Cancer, as a serious health problem, represents one
of the main causes of morbidity and mortality worldwide (1). Immunotherapy is a treatment method
that utilizes the body's own immune system to combat diseases and
has shown potential in cancer treatment. However, despite
significant success, immunotherapy still faces multiple challenges
(2). Genetic heterogeneity within
tumors may lead to inconsistent responses of tumor cells in
different regions to immunotherapy, while antigen loss makes it
difficult for the immune system to recognize and target all tumor
cells (3). Tumor cells may develop
resistance to immunotherapy through various mechanisms, such as
upregulation or downregulation of programmed death-ligand 1
(PD-L1), changes in the IFNγ signaling pathway and alterations in
metabolic pathways (4). For
instance, Choi et al (4)
found that the consumption of developmentally regulated GTP binding
protein 2 in melanoma cells not only increases the expression of
PD-L1 in tumor cells, but also increases the proportion of
IFN-expressing CD8 T cells in tumor-infiltrating immune cells.
Immunotherapy may cause immune-related adverse events (irAEs), such
as inflammatory reactions in the endocrine system, skin, digestive
tract and lungs (5). Therefore, it
is important to overcome these adverse factors by developing new
therapies (6).
In previous years, immunotherapy has been developed
to design effective treatment methods to enhance the specificity
and intensity of the immune system towards cancer (7). T cells are cellular effectors and
coordinators in cancer, which serve as adaptive immune cells
required for immune tolerance, host defense, immune memory and
homeostasis (8). A large amount of
data indicates that T cell activation, clone amplification and
effector differentiation are closely related to cell energy
metabolism (9). Based on studies of
immune checkpoint programmed cell death protein 1 (PD-1) and
cytotoxic T lymphocyte-associated protein 4 (CTLA-4), the
inhibition of immune checkpoints can more effectively activate T
cells and eliminate cancer cells (7) and the immune checkpoint inhibitors may
have important therapeutic value (6).
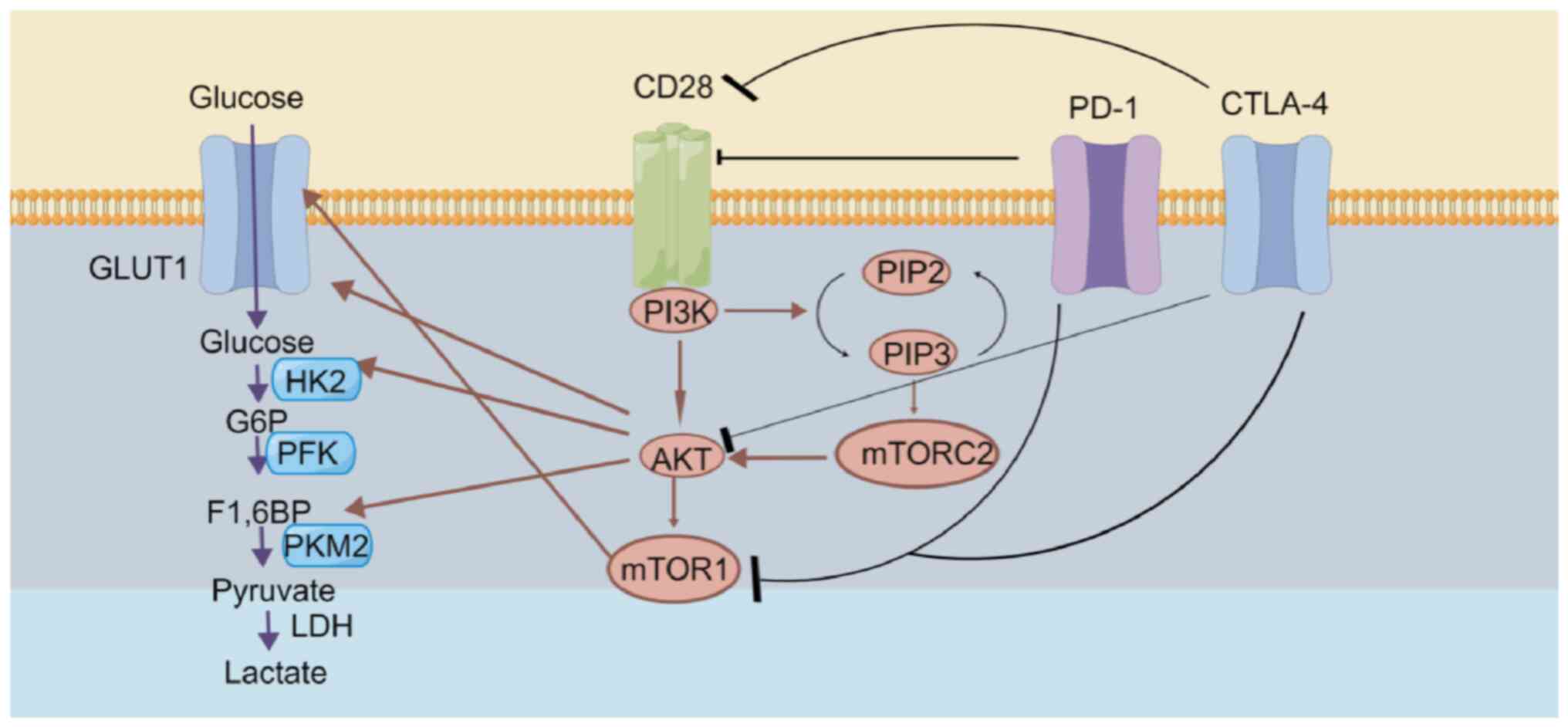
The PI3K/AKT/mTOR pathway represents a primary
signaling pathway that regulates processes, including cell
metabolism, apoptosis and proliferation (10,11).
The activation of PI3K/AKT can strengthen the nutrient intake and
energy production of CD8+T cells, whereas mTOR is
responsible for participating in regulating both innate and
adaptive immune systems and the biological effects of immune cell
stimulation (8,12). PI3K/AKT activation can promote PD-L1
expression and co-stimulate CD28, which can enhance T cell
activation and metabolism. Therefore, the present article mainly
explored the interaction and clinical application of the
PD-1/CTLA-4/CD28 and PI3K/AKT/mTOR pathways in T cells.
By recognizing and killing tumor cells, T
lymphocytes protect the body from cancer (13). According to the functions and
phenotypes, T lymphocytes can be mainly classified into immature T
cells (Tn) and memory T cells (Tm) (14). To be specific, Tn represents a
dormant mature T cell. Through the circulation between the blood
and secondary lymphoid organs, Tn exhibits immune surveillance
functions (15,16). In addition, Tm is included in
maintaining the rapid and long-term immune responses, also known as
the memory immune responses (17,18).
Situated between Tn and Tcm, Tscm is a significantly different
subset (19). Tn and T memory stem
cells (Tscm) can self-renew and differentiate into each subset of
memory and effector T cells, which are central memory T cells
(Tcm), Terminal effector T cells (Tte) and effector memory T cells
(Tem) (14). Usually, Tcm cells
exist in lymphoid organs and show no direct lytic function
(20). Tscm and Tcm go through
memory immune responses and quickly clone and proliferate to
generate Tem and Tte which particularly kill tumor cells (21).
The energy requirements for T cell proliferation and
differentiation are met through the prominent programming of
cellular metabolism, and the different phenotypes of T cells can
determine their different metabolic modes (22–24).
Initial T cells maintain the minimum ATP levels by uptaking basic
nutrients via oxidative phosphorylation (OXPHOS) while maintaining
basic metabolic needs by relying on fatty acid oxidation (FAO) and
glutamine metabolism (8). However,
under aerobic conditions, rapidly dividing T cells transition their
metabolism from oxidative phosphorylation to aerobic glycolysis and
glutamine breakdown (25). Despite
the existence of oxygen, glucose can still ferment into lactic
acid, which can enter the tricarboxylic acid cycle (the TCA cycle),
where the main carbon flux is converted from glucose to glutamine
(12,25). After encountering antigens, immature
T cells rapidly transform into effector T cells, which exhibit
increased nutrient absorption and glycolysis rates and a metabolic
activation state, mainly relying on OXPHOS and aerobic glycolysis
to maintain T cell adaptability and function (26,27).
Given the 10 enzymatic steps of glycolysis (which
converts glucose to pyruvate), some intermediates are generated for
the various biosynthetic pathways. That can involve de novo
fatty acid synthesis, the pentose phosphate pathway, hexosamine
biosynthesis and serine biosynthesis, where the pentose phosphate
pathway plays an important role in cell growth and offers primary
precursors for nucleotide synthesis (28,29).
Therefore, glycolysis is not only an energy production pathway in T
cells, but also the metabolic foundation for proliferating
synthetic organisms (30). Glucose
transporter 1 (GLUT1) is a key signaling molecule for T lymphocyte
activation and metabolism (31),
capturing glucose to convert it into lactic acid, which can be used
for oxidative phosphorylation even with sufficient oxygen (32). Glucose is first converted to
glucose-6-phosphate (G-6-P), then to fructose-6-phosphate (F-6-P)
and further to F-1,6-BP by the key regulatory factor PFK1 in
glycolysis (33). Next F-1,6-BP
enters the second part of glycolysis, which ultimately produces ATP
and pyruvate (34). Therefore,
glycolysis contributes much to T cell proliferation and
differentiation.
According to differences in structure and function,
PI3K is classified into categories I, II and III (35). To generate 3,4,5-triphosphate
phosphatidylinositol (PIP3), PI3K phosphorylates 4,5-diphosphate
phosphatidylinositol. PIP3 co-acts with target proteins [including
Akt and phosphoinositol dependent protein kinase (PDK1)] that
involve pleckstrin homologous domains on the inner lobe of the
plasma membrane (36). Also known
as protein kinase B (PKB), there are three subtypes in Akt, which
are Akt1/PKBα, Akt2/PKBβ and Akt3/PKBγ (37). To reach complete activation, Akt
should be respectively phosphorylated by PDK1 and mTOR complex 2
(mTORC2) (38). MTOR contributes
much as the key element of the two multi-subunit proteins that have
various functions. The complex is called mTORC1 and mTORC2
(39). In turn, MTORC1 activation
can control protein synthesis, metabolism and cell growth. MTOR is
fundamentally part of PI3K related kinases (PIKK), and most PIKK
members possess conserved domains (40).
The PI3K/Akt/mTOR signaling pathway exists in each
of the mammalian cells and exerts a significant impact on different
processes, including cell proliferation, metabolism,
differentiation and migration. In T cells, PI3Kδ and PI3Kγ subtypes
play an important role in development (41). In the process of thymogenesis,
absent or inactivated two isomers hinder the CD4 CD8 double
negative stage of T cell development. By contrast, PI3Kδ is a
subtype that can contribute significantly to PI3K signal
transduction in mature T cells (42). For CD8+T cells, IA type
PI3K can be mainly activated by tyrosine kinase-related receptors,
including T cell receptors (TCRs), cytokine receptors and
co-stimulatory receptors (43). The
PI3K/Akt signaling pathway in CD8+T cells is stimulated
by the signaling pathways triggered by cytokines IL-12, IL-2, IL-7,
IL-15 and IL-21 (44). IL-2 can
produce sustained high levels of PIP3, whereas IL-15 can relatively
stimulate PI3K weakly, which can cause lower levels of PIP3.
Conversely, chemokine receptors and other G protein coupled
receptors can activate IB-type PI3K. The mTOR pathway represents an
important regulatory factor for cell growth and proliferation and
is becoming an attractive target for cancer treatment (45). Apart from the cancer cells, mTOR
contributes much to mediating T cell activation and differentiation
(46).
T cells are activated via antigen recognition
through TCRs and co-stimulatory signals including CD28 (47). CD28 is considered a biosensor for T
cell metabolism (48). CD28
co-stimulation plays an important role in strengthening T cell
activation and metabolism and is antagonized via the inhibitory and
checkpoint immunotherapy receptors CTLA-4 and PD-1 (49).
In addition to revolutionizing cancer treatment,
cancer immunotherapy targeting the inhibitory co-receptors PD-1 and
CTLA-4 has also shown long-standing clinical benefits for different
tumor types (51,52). CTLA-4 inhibits T cell activation
while interacting with CD86 or CD80 through transduction of
inhibitory signals and/or the inhibition that involves
co-stimulatory protein CD28 (53,54).
CTLA-4 can also downregulate T cell glycolysis 1, and related
studies have shown that CTLA-4 blockade can affect the metabolic
adaptability of T cells in tumors related to tumor glycolysis
ability (48).
CD28 co-stimulation increases T cell synthesis and
metabolism, whereas CD28 family members PD-1 and CTLA-4 can inhibit
T cell metabolic reprogramming (55). PD-1 can inhibit glycolysis, promote
the FAO of endogenous lipids, and change nucleoside synthesis. In
addition, PD-1 negatively regulates T cells through changing
mitochondrial cristae formation, which hinders oxidative
phosphorylation (56). Besides,
CTLA-4 can inhibit Akt/mTORC1 on the CD28 signaling and
PI3K/Akt/mTORC1 signaling pathways, which can cause reduced
glycolysis and mitochondrial oxidative capacity (57,58).
AKT can regulate important enzymes in T cell metabolism, including
GLUT1, hexokinase 2 and pyruvate kinase isozymes M1/M2, and mTORC1
can promote the expression of GLUT1, which promotes the process of
T cell glycolysis (Fig. 1).
CTLA-4 is a key negative regulatory molecule
recruited to the cell membrane during T-cell activation and can
bind to helper molecules of the B7 family expressed by
antigen-presenting cells (59,60).
Since the binding of CTLA-4 effectively inhibits further activation
and proliferation of T cells, the progression of immune response
can be controlled and the occurrence of chronic autoimmune
inflammation can also be reduced (61). In cancer treatment, the blocking of
CTLA-4 can not only relieve the inhibition of anti-cancer T cells
but also trigger a new immune response. Related studies have shown
that ipilimumab is a therapeutic drug with primary clinical
significance, especially for patients in advanced stages (62,63).
Different from conventional treatment that directly kills cancer
cells and rapidly causes tumor volume reduction, it may take
several months for ipilimumab to stimulate T cell responses
(64). In addition, other
applications of ipizumab have also been pursued, especially in
combination with other immunotherapies, such as effective immune
checkpoint inhibitors, vaccine administration and small molecule
tyrosine kinase inhibitors (65).
The metabolism can affect the functions of the PD-1
pathway. In the cytoplasmic domain in PD-1, there are two tyrosine
motifs. Upon binding to ligands, the phosphorylation of tyrosine
residues of PD-1 occurs, which can cause the binding to The Src
homology 2 and other protein tyrosine phosphatases (PTP) (66). Then, PTP can phosphorylate kinases
and antagonize positive signals that occur via TCR and CD28, which
affects the downstream signaling pathways (67), such as Extracellular
signal-regulated kinase 1/2 (ERK) (68), PI3K-AKT (69), finally reducing T cell survival,
activation and proliferation and altering cytokine production and
metabolism.
Given the effectiveness of the PD-1 pathway blockade
monotherapy, improved response rates are associated with the
infiltration of CD8+T cells at the tumor margin and high
levels of PD-L1 expression (70).
However, PD-L1 expression is linked to treatment outcomes all the
time, as some PD-L1 tumors exhibit poor response to PD-1 pathway
blockade while others exhibit a good response. Therefore, multiple
biomarkers may play a more effective role in predicting responses
of anti-PD-1 monotherapy than PD-L1 expression levels alone
(71). In addition, in Renal Cell
Carcinoma, certain metabolic signals are linked to treatment
failure, which may be caused by the increased metabolic
adaptability of tumor cells. Conversely, the immunological markers
that involve BACH2 encode transcription factors regulating
differentiation and function of effector T cells and memory T
cells, as well as CCL3, which can encode chemokines in leukocyte
migration, and has a connection to successful RCC treatment of
PD-L1 (72,73). In the recent metastatic melanoma
data, there is a relationship between mesenchymal and ‘inhibitory
inflammatory’ transcriptional phenotypes and the efficacy of PD-1
inhibitors (74).
The mTOR pathway is a crucial regulatory factor for
innate (including the dendritic cells and macrophages) and adaptive
effector (including the T and B lymphocytes) immune cell
metabolism, proliferation and anti-inflammatory response (75). Since the mTOR pathway is usually
dysregulated in various solid tumors and hematological malignancy
types, mTOR inhibitors (mTORi) indicate the immunosuppressive
method that can prevent transplant rejection in transplant patients
(76) and the anti-tumor therapy,
which can be combined with immunotherapy and carefully adjusted for
the immunosuppressive dose (77).
Using mTORi and reducing the dosage of other immunosuppressive
drugs is related to improved overall survival in patients with
cancer. research indicates that T cell anergy can be maintained by
using mTORi and accompanying immune checkpoint inhibitor therapy.
Besides, mTORi stimulates the differentiation of immature T cells
into Tregs (78).
Hence, mTORi therapy decreases cancer progression in
various malignant tumors and exerts antitumor effects, which is
seen to be controversial (79–81).
Currently, only a few mTOR inhibitors are employed in clinical
practice. All approved mTOR inhibitors belong to the
first-generation mTOR inhibitors. Rapamycin is the first mTOR
inhibitor approved by the FDA and is currently suitable for
preventing organ rejection after organ transplantation. Also, it
can be used alone or in combination with calcineurin inhibitors or
corticosteroids (82,83) (Table
I).
Despite significant progress in the field of
immunotherapy for lung cancer, there are still a number of
challenges to be faced. The present article reviewed the components
of the PI3K/AKT/mTOR signaling pathway and the interactions between
the PD-1/CTLA-4/CD28 and the PI3K/AKT/mTOR pathways in T cells and
their impact on T cell metabolism and proliferation ability. CD28
can promote T cell glycolysis through activating the PI3K/Akt/mTOR
pathway, while CD28 is inhibited by PD-1 and CTLA4. Therefore, it
is necessary to further consider PI3K/AKT/mTOR pathway inhibitors
combined with PD-1/PD-L1 inhibition to regulate T cell metabolism
and proliferation, which can prevent and treat cancer.
Not applicable.
Funding: No funding was received.
Not applicable.
SCW designed the review, prepared the figure, wrote
the manuscript and made substantial contributions to conception and
design, acquisition of data, and analysis and interpretation of
data. CYL, CXY, YTJ and QC participated in the conception and
design of the study. DW, TG and GXH analysed and interpretated the
data. WTL, GZ, AQL, YX and YHL were involved in the conception and
design of the study and revised the manuscript. JCY revised the
manuscript. All authors read and approved the final manuscript.
Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global Cancer Statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shi H, Zhang W, Zhang L, Zheng Y and Dong
T: Comparison of different predictive biomarker testing assays for
PD-1/PD-L1 checkpoint inhibitors response: A systematic review and
network meta-analysis. Front Immunol. 14:12652022023. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Qiu J, Cheng Z, Jiang Z, Gan L, Zhang Z
and Xie Z: Immunomodulatory precision: A narrative review exploring
the critical role of immune checkpoint inhibitors in cancer
treatment. Int J Mol Sci. 25:54902024. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Choi SH, Mani M, Kim J, Cho WJ, Martin
TFJ, Kim JH, Chu HS, Jeong WJ, Won YW, Lee BJ, et al: DRG2 is
required for surface localization of PD-L1 and the efficacy of
anti-PD-1 therapy. Cell Death Discov. 10:2602024. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shen X, Yang J, Qian G, Sheng M, Wang Y,
Li G and Yan J: Treatment-related adverse events of immune
checkpoint inhibitors in clinical trials: A systematic review and
meta-analysis. Front Oncol. 14:13917242024. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Han Y, Liu D and Li L: PD-1/PD-L1 pathway:
Current researches in cancer. Am J Cancer Res. 10:727–742.
2020.PubMed/NCBI
|
|
7
|
Chen C, Wang Z and Qin Y: CRISPR/Cas9
system: Recent applications in immuno-oncology and cancer
immunotherapy. Exp Hematol Oncol. 12:952023. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Guo C, Chen S, Liu W, Ma Y, Li J, Fisher
PB, Fang X and Wang XY: Immunometabolism: A new target for
improving cancer immunotherapy. Adv Cancer Res. 143:195–253. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lim SA, Su W, Chapman NM and Chi H: Lipid
metabolism in T cell signaling and function. Nat Chem Biol.
18:470–481. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mafi S, Mansoori B, Taeb S, Sadeghi H,
Abbasi R, Cho WC and Rostamzadeh D: mTOR-Mediated regulation of
immune responses in cancer and tumor microenvironment. Front
Immunol. 12:7741032021. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He J, Zeng F, Jin XI, Liang L, Gao M, Li
W, Li G and Zhou Y: YWHAH activates the HMGA1/PI3K/AKT/mTOR
signaling pathway by positively regulating Fra-1 to affect the
proliferation of gastric cancer cells. Oncol Res. 31:615–630. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liang Y, Wang X, Wang H, Yang W, Yi P,
Soong L, Cong Y, Cai J, Fan X and Sun J: IL-33 activates mTORC1 and
modulates glycolytic metabolism in CD8+ T cells. Immunology.
165:61–73. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kruse B, Buzzai AC, Shridhar N, Braun AD,
Gellert S, Knauth K, Pozniak J, Peters J, Dittmann P, Mengoni M, et
al: CD4+ T cell-induced inflammatory cell death controls
immune-evasive tumours. Nature. 618:1033–1040. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Saxena A, Dagur PK and Biancotto A:
Multiparametric flow cytometry analysis of naïve, memory, and
effector T cells. Methods Mol Biol. 2032:129–140. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Notarbartolo S and Abrignani S: Human T
lymphocytes at tumor sites. Semin Immunopathol. 44:883–901. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Goronzy JJ and Weyand CM: Mechanisms
underlying T cell ageing. Nat Rev Immunol. 19:573–583. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zander R, Schauder D, Xin G, Nguyen C, Wu
X, Zajac A and Cui W: CD4+ T cell help is required for the
formation of a Cytolytic CD8+ T cell subset that protects against
chronic infection and cancer. Immunity. 51:1028–1042.e4. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Omilusik KD and Goldrath AW: Remembering
to remember: T cell memory maintenance and plasticity. Curr Opin
Immunol. 58:89–97. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fazeli P, Kalani M and Hosseini M: T
memory stem cell characteristics in autoimmune diseases and their
promising therapeutic values. Front Immunol. 14:12042312023.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Q, Sun Z and Chen L: Memory T cells:
Strategies for optimizing tumor immunotherapy. Protein Cell.
11:549–564. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang G, Liu A, Yang Y, Xia Y, Li W, Liu
Y, Zhang J, Cui Q, Wang D, Liu X, et al: Clinical predictive value
of naïve and memory T cells in advanced NSCLC. Front Immunol.
13:9963482022. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Reina-Campos M, Scharping NE and Goldrath
AW: CD8+ T cell metabolism in infection and cancer. Nat Rev
Immunol. 21:718–738. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wißfeld J, Werner A, Yan X, Ten Bosch N
and Cui G: Metabolic regulation of immune responses to cancer.
Cancer Biol Med. 19:1528–1542. 2022.PubMed/NCBI
|
|
24
|
Ma S, Ming Y, Wu J and Cui G: Cellular
metabolism regulates the differentiation and function of T-cell
subsets. Cell Mol Immunol. 21:419–435. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Araujo L, Khim P, Mkhikian H, Mortales CL
and Demetriou M: Glycolysis and glutaminolysis cooperatively
control T cell function by limiting metabolite supply to
N-glycosylation. Elife. 6:e213302017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kumar A and Chamoto K: Immune metabolism
in PD-1 blockade-based cancer immunotherapy. Int Immunol. 33:17–26.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li F, Liu H, Zhang D, Ma Y and Zhu B:
Metabolic plasticity and regulation of T cell exhaustion.
Immunology. 167:482–494. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Upadhyay S, Khan S and Hassan MI:
Exploring the diverse role of pyruvate kinase M2 in cancer:
Navigating beyond glycolysis and the Warburg effect. Biochim
Biophys Acta Rev Cancer. 1879:1890892024. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Barba I, Carrillo-Bosch L and Seoane J:
Targeting the Warburg effect in cancer: Where do we stand? Int J
Mol Sci. 25:31422024. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
von Meyenn L, Bertschi NL and Schlapbach
C: Targeting T cell metabolism in inflammatory skin disease. Front
Immunol. 10:22852019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang Z and Li X, Yang F, Chen C, Liu P,
Ren Y, Sun P, Wang Z, You Y, Zeng YX and Li X: DHHC9-mediated GLUT1
S-palmitoylation promotes glioblastoma glycolysis and
tumorigenesis. Nat Commun. 12:58722021. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gerriets VA, Kishton RJ, Nichols AG,
Macintyre AN, Inoue M, Ilkayeva O, Winter PS, Liu X, Priyadharshini
B, Slawinska ME, et al: Metabolic programming and PDHK1 control
CD4+ T cell subsets and inflammation. J Clin Invest. 125:194–207.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Adeva-Andany MM, Pérez-Felpete N,
Fernández-Fernández C, Donapetry-García C and Pazos-García C: Liver
glucose metabolism in humans. Biosci Rep. 36:e004162016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Icard P, Alifano M, Donnadieu E and Simula
L: Fructose-1,6-bisphosphate promotes PI3K and glycolysis in T
cells? Trends Endocrinol Metab. 32:540–543. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bilanges B, Posor Y and Vanhaesebroeck B:
PI3K isoforms in cell signalling and vesicle trafficking. Nat Rev
Mol Cell Biol. 20:515–534. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shen Q, Han Y, Wu K, He Y, Jiang X, Liu P,
Xia C, Xiong Q, Liu R, Chen Q, et al: MrgprF acts as a tumor
suppressor in cutaneous melanoma by restraining PI3K/Akt signaling.
Signal Transduct Target Ther. 7:1472022. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Akhtar N and Jabeen I: Pharmacoinformatic
approaches to design novel inhibitors of protein kinase B pathways
in cancer. Curr Cancer Drug Targets. 18:830–846. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun K, Luo J, Guo J, Yao X, Jing X and Guo
F: The PI3K/AKT/mTOR signaling pathway in osteoarthritis: A
narrative review. Osteoarthritis Cartilage. 28:400–409. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jhanwar-Uniyal M, Wainwright JV, Mohan AL,
Tobias ME, Murali R, Gandhi CD and Schmidt MH: Diverse signaling
mechanisms of mTOR complexes: mTORC1 and mTORC2 in forming a
formidable relationship. Adv Biol Regul. 72:51–62. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen Y and Zhou X: Research progress of
mTOR inhibitors. Eur J Med Chem. 208:1128202020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Dwyer CJ, Arhontoulis DC, Rangel Rivera
GO, Knochelmann HM, Smith AS, Wyatt MM, Rubinstein MP, Atkinson C,
Thaxton JE, Neskey DM and Paulos CM: Ex vivo blockade of PI3K gamma
or delta signaling enhances the antitumor potency of adoptively
transferred CD8+ T cells. Eur J Immunol. 50:1386–1399. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Preite S, Gomez-Rodriguez J, Cannons JL
and Schwartzberg PL: T and B-cell signaling in activated PI3K delta
syndrome: From immunodeficiency to autoimmunity. Immunol Rev.
291:154–173. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Chapman NM, Yoder AN, Barbón KM, Bilal MY,
Connolly SF and Houtman JC: Proline-rich tyrosine kinase 2 controls
PI3-kinase activation downstream of the T cell antigen receptor in
human T cells. J Leukoc Biol. 97:285–296. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Coppola C, Hopkins B, Huhn S, Du Z, Huang
Z and Kelly WJ: Investigation of the Impact from IL-2, IL-7, and
IL-15 on the growth and signaling of activated CD4+ T Cells. Int J
Mol Sci. 21:78142020. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mossmann D, Park S and Hall MN: mTOR
signalling and cellular metabolism are mutual determinants in
cancer. Nat Rev Cancer. 18:744–757. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Werlen G, Jain R and Jacinto E: MTOR
signaling and metabolism in Early T cell development. Genes
(Basel). 12:7282021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Wen L: The tumor immunotherapy effect of
dendritic cells carrying anti CD40 single chain antibodies and the
expression and role of CD40 on CD4+T cell surface D. Suzhou
University; 2012
|
|
48
|
Zappasodi R, Serganova I, Cohen IJ, Maeda
M, Shindo M, Senbabaoglu Y, Watson MJ, Leftin A, Maniyar R, Verma
S, et al: CTLA-4 blockade drives loss of Treg stability in
glycolysis-low tumours. Nature. 591:652–658. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Beckermann KE, Hongo R, Ye X, Young K,
Carbonell K, Healey DCC, Siska PJ, Barone S, Roe CE, Smith CC, et
al: CD28 costimulation drives tumor-infiltrating T cell glycolysis
to promote inflammation. JCI Insight. 5:e1387292020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sugiura D, Okazaki IM, Maeda TK, Maruhashi
T, Shimizu K, Arakaki R, Takemoto T, Ishimaru N and Okazaki T: PD-1
agonism by anti-CD80 inhibits T cell activation and alleviates
autoimmunity. Nat Immunol. 23:399–410. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Sun C, Mezzadra R and Schumacher TN:
Regulation and function of the PD-L1 checkpoint. Immunity.
48:434–452. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ribas A and Wolchok JD: Cancer
immunotherapy using checkpoint blockade. Science. 359:1350–1355.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Lin F, Chen Y, Huang B, Ruan S, Lin J,
Chen Z, Huang C and Zhao B: Application of immune checkpoint
inhibitors for resectable gastric/gastroesophageal cancer. Front
Pharmacol. 15:13915622024. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Oyewole-Said D, Konduri V, Vazquez-Perez
J, Weldon SA, Levitt JM and Decker WK: Beyond T-cells: Functional
characterization of CTLA-4 expression in immune and non-immune cell
types. Front Immunol. 11:6080242020. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xia S, Chen Q and Niu B: CD28: A new drug
target for immune disease. Curr Drug Targets. 21:589–598. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kumagai S, Koyama S, Itahashi K,
Tanegashima T, Lin YT, Togashi Y, Kamada T, Irie T, Okumura G, Kono
H, et al: Lactic acid promotes PD-1 expression in regulatory T
cells in highly glycolytic tumor microenvironments. Cancer Cell.
40:201–218.e9. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Palaskas NJ, Garcia JD, Shirazi R, Shin
DS, Puig-Saus C, Braas D, Ribas A and Graeber TG: Global alteration
of T-lymphocyte metabolism by PD-L1 checkpoint involves a block of
de novo nucleoside phosphate synthesis. Cell Discov. 5:622019.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Dong S, Guo X, Han F, He Z and Wang Y:
Emerging role of natural products in cancer immunotherapy. Acta
Pharm Sin B. 12:1163–1185. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Pulanco MC, Madsen AT, Tanwar A, Corrigan
DT and Zang X: Recent advancements in the B7/CD28 immune checkpoint
families: New biology and clinical therapeutic strategies. Cell Mol
Immunol. 20:694–713. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sharma P, Goswami S, Raychaudhuri D,
Siddiqui BA, Singh P, Nagarajan A, Liu J, Subudhi SK, Poon C, Gant
KL, et al: Immune checkpoint therapy-current perspectives and
future directions. Cell. 186:1652–1669. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Zhang J, Wei X, Zhang Q, Jiao X, Li K,
Geng M, Cao Y, Wang D, Cheng J and Yang J: Fish uses CTLA-4 immune
checkpoint to suppress mTORC1-Controlled T-cell glycolysis and
immunity. J Immunol. 212:1113–1128. 2024. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hellmann MD, Paz-Ares L, Bernabe Caro R,
Zurawski B, Kim SW, Carcereny Costa E, Park K, Alexandru A,
Lupinacci L, de la Mora Jimenez E, et al: Nivolumab plus Ipilimumab
in Advanced Non-Small-Cell Lung Cancer. N Engl J Med.
381:2020–2031. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
VanderWalde A, Bellasea SL, Kendra KL,
Khushalani NI, Campbell KM, Scumpia PO, Kuklinski LF, Collichio F,
Sosman JA, Ikeguchi A, et al: Ipilimumab with or without nivolumab
in PD-1 or PD-L1 blockade refractory metastatic melanoma: A
randomized phase 2 trial. Nat Med. 29:2278–2285. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Cascone T, Leung CH, Weissferdt A, Pataer
A, Carter BW, Godoy MCB, Feldman H, William WN Jr, Xi Y, Basu S, et
al: Neoadjuvant chemotherapy plus nivolumab with or without
ipilimumab in operable non-small cell lung cancer: The phase 2
platform NEOSTAR Trial. Nat Med. 29:593–604. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Grimm MO, Esteban E, Barthélémy P,
Schmidinger M, Busch J, Valderrama BP, Charnley N, Schmitz M,
Schumacher U, Leucht K, et al: Tailored immunotherapy approach with
nivolumab with or without nivolumab plus ipilimumab as
immunotherapeutic boost in patients with metastatic renal cell
carcinoma (TITAN-RCC): A multicentre, single-arm, phase 2 trial.
Lancet Oncol. 24:1252–1265. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Liu L, Cheng Y, Zhang Z, Li J, Geng Y, Li
Q, Luo D, Liang L, Liu W, Hu J and Ouyang W: Study on the
allosteric activation mechanism of SHP2 via elastic network models
and neural relational inference molecular dynamics simulation. Phys
Chem Chem Phys. 25:23588–23601. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Ai L, Xu A and Xu J: Roles of PD-1/PD-L1
pathway: Signaling, cancer, and beyond. Adv Exp Med Biol.
1248:33–59. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Guo YJ, Pan WW, Liu SB, Shen ZF, Xu Y and
Hu LL: ERK/MAPK signalling pathway and tumorigenesis. Exp Ther Med.
19:1997–2007. 2020.PubMed/NCBI
|
|
69
|
Quan Z, Yang Y, Zheng H, Zhan Y, Luo J,
Ning Y and Fan S: Clinical implications of the interaction between
PD-1/PD-L1 and PI3K/AKT/mTOR pathway in progression and treatment
of non-small cell lung cancer. J Cancer. 13:3434–3443. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Huang Q, Wu X, Wang Z, Chen X, Wang L, Lu
Y, Xiong D, Liu Q, Tian Y, Lin H, et al: The primordial
differentiation of tumor-specific memory CD8+ T cells as bona fide
responders to PD-1/PD-L1 blockade in draining lymph nodes. Cell.
185:4049–4066.e25. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Patel SP and Kurzrock R: PD-L1 Expression
as a predictive biomarker in cancer immunotherapy. Mol Cancer Ther.
14:847–856. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Karim S, Mirza Z, Chaudhary AG, Abuzenadah
AM, Gari M and Al-Qahtani MH: Assessment of radiation induced
therapeutic effect and cytotoxicity in cancer patients based on
transcriptomic profiling. Int J Mol Sci. 17:2502016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Ascierto ML, McMiller TL, Berger AE,
Danilova L, Anders RA, Netto GJ, Xu H, Pritchard TS, Fan J, Cheadle
C, et al: The intratumoral balance between metabolic and
immunologic gene expression is associated with Anti-PD-1 response
in patients with renal cell carcinoma. Cancer Immunol Res.
4:726–733. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Van Damme H, Dombrecht B, Kiss M, Roose H,
Allen E, Van Overmeire E, Kancheva D, Martens L, Murgaski A, Bardet
PMR, et al: Therapeutic depletion of CCR8+ tumor-infiltrating
regulatory T cells elicits antitumor immunity and synergizes with
anti-PD-1 therapy. J Immunother Cancer. 9:e0017492021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Zou Z, Tao T, Li H and Zhu X: mTOR
signaling pathway and mTOR inhibitors in cancer: Progress and
challenges. Cell Biosci. 10:312020. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lim LM, Kung LF, Kuo MC, Huang AM and Kuo
HT: Timing of mTORI usage and outcomes in kidney transplant
recipients. Int J Med Sci. 18:1179–1184. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Kumari A, Gesumaria L, Liu YJ, Hughitt VK,
Zhang X, Ceribelli M, Wilson KM, Klumpp-Thomas C, Chen L, McKnight
C, et al: mTOR inhibition overcomes RSK3-mediated resistance to BET
inhibitors in small cell lung cancer. JCI Insight. 8:e1566572023.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Daenthanasanmak A, Lin Y, Zhang M, Bryant
BR, Petrus MN, Bamford RN, Thomas CJ, Miljkovic MD, Conlon KC and
Waldmann TA: Enhanced efficacy of JAK1 inhibitor with mTORC1/C2
targeting in smoldering/chronic adult T cell leukemia. Transl
Oncol. 14:1009132021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bai S, Taylor SE, Jamalruddin MA,
McGonigal S, Grimley E, Yang D, Bernstein KA and Buckanovich RJ:
Targeting therapeutic resistance and multinucleate giant cells in
CCNE1-Amplified HR-Proficient ovarian cancer. Mol Cancer Ther.
21:1473–1484. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Sawaki M, Muramatsu Y, Togo K and Iwata H:
Real-world treatment patterns of subsequent therapy after
palbociclib in patients with advanced breast cancer in Japan
Breast. 70:1–7. 2023.PubMed/NCBI
|
|
81
|
Qiu HY, Wang PF and Zhang M: A patent
review of mTOR inhibitors for cancer therapy (2011–2020). Expert
Opin Ther Pat. 31:965–975. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Vidigal AC, de Lucena DD, Beyerstedt S and
Rangel ÉB: A comprehensive update of the metabolic and
toxicological considerations for immunosuppressive drugs used
during pancreas transplantation. Expert Opin Drug Metab Toxicol.
19:405–427. 2023. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Franzin R, Netti GS, Spadaccino F, Porta
C, Gesualdo L, Stallone G, Castellano G and Ranieri E: The use of
immune checkpoint inhibitors in oncology and the occurrence of AKI:
Where do we stand? Front Immunol. 11:5742712020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zhang Y and Zhang Z: The history and
advances in cancer immunotherapy: Understanding the characteristics
of tumor-infiltrating immune cells and their therapeutic
implications. Cell Mol Immunol. 17:807–821. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Catalano M, Iannone LF, Nesi G, Nobili S,
Mini E and Roviello G: Immunotherapy-related biomarkers:
Confirmations and uncertainties. Crit Rev Oncol Hematol.
192:1041352023. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Al-Hussainy A, Adams J, Simmons J and
Kennedy J: Immune checkpoint inhibitor associated diarrhoea. BMJ
Case Rep. 17:e2590572024. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Blum SM, Rouhani SJ and Sullivan RJ:
Effects of immune-related adverse events (irAEs) and their
treatment on antitumor immune responses. Immunol Rev. 318:167–178.
2023. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Ma S, Lei J and Lai X: Modeling tumour
heterogeneity of PD-L1 expression in tumour progression and
adaptive therapy. J Math Biol. 86:382023. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yi M, Zheng X, Niu M, Zhu S, Ge H and Wu
K: Combination strategies with PD-1/PD-L1 blockade: Current
advances and future directions. Mol Cancer. 21:282022. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Chen Z, Yue Z, Wang R, Yang K and Li S:
Nanomaterials: A powerful tool for tumor immunotherapy. Front
Immunol. 13:9794692022. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Li J, Zeng H, Li L, Yang Q, He L and Dong
M: Advanced generation therapeutics: Biomimetic nanodelivery system
for tumor immunotherapy. ACS Nano. 17:24593–24618. 2023. View Article : Google Scholar : PubMed/NCBI
|