genome-wide analysis of long noncoding
RNA expression profile in papillary thyroid carcinoma. Gene.
569:109–117. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Goldman MJ, Craft B, Hastie M, Repečka K,
McDade F, Kamath A, Banerjee A, Luo Y, Rogers D, Brooks AN, et al:
Visualizing and interpreting cancer genomics data via the Xena
platform. Nat Biotechnol. 38:675–678. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y and Wang X: MiRDB: An online
database for prediction of functional microRNA targets. Nucleic
Acids Res. 48:D127–D131. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:e050052015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dweep H and Gretz N: MiRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ye M, Dong S, Hou H, Zhang T and Shen M:
Oncogenic role of long noncoding RNAMALAT1 in thyroid cancer
progression through regulation of the miR-204/IGF2BP2/m6A-MYC
signaling. Mol Ther Nucleic Acids. 23:1–12. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang Z, Li H, Wang Z, Yang Y, Niu J, Liu
Y, Sun Z and Yin C: Microarray expression profile of long
non-coding RNAs in human lung adenocarcinoma. Thorac Cancer.
9:1312–1322. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu Q and Liu K: MiR-369-3p inhibits
tumorigenesis of hepatocellular carcinoma by binding to PAX6. J
Biol Regul Homeost Agents. 34:917–926. 2020.PubMed/NCBI
|
|
30
|
Liu P, Ma C, Wu Q, Zhang W, Wang C, Yuan L
and Xi X: MiR-369-3p participates in endometrioid adenocarcinoma
via the regulation of autophagy. Cancer Cell Int. 19:1782019.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Scalavino V, Liso M, Cavalcanti E, Gigante
I, Lippolis A, Mastronardi M, Chieppa M and Serino G: miR-369-3p
modulates inducible nitric oxide synthase and is involved in
regulation of chronic inflammatory response. Sci Rep. 10:159422020.
PubMed/NCBI
|
|
32
|
Dong L, Zhang Z, Xu J, Wang F, Ma Y, Li F,
Shen C, Liu Z, Zhang J, Liu C, et al: Consistency analysis of
microRNA-arm expression reveals microRNA-369-5p/3p as tumor
suppressors in gastric cancer. Mol Oncol. 13:1605–1620. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li P, Dong M and Wang Z: Downregulation of
TSPAN13 by miR-369-3p inhibits cell proliferation in papillary
thyroid cancer (PTC). Bosn J Basic Med Sci. 19:146–154.
2019.PubMed/NCBI
|
|
34
|
Gilbert-Sirieix M, Makoukji J, Kimura S,
Talbot M, Caillou B, Massaad C and Massaad-Massade L:
Wnt/beta-catenin signaling pathway is a direct enhancer of thyroid
transcription factor-1 in human papillary thyroid carcinoma cells.
PLoS One. 6:e222802011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sun J, Li B, Jia Z, Zhang A, Wang G, Chen
Z, Shang Z, Zhang C, Cui J and Yang W: RUNX3 inhibits glioma
survival and invasion via suppression of the beta-catenin/TCF-4
signaling pathway. J Neurooncol. 140:15–26. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang R, Cai J, Xie S, Zhao C, Wang Y, Cao
D and Li G: T cell factor 4 is involved in papillary thyroid
carcinoma via regulating long non-coding RNA HCP5. Technol Cancer
Res Treat. 19:15330338209832902020. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Adams JM: Ways of dying: Multiple pathways
to apoptosis. Genes Dev. 17:2481–2495. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gukovskaya AS and Pandol SJ: Cell death
pathways in pancreatitis and pancreatic cancer. Pancreatology.
4:567–586. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schultz DR and Harrington WJ Jr:
Apoptosis: Programmed cell death at a molecular level. Semin
Arthritis Rheum. 32:345–369. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cohausz O and Althaus FR: Role of PARP-1
and PARP-2 in the expression of apoptosis-regulating genes in HeLa
cells. Cell Biol Toxicol. 25:379–391. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Yu-Wei D, Li ZS, Xiong SM, Huang G, Luo
YF, Huo TY, Zhou MH and Zheng YW: Paclitaxel induces apoptosis
through the TAK1-JNK activation pathway. FEBS Open Biol.
10:1655–1667. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu X and Deng Y: Bax and BH3-domain-only
proteins in p53-mediated apoptosis. Front Biosci. 7:d151–d156.
2002. View Article : Google Scholar : PubMed/NCBI
|