Introduction
According to statistics regarding the global burden
of cancer released by the International Agency for Research on
Cancer, there were ~1.8 million cases of mortality resulting from
lung cancer worldwide in 2020, far surpassing those from other
cancer types and ranking first in terms of the cancer mortality
rate (1). As previously reported,
metabolic reprogramming is an important characteristic of cancer,
with both the Warburg effect and essential amino acid (such as
proline) metabolism alteration having been previously implicated in
this reprogramming (2). Proline has
been documented to exert protective effects against stress-induced
cell death and apoptosis in mammalian cells in culture to meet
their rapid proliferative needs (3,4). The
housekeeping enzyme that catalyzes the last step of proline
biosynthesis, pyrroline-5-carboxylate reductase 1 (PYCR1), utilizes
NAD(P)H as a cofactor to catalyze the transformation of
5-pyrrolinecarboxylic acid to proline (5). In addition, PYCR1 was previously found
to be one of the most commonly upregulated metabolic enzymes in
malignant tumors such as in lung, liver and prostate cancer, where
was also shown to be closely associated with the occurrence and
development of disease (6).
The PYCR1 gene is located on chromosome
17q2.3 and encodes a 33.4-kDa enzyme that consists of 319 amino
acids (7). Phang et al
(8) previously found that the PYCR1
protein is mainly localized to the mitochondria and forms a
decameric structure of two main structural domains, namely the N-
and C-terminal domains. The N-terminal domain folds to bind
NAD(P)H, whilst the C-terminal domain contains an α-helix to bind
other substrates, such as Pyrroline 5-carboxylate (P5C) or
L-Thiazolidine-4-carboxylate (T4C,thiaproline) (9,10).
When first discovered, PYCR1 gene mutations were found to be
closely associated with the loss of skin elasticity, premature
aging and antioxidant stress activation and mitochondrial
regulation (11–13). Bogner et al (6) previously analyzed the RNA-sequencing
data of 9,736 tumor and 8,587 normal tissues from 28 different
cancer types from the gene expression profiling database, GEPIA2.
Expression of the PYCR1 gene was found to be increased in
79% (22/28) of the cancer types tested, with a >4X increase
observed in 16 types of cancers compared with that in the
corresponding normal tissues. It has also previously been reported
that high expression levels of the PYCR1 gene in most solid
tumors may serve an oncogenic role by promoting tumor cell
proliferation whilst inhibiting apoptosis (6). By contrast, downregulation of the
PYCR1 gene with small interfering RNA (siRNA) has been
documented to inhibit cell proliferation and invasion whilst
promoting apoptosis in breast cancer (14).
Recombinant human (rh) TNF-related apoptosis
inducing ligand protein (TRAIL) has been reported to exert specific
antitumor properties in human melanoma WM793 and lung cancer cells
H460 whilst remaining non-toxic to normal cells (15,16).
However, a number of tumor cell types (such as colorectal cancer
cells and non-small cell lung cancer cells) can evade
TRAIL-mediated killing either directly (in a process known as
primary resistance) or by developing acquired resistance to TRAIL
(in a process known as secondary resistance) following treatment
(16,17). The long-term clinical efficacy of
TRAIL is therefore restricted by such drug resistance.
PYCR1 has also been implicated in drug resistance.
Meng et al (9) previously
found, via structural and biochemical analyses, that the catalytic
sites of human PYCR1 are located in a circular groove structured by
the N-terminal domain, which contains the conserved
dinucleotide-binding Rossmann motif (sequence, GXGXXA/G; part of
the Rossmann fold), and dimerization of the C-terminal domain. The
fine changes of PYCR1 result in an opening and closing of the
binding groove to facilitate the cofactor to enter and leave the
active site. Glu221 as a conformational switch for cofactor
selectivity for thiaproline dehydrogenase activity. Furthermore,
the thermal stability and fine changes of the PYCR1 enzyme was
associated with the oxidation of the antitumor drug, thiaproline.
In another study, She et al (18) found that silencing expression of the
PYCR1 gene significantly increased the sensitivity of lung
adenocarcinoma to cisplatin. Yan et al (19) also previously revealed that the
sensitivity of HCT116 and DLD1 colon cancer cells to 5-fluorouracil
was significantly increased following knockdown of PYCR1
expression. Ding et al (14)
found that PYCR1 is highly expressed in doxorubicin-resistant
breast cancer cells, while downregulation of PYCR1 expression using
siRNA transfection significantly enhanced the cytotoxicity of
doxorubicin. However, the relevance and molecular mechanism
underlying the function of the PYCR1 gene, in addition to
its potential role in TRAIL resistance, remains unclear.
In the present study, the association between the
PYCR1 gene and TRAIL resistance was explored.
Materials and methods
Reagents and plasmids
rhTRAIL was purchased from Sino Biological, Inc.
(cat. no. 10409-HNAE). A negative control siRNA with no homology to
other genes (‘NC’ hereafter) was used in RNA interference (Suzhou
GenePharma Co., Ltd.). siRNA1 or siRNA2 against PYCR1 gene
(‘siRNA1’ or ‘siRNA2’ hereafter) were used to knock down PYCR1
expression in H1299 cells(Suzhou GenePharma Co., Ltd.). The
pGCMV/MCS/Neo vector (‘pEX-3’ hereafter) was used as the control
vector (Suzhou GenePharma Co., Ltd.). The full-length PYCR1
gene was inserted between the SalI/BamHI cloning
sites of pEX-3 to construct the PYCR1-overexpression vector
(‘pEX-3-PYCR1’ hereafter) (Suzhou GenePharma Co., Ltd.).
Cells
The human bronchial epithelial cell line, HBE, and
the human non-small cell lung cancer (NSCLC) cell lines, H460
(TRAIL-sensitive) and H1299 (TRAIL-resistant cell line) were
purchased from the Type Culture Collection of the Chinese Academy
of Sciences (Shanghai, China), and preserved by Hubei Key
Laboratory of Tumor Microenvironment and Immunotherapy at China
Three Gorges University (Yichang, China). The cells were cultured
under 5% CO2 at 37°C in a humidified incubator in
RPMI-1640 medium (cat. no. PM150110; Procell Life Science &
Technology Co., Ltd.) containing 10% (v/v) fetal bovine serum (cat.
no. 164210; Procell Life Science & Technology Co., Ltd.) and
Penicillin-Streptomycin Solution (cat. no. PB180120; Procell Life
Science & Technology Co., Ltd.).
RNA interference, plasmids
transfection and generation of stable cell lines
First, PYCR1 expression was knocked down by
siRNA transfection. siRNAs were used to knock down PYCR1
expression in H1299 cells. The sequences of the negative control
(NC) and siRNA against PYCR1 (siRNA1 and siRNA2) are listed
in Table SI. Cells were first
seeded into 6-well plates and transfected once the density reached
60–70%. Following the manufacturer's protocol, TurboFect
transfection reagent (cat. no. R0531; Thermo Fisher Scientific,
Inc.) was used for transient transfection. A total of
1×105 adherent cells were cultured with 2 ml RPMI-1640
medium containing 10% (v/v) fetal bovine serum for 16 h at 37°C
before transfection. The transfection mixture was prepared
immediately before transfection. Specifically, 2 µg DNA was diluted
in 200 µl serum-free RPMI-1640 medium. For this, the transfection
reagent was gently mixed before being immediately added (4 µl) to
the diluted DNA mixture by pipetting. This mixture was then
incubated at room temperature for 15–20 min. Subsequently, 200 µl
of the transfection reagent + DNA mixture was added to each well.
The growth medium was not removed from the cells prior to adding
the transfection reagent + DNA mixture. The plate was then gently
shaken immediately after the addition of the transfection mixture
to evenly distribute the complexes. The cells were incubated under
5% CO2 at 37°C for 48 h, before transfection efficacy
was assessed by RT-qPCR or western blotting. Then cells were
divided into four groups and named NC, siRNA1, NC+TRAIL and
siRNA1+TRAIL. In siRNA1+TRAIL group, H1299 cells were transfected
by siRNA1 and then treated with 50 ng/ml TRAIL at 37°C for 48 h.
The expression of PYCR1, DR4 and DR5 gene were detected by RT-qPCR
or western blotting.
To determine explain the association between
PYCR1 and TRAIL resistance, transgenic cell line with stable
overexpression of PYCR1 was constructed. Following
transfection as aforementioned, the cells were transfected with
pEX-3 or pEX-3-PYCR1 vector and then selectively screened using
G418 sulfate solution (cat. no. G4024; Wuhan Servicebio Technology
Co., Ltd.). A G418 screening gradient (100–1,000 µg/ml) was added
to untransfected H1299 cells at 37°C, with medium + G418 changes
every 3–5 days. Subsequently, 800 µg/ml was found to be the optimal
G418 screening concentration since this concentration killed all
cells within 14 days. Following transfection of H1299 cells with
the overexpression plasmids, the cells were incubated with 800
µg/ml G418 for 14 days. Screening was then terminated for clone
amplification, whereby a cell suspension of each clone was prepared
by diluting the cells to 1 cell/10 µl with RPMI-1640 medium to a
96-well plate. Cells were divided into four groups: pEX-3,
pEX-3-PYCR1, pEX-3+TRAIL and pEX-3-PYCR1+TRAIL. In
pEX-3-PYCR1+TRAIL group, the PYCR1 overexpression stable transgenic
cell line was treated with 50 ng/ml TRAIL at 37°C for 48 h. Then
the expression of PYCR1, DR4 and DR5 gene were detected by RT-qPCR
or western blotting.
Detection of PYCR1,DR4 and DR5 gene
expression by reverse transcription-quantitative PCR (RT-qPCR)
At first, the difference expression of PYCR1 gene in
HBE, H1299 and H460 cells were detected by RT-qPCR. Total RNA of
those cells were extracted using TRIzol (cat. no. 15596-026;
Invitrogen; Thermo Fisher Scientific, Inc.) respectively, before
the RNA concentration and integrity of each sample was determined
using a Nanodrop spectrophotometer (Thermo Fisher Scientific,
Inc.). The first-strand synthesis of cDNA was conducted using the
Revert Aid First Strand cDNA Synthesis kit (cat. no. K1622; Thermo
Fisher Scientific, Inc. and reacted in gene amplification PCR
apparatus (Biometra Tone 96, Jena, Germany) at 42°C for 60 min and
then at 70°C for 10 min. qPCR was then performed using the cDNA as
a template and a HiScript II One Step qRT-PCR SYBR Green Kit (cat.
no. Q221-01; Vazyme Biotech Co., Ltd.) on a CFX96 qRT-PCR detection
system (Applied Biosystems; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol (pre denaturation for 5
min at 94°C; denaturation for 30 sec at 94°C, annealing for 30 sec
at 55°C, extension for 45 sec at 72°C, 35 cycles; final extension
for 10 min). The sequences of the primers used are listed in
Table SII. A total of three
replicate wells were set up for each sample. The 2−ΔΔCq
method was used to analyze the fold-change relative to that of
β-actin (20). Following the
same experimental procedure as aforementioned, the expression of
PYCR1, DR4 and DR5 gene after knockdown and overexpression of PYCR1
were detested by RT-qPCR.
Western blotting analysis
Expression of PYCR1 in HBE, H1299 and H460
cells were detected by Western blotting analysis. The whole cell
lysates of those three cells were extracted using RIPA (cat. no.
G2002; Wuhan Servicebio Technology Co., Ltd.) supplemented with
Protease Inhibitor Cocktail (cat. no. B14001; Selleck Chemicals).
The concentration of proteins was determined using the BCA Protein
Quantification Kit (cat. no. E112; Vazyme Biotech Co., Ltd.). In
total, 20–30 µg protein per lane was separated by SDS-PAGE (10%
gel), before transfer onto PVDF membranes. After blocking the
membranes for 2 h with TBST (TBS+0.1% Tween) containing 5% skimmed
milk at room temperature, the membranes were incubated with primary
antibodies (Table SIII) diluted
with the same solution at 4°C overnight. After washing with TBST
three times to remove unbound primary antibody, IgG (H + L)-HRP
secondary antibody was incubated with the membrane at 37°C for 1 h
(Table SIV). Enhanced
chemiluminescence reagent (cat. no. MA0186; Dalian Meilun Biology
Technology Co., Ltd.) was then used to visualize proteins, which
was performed on a ChemiScope 6100 gel imaging system (Shanghai
Clinx Scientific Instrument Co., LTD) and analyzed by
ChemiScope11.14.0 software (Shanghai Clinx Scientific Instrument
Co., LTD) and Image J 1.48 software(National Institutes of Health).
β-actin was used as the internal reference protein.
TRAIL can specifically induce apoptosis in tumor
cells, whereas the overexpression of anti-apoptotic proteins or
downregulation of pro-apoptotic proteins will lead to TRAIL
resistance (21). Therefore, the
expression of apoptosis-related proteins were measured by western
blotting analysis after PYCR1 knockdown or overexpression
and following TRAIL treatment. Finally, western blotting analysis
were conducted to test the expression DR4 and DR5.
The detailed experimental western blotting procedure is similar to
aforementioned.
TRAIL sensitivity detection by Cell
Counting Kit-8 (CCK-8) assay
The viability of cells was monitored using a CCK-8
kit (cat. no. CK04; Dojindo Laboratories, Inc.) following the
manufacturer's protocols. After PYCR1 knockdown or
overexpression, single-cell suspensions were inoculated in 96-well
overnight after cell counting, ensuring that each well contained
200 µl medium and 1×104 cells. Different concentrations
of TRAIL solution (0, 200, 400, 600, 800 and 1,000 ng/ml) were
prepared and added to the cells in each well. The 96-well plates
were then further incubated at 37°C for 48 h. A microplate reader
(Bio-Rad Laboratories, Inc.) was then used to measure the optical
density at 450 nm of each well following a 1 h incubation with 10%
(v/v) CCK-8 reagent at 37°C. The relative cell viability was
calculated using the following formula: Cell viability (%)=[optical
density (OD)treated/ODuntreated) ×100%. Each
experiment was conducted three times with five replicates.
Flow cytometry to detect apoptosis and
the expression of DR4/5 proteins on the cell membrane surface
Following transfection with NC or siRNA1 for 48 h,
H1299 cells were treated with 50 ng/ml TRAIL for 48 h at 37°C. The
Annexin-V-FITC/PI staining method was used to detect the effect of
PYCR1 knockdown on apoptosis. The cells were harvested using
trypsin (EDTA-free) and washed twice with PBS. Annexin-V-FITC/PI
kit (cat. no. BB4102; BestBio) was used to measure the extent of
apoptosis according to the manufacturer's protocols. A single cell
suspension was created by adding 400 µl binding buffer to
1×106 cells/ml, then 5 µl Annexin-V-FITC fluorescent
probe was incubated with the cell suspension for 20 min at 4°C
whilst being protected from light. Subsequently, 3 µl 20 mg/ml PI
was added, followed by 5 min incubation at room temperature
protected from light. A blank control is unstained cell with no
Annexin V-FITC and no PI and used to regulate voltage. And control
groups containing only Annexin-V-FITC or PI was set up to adjust
the compensation. The prepared samples were then assayed separately
on BD FACSVerse flow cytometer (BD Biosciences), analysed by BD
FACSuite™ 1.0.3 Software(BD Biosciences)and FlowJo 10 software
(FlowJo LLC).
The binding of TRAIL to DR4 and DR5 on the cell
membrane is a key step in the initiation of programmed cell death.
Therefore, the degree of DR membrane expression can be directly
measured as an indicator of sensitivity to TRAIL (22). Following transfection with siRNA,
each sample was collected and treated with TRAIL, before the
expression of DRs on the cell membranes was detected by flow
cytometry. A total of 1×106 cells were resuspended in 50
µl PBS containing 1% goat serum (cat. no. G1208-5ML; Wuhan
Servicebio Technology Co., Ltd.) and incubated for 30 min at room
temperature. After three washes with PBS, the cells were incubated
with primary antibodies (Table
SIII) at 4°C overnight. The cells were then washed three times
with PBS and incubated with the anti-Rabbit-IgG(H+L)-Cy3 secondary
antibody (Table SIV) at room
temperature for 30 min. An additional negative control was
established to set up the instrument until spontaneous fluorescence
was detected in the range of the negative gate. For this additional
negative control, cells were collected following transfection with
NC siRNA and treated as aforementioned but without the use of
primary antibody. Fluorescence intensity was then detected using
flow cytometry (BD FACSVerse, BD Biosciences). Each set of
experiments was repeated three times. The remaining samples were
then tested sequentially under the same instrument voltage and
gating strategy. The fluorescent intensity of samples was
considered to be directly proportional to the density of DR. All
histograms of the treatment samples were overlayed using the FlowJo
10 software (FlowJo LLC) to compare the expression of DRs on the
membrane surface. The proportion of positive cells in each treated
sample was analyzed statistically and visualized by column
diagrams.
To investigate the effect of PYCR1 overexpression on
cell apoptosis and the cytomembrane expression of DR4/5, the same
experiment was performed according to the aforementioned method
following the establishment of an PYCR1 overexpression stable
transgenic cell line through transfecting cells with pEX-3 control
or pEX-3-PYCR1 vectors and screening with G418. These cells were
then treated with TRAIL and the results analyzed by flow cytometry.
For the additional negative control to set up the instrument in
cytomembrane expression of DR4/5, cells collected following
transfection with the pEX-3 control vector were treated as
aforementioned but without the use of primary antibody.
Statistical analysis
Each experiment was repeated three times under the
same conditions. The results are presented as the mean ± standard
deviation. SPSS (version 24; IBM SPSS Advanced Statistics 24
Software; IBM Corp.) and GraphPad Prism (version 7.01; GraphPad
Software; Dotmatics) were used for data analyses. One-way ANOVA
followed by LSD post-hoc comparison was used to compare the
difference among three groups. Two-way ANOVA followed by Bonferroni
post-hoc test was used for cell viability analysis, to reflect the
influence of PYCR1 gene overexpression or knockdown on TRAIL
sensitivity. Differences between two groups were analyzed with
unpaired Student's t-tests. P<0.05 was considered to indicate a
statistically significant difference.
Results
PYCR1 may regulate TRAIL
sensitivity
To evaluate whether PYCR1 can regulate TRAIL
resistance in NSCLC cells, the PYCR1 protein expression levels in
HBE, H460 and H1299 cells were measured using western blotting. In
previous studies, H460 was reported to be a TRAIL sensitive cell
line, whilst H1299 was reported to be a TRAIL resistant cell line
(17,22,23).
PYCR1 expression was found to be increased in the two NSCLC cell
lines tested when compared with the HBE human bronchial epithelial
cell line (Fig. 1A and B). In
addition, PYCR1 expression was found to be increased in the
TRAIL-resistant cell line, H1299, when compared with the
TRAIL-sensitive cell line, H460. These observations suggested that
PYCR1 may be associated with NSCLC tumorigenesis and TRAIL
resistance. Therefore, PYCR1 expression was subsequently
overexpressed or knocked down to determine the effects on TRAIL
sensitivity in NSCLC cells.
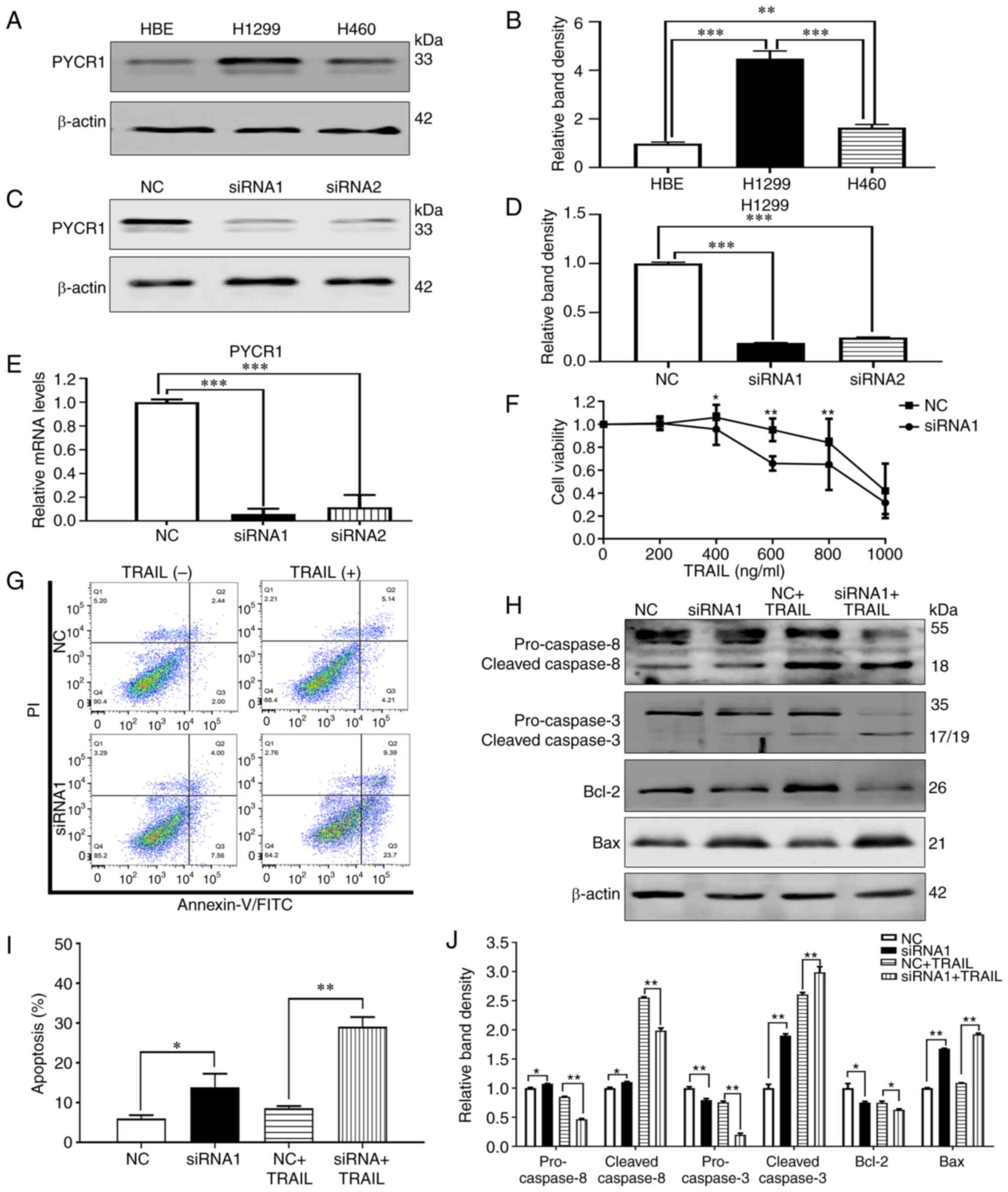 | Figure 1.Knockdown of PYCR1 enhances TRAIL
sensitivity in TRAIL-resistant H1299 cells. (A) PYCR1 protein
expression levels were detected by western blotting in HBE, H1299
and H460 cells, which were the (B) quantified and normalized to
β-actin. (C) Western blotting was used to detect the PYCR1 protein
expression levels in H1299 cells transfected with siRNA1/2 and NC,
which were then (D) quantified and normalized to β-actin. (E)
Relative PYCR1 mRNA expression levels in H1299 cells transfected
with siRNA1/2 or NC were detected by reverse
transcription-quantitative PCR. (F) The viability of H1299 cells
transfected with siRNA1 or NC for 48 h was measured using a Cell
Counting Kit-8 assay. (G) Representative Annexin V-FITC/PI flow
cytometry plots of H1299 cells transfected with siRNA1 or NC and
treated with 50 ng/ml TRAIL, which were then (I) quantified. (H)
Apoptosis marker protein expression levels in H1299 cells
transfected with siRNA1 or NC and treated with 50 ng/ml TRAIL
treatment were detected using western blotting, which were (J)
quantified and normalized to β-actin. Data are presented as the
mean ± SD from three different experiments. *P<0.05,
**P<0.01, ***P<0.001. PYCR1, pyrroline-5-carboxylate
reductase 1; TRAIL, TNF-related apoptosis-inducing ligand; NC,
negative control; siRNA, small interfering RNA. |
Knocking down PYCR1 by siRNA enhances
TRAIL sensitivity in H1299 cells
To clarify the effects of siRNA transfection on
PYCR1 mRNA and protein expression in H1299 cells, the cells were
first transfected with siRNA1 and siRNA2 before the expression
levels of PYCR1 mRNA and protein were detected by RT-qPCR and
western blotting. The results showed that siRNA1 transfection
reduced ~95% PYCR1 mRNA expression and ~81% PYCR1 protein
level in H1299 cells after 48 h, with its inhibitory effects more
pronounced compared with those mediated by siRNA2 (Fig. 1C-E). Therefore, siRNA1 was chosen
for subsequent experiments. Following transfection, the cells were
treated with various doses of TRAIL for 48 h. Knocking down PYCR1
expression resulted in an increased sensitivity of H1299 to TRAIL
after 48 h (Fig. 1F). In our
previous study (23), it was found
that 50 ng/ml TRAIL can induce apoptosis in H460 (TRAIL-sensitive
cell), while the effect was less obvious on H1299 (TRAIL-resistant
cell line) at the same concentration. Therefore, if H1299 cells
respond to 50 ng/ml TRAIL after knockdown of PYCR1 gene, it
indicates that the PYCR1 gene may be involved in TRAIL resistance.
So the degree of apoptosis induced by TRAIL was also observed to be
significantly increased after PYCR1 knockdown followed by 50 ng/ml
TRAIL treatment for 48 h (Fig. 1G and
I) compared with NC cells. These findings suggested that PYCR1
may mediate TRAIL resistance in a PYCR1 knockdown-reversible
manner. Changes in the expression levels of proteins associated
with the DR pathway (exogenous apoptotic pathway) and the
mitochondrial pathway (endogenous apoptotic pathway) were then
measured by western blotting as markers of sensitivity to TRAIL.
After PYCR1 knockdown by siRNA1 and treatment with 50 ng/ml TRAIL
for 48 h, the expression levels of apoptosis markers, Caspase 3,
Caspase 8, Bax and Bcl-2, were detected. The results showed that
knocking down PYCR1 expression resulted in the significant
activation of the executioner apoptotic protein, Caspase 3, with
the simultaneous activation of the exogenous apoptotic marker,
Caspase 8 (Fig. 1H and J). In
addition, expression of Bax, which belongs to the endogenous
apoptotic pathway, was increased whereas Bcl-2 was downregulated
following PYCR1 knockdown. The changes of apoptotic rate and
protein in siRNA1+TRAIL group were significant compared with
NC+TRAIL group. These results suggested that knocking down PYCR1
expression enhances TRAIL sensitivity by activating both the
exogenous and endogenous apoptotic pathways.
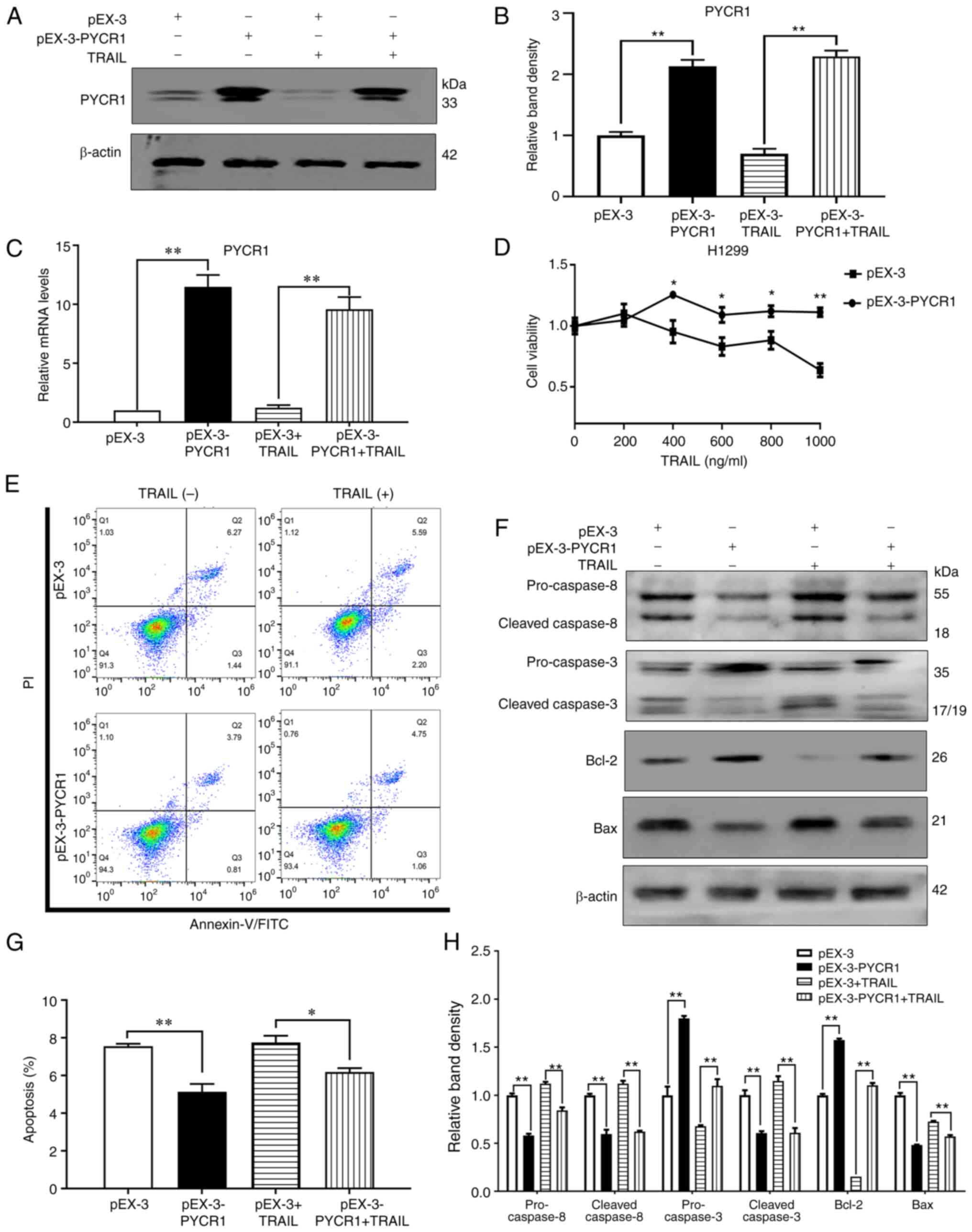
Overexpression of PYCR1 promotes TRAIL
resistance in H1299 cells
To analyze the relationship between PYCR1 and TRAIL
sensitivity, TRAIL-resistant H1299 cells were transfected with
pEX-3-PYCR1. After transfection and G418 screening, the PYCR1 mRNA
and protein expression levels were found to be significantly
upregulated in H1299 cells (Fig.
2A-C). According to the results of the CCK-8 assay,
overexpression of PYCR1 promoted TRAIL resistance in H1299 cells
(Fig. 2D). The apoptosis rate of
H1299 cells was also found to be reduced after PYCR1 overexpression
(Fig. 2E and G). In addition, the
protein expression levels of cleaved-caspase 8 and cleaved-caspase
3 were significantly decreased following PYCR1 overexpression
(Fig. 2F and H). Expression of the
pro-apoptotic protein, Bax, was found to be downregulated, whereas
the anti-apoptotic protein, Bcl-2, was upregulated. The changes of
apoptotic rate and protein in pEX-3-PYCR1+TRAIL group were
significant compared with pEX-3-PYCR1 group. These findings
suggested that PYCR1 expression is negatively associated with TRAIL
sensitivity, such that downregulation of PYCR1 expression using
siRNA transfection increased TRAIL sensitivity by activating the
apoptotic pathway. Conversely, NSCLC cells may ‘escape’ from
TRAIL-induced apoptosis by PYCR1 gene upregulation, resulting in
TRAIL resistance.
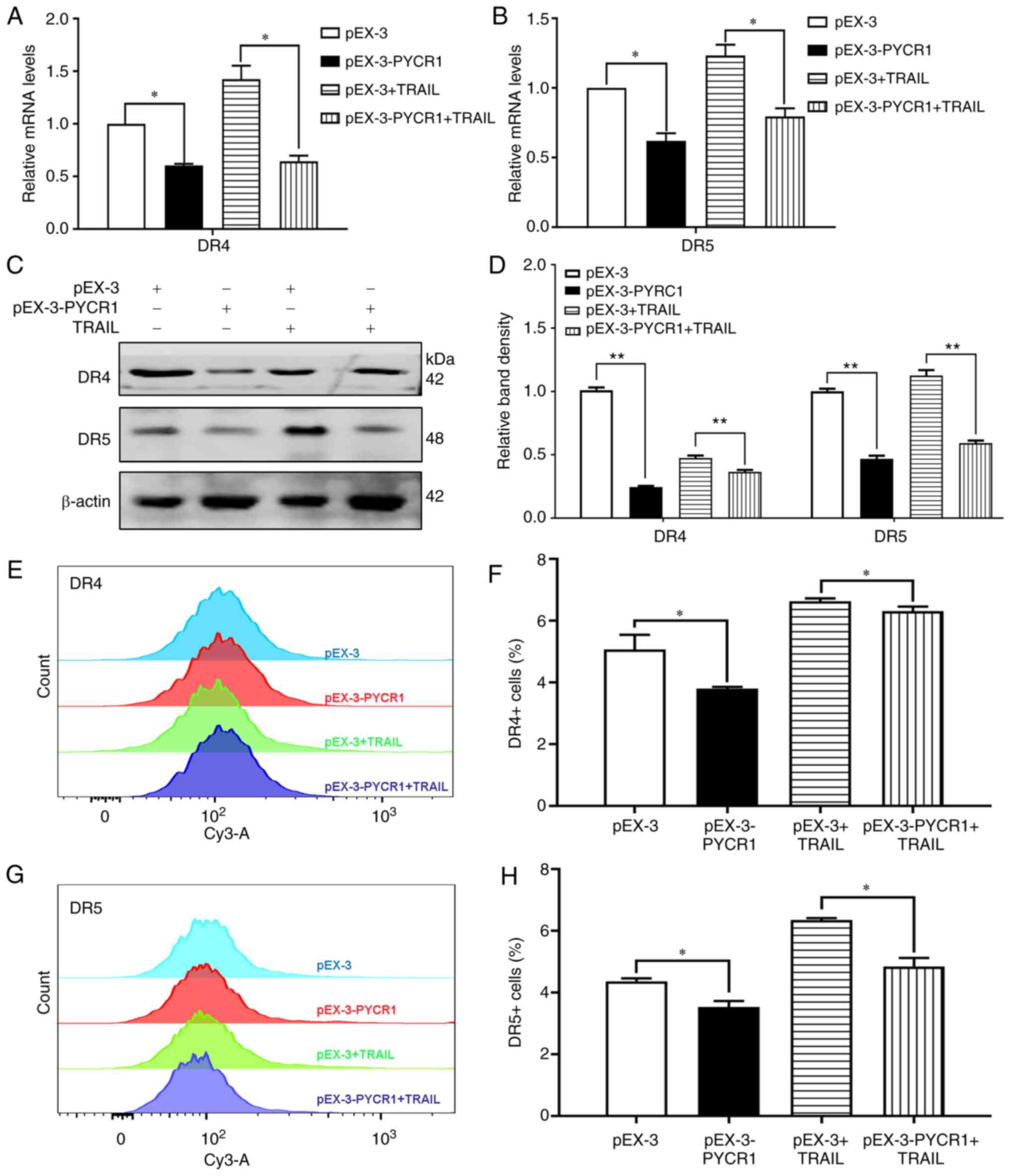
PYCR1 reduces DR4/5 expression in
NSCLC cells
A number of studies have previously demonstrated
that TRAIL triggers apoptosis after binding to DR4/5, which are
located on the plasma membrane (14,15).
Therefore, to assess the effects of PYCR1 on DR4/5 in the context
of TRAIL sensitivity, the membrane expression of these DRs was
measured following PYCR1 knockdown or overexpression. According to
the RT-qPCR and western blotting results, the DR4/5 mRNA and
protein levels were increased following the knockdown of PYCR1
(Fig. 3A-D). The changes of DR4/5
protein in siRNA1+TRAIL group were significant compared with
NC+TRAIL group. Subsequent flow cytometry assays revealed that
knocking down PYCR1 expression increased the cell membrane levels
of DR4/5 (Fig. 3E-H). In addition,
knocking down PYCR1 promoted the cell membrane expression of DR4/5,
where they can bind to TRAIL more efficiently to induce apoptosis
by activating pro-apoptotic caspase proteins, thereby reversing
TRAIL resistance.
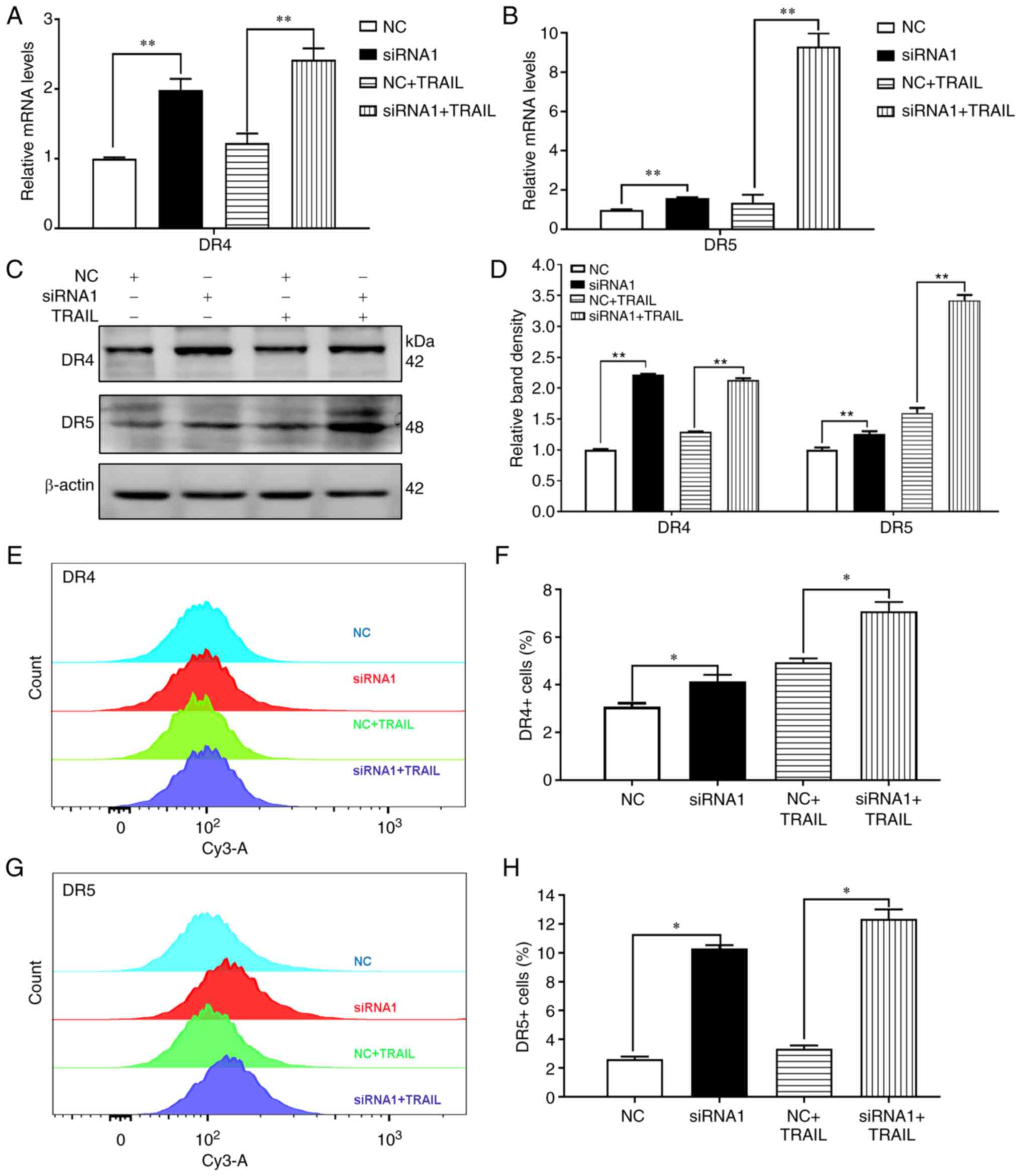
By contrast, compared with pEX-3-PYCR1 group, the
DR4/5 mRNA and protein expression levels were decreased in
pEX-3-PYCR1+TRAIL group (Fig.
4A-D). Decreased DR4/5 expression in the cell membrane was also
observed by flow cytometry following overexpression of
PYCR1(Fig. 4E-H). These
observations suggested that increased expression of PYCR1 not only
decreased the expression of DR4/5, but also inhibited translocation
of DR4/5 to the cytomembrane, thereby promoting resistance to
TRAIL.
Discussion
The lung cancer incidence and mortality rates have
been increasing rapidly, with >85% of lung cancer cases being
diagnosed as NSCLC and ~75% of cases being diagnosed at the
advanced stages on first presentation (24). At present, chemotherapy and targeted
therapy are the main treatment strategies for advanced lung cancer.
However, the majority of the currently available chemotherapeutic
reagents function by inducing tumor cell death through blocking DNA
replication and cell division, which causes DNA damage and cellular
stress. Therefore, these agents can also kill normal cells and
cause adverse effects. Although drugs targeting EGFR and anaplastic
lymphoma kinase (ALK) mutations are available, the number of
patients who can benefit from such treatments is limited. Ceritinib
targets patients with lung cancer harboring ALK mutations, which
only represent 3–5% of all patients with lung cancer (25). Therefore, a demand remains to
identify novel therapeutic strategies to selectively kill tumor
cells in patients with lung cancer.
TRAIL has the ability to induce cancer cell
apoptosis whilst selectively preserving normal cells (14,15).
Mechanistically, TRAIL binding to DR4/5 induces the aggregation of
Fas-associated death domain and Caspase-8, which induces apoptosis
in the cell of interest through the DR pathway (exogenous apoptotic
pathway) and the mitochondrial pathway (endogenous apoptotic
pathway) (26). In addition, TRAIL
also binds to decoy receptors, DcR1/2, but DcR1/2 is only expressed
in normal cells and since its intracellular segment lacks an intact
structural death domain, apoptotic signaling will not be activated
following binding (27,28). Therefore, TRAIL specifically induces
apoptosis in tumor cells but the DcR1/2 expressed on the surfaces
of normal cells will protect normal cells from TRAIL-mediated
killing (29). However, the
long-term clinical efficacy of TRAIL is restricted by drug
resistance, the mechanism of which has remained elusive. TRAIL
resistance has been documented to be caused by various factors,
such as protein synthesis disorders (30), decreased DR expression and increased
anti-apoptotic protein expression (21). The binding of TRAIL to its receptor
is the first step in TRAIL-induced apoptotic signaling. The cell
membrane localization of DR4/5, but not the absolute expression of
these DRs, is the main determinant of TRAIL sensitivity (16). Previous studies have shown that
TRAIL can induce the translocation of DRs to the lipid rafts of
sensitive cells, which does not occur in drug-resistant cells
(16,21). The present study reported that PYCR1
expression is negatively associated with TRAIL sensitivity by
reducing the plasma membrane expression of DR4/5 in NSCLC
cells.
PYCR1 may be an important molecule in the regulation
of TRAIL resistance in H1299 cells. PYCR1, a key enzyme in proline
biosynthesis (5), has been reported
to be highly expressed in lung cancer cells and tissues compared
with normal cells and adjacent tissues (18,31,32).
High levels of PYCR1 expression were found to be associated with a
poorer prognosis (31).
Furthermore, PYCR1 overexpression was previously found to promote
tumor cell resistance to cisplatin (18) and 5-fluorouracil (19). Knocking down PYCR1 expression was
also reported to reverse fluorouracil resistance by downregulating
the expression of the anti-apoptotic protein, Bcl-2 (19,20),
multidrug resistance-associated protein and p-glycoprotein
(18). The present study also
observed increased PYCR1 expression in TRAIL-resistant H1299 cells.
Knocking down PYCR1 expression using siRNA reversed this TRAIL
resistance suggesting that increased PYCR1 expression may promote
TRAIL resistance. This effect was likely achieved by regulating the
expression of anti-apoptotic proteins and/or DRs. Oudaert et
al (33) previously found that
multiple myeloma cells from patients with relapsed/refractory
cancer had significantly upregulated PYCR1 expression, whereas
knockdown of PYCR1 expression reduced cell viability and inhibited
cell proliferation by decreasing AKT, p42/44 MAPK, c-MYC and mTOR
activation. In addition, Cai et al (31) found that PYCR1 was highly expressed
in patients with NSCLC, which was in turn associated with a poorer
overall survival rate and higher TNM stages. Furthermore,
overexpression of PYCR1 promoted cell proliferation and inhibited
apoptosis by increasing Cyclin D1 and Bcl-xl expression. PYCR1
knockdown also resulted in cell cycle arrest in the G1 phase, which
inhibited cell proliferation and induced apoptosis. In another
study, Nilsson et al (34)
hypothesized that cancer cells may require high levels of PYCR1
expression to sustain growth and provide oxidative stress capacity.
As the housekeeping enzyme that catalyzes the last step in proline
biosynthesis, PYCR1 is mainly localized in the mitochondria and
utilizes NAD(P)H as a cofactor to catalyze the conversion of
5-pyrrolinecarboxylic acid to proline (5). Schwörer et al (35) also previously found that TGF-β
treatment elevated the expression of PYCR1 in the mitochondrial
proline biosynthetic pathway in mouse NIH-3T3 fibroblasts. Liu
et al (36) demonstrated
that PYCR1 is a target of the oncogene, c-MYC, which promotes the
proliferation of P493 human Burkitt lymphoma cells and PC3 human
prostate cancer cells through the regulation of proline metabolism
proliferation. Knocking down c-MYC expression resulted in a
decrease in PYCR1 expression and an increase in other proline
metabolizing enzymes.
Previous reports concluded that PYCR1 promotes the
development of lung cancer and that high PYCR1 expression was
associated with poor prognosis (31,32).
In the present study, an association between PYCR1 expression and
TRAIL sensitivity in NSCLC was found via PYCR1 gene overexpression
or knockdown experiments. The results of the present study verified
that PYCR1 mediates TRAIL resistance. PYCR1 may inhibit the
membrane expression of DRs for more efficient binding to TRAIL and
activate the downstream cysteine protease family of pro-apoptotic
molecules to induce apoptosis and reverse TRAIL resistance. In
conclusion, downregulation of PYCR1 may therefore be an effective
therapeutic strategy for promoting TRAIL sensitivity in NSCLC.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Ms. Jianlin Yang and
Ms. Yaling Fu (Hubei Key Laboratory of Tumor Microenvironment and
Immunotherapy, China Three Gorges University, Yichang, China) for
their technical assistance.
Funding
The present study was funded by Chinese National Natural Science
Foundation (grant no. 81374024), Scientific Research Program of
Hubei Provincial Department of Education (grant no. B2022033), The
2023-2024 Annual Scientific Research Project of Traditional Chinese
Medicine from Hubei Provincial Administration of Traditional
Chinese Medicine (grant no. ZY2023F145), Three Gorges University
Open Fund of Research Center for Basic and Clinical Pathology
(grant no. RHKFBL2022-01), Three Gorges University High-level
Talent Research Start-up Foundation (grant no. 8220309) and the
Open Fund of Hubei Key Laboratory of Tumor Microenvironment and
Immunotherapy (Three Gorges University; grant no. 2023KZL015).
Availability of data and materials
The data generated in the present study may be
requested from the corresponding author.
Authors' contributions
CYo, JH, YS and YH conceived and designed the
research. CYo, JH, LW, ZH, XZ and CYi performed the experiments.
CYo, CC, DS and CYi interpreted the results. CYo, JH, DS ZH and XZ
analyzed the data. CYo, DS, LW, ZH, and XZ prepared the figures.
CYo, JH, YS and YH searched the literature and drafted the
manuscript. YS and YH edited and revised the manuscript. All
authors have read and approved the final manuscript. CYo, JH and
CYi confirm the authenticity of all the raw data.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cao W, Chen HD, Yu YW, Li N and Chen WQ:
Changing profiles of cancer burden worldwide and in China: A
secondary analysis of the global cancer statistics 2020. Chin Med J
(Engl). 134:783–791. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Z, Wu X, Chen HN and Wang K: Amino
acid metabolic reprogramming in tumor metastatic colonization.
Front Oncol. 13:11231922023. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Krishnan N, Dickman MB and Becker DF:
Proline modulates the intracellular redox environment and protects
mammalian cells against oxidative stress. Free Radic Biol Med.
44:671–681. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pandhare J, Donald SP, Cooper SK and Phang
JM: Regulation and function of proline oxidase under nutrient
stress. J Cell Biochem. 107:759–768. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bogner AN, Stiers KM and Tanner JJ:
Structure, biochemistry, and gene expression patterns of the
proline biosynthetic enzyme pyrroline-5-carboxylate reductase
(PYCR), an emerging cancer therapy target. Amino Acids.
53:1817–1834. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu CA, Williams DB, Zhaorigetu S, Khalil
S, Wan G and Valle D: Functional genomics and SNP analysis of human
genes encoding proline metabolic enzymes. Amino Acids. 35:655–664.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Phang JM, Liu W and Zabirnyk O: Proline
metabolism and microenvironmental stress. Annu Rev Nutr.
30:441–463. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Meng Z, Lou Z, Liu Z, Li M, Zhao X,
Bartlam M and Rao Z: Crystal structure of human
pyrroline-5-carboxylate reductase. J Mol Biol. 359:1364–1377. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Christensen EM, Patel SM, Korasick DA,
Campbell AC, Krause KL, Becker DF and Tanner JJ: Resolving the
cofactor-binding site in the proline biosynthetic enzyme human
pyrroline-5-carboxylate reductase 1. J Biol Chem. 292:7233–7243.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dimopoulou A, Fischer B, Gardeitchik T,
Schroter P, Kayserili H, Schlack C, Li Y, Brum JM, Barisic I,
Castori M, et al: Genotype-phenotype spectrum of PYCR1-related
autosomal recessive cutis laxa. Mol Genet Metab. 110:352–361. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Scherrer DZ, Baptista MB, Matos AH,
Maurer-Morelli CV and Steiner CE: Mutations in PYCR1 gene in three
families with autosomal recessive cutis laxa, type 2. Eur J Med
Genet. 56:336–339. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yasuda T, Kaji Y, Agatsuma T, Niki T,
Arisawa M, Shuto S, Ariga H and Iguchi-Ariga SM: DJ-1 cooperates
with PYCR1 in cell protection against oxidative stress. Biochem
Biophys Res Commun. 436:289–294. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding J, Kuo ML, Su L, Xue L, Luh F, Zhang
H, Wang J, Lin TG, Zhang K, Chu P, et al: Human mitochondrial
pyrroline-5-carboxylate reductase 1 promotes invasiveness and
impacts survival in breast cancers. Carcinogenesis. 38:519–531.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wiley SR, Schooley K, Smolak PJ, Din WS,
Huang CP, Nicholl JK, Sutherland GR, Smith TD, Rauch C and Smith
CA: Identification and characterization of a new member of the TNF
family that induces apoptosis. Immunity. 3:673–682. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hao C, Song JH, Hsi B, Lewis J, Song DK,
Petruk KC, Tyrrell DL and Kneteman NM: TRAIL inhibits tumor growth
but is nontoxic to human hepatocytes in chimeric mice. Cancer Res.
64:8502–8506. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jong K, Mohamed E and Ibrahim ZA: Escaping
cell death via TRAIL decoy receptors: A systematic review of their
roles and expressions in colorectal cancer. Apoptosis. 27:787–799.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
She Y, Mao A, Li F and Wei X: P5CR1
protein expression and the effect of gene-silencing on lung
adenocarcinoma. Peer J. 7:e69342019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yan K, Xu X, Wu T, Li J, Cao G, Li Y and
Ji Z: Knockdown of PYCR1 inhibits proliferation, drug resistance
and EMT in colorectal cancer cells by regulating STAT3-Mediated p38
MAPK and NF-kappaB signalling pathway. Biochem Biophys Res Commun.
520:486–491. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mert U and Sanlioglu AD: Intracellular
localization of DR5 and related regulatory pathways as a mechanism
of resistance to TRAIL in cancer. Cell Mol Life Sci. 74:245–255.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ouyang W, Yang C, Liu Y, Xiong J, Zhang J,
Zhong Y, Zhang G, Zhou F, Zhou Y and Xie C: Redistribution of DR4
and DR5 in lipid rafts accounts for the sensitivity to TRAIL in
NSCLC cells. Int J Oncol. 39:1577–1586. 2011.PubMed/NCBI
|
|
23
|
You C, Sun Y, Zhang S, Tang G, Zhang N, Li
C, Tian X, Ma S, Luo Y, Sun W, et al: Trichosanthin enhances
sensitivity of non-small cell lung cancer (NSCLC) TRAIL-resistance
cells. Int J Biol Sci. 14:217–227. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao C, D'Amico T, Demmy T, Dunning J,
Gossot D, Hansen H, He J, Jheon S, Petersen RH, Sihoe A, et al:
Surgery versus SABR for resectable non-small-cell lung cancer.
Lancet Oncol. 16:e370–e371. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kischkel FC, Lawrence DA, Chuntharapai A,
Schow P, Kim KJ and Ashkenazi A: Apo2L/TRAIL-dependent recruitment
of endogenous FADD and caspase-8 to death receptors 4 and 5.
Immunity. 12:611–620. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ashkenazi A: Directing cancer cells to
self-destruct with pro-apoptotic receptor agonists. Nat Rev Drug
Discov. 7:1001–1012. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Johnstone RW, Frew AJ and Smyth MJ: The
TRAIL apoptotic pathway in cancer onset, progression and therapy.
Nat Rev Cancer. 8:782–798. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Maksimovic-Ivanic D, Stosic-Grujicic S,
Nicoletti F and Mijatovic S: Resistance to TRAIL and how to
surmount it. Immunol Res. 52:157–168. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fan S, Li Y, Yue P, Khuri FR and Sun SY:
The eIF4E/eIF4G interaction inhibitor 4EGI-1 augments
TRAIL-mediated apoptosis through c-FLIP down-regulation and DR5
induction independent of inhibition of cap-dependent protein
translation. Neoplasia. 12:346–356. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cai F, Miao Y, Liu C, Wu T, Shen S, Su X
and Shi Y: Pyrroline-5-carboxylate reductase 1 promotes
proliferation and inhibits apoptosis in non-small cell lung cancer.
Oncol Lett. 15:731–740. 2018.PubMed/NCBI
|
|
32
|
Xue W, Wu K, Guo X, Chen C, Huang T, Li L,
Liu B, Chang H and Zhao J: The pan-cancer landscape of glutamate
and glutamine metabolism: A comprehensive bioinformatic analysis
across 32 solid cancer types. Biochim Biophys Acta Mol Basis Dis.
1870:1669822024. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Oudaert I, Satilmis H, Vlummens P, De
Brouwer W, Maes A, Hose D, De Bruyne E, Ghesquiere B, Vanderkerken
K, De Veirman K and Menu E: Pyrroline-5-carboxylate reductase 1: A
novel target for sensitizing multiple myeloma cells to bortezomib
by inhibition of PRAS40-mediated protein synthesis. J Exp Clin
Cancer Res. 41:452022. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nilsson R, Jain M, Madhusudhan N, Sheppard
NG, Strittmatter L, Kampf C, Huang J, Asplund A and Mootha VK:
Metabolic enzyme expression highlights a key role for MTHFD2 and
the mitochondrial folate pathway in cancer. Nat Commun. 5:31282014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schwörer S, Berisa M, Violante S, Qin W,
Zhu J, Hendrickson RC, Cross JR and Thompson CB: Proline
biosynthesis is a vent for TGFβ-induced mitochondrial redox stress.
EMBO J. 39:e1033342020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu W, Le A, Hancock C, Lane AN, Dang CV,
Fan TW and Phang JM: Reprogramming of proline and glutamine
metabolism contributes to the proliferative and metabolic responses
regulated by oncogenic transcription factor c-MYC. Proc Natl Acad
Sci USA. 109:8983–8988. 2012. View Article : Google Scholar : PubMed/NCBI
|