C-C motif chemokine ligand 2 (CCL2), is a key member
of the CC subfamily of chemokines, which was purified and
identified in 1989 by Yoshimura et al (1). CCL2 exerts chemotactic effects on
monocytes, macrophages and T lymphocytes (2), and is secreted by activated cells
through autocrine or paracrine methods. Cytokines in the tumor
microenvironment (TME), including TNFα, lipopolysaccharide, IL-1,
IL-6 and transforming growth factor-β (TGF-β), stimulate tumor
cells to produce CCL2 (2). C-C
motif chemokine receptor 2 (CCR2), the main receptor of CCL2, is a
G protein-coupled receptor expressed in both immune and tumor cells
(2–4). It plays an essential role in different
aspects of tumor cell biology, including the regulation of
proliferation, angiogenesis, immune response and migration of cells
in inflammatory environments (5).
CCL2 exerts its biological effects mainly by combining with CCR2
(2,6).
Studies have found that numerous types of tumor
cells, including myeloma, breast cancer, prostate cancer and
melanoma cells, express CCR2 and secrete high levels of CCL2
(7–9). CCL2 thus promotes tumor cell
proliferation and survival through autocrine or paracrine pathways,
participating in the regulation of tumor immune tolerance, inducing
tumor angiogenesis, and promoting tumor invasion and metastasis
(10).
The ‘seed and soil’ hypothesis posited that, during
metastasis, tumor cells not only adapt to the recipient
microenvironment but also actively remodel it to facilitate
colonization (16,17). The chemokine-receptor crosstalk
represents a pivotal research frontier in tumor metastasis biology.
Its multifaceted roles include tumor cell proliferation,
chemoresistance, migratory/invasive capacity and organotropic
metastasis, alongside regulatory effects on angiogenesis and
lymphangiogenesis. Growth factors, circulatory hypoxia, antitumor
drugs and radiation therapy stimulate tumor cells to secrete more
CCL2, induce the establishment of an immunosuppressive TME through
the CCL2/CCR2 signaling axis, and promote tumor progression and
metastasis. de Visser reviewed the importance of the TME in every
stage of cancer progression, including tumor initiation,
progression, invasion, infiltration, metastasis, diffusion and
growth (18). TME represents a
highly complex and dynamic ecosystem, primarily composed of three
major cellular components: myeloid cells (including
tumor-associated macrophages [TAMs], myeloid-derived suppressor
cells [MDSCs], dendritic cells [DCs], and tumor-associated
neutrophils [TANs]), lymphoid cells (such as T cells, B cells,
natural killer [NK] cells, and innate lymphoid cells [ILCs]), and
stromal components (cancer-associated fibroblasts [CAFs] and
endothelial cells) (18). While
these host-derived cells were historically dismissed as passive
bystanders in tumorigenesis, emerging evidence highlights their key
roles in driving cancer pathogenesis (18). The cellular architecture and
functional phenotype of the TME exhibit pronounced heterogeneity
dictated by primary tumor location, cancer cell-intrinsic
properties, disease stage and patient-specific factors (18). Deciphering the crosstalk among tumor
cell-autonomous signals, microenvironmental cues and systemic
regulatory networks is key for the rationale-driven design of
next-generation cancer therapeutics. The present study summarized
the role of the CCL2/CCR2 signaling axis in tumor progression and
metastasis.
CCL2 orchestrates antitumor immune responses by
promoting immune cell recruitment and surveillance. In vitro
studies have shown that diverse tumor cell lines chemoattract
CD8+ and CD4+ T lymphocytes through CCL2
secretion (19,20). For example, in a previous study of
nude mice, ovarian cancer cells engineered to express CCL2 induced
robust monocyte infiltration at the injection site, resulting in
localized tumor growth inhibition (20) Clinically, plasma CCL2 levels in
patients with pancreatic cancer have been shown to be negatively
correlated with tumor proliferation markers (21). In addition, in colorectal cancer
preclinical models (22–24), the targeted modulation of
tumor-derived chemokines enhances immune cell infiltration and
suppresses tumor progression.
CCL2 serves a pivotal role in the metastatic
cascade, particularly during epithelial-mesenchymal transition
(EMT). Preclinical studies have shown EMT-induced cancer cells
upregulate CCL2 expression (22–25),
which in turn attracts neutrophils to the TME. While neutrophils
are traditionally associated with tumor-promoting inflammation,
evidence has highlighted their potential to exert cytotoxic effects
against cancer cells in specific contexts (26–28).
Tumor-associated neutrophils (TANs) have two phenotypes. ‘N2’ is
known as a pro-tumorigenic phenotype, as it exerts a
pro-tumorigenic effect at the primary site by secreting oncogenic
factors, promoting primary tumor growth and angiogenesis,
increasing extracellular matrix degradation and suppressing immune
responses (29–33). ‘N1’ is an antitumorigenic phenotype.
These neutrophil phenotype switches are contingent on TGF-β
signaling, as the neutralization of TGF-β has been shown to induce
a shift from ‘N2’ pro-tumorigenic to ‘N1’ antitumorigenic
neutrophil states (31).
Tumor-entrained neutrophils suppress lung metastatic seeding via
H2O2-dependent cytotoxic mechanisms, with
tumor-derived CCL2 serving as a key mediator of granulocyte
colony-stimulating factor-stimulated neutrophil recruitment for
optimal anti-metastatic function (28). A preclinical study of mouse breast
cancer models further demonstrated that neutrophils accumulate in
the lungs prior to the arrival of metastatic cells, establishing a
pre-metastatic surveillance niche (28).
TANs exhibit dual roles in cancer progression, with
N1-polarized TANs exerting antitumor effects and N2-polarized TANs
promoting tumorigenesis. In hepatocellular carcinoma (HCC),
cancer-associated fibroblasts (CAFs) secrete cardiotrophin-like
cytokine factor 1 (CLCF1), which drives tumor stemness via C-X-C
motif chemokine ligand 6/TGF-β and recruits immunosuppressive N2
TANs, forming a pro-tumorigenic feedback loop. Conversely, in lung
cancer, C-X-C chemokine receptor type 2 (CXCR2) inhibition shifts
TANs toward an N1 phenotype, decreasing immunosuppressive factors
(arginase 1, TGF-β) and enhancing CD8+ T-cell activity,
thereby improving antitumor immunity and chemotherapy response
(34,35). These findings highlight TAN
polarization as a critical determinant of tumor fate, suggesting
therapeutic strategies targeting N2 TANs (such asCLCF1/ERK
inhibition in HCC) or promoting N1 TANs (for example via CXCR2
blockade in lung cancer) may modulate the TME.
TILs serve as both prognostic biomarkers in cancer
progression and key effectors in immunotherapeutic responses
(26). The CCL2/CCR2 signaling axis
orchestrates TIL recruitment to the TME, while CCL2 modulates the
anti-cancer functions of T lymphocytes and dendritic cells, thereby
potentiating antitumor immunity (26).
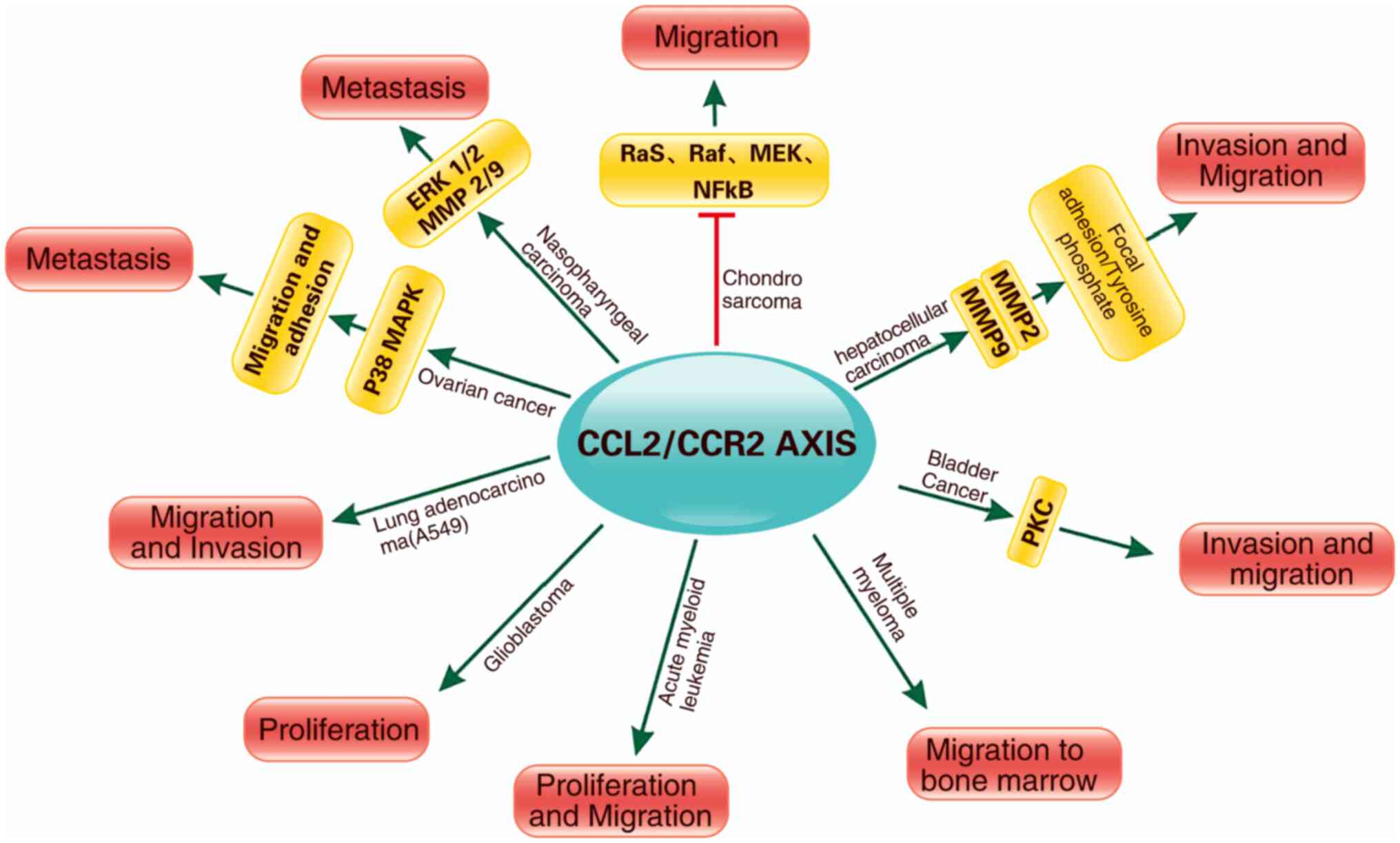
CCL2 promotes chondrosarcoma cell migration by
upregulating MMP9 expression and engaging CCR2, thereby inhibiting
Ras/Raf-1/MEK/ERK and NFκB signaling cascades (53). In HCC cell model, CCL2/CCR2
engagement triggers focal adhesion kinase tyrosine phosphorylation
at Y397, promoting the recruitment of Src family kinases and
activation of MMP2/9 through the ERK1/2 signaling cascade (54). Concurrently, calcium ion flux
mediated by CCR2 activates the calcineurin-nuclear factor of
activated T cells pathway, upregulating MMP9 transcription and
enhancing extracellular matrix degradation (55). Sustained CCL2 secretion also
polarizes tumor-associated macrophages (TAMs) toward the M2
phenotype, fostering a pro-inflammatory milieu rich in IL-6 and
TNF-α that supports cancer cell clonal expansion (56–59).
This multifaceted signaling network has been implicated in
metastatic progression in multiple types of cancer, including
cervical and breast malignancy (56,58).
In breast cancer, the CCL2/CCR2 axis regulates cellular motility
and survival, thereby driving metastatic dissemination (60,61).
Similarly, in prostate cancer, CCL2/CCR2 signaling governs
proliferation, apoptosis resistance and invasive potential
(62). In bladder cancer, the
activation of this axis enhances migration and invasion via protein
kinase C activation and tyrosine phosphorylation, independent of
its effects on cell proliferation (63). Blockade of CCL2/CCR2 signaling
markedly impairs the bone marrow homing of multiple myeloma cells,
underscoring its role in hematological malignancy metastasis
(64). In nasopharyngeal carcinoma
(NPC), CCL2/CCR2 signaling promotes metastasis via ERK1/2-MMP2/9
pathway activation (65). In
colorectal cancer, alcohol exposure upregulates CCL2/CCR2 signaling
via the glycogen synthase kinase 3β/β-catenin pathway, facilitating
metastatic progression (66).
Epithelial ovarian cancer exploits CCL2/CCR2 signaling to promote
peritoneal dissemination, as demonstrated by the enhanced migration
and adhesion of ovarian cancer cells following CCL2 stimulation, a
process mediated by the P38 MAPK pathway (67,68).
The CCL2/CCR2 signaling pathway drives the
proliferation and migration of acute myeloid leukemia cells, as
demonstrated by in vitro and in vivo studies
(69). In glioblastoma,
tumor-secreted CCL2 orchestrates monocyte recruitment, establishing
a pro-tumorigenic microenvironment that fosters neoplastic growth
(70). Leveraging this mechanistic
insight, preclinical investigation have explored CCR2 antagonists
as novel antitumor agents. For example, the pharmacological
blockade of CCR2 in lung adenocarcinoma A549 cells attenuates their
migratory and invasive capabilities (Fig. 1; Table
I) (71).
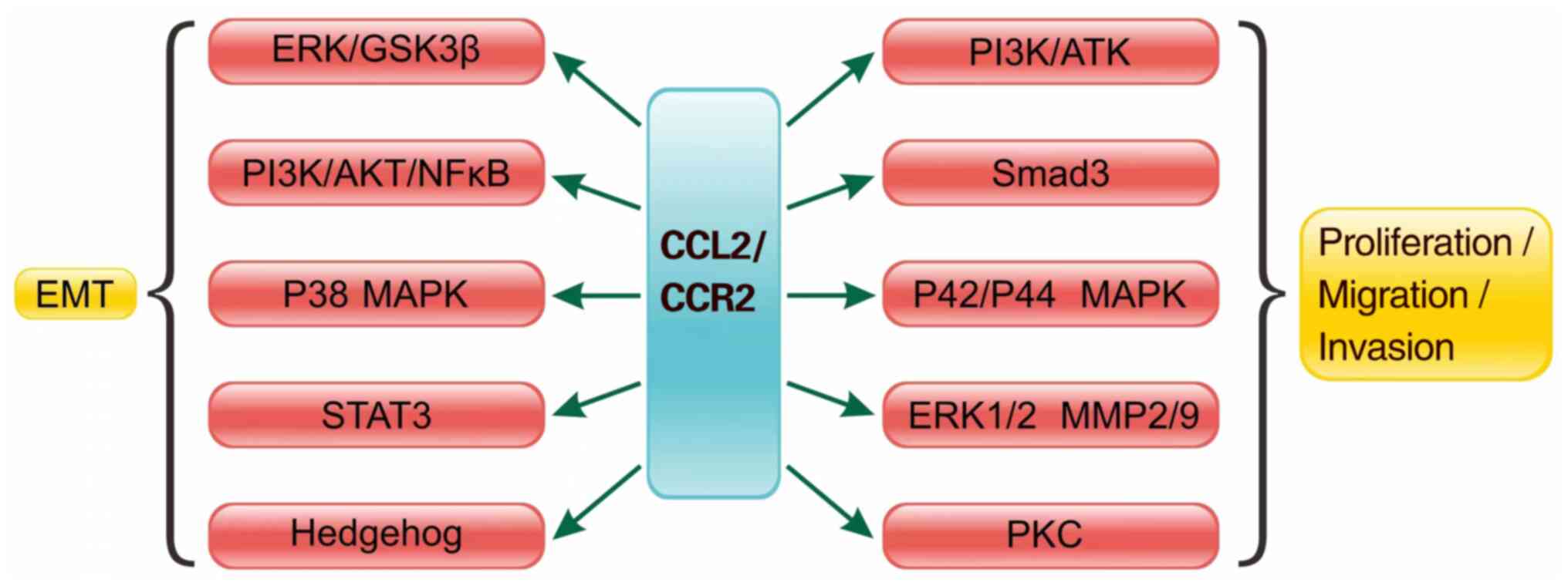
EMT is the initial step of tumor metastasis, and is
involved in the progression and metastasis of various types of
cancer (72,73). The CCL2/CCR2 signaling axis promotes
EMT in liver cancer through MMP2 (74). There is a growing body of data
describing a direct stimulatory effect of CCL2 on tumor epithelial
cells (8,75). Notably, interruption of the
CCL2/CCR2/STAT3 pathway can inhibit the EMT and migration of
prostate cancer cells, inhibiting the progression and metastasis of
prostate cancer (75). Furthermore,
the TME enhances bladder cancer metastasis by modulating estrogen
receptor (ER)β/CCL2/CCR2 EMT/MMP9 signaling following mast cell
recruitment (76). The CCL2/CCR2
pathway induces the invasion and EMT of HCC in vitro by
activating the Hedgehog pathway (Fig.
2) (77).
Tumor angiogenesis requires the production of
angiogenic factors by tumor cells and stromal components. CCL2
directly promotes angiogenesis via the activation of
CCR2-expressing vascular endothelial cells (78–80).
Concurrently, the CCL2/CCR2 signaling axis enhances vascular
permeability, facilitating efficient extravasation of tumor cells
and metastatic niche formation (81). Human endothelial cells express CCR2,
which promotes tumor angiogenesis and progression after binding to
CCL2. Several clinical studies have shown that CCL2 may be a
biomarker of tumor angiogenesis (82–84).
The activation of the CCL2/CCR2 signaling axis in the TME promotes
tumor angiogenesis (83). CCL2
directly interacts with CCR2 on the surface of endothelial cells,
resulting in increased vessel sprouting and angiogenesis (84). As a chemokine produced in abundance
by certain types of tumor, such as hepatocellular carcinoma (HCC),
glioblastoma, and Small Cell Lung Cancer (SCLC) (85), it can also directly promote tumor
progression. Therefore, therapy using MCP-1 antagonists in
combination with other angiogenesis inhibitors may suppress tumor
growth (86).
In contrast to earlier assumptions, emerging
evidence has indicated that tumor vascular endothelial cells lack
CCR2 expression (87,88). Instead, CCL2 drives angiogenesis
indirectly by recruiting TAMs and increasing VEGF-A production in
these cells (89–92). In addition, CCL2 enhances cancer
cell autonomous VEGF secretion, further contributing to
neovascularization. In melanoma, this axis is amplified by
autocrine/paracrine loops, wherein CCL2 and its receptor CCR2 are
co-expressed (93,94). AM-derived TNFα and IL-1α synergize
with VEGF to promote endothelial cell activation, thereby
accelerating early-stage tumor angiogenesis and growth (9).
The TME is a dynamically complex ecosystem, where
inflammatory networks composed of immune cells and their secretory
products influence cancer biology and progression (95). Chemokine-chemokine receptor
interactions are key to recruiting inflammatory cells into the TME,
with CCL2 serving as a key driver of inflammatory monocyte
accumulation (96). Inflammatory
monocytes exhibit high expression of CCR2 (26), while other CCR2-expressing
leukocytes, including CD8+ effector T cells and
CD4+ regulatory T cells (Tregs) (19,97)
and myeloid-derived suppressor cells (MDSCs) (98), are enriched in inflamed tumor
tissues. The CCL2/CCR2 pathway orchestrates metastatic
microenvironment formation, exerting pro-tumorigenic and
pro-metastatic functions in numerous types of cancer (99,100).
In sarcoma and breast cancer, the CCL2/CCR2 axis
mediates the recruitment of TAMs and MDSCs to the TME (89). T cells infiltrate tumors in an
antigen-specific manner, with IFN-γ secreted by early-invading T
cells inducing tissue macrophages to increase CCL2 expression
(101,102). This creates a positive feedback
loop wherein CCL2 recruits additional T cells and macrophages via
CCR2 signaling, enhancing immune cell infiltration. CCR2 expression
is detectable in tumor-infiltrating immune cells, supporting the
role of active chemotactic recruitment (101,102). In NPC, this mechanism is
exemplified by T cell-derived IFN-γ activating macrophages to
secrete CCL2, thereby amplifying T-cell and macrophage accumulation
via the CCL2/CCR2 pathway (103).
CAFs, as a key component of the TME, represent
activated fibroblast populations that impact stromal compartment
remodeling within the TME via collagen deposition and MMP secretion
(104). CAFs constitute one of the
most prevalent cell types in the tumor stroma (105), and evidence has highlighted their
tumor-promoting functions, including accelerating tumor
proliferation, facilitating metastatic progression and shielding
tumors from therapeutic agent penetration (106–108).
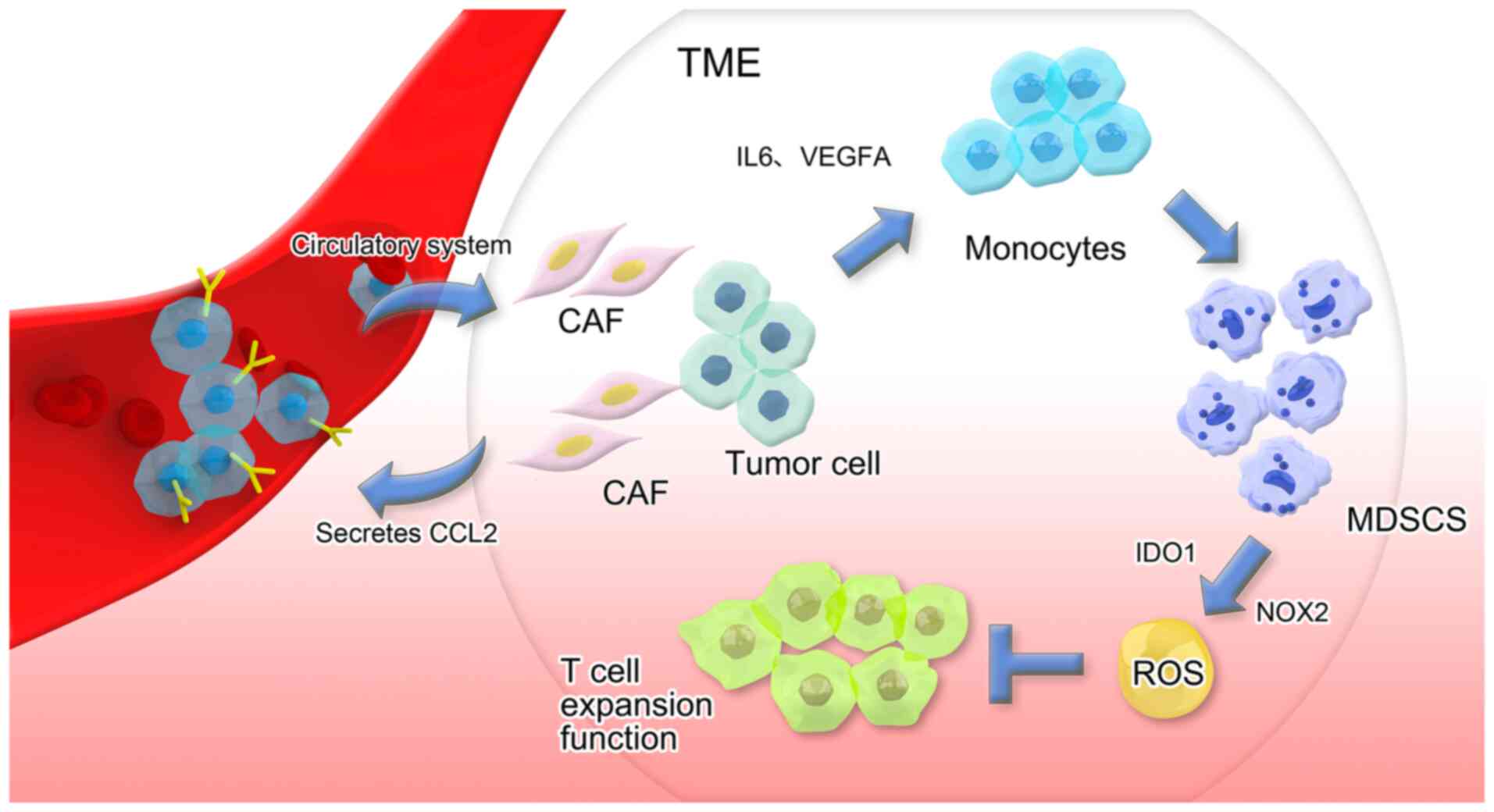
Accumulating evidence has highlighted the crosstalk
between CAFs and immunosuppressive cell lineages, which is
primarily due to the immune-modulatory functions of CAFs (109–111). CAFs secreting CCL2 facilitate the
recruitment of CCR2+ monocytes from the bloodstream into
the TME, where direct cell-cell interactions drive monocyte
differentiation into MDSCs. CAF-educated MDSCs suppress T-cell
proliferation via upregulation of NADPH oxidase 2 and indoleamine
2,3-dioxygenase 1, leading to excessive reactive oxygen species
production that inhibits immune effector function (Fig. 3) (112).
Crosstalk with cancer cells drives CCL2 upregulation
in CAFs, which contributes to metastatic niche establishment during
early tumor progression and modulates broader tumor functionality
(113–118). In a murine liver tumor model,
fibroblast STAT3/CCL2 signaling has been shown to enhance MDSC
recruitment, thereby promoting tumor growth (119). In addition, the CCL2/CCR2
signaling pathway facilitates early breast cancer survival and
invasion via a fibroblast-mediated mechanism (120).
A previous study demonstrated that discoidin domain
receptor 1 (DDR1) orchestrates an immunosuppressive TME by
activating CAFs to secrete CCL2 and IL-6. This signaling axis
exhibits dual regulatory effects by recruiting immunosuppressive
cells, including MDSCs and TAMs, and facilitating extracellular
matrix remodeling through enhanced collagen deposition and MMP9
activation. These findings demonstrate the key role of the
DDR1/CCL2/IL-6 pathway in CAF-mediated immunosuppression, providing
a potential therapeutic strategy to target tumor fibrosis and
immune evasion (121,122).
CCL2 primarily regulates the directional migration
and invasive infiltration of reticuloendothelial system cells, with
a focus on monocyte/macrophage phenotypes (123). CCL2 induces monocytes to exit the
bloodstream and extravasate into peripheral tissue (71), where they differentiate into
tissue-resident macrophages. The CCL2/CCR2 signaling pathway has
been implicated in recruiting macrophages in various types of human
cancer, including those originating in the bladder, cervix, ovary,
lung and breast (124–127). TAMs, a major component of
infiltrating inflammatory cells, are regulated by the CCL2/CCR2
pathway through a macrophage-dependent mechanism sustained by
positive feedback loops (61,128).
As a key chemokine system mediating blood cell recruitment,
particularly macrophages, into tissues (11,128–130), CCL2/CCR2 signaling drives tumor
progression. M2-polarized TAMs influence tumor progression and
metastasis (131), and CCL2
secretion recruits TAMs that mediate metastatic phenotypes in
ER-negative breast cancer (64).
TAMs drive prostate cancer metastasis via activation
of the CCL2/CCR2 signaling pathway (132). Phenotypically, tumor-infiltrating
macrophages can be tumor-supportive (M2) or function in tumor
immune surveillance (M1). TAMs of the M1 type lead to a better
prognosis, whereas TAMs of the M2 type lead to a poorer prognosis.
Tumors associated with M2-type TAMs include breast, ovarian and
prostate cancer (133–135). TAM recruitment is dependent on the
CCL2/CCR2 signaling axis and the formation of a tumor-supportive
microenvironment depends on altered cellular dynamics following the
interaction of CCL2, T cells and monocytes (136).
TAMs serve a pivotal role in driving hormone
resistance in prostate cancer cells (137). Within the prostate TME,
M2-polarized TAMs exert pro-tumorigenic effects. Previous studies
(78,138) employed a PC3 cell xenograft model
to show that CCL2 increases in vivo prostate tumor growth
and metastasis by enhancing TAM recruitment and angiogenesis.
Investigations (139,140) have also revealed that M2-phenotype
TAMs influence cancer progression and metastasis; in human lung
cancer, CCL2/CCR2 signaling promotes tumor cell proliferation,
migration and M2 polarization of TAMs (131). In pancreatic ductal
adenocarcinoma, the CCL2/CCR2 axis recruits TAMs to establish an
immunosuppressive TME (141).
Conversely, anti-CCL2 antibody treatment in breast cancer xenograft
models has been shown to diminish macrophage infiltration and tumor
growth (142,143).
The CCL2/CCR2 pathway is indispensable for
monocyte/macrophage recruitment. The therapeutic interruption of
this pathway suppresses inflammatory monocyte recruitment, TAM
infiltration and M2 polarization, thereby reversing the
immunosuppressive state of the TME and activating antitumor
CD8+ T-cell responses (39). Platelet-derived growth
factor-BB-mediated autocrine signaling drives CCL2 secretion, which
recruits macrophages via the CCL2/CCR2 axis to facilitate lung
cancer cell invasion (144).
Targeting CCL2/CCR2 signaling in tumor-infiltrating macrophages has
emerged as a promising therapeutic strategy for HCC (145). Preclinical studies (26,93,146,147) have also shown that genetic
knockout of CCR2 or pharmacological blockade with CCR2 antagonists
inhibits malignant growth and metastasis, decreases postoperative
recurrence and improves survival rates. In pancreatic
adenocarcinoma, cancer cells exploit chemokine pathways,
particularly CCL2/CCR2, to establish an immunosuppressive niche
(146). Blocking TAM recruitment
inhibits murine breast cancer growth (148). In addition, the JAK2/STAT3
signaling pathway promotes M2-like macrophage polarization, which
drives gastric cancer metastasis via EMT (149).
Tregs, a specialized subset of T cells, are key for
maintaining peripheral self-tolerance and preventing
immunopathological responses (150). Tumors exploit immune evasion
mechanisms to sustain uncontrolled growth, with high intratumoral
Treg abundance contributing to the establishment of
immunosuppressive microenvironments. Tregs exert suppressive
effects on T-cell proliferation and cytotoxic function (151,152). The infiltration of Tregs in tumors
is influenced by both in situ generation (mediated by
cytokine secretion) and peripheral recruitment (driven by chemokine
signaling) (153). Tumor-derived
CCL2 has been implicated in inducing Treg migration into the TME.
For example, in glioma local immunosuppression is enhanced by
selectively recruiting CCL2/CCR2-dependent Tregs (129), while colorectal cancer cells
secrete CCL2 that binds to CCR2 on cytotoxic T lymphocytes (CTLs),
paradoxically promoting CTL migration to tumors (44).
Tumors employ diverse evasion strategies to
circumvent immune recognition and elimination. Intratumoral
immunosuppressive MDSCs represent a heterogeneous population of
immature myeloid cells originating from bone marrow progenitors
(154–156). MDSCs exert multifaceted
tumor-promoting activities (157).
The CCL2/CCR2 signaling pathway governs the recruitment of
myelosuppressive cells to tumor sites. Studies have demonstrated
that CCR2 is universally expressed on MDSCs, the primary drivers of
tumor immune evasion, and blocking CCL2/CCR2 signaling inhibits
MDSC migration and tumor growth facilitated by these cells
(31,112,158–160). Collectively, these data
demonstrate a key role for the CCL2/CCR2 pathway in regulating MDSC
trafficking. CCL2/CCR2 interactions also recruit other
immunosuppressive cells, including monocytes, to form a
pro-metastatic microenvironment (26,98).
The immunosuppressive Treg-MDSC network drives
immune exclusion and immune checkpoint inhibitor (ICI) resistance.
In head and neck squamous cell carcinoma, semaphorin (SEMA)4D
blockade using pepinemab disrupts MDSC recruitment while enhancing
T-cell infiltration (KEYNOTE-B84 trial) (161). In bladder cancer, gemcitabine/BCG
decreases IL-6-mediated MDSC suppression. Prostate cancer subtyping
has revealed TGF-β-enriched, immune-excluded tumors (stage I) with
a poor ICI response vs. inflamed subtypes (S-IV). Targeting this
axis (via SEMA4D inhibition or myeloid modulation) represents a
promising therapeutic strategy (162).
The CCL2/CCR2 signaling pathway serves a central
role in shaping the TME by regulating both tumor progression and
antitumor immunity (2). Through the
activation of the PI3K/AKT, MAPK and EMT pathways (Fig. 2), the CCL2/CCR2 signaling pathway
promotes tumor cell proliferation, invasion and metastasis.
Paradoxically, it also mediates antitumor effects by recruiting
immune cells and activating immunosurveillance mechanisms (143). This dual functionality stems from
its ability to recruit diverse immune populations. While MDSCs,
TAMs and Tregs establish an immunosuppressive TME that facilitates
metastasis, the CCL2/CCR2 pathway simultaneously promotes TIL
infiltration and enhances antitumor lymphocyte function (Fig. 4) (163). In melanoma, CCL2/CCR2 signaling
drives resistance to BRAF/MEK inhibitors by expanding MDSCs and
suppressing CD8+ T cells (164–168). The TNF-related apoptosis-inducing
ligand-CCL2 axis further reinforces this resistance by polarizing
monocytes toward MDSCs and M2-like macrophages, facilitating tumor
progression (169,170).
The therapeutic targeting of the CCL2/CCR2 axis
involves multiple strategies with distinct mechanisms and clinical
implications. Small-molecule CCR2 antagonists (such as PF-04136309
and BMS-813160) block immunosuppressive MDSC/TAM recruitment and
restore antitumor immunity in preclinical models (164,165,169,176), with ongoing clinical trials
(177,178) evaluating their efficacy in
combination with chemotherapy or immunotherapy (trial no.
NCT03184870). While anti-CCL2 antibodies (carlumab/CNTO-888) have
demonstrated partial responses in early trials, compensatory CCL2
upregulation limits their efficacy, prompting the exploration of
combination strategies with programmed cell death-1 (PD-1)/CTLA-4
inhibitors to overcome resistance (172). Dual targeting approaches
simultaneously inhibiting CCL2 and CCR2 (CCL2-trapping agents and
CCR2 antagonists) may prevent microenvironmental bypass mechanisms,
particularly in BRAF inhibitor-resistant melanoma (166–168). Furthermore, elevated CCL2/MDSC
signatures could serve as predictive biomarkers for patient
selection, with studies (179–181) investigating liquid biopsy-based
monitoring (CCR2+ exosomes) (170).
The net biological effect of the CCL2/CCR2 axis is
fundamentally context-dependent, governed by four intersecting
factors: i) Temporal dynamics, where early-phase immune
surveillance progressively shifts to late-stage TME remodeling; ii)
spatial heterogeneity across distinct tumor niches; iii) immune
cell composition ratios, particularly the M1/M2 macrophage
equilibrium; and iv) host genetic background, including CCR2
isoform expression patterns. This necessitates identification of
the biological threshold where pro-tumor effects supersede
antitumor mechanisms. This may be regulated by three factors: i)
CCL2 concentration thresholds, ii) specific immune cell
infiltration ratio and iii) hypoxia-induced pathway activation
states. These mechanistic insight may inform clinical strategies,
as evidenced by Phase II trial (182) (trial no. NCT03184870) combining
anti-CCL2 agents with PD-1 inhibitors in non-small cell lung
cancer, which aim to therapeutically manipulate this balance.
While preclinical findings have provided
mechanistic insights into the CCL2/CCR2 pathway in cancer
progression, direct clinical validation in human malignancies
remains limited. Although studies (183,184) have investigated CCR2 antagonists
(PF-04136309) or CCL2 inhibitors (CNTO-888), most therapeutic
strategies have focused on an individual target blockade rather
than coordinated modulation of the CCL2/CCR2 interaction. Future
studies should prioritize the following: i) Comprehensive CCL2/CCR2
co-expression profiling in patient-derived samples, ii)
longitudinal assessment of axis activation during CCR2-targeted
therapy and iii) development of novel dual-targeting approaches to
elucidate the clinical relevance of this chemokine axis in human
cancer.
The present study provides a rationale for the
clinical evaluation of CCL2/CCR2 axis inhibitors in combination
with existing immunotherapies, offering potential for
treatment-resistant malignancies, including melanoma. Future
studies should focus on optimizing combination strategies and
identifying predictive biomarkers for patient stratification.
Not applicable.
The present study was supported by the Scientific Research Fund
project of Education Department of Yunnan Province (grant no.
2025J0181).
Not applicable.
YZ, BF, HY and ZY conceived and designed the study.
YZ wrote the manuscript. HY constructed figures. GC and ZH designed
and implemented the comprehensive literature search strategy,
established rigorous eligibility criteria for study inclusion,
synthesized key findings to construct the theoretical framework,
made substantial contributions to both the Introduction and
Discussion sections and actively participated in critical revision
of the manuscript with particular focus on ensuring the accuracy
and optimal presentation of figures. Specifically, GC conceived the
core visual framework for all figures, which was instrumental in
interpreting the study's conceptual approach and ZH established the
comparative logic and data hierarchy in figures, ensuring perfect
alignment with the manuscript's analytical narrative. YiL and XM
acquired full-text articles and independently screened literature
(resolving discrepancies through consensus), organized references
(using EndNote), revised the Discussion section for clarity and
clinical relevance, and oordinated revision feedback among all
co-authors to ensure consistency. TW conducted supplementary
literature searches to validate results, provided visualization
support (including figure and table design), revised the Discussion
section with critical intellectual input and helped coordinate
final revisions. WW, LC, LH, YaL, DL, XC and YY reviewed the
manuscript. Data authentication is not applicable. All authors have
read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Yoshimura T, Robinson EA, Tanaka S,
Appella E, Kuratsu J and Leonard EJ: Purification and amino acid
analysis of two human glioma-derived monocyte chemoattractants. J
Exp Med. 169:1449–1459. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yoshimura T: The production of monocyte
chemoattractant protein-1 (MCP-1)/CCL2 in tumor microenvironments.
Cytokine. 98:71–78. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kurihara T, Warr G, Loy J and Bravo R:
Defects in macrophage recruitment and host defense in mice lacking
the CCR2 chemokine receptor. J Exp Med. 186:1757–1762. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mackay CR: Chemokines: Immunology's high
impact factors. Nat Immunol. 2:95–101. 2001. View Article : Google Scholar
|
|
5
|
Strieter RM: Chemokines: Not just
leukocyte chemoattractants in the promotion of cancer. Nat Immunol.
2:285–286. 2001. View
Article : Google Scholar
|
|
6
|
Lim SY, Yuzhalin AE, Gordon-Weeks AN and
Muschel RJ: Targeting the CCL2-CCR2 signaling axis in cancer
metastasis. Oncotarget. 7:28697–28710. 2016. View Article : Google Scholar
|
|
7
|
Loberg RD, Day LL, Harwood J, Ying C, St
John LN, Giles R, Neeley CK and Pienta KJ: CCL2 is a potent
regulator of prostate cancer cell migration and proliferation.
Neoplasia. 8:578–586. 2006. View Article : Google Scholar
|
|
8
|
Craig MJ and Loberg RD: CCL2 (Monocyte
Chemoattractant Protein-1) in cancer bone metastases. Cancer
Metastasis Rev. 25:611–619. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Koga M, Kai H, Egami K, Murohara T, Ikeda
A, Yasuoka S, Egashira K, Matsuishi T, Kai M, Kataoka Y, et al:
Mutant MCP-1 therapy inhibits tumor angiogenesis and growth of
malignant melanoma in mice. Biochem Biophys Res Commun.
365:279–284. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Q, Zhang H, Jiang X, Qian C, Liu Z and
Luo D: Factors involved in cancer metastasis: A better
understanding to ‘seed and soil’ hypothesis. Mol Cancer.
16:1762017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deshmane SL, Kremlev S, Amini S and Sawaya
BE: Monocyte chemoattractant protein-1 (MCP-1): An overview. J
Interferon Cytokine Res. 29:313–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Charo IF and Ransohoff RM: The many roles
of chemokines and chemokine receptors in inflammation. N Engl J
Med. 354:610–621. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Raman D, Baugher PJ, Thu YM and Richmond
A: Role of chemokines in tumor growth. Cancer Lett. 256:137–165.
2007. View Article : Google Scholar
|
|
14
|
Balkwill FR: The chemokine system and
cancer. J Pathol. 226:148–157. 2012. View Article : Google Scholar
|
|
15
|
Frederick MJ and Clayman GL: Chemokines in
cancer. Expert Rev Mol Med. 3:1–18. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Paget S: The distribution of secondary
growths in cancer of the breast. 1889. Cancer Metastasis Rev.
8:98–101. 1989.PubMed/NCBI
|
|
17
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar
|
|
18
|
de Visser KE and Joyce JA: The evolving
tumor microenvironment: From cancer initiation to metastatic
outgrowth. Cancer Cell. 41:374–403. 2023. View Article : Google Scholar
|
|
19
|
Brown CE, Vishwanath RP, Aguilar B, Starr
R, Najbauer J, Aboody KS and Jensen MC: Tumor-derived chemokine
MCP-1/CCL2 is sufficient for mediating tumor tropism of adoptively
transferred T cells. J Immunol. 179:3332–3341. 2007. View Article : Google Scholar
|
|
20
|
Rollins BJ and Sunday ME: Suppression of
tumor formation in vivo by expression of the JE gene in malignant
cells. Mol Cell Biol. 11:3125–3131. 1991. View Article : Google Scholar
|
|
21
|
Monti P, Leone BE, Marchesi F, Balzano G,
Zerbi A, Scaltrini F, Pasquali C, Calori G, Pessi F, Sperti C, et
al: The CC chemokine MCP-1/CCL2 in pancreatic cancer progression:
regulation of expression and potential mechanisms of antimalignant
activity. Cancer Res. 63:7451–7461. 2003.PubMed/NCBI
|
|
22
|
Gunderson AJ, Yamazaki T, McCarty K, Fox
N, Phillips M, Alice A, Blair T, Whiteford M, O'Brien D, Ahmad R,
et al: TGFβ suppresses CD8(+) T cell expression of CXCR3 and tumor
trafficking. Nat Commun. 11:17492020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qin J, Gong Q, Zhou C, Xu J, Cheng Y, Xu
W, Zhu D, Liu Y, Zhang Y, Wang Y, et al: Differential expression
pattern of CC chemokine receptor 7 guides precision treatment of
hepatocellular carcinoma. Signal Transduct Target Ther. 10:2292025.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei XL, Zhang Y, Zhao HY, Fang WF, Luo HY,
Qiu MZ, He MM, Zou BY, Xie J, Jin CL, et al: Safety and clinical
activity of SHR7390 monotherapy or combined with camrelizumab for
advanced solid tumor: Results from two phase I trials. Oncologist.
28:e36–e44. 2023. View Article : Google Scholar
|
|
25
|
Mestdagt M, Polette M, Buttice G, Noël A,
Ueda A, Foidart JM and Gilles C: Transactivation of MCP-1/CCL2 by
beta-catenin/TCF-4 in human breast cancer cells. Int J Cancer.
118:35–42. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qian BZ, Li J, Zhang H, Kitamura T, Zhang
J, Campion LR, Kaiser EA, Snyder LA and Pollard JW: CCL2 recruits
inflammatory monocytes to facilitate breast-tumour metastasis.
Nature. 475:222–225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kitamura T, Qian BZ, Soong D, Cassetta L,
Noy R, Sugano G, Kato Y, Li J and Pollard JW: CCL2-induced
chemokine cascade promotes breast cancer metastasis by enhancing
retention of metastasis-associated macrophages. J Exp Med.
212:1043–1059. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Granot Z, Henke E, Comen EA, King TA,
Norton L and Benezra R: Tumor entrained neutrophils inhibit seeding
in the premetastatic lung. Cancer Cell. 20:300–314. 2011.
View Article : Google Scholar
|
|
29
|
Pekarek LA, Starr BA, Toledano AY and
Schreiber H: Inhibition of tumor growth by elimination of
granulocytes. J Exp Med. 181:435–440. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shojaei F, Singh M, Thompson JD and
Ferrara N: Role of Bv8 in neutrophil-dependent angiogenesis in a
transgenic model of cancer progression. Proc Natl Acad Sci USA.
105:2640–2645. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Youn JI, Nagaraj S, Collazo M and
Gabrilovich DI: Subsets of myeloid-derived suppressor cells in
tumor-bearing mice. J Immunol. 181:5791–5802. 2008. View Article : Google Scholar
|
|
32
|
Nozawa H, Chiu C and Hanahan D:
Infiltrating neutrophils mediate the initial angiogenic switch in a
mouse model of multistage carcinogenesis. Proc Natl Acad Sci USA.
103:12493–12498. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
De Larco JE, Wuertz BR and Furcht LT: The
potential role of neutrophils in promoting the metastatic phenotype
of tumors releasing interleukin-8. Clin Cancer Res. 10:4895–4900.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng Y, Mo F, Li Q, Han X, Shi H, Chen S,
Wei Y and Wei X: Targeting CXCR2 inhibits the progression of lung
cancer and promotes therapeutic effect of cisplatin. Mol Cancer.
20:622021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Song M, He J, Pan QZ, Yang J, Zhao J,
Zhang YJ, Huang Y, Tang Y, Wang Q, He J, et al: Cancer-associated
fibroblast-mediated cellular crosstalk supports hepatocellular
carcinoma progression. Hepatology. 73:1717–1735. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Dutta P, Sarkissyan M, Paico K, Wu Y and
Vadgama JV: MCP-1 is overexpressed in triple-negative breast
cancers and drives cancer invasiveness and metastasis. Breast
Cancer Res Treat. 170:477–486. 2018. View Article : Google Scholar
|
|
37
|
Parween F, Singh SP, Kathuria N, Zhang HH,
Ashida S, Otaizo-Carrasquero FA, Shamsaddini A, Gardina PJ, Ganesan
S, Kabat J, et al: Migration arrest and transendothelial
trafficking of human pathogenic-like Th17 cells are mediated by
differentially positioned chemokines. Nat Commun. 16:19782025.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dey D, Pal S, Chakraborty BC, Baidya A,
Bhadra S, Ghosh R, Banerjee S, Ahammed SKM, Chowdhury A and Datta
S: Multifaceted defects in monocytes in different phases of chronic
hepatitis B virus infection: Lack of restoration after antiviral
therapy. Microbiol Spectr. 10:e01939222022. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li X, Yao W, Yuan Y, Chen P, Li B, Li J,
Chu R, Song H, Xie D, Jiang X and Wang H: Targeting of
tumour-infiltrating macrophages via CCL2/CCR2 signalling as a
therapeutic strategy against hepatocellular carcinoma. Gut.
66:157–167. 2017. View Article : Google Scholar
|
|
40
|
Guerra N, Tan YX, Joncker NT, Choy A,
Gallardo F, Xiong N, Knoblaugh S, Cado D, Greenberg NM and Raulet
DH: NKG2D-deficient mice are defective in tumor surveillance in
models of spontaneous malignancy. Immunity. 28:571–580. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lujambio A, Akkari L, Simon J, Grace D,
Tschaharganeh DF, Bolden JE, Zhao Z, Thapar V, Joyce JA,
Krizhanovsky V and Lowe SW: Non-cell-autonomous tumor suppression
by p53. Cell. 153:449–460. 2013. View Article : Google Scholar
|
|
42
|
Iannello A, Thompson TW, Ardolino M, Lowe
SW and Raulet DH: p53-dependent chemokine production by senescent
tumor cells supports NKG2D-dependent tumor elimination by natural
killer cells. J Exp Med. 210:2057–2069. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nagai M and Masuzawa T: Vaccination with
MCP-1 cDNA transfectant on human malignant glioma in nude mice
induces migration of monocytes and NK cells to the tumor. Int
Immunopharmacol. 1:657–664. 2001. View Article : Google Scholar
|
|
44
|
Berencsi K, Rani P, Zhang T, Gross L,
Mastrangelo M, Meropol NJ, Herlyn D and Somasundaram R: In vitro
migration of cytotoxic T lymphocyte derived from a colon carcinoma
patient is dependent on CCL2 and CCR2. J Transl Med. 9:332011.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Nakasone Y, Fujimoto M, Matsushita T,
Hamaguchi Y, Huu DL, Yanaba M, Sato S, Takehara K and Hasegawa M:
Host-derived MCP-1 and MIP-1α regulate protective anti-tumor
immunity to localized and metastatic B16 melanoma. Am J Pathol.
180:365–374. 2012. View Article : Google Scholar
|
|
46
|
Joyce JA and Pollard JW:
Microenvironmental regulation of metastasis. Nat Rev Cancer.
9:239–252. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang J, Sarkar S, Cua R, Zhou Y, Hader W
and Yong VW: A dialog between glioma and microglia that promotes
tumor invasiveness through the CCL2/CCR2/interleukin-6 axis.
Carcinogenesis. 33:312–319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bogenrieder T and Herlyn M: Axis of evil:
molecular mechanisms of cancer metastasis. Oncogene. 22:6524–6536.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rafei M, Deng J, Boivin MN, Williams P,
Matulis SM, Yuan S, Birman E, Forner K, Yuan L, Castellino C, et
al: A MCP1 fusokine with CCR2-specific tumoricidal activity. Mol
Cancer. 10:1212011. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Johnson Z, Power CA, Weiss C, Rintelen F,
Ji H, Ruckle T, Camps M, Wells TN, Schwarz MK, Proudfoot AE and
Rommel C: Chemokine inhibition-why, when, where, which and how?
Biochem Soc Trans. 32:366–377. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mellado M, Rodríguez-Frade JM, Aragay A,
del Real G, Martín AM, Vila-Coro AJ, Serrano A, Mayor F Jr and
Martínez-A C: The chemokine monocyte chemotactic protein 1 triggers
Janus kinase 2 activation and tyrosine phosphorylation of the CCR2B
receptor. J Immunol. 161:805–813. 1998. View Article : Google Scholar
|
|
52
|
Jiménez-Sainz MC, Fast B, Mayor F Jr and
Aragay AM: Signaling pathways for monocyte chemoattractant protein
1-mediated extracellular signal-regulated kinase activation. Mol
Pharmacol. 64:773–782. 2003. View Article : Google Scholar
|
|
53
|
Tang CH and Tsai CC: CCL2 increases MMP-9
expression and cell motility in human chondrosarcoma cells via the
Ras/Raf/MEK/ERK/NF-κB signaling pathway. Biochem Pharmacol.
83:335–344. 2012. View Article : Google Scholar
|
|
54
|
Dagouassat M, Suffee N, Hlawaty H, Haddad
O, Charni F, Laguillier C, Vassy R, Martin L, Schischmanoff PO,
Gattegno L, et al: Monocyte chemoattractant protein-1 (MCP-1)/CCL2
secreted by hepatic myofibroblasts promotes migration and invasion
of human hepatoma cells. Int J Cancer. 126:1095–1108. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Schröer N, Pahne J, Walch B, Wickenhauser
C and Smola S: Molecular pathobiology of human cervical high-grade
lesions: Paracrine STAT3 activation in tumor-instructed myeloid
cells drives local MMP-9 expression. Cancer Res. 71:87–97. 2011.
View Article : Google Scholar
|
|
56
|
Balkwill F and Mantovani A: Inflammation
and cancer: Back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar
|
|
57
|
Pollard JW: Tumour-educated macrophages
promote tumour progression and metastasis. Nat Rev Cancer. 4:71–78.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
van Kempen LC, de Visser KE and Coussens
LM: Inflammation, proteases and cancer. Eur J Cancer. 42:728–734.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yumimoto K, Sugiyama S, Mimori K and
Nakayama KI: Potentials of C-C motif chemokine 2-C-C chemokine
receptor type 2 blockers including propagermanium as anticancer
agents. Cancer Sci. 110:2090–2099. 2019. View Article : Google Scholar
|
|
60
|
Fang WB, Jokar I, Zou A, Lambert D,
Dendukuri P and Cheng N: CCL2/CCR2 chemokine signaling coordinates
survival and motility of breast cancer cells through Smad3 protein-
and p42/44 mitogen-activated protein kinase (MAPK)-dependent
mechanisms. J Biol Chem. 287:36593–36608. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Yao M, Fang W, Smart C, Hu Q, Huang S,
Alvarez N, Fields P and Cheng N: CCR2 chemokine receptors enhance
growth and cell-cycle progression of breast cancer cells through
SRC and PKC Activation. Mol Cancer Res. 17:604–617. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Gao J, Wang A, Zhang M, Li H, Wang K, Han
Y, Wang Z, Shi C and Wang W: RNAi targeting of CCR2 gene expression
induces apoptosis and inhibits the proliferation, migration, and
invasion of PC-3M cells. Oncol Res. 21:73–82. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chiu HY, Sun KH, Chen SY, Wang HH, Lee MY,
Tsou YC, Jwo SC, Sun GH and Tang SJ: Autocrine CCL2 promotes cell
migration and invasion via PKC activation and tyrosine
phosphorylation of paxillin in bladder cancer cells. Cytokine.
59:423–432. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Vande Broek I, Asosingh K, Vanderkerken K,
Straetmans N, Van Camp B and Van Riet I: Chemokine receptor CCR2 is
expressed by human multiple myeloma cells and mediates migration to
bone marrow stromal cell-produced monocyte chemotactic proteins
MCP-1, −2 and −3. Br J Cancer. 88:855–862. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Yang J, Lv X, Chen J, Xie C, Xia W, Jiang
C, Zeng T, Ye Y, Ke L, Yu Y, et al: CCL2-CCR2 axis promotes
metastasis of nasopharyngeal carcinoma by activating ERK1/2-MMP2/9
pathway. Oncotarget. 7:15632–15647. 2016. View Article : Google Scholar
|
|
66
|
Xu M, Wang S, Qi Y, Chen L, Frank JA, Yang
XH, Zhang Z, Shi X and Luo J: Role of MCP-1 in alcohol-induced
aggressiveness of colorectal cancer cells. Mol Carcinog.
55:1002–1011. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yasui H, Kajiyama H, Tamauchi S, Suzuki S,
Peng Y, Yoshikawa N, Sugiyama M, Nakamura K and Kikkawa F: CCL2
secreted from cancer-associated mesothelial cells promotes
peritoneal metastasis of ovarian cancer cells through the P38-MAPK
pathway. Clin Exp Metastasis. 37:145–158. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Furukawa S, Soeda S, Kiko Y, Suzuki O,
Hashimoto Y, Watanabe T, Nishiyama H, Tasaki K, Hojo H, Abe M and
Fujimori K: MCP-1 promotes invasion and adhesion of human ovarian
cancer cells. Anticancer Res. 33:4785–4790. 2013.PubMed/NCBI
|
|
69
|
Macanas-Pirard P, Quezada T, Navarrete L,
Broekhuizen R, Leisewitz A, Nervi B and Ramírez PA: The CCL2/CCR2
axis affects transmigration and proliferation but not resistance to
chemotherapy of acute myeloid leukemia cells. PLoS One.
12:e01688882017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Salacz ME, Kast RE, Saki N, Brüning A,
Karpel-Massler G and Halatsch ME: Toward a noncytotoxic
glioblastoma therapy: Blocking MCP-1 with the MTZ Regimen. Onco
Targets Ther. 9:2535–2545. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
An J, Xue Y, Long M, Zhang G, Zhang J and
Su H: Targeting CCR2 with its antagonist suppresses viability,
motility and invasion by downregulating MMP-9 expression in
non-small cell lung cancer cells. Oncotarget. 8:39230–39240. 2017.
View Article : Google Scholar
|
|
72
|
Giannelli G, Koudelkova P, Dituri F and
Mikulits W: Role of epithelial to mesenchymal transition in
hepatocellular carcinoma. J Hepatol. 65:798–808. 2016. View Article : Google Scholar
|
|
73
|
van Zijl F, Zulehner G, Petz M, Schneller
D, Kornauth C, Hau M, Machat G, Grubinger M, Huber H and Mikulits
W: Epithelial-mesenchymal transition in hepatocellular carcinoma.
Future Oncol. 5:1169–1179. 2009. View Article : Google Scholar
|
|
74
|
Li H, Li H, Li XP, Zou H, Liu L, Liu W and
Duan T: C-C chemokine receptor type 2 promotes
epithelial-to-mesenchymal transition by upregulating matrix
metalloproteinase-2 in human liver cancer. Oncol Rep. 40:2734–2741.
2018.PubMed/NCBI
|
|
75
|
Izumi K, Fang LY, Mizokami A, Namiki M, Li
L, Lin WJ and Chang C: Targeting the androgen receptor with siRNA
promotes prostate cancer metastasis through enhanced macrophage
recruitment via CCL2/CCR2-induced STAT3 activation. EMBO Mol Med.
5:1383–1401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Rao Q, Chen Y, Yeh CR, Ding J, Li L, Chang
C and Yeh S: Recruited mast cells in the tumor microenvironment
enhance bladder cancer metastasis via modulation of ERβ/CCL2/CCR2
EMT/MMP9 signals. Oncotarget. 7:7842–7855. 2016. View Article : Google Scholar
|
|
77
|
Zhuang H, Cao G, Kou C and Liu T:
CCL2/CCR2 axis induces hepatocellular carcinoma invasion and
epithelial-mesenchymal transition in vitro through activation of
the Hedgehog pathway. Oncol Rep. 39:21–30. 2018.PubMed/NCBI
|
|
78
|
Koide N, Nishio A, Sato T, Sugiyama A and
Miyagawa S: Significance of macrophage chemoattractant protein-1
expression and macrophage infiltration in squamous cell carcinoma
of the esophagus. Am J Gastroenterol. 99:1667–1674. 2004.
View Article : Google Scholar
|
|
79
|
Salcedo R, Ponce ML, Young HA, Wasserman
K, Ward JM, Kleinman HK, Oppenheim JJ and Murphy WJ: Human
endothelial cells express CCR2 and respond to MCP-1: direct role of
MCP-1 in angiogenesis and tumor progression. Blood. 96:34–40. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Hong KH, Ryu J and Han KH: Monocyte
chemoattractant protein-1-induced angiogenesis is mediated by
vascular endothelial growth factor-A. Blood. 105:1405–1407. 2025.
View Article : Google Scholar
|
|
81
|
Wyler L, Napoli CU, Ingold B, Sulser T,
Heikenwälder M, Schraml P and Moch H: Brain metastasis in renal
cancer patients: Metastatic pattern, tumour-associated macrophages
and chemokine/chemoreceptor expression. Br J Cancer. 110:686–694.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Valković T, Babarović E, Lučin K, Štifter
S, Aralica M, Seili-Bekafigo I, Duletić-Načinović A and Jonjić N:
Plasma levels of monocyte chemotactic protein-1 are associated with
clinical features and angiogenesis in patients with multiple
myeloma. Biomed Res Int. 2016:78705902016.
|
|
83
|
Li F, Kitajima S, Kohno S, Yoshida A,
Tange S, Sasaki S, Okada N, Nishimoto Y, Muranaka H, Nagatani N, et
al: Retinoblastoma inactivation induces a protumoral
microenvironment via enhanced CCL2 secretion. Cancer Res.
79:3903–3915. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Li X, Loberg R, Liao J, Ying C, Snyder LA,
Pienta KJ and McCauley LK: A destructive cascade mediated by CCL2
facilitates prostate cancer growth in bone. Cancer Res.
69:1685–1692. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Balkwill F: Cancer and the chemokine
network. Nat Rev Cancer. 4:540–550. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Niu J, Azfer A, Zhelyabovska O, Fatma S
and Kolattukudy PE: Monocyte chemotactic protein (MCP)-1 promotes
angiogenesis via a novel transcription factor, MCP-1-induced
protein (MCPIP). J Biol Chem. 283:14542–14551. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ohta M, Kitadai Y, Tanaka S, Yoshihara M,
Yasui W, Mukaida N, Haruma K and Chayama K: Monocyte
chemoattractant protein-1 expression correlates with macrophage
infiltration and tumor vascularity in human esophageal squamous
cell carcinomas. Int J Cancer. 102:220–224. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Kuroda T, Kitadai Y, Tanaka S, Yang X,
Mukaida N, Yoshihara M and Chayama K: Monocyte chemoattractant
protein-1 transfection induces angiogenesis and tumorigenesis of
gastric carcinoma in nude mice via macrophage recruitment. Clin
Cancer Res. 11:7629–7636. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Goede V, Brogelli L, Ziche M and Augustin
HG: Induction of inflammatory angiogenesis by monocyte
chemoattractant protein-1. Int J Cancer. 82:765–770. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Low-Marchelli JM, Ardi VC, Vizcarra EA,
van Rooijen N, Quigley JP and Yang J: Twist1 induces CCL2 and
recruits macrophages to promote angiogenesis. Cancer Res.
73:662–671. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Varney ML, Olsen KJ, Mosley RL and Singh
RK: Paracrine regulation of vascular endothelial growth factor-a
expression during macrophage-melanoma cell interaction: Role of
monocyte chemotactic protein-1 and macrophage colony-stimulating
factor. J Interferon Cytokine Res. 25:674–683. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Jetten N, Verbruggen S, Gijbels MJ, Post
MJ, De Winther MP and Donners MM: Anti-inflammatory M2, but not
pro-inflammatory M1 macrophages promote angiogenesis in vivo.
Angiogenesis. 17:109–118. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Bonapace L, Coissieux MM, Wyckoff J, Mertz
KD, Varga Z, Junt T and Bentires-Alj M: Cessation of CCL2
inhibition accelerates breast cancer metastasis by promoting
angiogenesis. Nature. 515:130–133. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Liu L, Li Y and Li B: Interactions between
cancer cells and tumor-associated macrophages in tumor
microenvironment. Biochim Biophys Acta Rev Cancer. 1880:1893442025.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Biswas SK and Mantovani A: Macrophage
plasticity and interaction with lymphocyte subsets: Cancer as a
paradigm. Nat Immunol. 11:889–896. 2010. View Article : Google Scholar
|
|
96
|
Allavena P, Germano G, Marchesi F and
Mantovani A: Chemokines in cancer related inflammation. Exp Cell
Res. 317:664–673. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Fridlender ZG, Buchlis G, Kapoor V, Cheng
G, Sun J, Singhal S, Crisanti MC, Wang LC, Heitjan D, Snyder LA and
Albelda SM: CCL2 blockade augments cancer immunotherapy. Cancer
Res. 70:109–118. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Huang B, Lei Z, Zhao J, Gong W, Liu J,
Chen Z, Liu Y, Li D, Yuan Y, Zhang GM and Feng ZH: CCL2/CCR2
pathway mediates recruitment of myeloid suppressor cells to
cancers. Cancer Lett. 252:86–92. 2007. View Article : Google Scholar
|
|
99
|
Samaniego R, Estecha A, Relloso M, Longo
N, Escat JL, Longo-Imedio I, Avilés JA, del Pozo MA, Puig-Kröger A
and Sánchez-Mateos P: Mesenchymal contribution to recruitment,
infiltration, and positioning of leukocytes in human melanoma
tissues. J Invest Dermatol. 133:2255–2264. 2013. View Article : Google Scholar
|
|
100
|
Roblek M, Strutzmann E, Zankl C, Adage T,
Heikenwalder M, Atlic A, Weis R, Kungl A and Borsig L: Targeting of
CCL2-CCR2-glycosaminoglycan axis using a CCL2 decoy protein
attenuates metastasis through inhibition of tumor cell seeding.
Neoplasia. 18:49–59. 2016. View Article : Google Scholar
|
|
101
|
Zhu Y, Yan P, Wang R, Lai J, Tang H, Xiao
X, Yu R, Bao X, Zhu F, Wang K, et al: Opioid-induced fragile-like
regulatory T cells contribute to withdrawal. Cell. 186:591–606.e23.
2023. View Article : Google Scholar
|
|
102
|
Kong X, Wu S, Dai X, Yu W, Wang J, Sun Y,
Ji Z, Ma L, Dai X, Chen H, et al: A comprehensive profile of
chemokines in the peripheral blood and vascular tissue of patients
with Takayasu arteritis. Arthritis Res Ther. 24:492022. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Tang KF, Tan SY, Chan SH, Chong SM, Loh
KS, Tan LK and Hu H: A distinct expression of CC chemokines by
macrophages in nasopharyngeal carcinoma: implication for the
intense tumor infiltration by T lymphocytes and macrophages. Hum
Pathol. 32:42–49. 2001. View Article : Google Scholar
|
|
104
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Gok Yavuz B, Gunaydin G, Gedik ME,
Kosemehmetoglu K, Karakoc D, Ozgur F and Guc D: Cancer associated
fibroblasts sculpt tumour microenvironment by recruiting monocytes
and inducing immunosuppressive PD-1(+) TAMs. Sci Rep. 9:31722019.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
LeBleu VS and Kalluri R: A peek into
cancer-associated fibroblasts: origins, functions and translational
impact. Dis Model Mech. 11:dmm0294472018. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Loeffler M, Krüger JA, Niethammer AG and
Reisfeld RA: Targeting tumor-associated fibroblasts improves cancer
chemotherapy by increasing intratumoral drug uptake. J Clin Invest.
116:1955–1962. 2006. View Article : Google Scholar
|
|
108
|
Valkenburg KC, de Groot AE and Pienta KJ:
Targeting the tumour stroma to improve cancer therapy. Nat Rev Clin
Oncol. 15:366–381. 2018. View Article : Google Scholar
|
|
109
|
Costa A, Kieffer Y, Scholer-Dahirel A,
Pelon F, Bourachot B, Cardon M, Sirven P, Magagna I, Fuhrmann L,
Bernard C, et al: Fibroblast heterogeneity and immunosuppressive
environment in human breast cancer. Cancer Cell. 33:463–479.e10.
2018. View Article : Google Scholar
|
|
110
|
Cho H, Seo Y, Loke KM, Kim SW, Oh SM, Kim
JH, Soh J, Kim HS, Lee H, Kim J, et al: Cancer-stimulated CAFs
enhance monocyte differentiation and protumoral TAM activation via
IL6 and GM-CSF secretion. Clin Cancer Res. 24:5407–5421. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Mace TA, Ameen Z, Collins A, Wojcik S,
Mair M, Young GS, Fuchs JR, Eubank TD, Frankel WL, Bekaii-Saab T,
et al: Pancreatic cancer-associated stellate cells promote
differentiation of myeloid-derived suppressor cells in a
STAT3-dependent manner. Cancer Res. 73:3007–3018. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Xiang H, Ramil CP, Hai J, Zhang C, Wang H,
Watkins AA, Afshar R, Georgiev P, Sze MA, Song XS, et al:
Cancer-associated fibroblasts promote immunosuppression by inducing
ROS-generating monocytic MDSCs in lung squamous cell carcinoma.
Cancer Immunol Res. 8:436–450. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Jung DW, Che ZM and Kim J, Kim K, Kim KY,
Williams D and Kim J: Tumor-stromal crosstalk in invasion of oral
squamous cell carcinoma: A pivotal role of CCL7. Int J Cancer.
127:332–344. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Barbai T, Fejős Z, Puskas LG, Tímár J and
Rásó E: The importance of microenvironment: the role of CCL8 in
metastasis formation of melanoma. Oncotarget. 6:29111–29128. 2015.
View Article : Google Scholar
|
|
115
|
Tsuyada A, Chow A, Wu J, Somlo G, Chu P,
Loera S, Luu T, Li AX, Wu X, Ye W, et al: CCL2 mediates cross-talk
between cancer cells and stromal fibroblasts that regulates breast
cancer stem cells. Cancer Res. 72:2768–2779. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Higashino N, Koma YI, Hosono M, Takase N,
Okamoto M, Kodaira H, Nishio M, Shigeoka M, Kakeji Y and Yokozaki
H: Fibroblast activation protein-positive fibroblasts promote tumor
progression through secretion of CCL2 and interleukin-6 in
esophageal squamous cell carcinoma. Lab Invest. 99:777–792. 2019.
View Article : Google Scholar
|
|
117
|
Pausch TM, Aue E, Wirsik NM, Freire Valls
A, Shen Y, Radhakrishnan P, Hackert T, Schneider M and Schmidt T:
Metastasis-associated fibroblasts promote angiogenesis in
metastasized pancreatic cancer via the CXCL8 and the CCL2 axes. Sci
Rep. 10:54202020. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Vickman RE, Broman MM, Lanman NA, Franco
OE, Sudyanti PAG, Ni Y, Ji Y, Helfand BT, Petkewicz J, Paterakos
MC, et al: Heterogeneity of human prostate carcinoma-associated
fibroblasts implicates a role for subpopulations in myeloid cell
recruitment. Prostate. 80:173–185. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Yang X, Lin Y, Shi Y, Li B, Liu W, Yin W,
Dang Y, Chu Y, Fan J and He R: FAP promotes immunosuppression by
cancer-associated fibroblasts in the tumor microenvironment via
STAT3-CCL2 signaling. Cancer Res. 76:4124–4135. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Brummer G, Acevedo DS, Hu Q, Portsche M,
Fang WB, Yao M, Zinda B, Myers M, Alvarez N, Fields P, et al:
Chemokine signaling facilitates early-stage breast cancer survival
and invasion through fibroblast-dependent mechanisms. Mol Cancer
Res. 16:296–308. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Majo S and Auguste P: The Yin and Yang of
discoidin domain receptors (DDRs): Implications in tumor growth and
metastasis development. Cancers (Basel). 13:17252021. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Scordamaglia D, Talia M, Cirillo F,
Zicarelli A, Mondino AA, De Rosis S, Di Dio M, Silvestri F, Meliti
C, Miglietta AM, et al: Interleukin-1β mediates a tumor-supporting
environment prompted by IGF1 in triple-negative breast cancer
(TNBC). J Transl Med. 23:6602025. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Vakilian A, Khorramdelazad H, Heidari P,
Sheikh Rezaei Z and Hassanshahi G: CCL2/CCR2 signaling pathway in
glioblastoma multiforme. Neurochem Int. 103:1–7. 2017. View Article : Google Scholar
|
|
124
|
Zijlmans HJ, Fleuren GJ, Baelde HJ, Eilers
PH, Kenter GG and Gorter A: The absence of CCL2 expression in
cervical carcinoma is associated with increased survival and loss
of heterozygosity at 17q11.2. J Pathol. 208:507–517. 2006.
View Article : Google Scholar
|
|
125
|
Yoshidome H, Kohno H, Shida T, Kimura F,
Shimizu H, Ohtsuka M, Nakatani Y and Miyazaki M: Significance of
monocyte chemoattractant protein-1 in angiogenesis and survival in
colorectal liver metastases. Int J Oncol. 34:923–930. 2009.
|
|
126
|
Zhang J, Patel L and Pienta KJ: CC
chemokine ligand 2 (CCL2) promotes prostate cancer tumorigenesis
and metastasis. Cytokine Growth Factor Rev. 21:41–48. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Soria G, Ofri-Shahak M, Haas I,
Yaal-Hahoshen N, Leider-Trejo L, Leibovich-Rivkin T, Weitzenfeld P,
Meshel T, Shabtai E, Gutman M and Ben-Baruch A: Inflammatory
mediators in breast cancer: coordinated expression of TNFα &
IL-1β with CCL2 & CCL5 and effects on epithelial-to-mesenchymal
transition. BMC Cancer. 11:1302011. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Jin G, Kawsar HI, Hirsch SA, Zeng C, Jia
X, Feng Z, Ghosh SK, Zheng QY, Zhou A, McIntyre TM and Weinberg A:
An antimicrobial peptide regulates tumor-associated macrophage
trafficking via the chemokine receptor CCR2, a model for
tumorigenesis. PLoS One. 5:e109932010. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Vasco C, Canazza A, Rizzo A, Mossa A,
Corsini E, Silvani A, Fariselli L, Salmaggi A and Ciusani E:
Circulating T regulatory cells migration and phenotype in
glioblastoma patients: An in vitro study. J Neurooncol.
115:353–363. 2013. View Article : Google Scholar
|
|
130
|
del Pozo MA, Cabañas C, Montoya MC, Ager
A, Sánchez-Mateos P and Sánchez-Madrid F: ICAMs redistributed by
chemokines to cellular uropods as a mechanism for recruitment of T
lymphocytes. J Cell Biol. 137:493–508. 1997. View Article : Google Scholar
|
|
131
|
Schmall A, Al-Tamari HM, Herold S,
Kampschulte M, Weigert A, Wietelmann A, Vipotnik N, Grimminger F,
Seeger W, Pullamsetti SS and Savai R: Macrophage and cancer cell
cross-talk via CCR2 and CX3CR1 is a fundamental mechanism driving
lung cancer. Am J Respir Crit Care Med. 191:437–447. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Maolake A, Izumi K, Shigehara K,
Natsagdorj A, Iwamoto H, Kadomoto S, Takezawa Y, Machioka K,
Narimoto K, Namiki M, et al: Tumor-associated macrophages promote
prostate cancer migration through activation of the CCL22-CCR4
axis. Oncotarget. 8:9739–9751. 2017. View Article : Google Scholar
|
|
133
|
Qian BZ and Pollard JW: Macrophage
diversity enhances tumor progression and metastasis. Cell.
141:39–51. 2010. View Article : Google Scholar
|
|
134
|
DeNardo DG, Barreto JB, Andreu P, Vasquez
L, Tawfik D, Kolhatkar N and Coussens LM: CD4(+) T cells regulate
pulmonary metastasis of mammary carcinomas by enhancing protumor
properties of macrophages. Cancer Cell. 16:91–102. 2009. View Article : Google Scholar
|
|
135
|
Reinartz S, Schumann T, Finkernagel F,
Wortmann A, Jansen JM, Meissner W, Krause M, Schwörer AM, Wagner U,
Müller-Brüsselbach S and Müller R: Mixed-polarization phenotype of
ascites-associated macrophages in human ovarian carcinoma:
Correlation of CD163 expression, cytokine levels and early relapse.
Int J Cancer. 134:32–42. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
van Attekum MHA, van Bruggen JAC, Slinger
E, Lebre MC, Reinen E, Kersting S, Eldering E and Kater AP: CD40
signaling instructs chronic lymphocytic leukemia cells to attract
monocytes via the CCR2 axis. Haematologica. 102:2069–2076. 2017.
View Article : Google Scholar
|
|
137
|
Zhu P, Baek SH, Bourk EM, Ohgi KA,
Garcia-Bassets I, Sanjo H, Akira S, Kotol PF, Glass CK, Rosenfeld
MG and Rose DW: Macrophage/cancer cell interactions mediate hormone
resistance by a nuclear receptor derepression pathway. Cell.
124:615–629. 2006. View Article : Google Scholar
|
|
138
|
Loberg RD, Ying C, Craig M, Yan L, Snyder
LA and Pienta KJ: CCL2 as an important mediator of prostate cancer
growth in vivo through the regulation of macrophage infiltration.
Neoplasia. 9:556–562. 2007. View Article : Google Scholar
|
|
139
|
Ye J, Yang Y, Jin J, Ji M, Gao Y, Feng Y,
Wang H, Chen X and Liu Y: Targeted delivery of chlorogenic acid by
mannosylated liposomes to effectively promote the polarization of
TAMs for the treatment of glioblastoma. Bioact Mater. 5:694–708.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Liu F, Li X, Zhang Y, Ge S, Shi Z, Liu Q
and Jiang S: Targeting tumor-associated macrophages to overcome
immune checkpoint inhibitor resistance in hepatocellular carcinoma.
J Exp Clin Cancer Res. 44:2272025. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Nywening TM, Wang-Gillam A, Sanford DE,
Belt BA, Panni RZ, Cusworth BM, Toriola AT, Nieman RK, Worley LA,
Yano M, et al: Targeting tumour-associated macrophages with CCR2
inhibition in combination with FOLFIRINOX in patients with
borderline resectable and locally advanced pancreatic cancer: A
single-centre, open-label, dose-finding, non-randomised, phase 1b
trial. Lancet Oncol. 17:651–662. 2016. View Article : Google Scholar
|
|
142
|
Fujimoto H, Sangai T, Ishii G, Ikehara A,
Nagashima T, Miyazaki M and Ochiai A: Stromal MCP-1 in mammary
tumors induces tumor-associated macrophage infiltration and
contributes to tumor progression. Int J Cancer. 125:1276–1284.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Bianconi V, Sahebkar A, Atkin SL and Pirro
M: The regulation and importance of monocyte chemoattractant
protein-1. Curr Opin Hematol. 25:44–51. 2018. View Article : Google Scholar
|
|
144
|
Ding M, He SJ and Yang J: MCP-1/CCL2
mediated by autocrine loop of PDGF-BB promotes invasion of lung
cancer cell by recruitment of macrophages via CCL2-CCR2 Axis. J
Interferon Cytokine Res. 39:224–232. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Avila MA and Berasain C: Targeting
CCL2/CCR2 in tumor-infiltrating macrophages: A tool emerging out of
the box against hepatocellular carcinoma. Cell Mol Gastroenterol
Hepatol. 7:293–294. 2019. View Article : Google Scholar
|
|
146
|
Nywening TM, Belt BA, Cullinan DR, Panni
RZ, Han BJ, Sanford DE, Jacobs RC, Ye J, Patel AA, Gillanders WE,
et al: Targeting both tumour-associated CXCR2(+) neutrophils and
CCR2(+) macrophages disrupts myeloid recruitment and improves
chemotherapeutic responses in pancreatic ductal adenocarcinoma.
Gut. 67:1112–1123. 2018. View Article : Google Scholar
|
|
147
|
Sanford DE, Belt BA, Panni RZ, Mayer A,
Deshpande AD, Carpenter D, Mitchem JB, Plambeck-Suess SM, Worley
LA, Goetz BD, et al: Inflammatory monocyte mobilization decreases
patient survival in pancreatic cancer: A role for targeting the
CCL2/CCR2 axis. Clin Cancer Res. 19:3404–3415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Yin S, Wang N, Riabov V, Mossel DM,
Larionova I, Schledzewski K, Trofimova O, Sevastyanova T, Zajakina
A, Schmuttermaier C, et al: SI-CLP inhibits the growth of mouse
mammary adenocarcinoma by preventing recruitment of
tumor-associated macrophages. Int J Cancer. 146:1396–1408. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Li W, Zhang X, Wu F, Zhou Y, Bao Z, Li H,
Zheng P and Zhao S: Gastric cancer-derived mesenchymal stromal
cells trigger M2 macrophage polarization that promotes metastasis
and EMT in gastric cancer. Cell Death Dis. 10:9182019. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Crane CA, Ahn BJ, Han SJ and Parsa AT:
Soluble factors secreted by glioblastoma cell lines facilitate
recruitment, survival, and expansion of regulatory T cells:
Implications for immunotherapy. Neuro Oncol. 14:584–595. 2012.
View Article : Google Scholar
|
|
151
|
Marangoni F, Zhakyp A, Corsini M, Geels
SN, Carrizosa E, Thelen M, Mani V, Prüßmann JN, Warner RD, Ozga AJ,
et al: Expansion of tumor-associated Treg cells upon disruption of
a CTLA-4-dependent feedback loop. Cell. 184:3998–4015.e19. 2021.
View Article : Google Scholar
|
|
152
|
Rodrigues PF, Wu S, Trsan T, Panda SK,
Fachi JL, Liu Y, Du S, de Oliveira S, Antonova AU, Khantakova D, et
al: Rorγt-positive dendritic cells are required for the induction
of peripheral regulatory T cells in response to oral antigens.
Cell. 188:2720–2737.e22. 2025. View Article : Google Scholar
|
|
153
|
Filaci G, Fenoglio D, Fravega M, Ansaldo
G, Borgonovo G, Traverso P, Villaggio B, Ferrera A, Kunkl A, Rizzi
M, et al: CD8+ CD28- T regulatory lymphocytes inhibiting T cell
proliferative and cytotoxic functions infiltrate human cancers. J
Immunol,. 179:4323–4334. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Serafini P, Borrello I and Bronte V:
Myeloid suppressor cells in cancer: Recruitment, phenotype,
properties, and mechanisms of immune suppression. Semin Cancer
Biol. 16:53–65. 2006. View Article : Google Scholar
|
|
155
|
Serafini P, De Santo C, Marigo I,
Cingarlini S, Dolcetti L, Gallina G, Zanovello P and Bronte V:
Derangement of immune responses by myeloid suppressor cells. Cancer
Immunol Immunother. 53:64–72. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Zea AH, Rodriguez PC, Atkins MB, Hernandez
C, Signoretti S, Zabaleta J, McDermott D, Quiceno D, Youmans A,
O'Neill A, et al: Arginase-producing myeloid suppressor cells in
renal cell carcinoma patients: A mechanism of tumor evasion. Cancer
Res. 65:3044–3048. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Ugel S, De Sanctis F, Mandruzzato S and
Bronte V: Tumor-induced myeloid deviation: when myeloid-derived
suppressor cells meet tumor-associated macrophages. J Clin Invest.
125:3365–3376. 2015. View Article : Google Scholar
|
|
158
|
Kumar V, Cheng P, Condamine T, Mony S,
Languino LR, McCaffrey JC, Hockstein N, Guarino M, Masters G,
Penman E, et al: CD45 phosphatase inhibits STAT3 transcription
factor activity in myeloid cells and promotes tumor-associated
macrophage differentiation. Immunity. 44:303–315. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Flavell RA, Sanjabi S, Wrzesinski SH and
Licona-Limón P: The polarization of immune cells in the tumour
environment by TGFbeta. Nat Rev Immunol. 10:554–567. 2010.
View Article : Google Scholar
|
|
160
|
Panni RZ, Sanford DE, Belt BA, Mitchem JB,
Worley LA, Goetz BD, Mukherjee P, Wang-Gillam A, Link DC, Denardo
DG, et al: Tumor-induced STAT3 activation in monocytic
myeloid-derived suppressor cells enhances stemness and mesenchymal
properties in human pancreatic cancer. Cancer Immunol Immunother.
63:513–528. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Feigin A, vans EE, Fisher TL, Leonard JE,
Smith ES, Reader A, Mishra V, Manber R, Walters KA, Kowarski L, et
al: Pepinemab antibody blockade of SEMA4D in early Huntington's
disease: A randomized, placebo-controlled, phase 2 trial. Nat Med.
28:2183–2193. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Lin BB, Lei HQ, Xiong HY, Fu X, Shi F,
Yang XW, Yang YF, Liao GL, Feng YP, Jiang DG and Pang J:
MicroRNA-regulated transcriptome analysis identifies four major
subtypes with prognostic and therapeutic implications in prostate
cancer. Comput Struct Biotechnol J. 19:4941–4953. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Korbecki J, Kojder K, Simińska D,
Bohatyrewicz R, Gutowska I, Chlubek D and Baranowska-Bosiacka I: CC
chemokines in a tumor: A review of pro-cancer and anti-cancer
properties of the ligands of receptors CCR1, CCR2, CCR3, and CCR4.
Int J Mol Sci. 21:84122020. View Article : Google Scholar
|
|
164
|
Steinberg SM, Zhang P, Malik BT, Boni A,
Shabaneh TB, Byrne KT, Mullins DW, Brinckerhoff CE, Ernstoff MS,
Bosenberg MW and Turk MJ: BRAF inhibition alleviates immune
suppression in murine autochthonous melanoma. Cancer Immunol Res.
2:1044–1050. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Ho PC, Meeth KM, Tsui YC, Srivastava B,
Bosenberg MW and Kaech SM: Immune-based antitumor effects of BRAF
inhibitors rely on signaling by CD40L and IFNγ. Cancer Res.
74:3205–3217. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Cooper ZA, Reuben A, Spencer CN, Prieto
PA, Austin-Breneman JL, Jiang H, Haymaker C, Gopalakrishnan V,
Tetzlaff MT, Frederick DT, et al: Distinct clinical patterns and
immune infiltrates are observed at time of progression on targeted
therapy versus immune checkpoint blockade for melanoma.
Oncoimmunology. 5:e11360442016. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Hugo W, Shi H, Sun L, Piva M, Song C, Kong
X, Moriceau G, Hong A, Dahlman KB, Johnson DB, et al: Non-genomic
and immune evolution of melanoma acquiring MAPKi resistance. Cell.
162:1271–1285. 2015. View Article : Google Scholar
|
|
168
|
Steinberg SM, Shabaneh TB, Zhang P,
Martyanov V, Li Z, Malik BT, Wood TA, Boni A, Molodtsov A, Angeles
CV, et al: Myeloid cells that impair immunotherapy are restored in
melanomas with acquired resistance to BRAF inhibitors. Cancer Res.
77:1599–1610. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Pavon LF, Sibov TT, de Souza AV, da Cruz
EF, Malheiros SMF, Cabral FR, de Souza JG, Boufleur P, de Oliveira
DM, de Toledo SRC, et al: Tropism of mesenchymal stem cell toward
CD133(+) stem cell of glioblastoma in vitro and promote tumor
proliferation in vivo. Stem Cell Res Ther. 9:3102018. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Tu MM, Abdel-Hafiz HA, Jones RT, Jean A,
Hoff KJ, Duex JE, Chauca-Diaz A, Costello JC, Dancik GM, Tamburini
BAJ, et al: Inhibition of the CCL2 receptor, CCR2, enhances tumor
response to immune checkpoint therapy. Commun Biol. 3:7202020.
View Article : Google Scholar
|
|
171
|
Vesely MD and Chen L: Normalization cancer
immunotherapy for melanoma. J Invest Dermatol. 140:1134–1142. 2020.
View Article : Google Scholar
|
|
172
|
Sanmamed MF and Chen L: A paradigm shift
in cancer immunotherapy: From enhancement to normalization. Cell.
175:313–326. 2018. View Article : Google Scholar
|
|
173
|
Ashkenazi A, Pai RC, Fong S, Leung S,
Lawrence DA, Marsters SA, Blackie C, Chang L, McMurtrey AE, Hebert
A, et al: Safety and antitumor activity of recombinant soluble Apo2
ligand. J Clin Invest. 104:155–162. 1999. View Article : Google Scholar
|
|
174
|
Gong Y, Fan Z, Luo G, Yang C, Huang Q, Fan
K, Cheng H, Jin K, Ni Q, Yu X and Liu C: The role of necroptosis in
cancer biology and therapy. Mol Cancer. 18:1002019. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Lemke J, von Karstedt S, Zinngrebe J and
Walczak H: Getting TRAIL back on track for cancer therapy. Cell
Death Differ. 21:1350–1364. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Knight DA, Ngiow SF, Li M, Parmenter T,
Mok S, Cass A, Haynes NM, Kinross K, Yagita H, Koya RC, et al: Host
immunity contributes to the anti-melanoma activity of BRAF
inhibitors. J Clin Invest. 123:1371–1381. 2013. View Article : Google Scholar
|
|
177
|
Grierson PM, Wolf C, Suresh R, Wang-Gillam
A, Tan BR, Ratner L, Oppelt P, Aranha O, Frith A, Pedersen KS, et
al: Neoadjuvant BMS-813160, nivolumab, gemcitabine, and
nab-paclitaxel for patients with pancreatic cancer. Clin Cancer
Res. 31:3644–3651. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
La Cruz TE, González-Bobes F, Eastgate MD,
Sfouggatakis C, Zheng B, Kopp N, Xiao Y, Fan Y, Galindo KA,
Pathirana C, et al: Scalable asymmetric synthesis of the all cis
triamino cyclohexane core of BMS-813160. J Org Chem. 87:1996–2011.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Keklikoglou I, Cianciaruso C, Güç E,
Squadrito ML, Spring LM, Tazzyman S, Lambein L, Poissonnier A,
Ferraro GB, Baer C, et al: Chemotherapy elicits pro-metastatic
extracellular vesicles in breast cancer models. Nat Cell Biol.
21:190–202. 2019. View Article : Google Scholar
|
|
180
|
Lima LG, Ham S, Shin H, Chai EPZ, Lek ESH,
Lobb RJ, Müller AF, Mathivanan S, Yeo B, Choi Y, et al: Tumor
microenvironmental cytokines bound to cancer exosomes determine
uptake by cytokine receptor-expressing cells and biodistribution.
Nat Commun. 12:35432021. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Tarale P, Chaudhary S, Mukherjee S and
Sarkar DK: Ethanol-activated microglial exosomes induce MCP1
signaling mediated death of stress-regulatory proopiomelanocortin
neurons in the developing hypothalamus. J Neuroinflammation.
21:2792024. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Luan S, Zhao Y, Yu Y, Xu J, Xu J, Ren T,
Tang X and Xie L: The relevance of B7-H3 and tumor-associated
macrophages in the tumor immune microenvironment of solid tumors:
Recent advances. Am J Transl Res. 17:2835–2849. 2025. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Zheng Y, Zacarías NV, de Vries H, Han GW,
Gustavsson M, Dabros M, Zhao C, Cherney RJ, Carter P, Stamos D, et
al: Structure of CC chemokine receptor 2 with orthosteric and
allosteric antagonists. Nature. 540:458–461. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
José R, Williams A, Sulikowski M, Brealey
D, Brown J and Chambers R: Regulation of neutrophilic inflammation
in lung injury induced by community-acquired pneumonia. Lancet. 385
(Suppl 1):S522015.
|