Introduction
MicroRNAs (miRNAs) are a class of endogenous, small
non-protein coding single-stranded RNA molecules, which are key
post-transcriptional regulators of gene expression in metazoans and
plants. In animals, miRNAs have regulatory effects through binding
loosely complimentary sequences within the 3′-untranslated regions
(3′UTRs) of their mRNA targets (1,2). In
silico prediction models suggest that miRNAs may be responsible
for the regulation of more than one-third of all human genes.
Functional studies indicate that miRNAs are important to several
fundamental biological processes, including proliferation,
apoptosis, development, and cellular differentiation (3,4).
By negatively regulating their mRNA targets to
either degradation or translational repression, miRNAs have the
capacity to function as either oncogenes or tumor suppressors
(2). Recent findings have suggested
that miRNAs not only are important biomarkers, but also might be
promising therapeutic targets for various diseases (5–8).
Emerging evidence has demonstrated that aberrant expression levels
of miRNAs are involved in glioblastoma multiforme (GBM) initiation
and progression (9). Notably,
miR-21 is almost invariably overexpressed in GBM, resulting in
enhanced cell motility, migration and decreased apoptosis (10,11).
Alternately, important down-regulated miRNAs have also been
identified in glioblastoma, such as miR-128 and miR-7 (12,13).
It has been demonstrated that miR-128 targets Bmi-1 and reduces
cellular proliferation and self-renewal of glioma stem cells
(13).
In this study, the expression of miRNA-205 in glioma
cell lines and the tissues specimens from glioma patients with
certain grades was studied by real-time PCR analysis. Further
investigation revealed that in glioma cell lines, miRNA-205
functioned as a tumor suppressor and overexpression of miRNA-205
reduced cell proliferation, induced G0/G1 phase arrest, decreased
cell invasive capacity and increased apoptosis. We further
demonstrated that miRNA-205 could specifically suppress expression
of VEGF-A by directly interacting with the putative miRNA-205
binding site at the 3′-UTR. Our findings will help to elucidate the
functions of miRNAs and their roles in tumorigenesis.
Materials and methods
Cell lines and tumor specimens
Human glioma cell lines, H4, U87, LN229 and U251,
were purchased from Chinese Academy of Sciences Cell Bank. All
glioma cell lines were maintained in a 37°C, 5% CO2
incubator in DMEM medium supplemented with 10% fetal bovine serum
(Invitrogen, CA, USA) and 1% penicillin-streptomycin (Invitrogen).
Cells were routinely passaged at 2–3 day intervals. Tissue samples
from human glioma and normal brain tissues were obtained from
Shandong Cancer Hospital and Institute (Jinan, China). The
histopathologic diagnoses were determined using WHO criteria and
evaluated by the hospital's pathologist using both morphologic
criteria and immunocytochemistry. Written consent of tissue
donation for research purposes was obtained from the patients
before tissue collection. Twenty-five samples were used for this
research with 5 samples for each group, including primary grade
pilocytic astrocytomas (WHO I), grade II astrocytoma (WHO II),
grade III anaplastic astrocytomas (WHO III), grade IV Glioblastoma
Multiforme (WHO IV) and normal brain tissues derived from the
temporal lobes and saddle area of the patients with arachnoid cyst
(AC) after surgery.
RNA isolation and real-time quantitative
RT-PCR
Total RNA from the frozen tissue specimens and
cultured cells was isolated using the TRIzol kit (Invitrogen)
following to the manufacturer's instructions. RNA quantity was
determined by UV measurement of OD 260/280 nm using the NanoDrop
2000 instrument (Thermo Scientific, FL, USA). To quantitate the
expression level of mature miRNA-205, the isolated RNA was reverse
transcribed and amplified using the mirVana™ qRT-PCR miRNA
detection kit (Ambion) according to the manufacturer's protocol.
PCR reactions were performed using an MJ-real-time PCR (Bio-Rad,
Hercules, CA, USA) system with the following conditions: 95°C, 10
min for 1 cycle, then 95°C, 15 sec, 60°C, 1 min for 40 cycles.
Signals were detected at the end of each cycle. The U6 small
nuclear RNA was amplified as a loading control. The primers for
this U6 internal control were purchased from Ambio. Relative
quantification was conducted using amplification efficiencies
derived from cDNA standard curves and obtained relative gene
expression. Data were shown as fold change (2−ΔΔCt) and
analyzed initially using Opticon Monitor Analysis Software V2.02
software (MJ Research, Waltham, MA, USA). Real-time PCR for VEGF-A
was performed using the MJ-real-time PCR System (Bio-Rad) with the
QuantiTect SYBR Green PCR mixture (Invitrogen). β-actin was used as
control. Amplification conditions were: 95°C, 3 min, 95°C, 30 sec,
60°C, 30 sec, 72°C, 40 sec, for 40 cycles, and 72°C, 8 min for
extension.
Transfection
miRNA-205 mimics and inhibitor and non-targeting
control were obtained from Dharmacon. Cells were transfected using
Lipofectamine 2000 reagent (Invitrogen) at the time of 70%
confluent. Transfection complexes were prepared according to the
manufacturer's instructions and added directly to the glioma cells
to a final oligonucleotide concentration of 50 nmol/l. Transfection
medium was replaced 8-h post-transfection.
MTT proliferation assay
The capacity for cellular proliferation was measured
with a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(MTT) assay. U87 and LN229 cells were plated at 104
cells per well in 96-well plates with six replicate wells for each
condition, transfected with oligonucleotides, and assayed 48-h
post-transfection. The cells were then incubated with 20 μl of MTT
(5 mg/ml) for 4 h at 37°C and 200 μl of DMSO was added to
solubilize the crystals for 20 min at room temperature. The optical
density was determined with a spectrophotometer [Multiskan MK3
(Thermo)] at a wavelength of 570 nm. Cell growth inhibition rates
formula is (AC-AT)/ACx100% (AC, Absorbance value of the blank
control group; AT, Absorbance value of the experimental group).
Anchorage-independent growth assay
After 24 h of transfection, U87 and LN229 cells
(5×102) were suspended in 2 ml of 0.3% agarose with DMEM
medium containing 12% FBS and plated into six-well plates on top of
an existing layer of 0.6% agarose prepared with the same medium.
The plates were incubated at 37°C in a 5% CO2 incubator.
After four weeks, cell colonies were fixed with methanol and
stained with 0.1% crystal violet for 10 min. Then, the colonies
were captured with Olympus SZX12 and Qcapture Pro software
(Olympus). Cell colonies >0.1 mm in diameter were counted under
a microscope. Each assay was performed in triplicate on four
independent occasions.
Cell cycle analysis
For cell cycle analysis, 48 h after transfection,
the adhered cells were obtained by trypsinization and pooled with
the floating cells and centrifuged at 1000 rpm for 5 min and then
incubated with RNase at 37°C for 30 min. A total of 104
nuclei were examined by a FACS Calibur flow cytometer and DNA
histograms were analyzed by Modifit software (Becton Dickinson,
Franklin Lakes, NJ, USA). Experiments were performed in triplicate.
Results are presented as percentage of cells in each phase.
Apoptosis assays
The Annexin V-FITC Apoptosis Detection kit I (Abcam,
USA) was used to detect and quantify apoptosis by flow cytometry.
In brief, cells were harvested 48 h after transfection and
collected by centrifugation for 5 min at 800 × g. Cells were
resuspended at a density of 1×106 cells/ml in 1X binding
buffer, stained with FITC-labeled Annexin V for 5 min and
immediately analyzed by FACScan Flow Cytometer (Becton Dickinson,
San Jose, CA, USA). The data obtained were analyzed using CellQuest
software.
Transwell invasion assay
Matrigel invasion assay was performed using a
24-well invasion chamber system (BD Biosciences, Bedford, MA) with
polycarbonic membrane (diameter: 6.5 mm, pore size 8 μm). Cells
were plated on the top of matrigel-coated invasion chambers in a
serum-free DMEM. As a chemo-attractant, DMEM containing 20% of FBS
was added to the lower compartment of the chamber. The cells were
incubated for 48 h. Invasion of cells to the underside of the
Matrigel-coated membrane was detected by staining the cells with
Mayer's hematoxylin solution and visualizing the cells under a
microscope. After staining, cells were counted under a microscope
in four random fields (magnification, ×100) and results were
expressed in the form of a bar graph. Assays were done in
triplicate for each experiment, and each experiment was repeated
three times.
Western blot analysis
Cells were washed with pre-chilled
phosphate-buffered saline (PBS) three times. The cells were then
solubilized in 1% Nonidet P-40 lysis buffer (20 mM Tris, pH 8.0,
137 mM NaCl, 1% Nonidet P-40, 10% glycerol, 1 mM CaCl2,
1 mM MgCl2, 1 mM phenylmethylsulfonyl fluoride, 1 mM
sodium fluoride, 1 mM sodium orthovanadate, and a protease
inhibitor mixture). Total protein lysates were separated by
SDS-PAGE. The separate proteins were transferred to PVDF membranes.
The blot was incubated with primary antibody detecting VEGF-A
(Santa Cruz; 1:1000 dilution), followed by incubation with
HRP-conjugated secondary antibody. The specific protein was
detected using a super signal protein detection kit (Pierce). After
washing with stripping buffer, the PVDF membrane was reprobed with
antibody against GAPDH (Santa Cruz, 1:1000 dilution).
Synthesis of luciferase reporter
constructs
Luciferase reporters were generated based on the
firefly luciferase expressing vector pGL3-control (Promega).
pGL3-WT-VEGF-A-3′UTR-Luc reporter was created by ligation of PCR
products of 3′UTR of VEGF-A into the XbaI site of the pGL3
control vector. The primers for PCR amplification are:
VEGF-A-3′UTR-Forward: 5′-ATC TCA GCA TGC CTG GTC AGT TAC CTA CTA
ATA GCG GGC CTG-3′ and VEGF-A-3′UTR-Reverse: 5′-GCC CTG AGT GCT GAG
CGA TCA AGT GTC ATT TGA CGT ATC GCT-3′. pGL3-MUT-VEGF-A-3′UTR-Luc
reporter was generated from pGL3-WT-VEGF-A-3′UTR-Luc reporter by
deleting the binding site for miRNA-205.
Luciferase activity assay
Cells were seeded in 24-well plates at
5×104 cells per well the day before transfection.
Luciferase reporter (500 ng), 50 pmol (miRNA-205 mimics or NC) and
40 ng of pRL-TK were added in each well. Cells were collected 48 h
after transfection and analyzed using the Dual-Luciferase Reporter
Assay System (Promega) and Centro LB 960 (Berthold).
Statistical analysis
SPSS10.0 was used for statistical analysis. One-way
analysis of variance (ANOVA) and χ2 test was used to
analyze the significance between groups. The LSD method of multiple
comparisons with parental and control vector groups was used when
the probability for ANOVA was statistically significant.
Statistical significance was determined at P<0.01.
Results
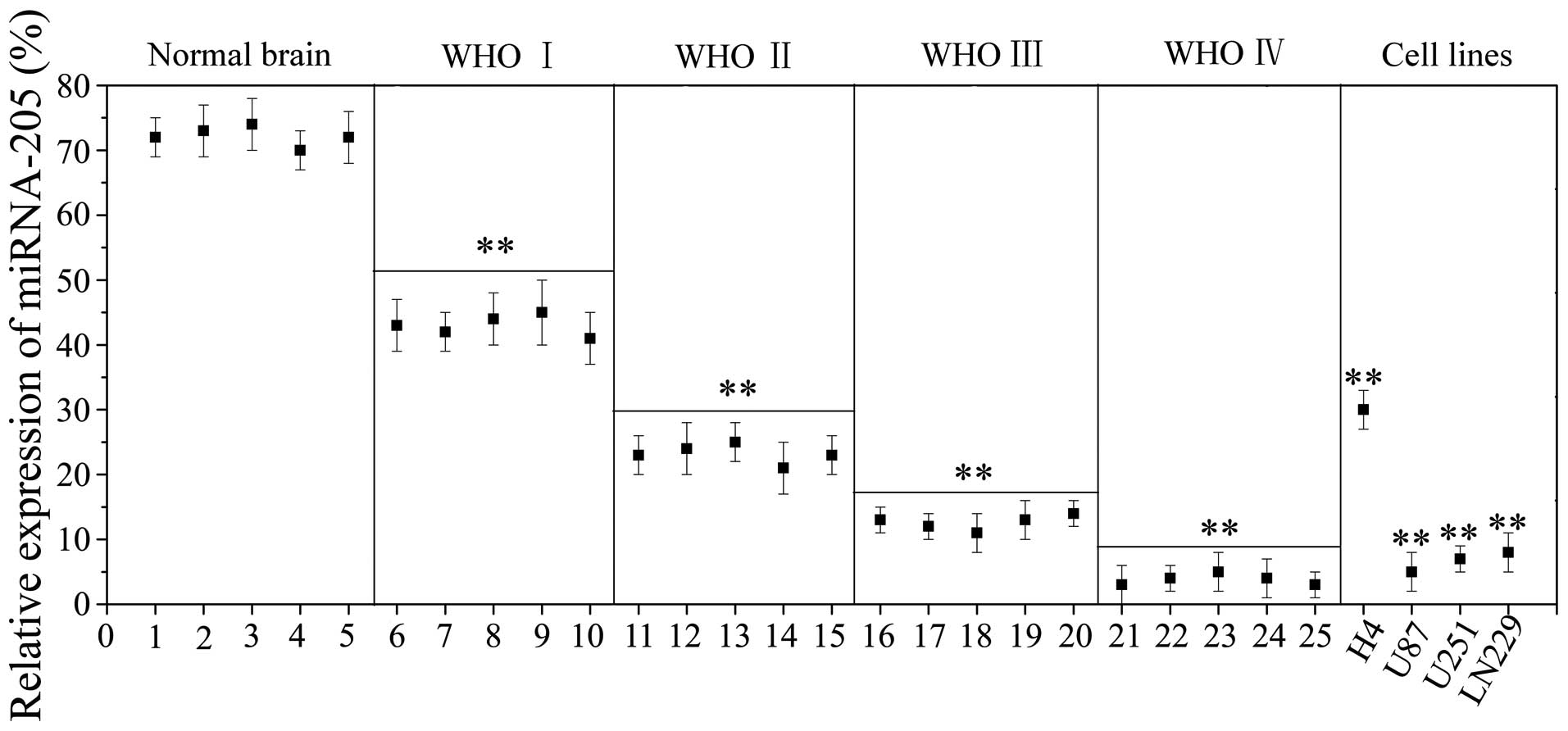
miRNA-205 is down-regulated in glioma
cell lines and tissue specimens
Previously, it has been reported that miRNA-205 is
down-regulated in breast tumor tissues and breast cancer cell lines
(14). However, the expression of
miRNA-205 in tissues of glioma patients has not been well
documented. To assess its relevance in glioma tumorigenesis, we
determined miRNA-205 levels in tumors of different grades compared
to normal brain by quantitative RT-PCR (qRT-PCR). The results
showed that in normal brain tissues, miRNA-205 exhibited a relative
high level expression, whereas the expression of miRNA-205 was
significantly (P<0.01) down-regulated in glioma samples (WHO I,
II, III and IV). The expression of miRNA-205 was negatively
correlated with tumor grade. We also examined expression levels of
miRNA-205 in glioma cell lines (H4, U87, LN229 and U251), and
normal brain tissues as control. They demonstrated the same
expression patterns as miRNA-205 in primary tumors and the normal
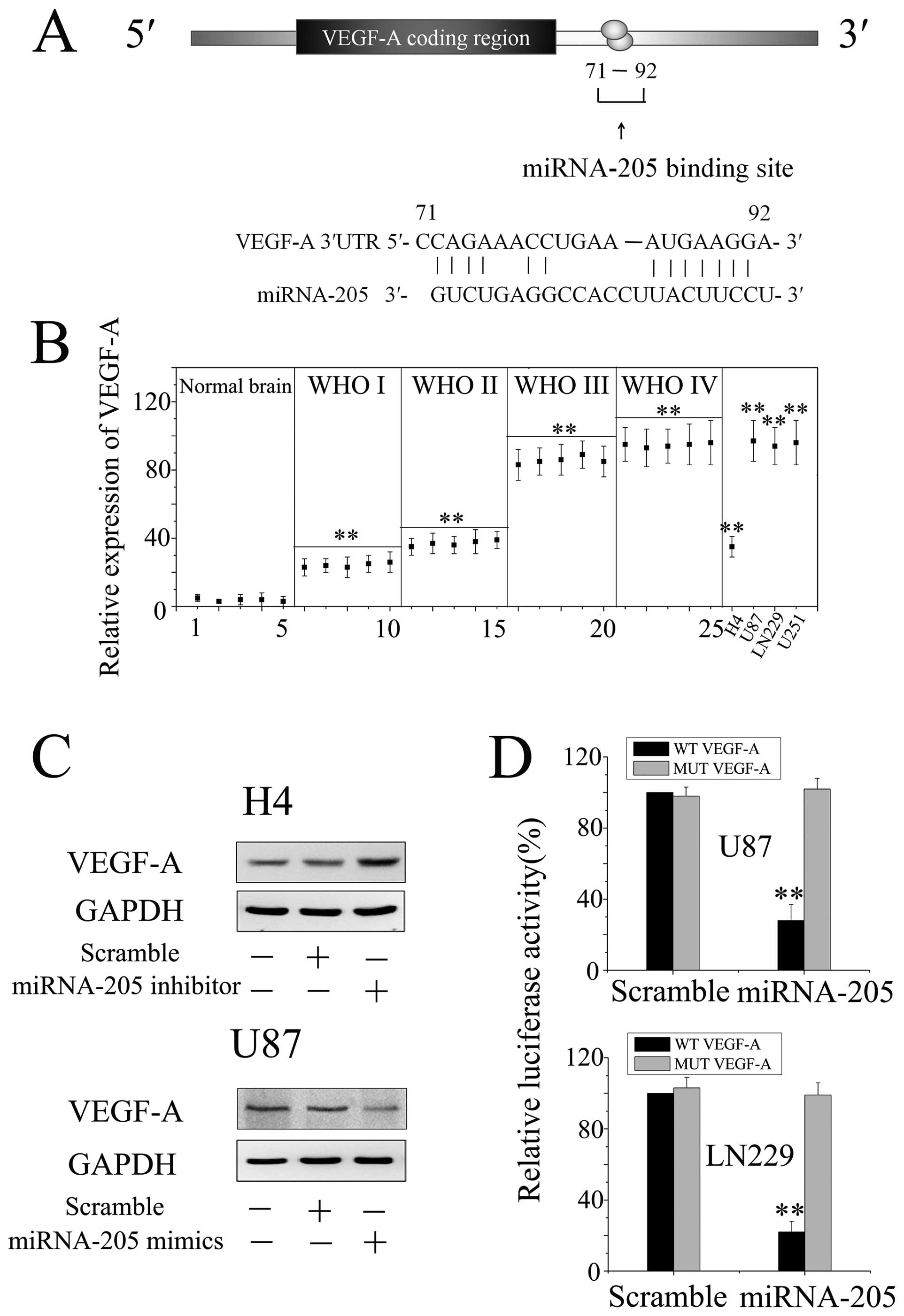
tissues (Fig. 1). The significant
suppression of miRNA-205 expression in tumors and cancer cell lines
suggests a tumor suppressor role in glioma.
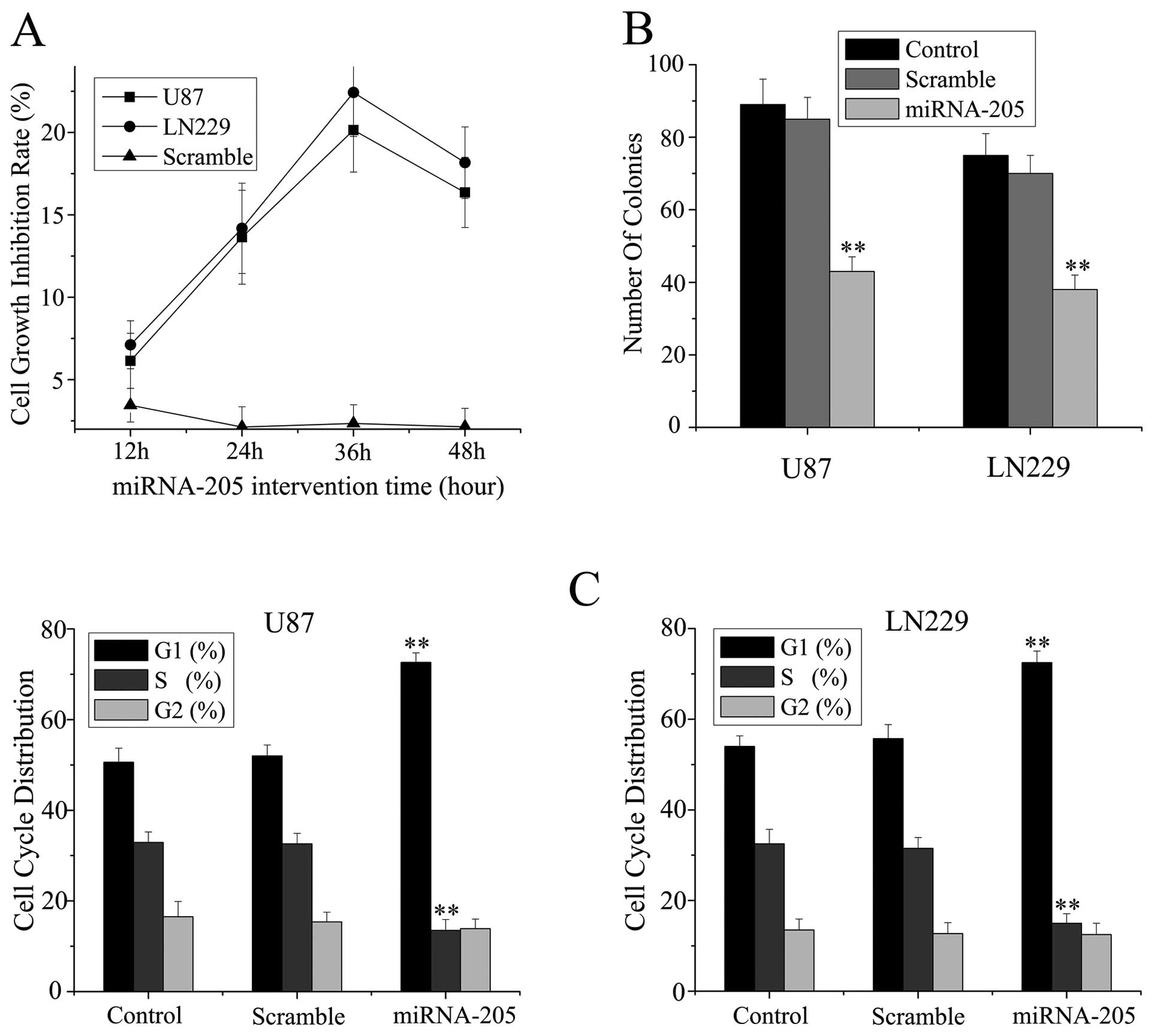
miRNA-205 inhibits the proliferation of
glioma cells in vitro
The significant reduction of miRNA-205 expression in
glioma cell lines and tissue specimens prompted us to explore the
possible biological significance of miRNA-205 in tumorigenesis.
The in vitro growth ability of glioma cells
was determined by MTT assay. About 13.64±2.85% and 14.18±2.47%
inhibition rates of miRNA-205 transfectants in U87 and LN229 cells
at 24 h time point were shown in Fig.
2A and the maximum inhibition rate was at 36-h time point.
These results imply that miRNA-205 might function as a tumor
suppressor in glioma cells in vitro.
To determine whether the inhibition of growth
induced by miRNA-205 in cells was anchorage-independent, the cells
were plated on soft agar 24 h after RNA transfection in U87 and
LN229 cells. After four weeks, the cells transfected with miRNA-205
mimics formed significantly fewer colonies on soft agar than
control and scramble treated cells (Fig. 2B).
To further examine whether the decrease in
proliferation of U87 and LN229 cells reflected a cell cycle arrest,
cell cycle progression was analyzed by propidium iodide staining
and flow cytometric analysis. The results revealed that U87 and
LN229 cells transfected with miRNA-205 mimics had an obvious cell
cycle arrest at the G0/G1 phase (Fig.
2C). These results suggest that miRNA-205 induces cell cycle
arrest and inhibits proliferation of glioma cells.
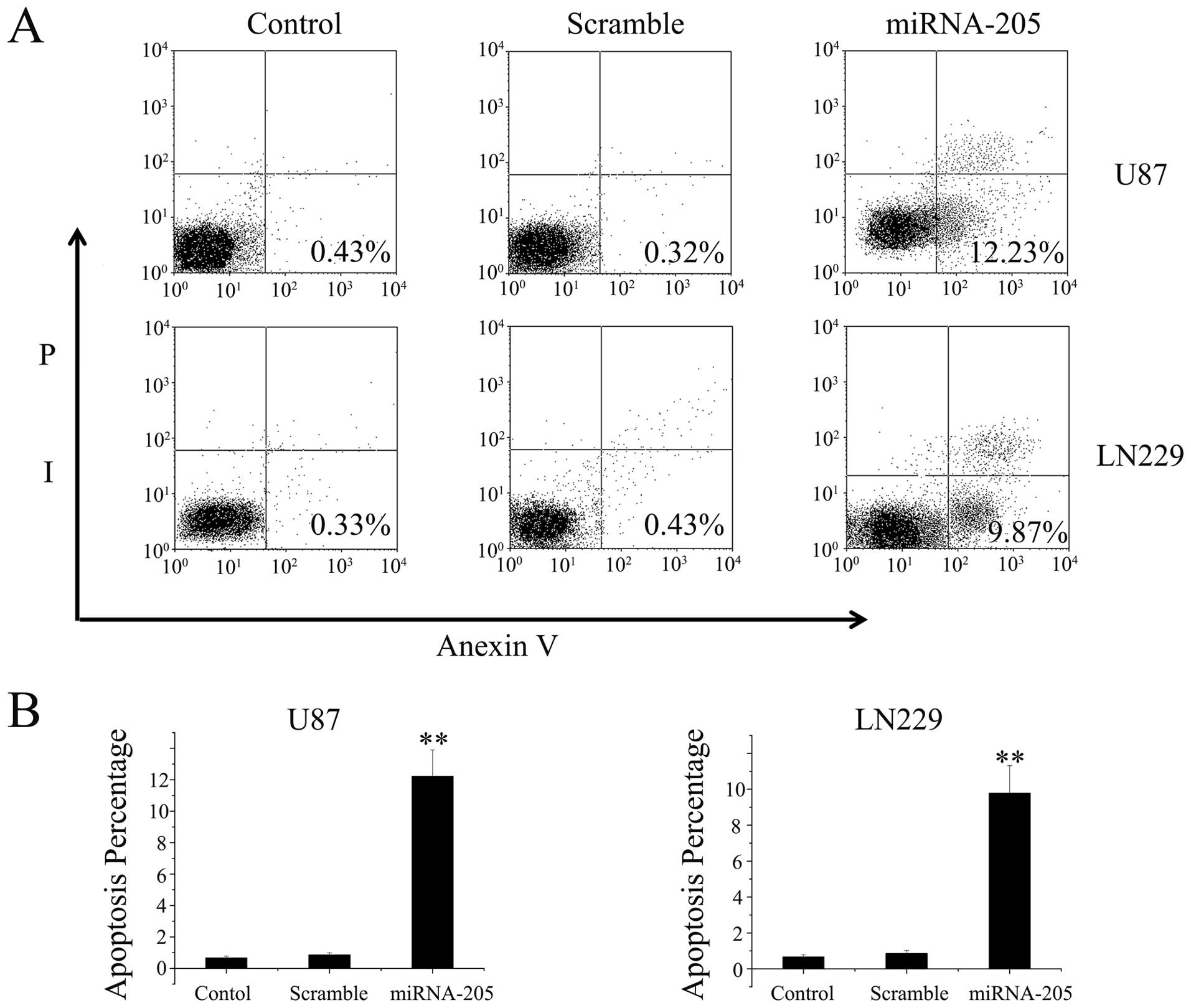
miRNA-205 induces apoptosis in glioma
cell lines
We also analyzed the effect of miRNA-205 on
apoptosis in glioma cells by conducting Annexin V and PI double
staining. The Annexin V-positive early-phase apoptotic cells were
significantly increased in cells transfected with miRNA-205 mimics
oligonucleotide when compared with untreated or scramble controls
cells (Fig. 3A). Percentages of
apoptotic cells are shown in the histogram (Fig. 3B). Annexin-V-FITC/PI double staining
assay showed that miRNA-205 induced apoptosis of U87 and LN229
cells.
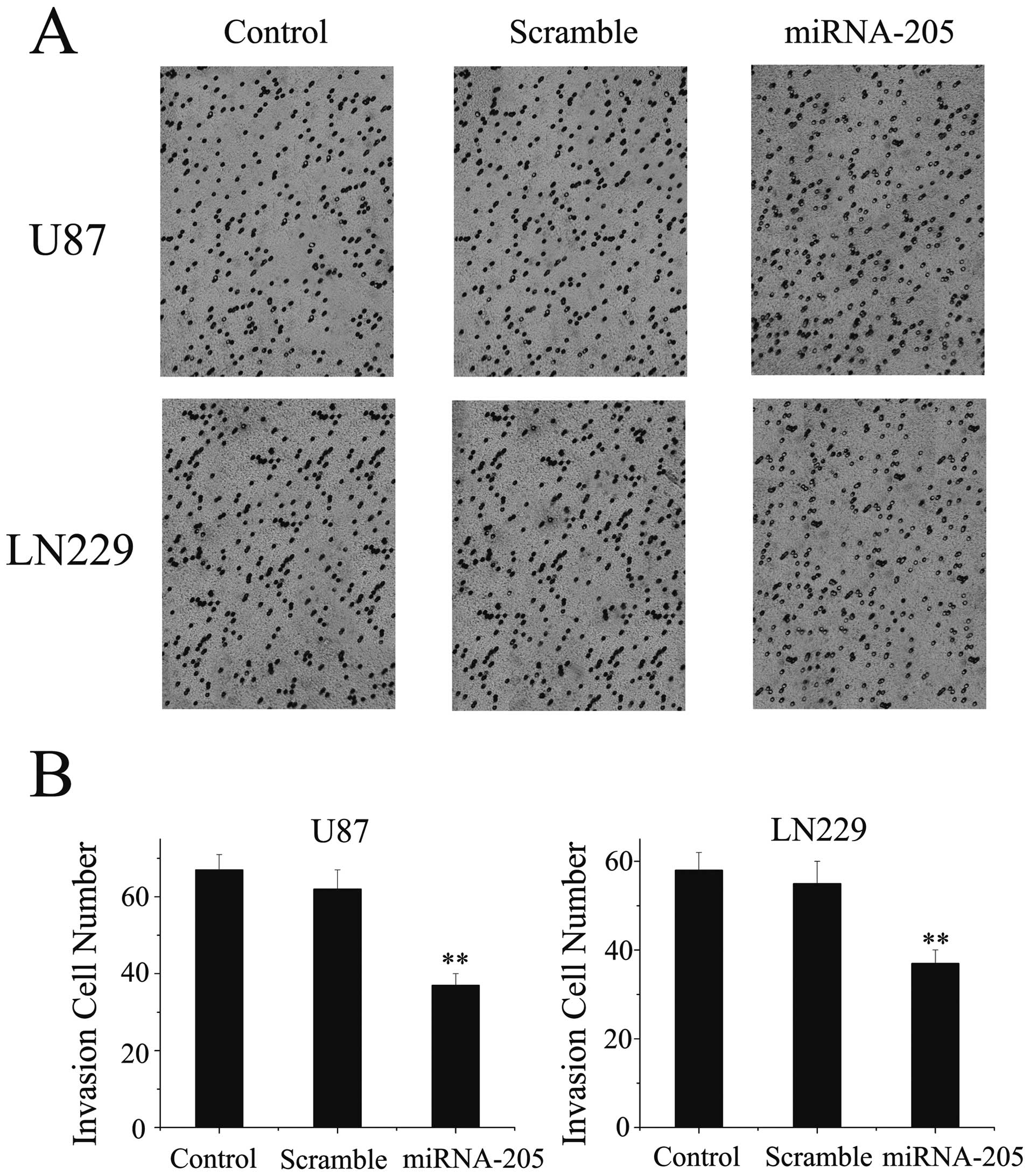
miRNA-205 depresses the invasion of
glioma cells in vitro
Invasive growth is an important biological
characteristic of malignant glioma cells. To evaluate the impact of
miRNA-205 on invasive ability of U87 and LN229 cells, we employed
transwell matrigel invasion assay. In U87 cells, miRNA-205
inhibited invasive activity by ~50%, as 36.24±4.12 cells/field
invaded the matrigel layer compared to 68.78±3.25 and 64.54±3.47
cells/field in the control and scramble-treated groups,
respectively. Similarly, miRNA-205 significantly inhibited invasive
activity of LN229 cells, as 36.26±4.02 cells/field invaded the
matrigel layer compared to 58.32±3.46 and 56.57±3.12 cells/field in
the control and scramble-treated groups, respectively (Fig. 4). These results demonstrate that
miRNA-205 significantly reduces glioblastoma cell invasion
capacity.
VEGF-A is a potential target of miRNA-205
in glioma cells
An obstacle to understanding miRNA function has been
the relative lack of experimentally validated targets (15). To understand the molecular
mechanisms by which miRNA-205 inhibited glioma cells growth and
invasion, we searched for putative miRNA-205 targets as predicted
by the commonly cited programs such as TargetScan, miRanda and
PicTar and found 3′UTR of VEGF-A containing the highly conserved
putative miRNA-205 binding sites (Fig.
5A). We detected the expression of VEGF-A in normal brain,
glioma specimens of different grades, and glioma cell lines.
Herein, we found that the expression of VEGF-A was significantly
elevated with the ascending order of glioma grade (P<0.01),
accompanying the decrease of miRNA-205 (Fig. 5B).
Further, we knocked down expression of miRNA-205 in
H4 cells, which exhibited elevated level of miR-205, and
ectopically expressed miRNA-205 in U87 cells with low endogenous
miRNA-205 expression (Fig. 1).
Western blot analysis showed that VEGF-A expression was
up-regulated in H4 cells with knockdown of miR-205, whereas
down-regulated in U87 cells overexpressing miR-205 (Fig. 5C), compared to control or scramble
treated cells. Moreover, we created pGL3-WT-VEGF-A-3′UTR and
pGL3-MUT-VEGF-A-3′UTR luciferase reporters. Reporter assay revealed
that overexpression of miRNA-205 significantly suppressed the
activity of pGL3-WT-VEGF-A-3′UTR plasmid in U87 and LN229 cells,
without change in luciferase activity of pGL3-MUT-VEGF-A-3′ UTR
plasmid (Fig. 5D). These results
indicate that miRNA-205 directly modulate VEGF-A expression by
binding 3′UTR of VEGF-A in glioma cells.
Discussion
Glioma is the most common type of malignant primary
intracranial tumor. The most frequent and most malignant glioma is
glioblastoma [World Health Organization (WHO) grade IV].
Glioblastoma is characterized by the hallmarks of cellular
heterogeneity, rapid proliferation, angiogenesis, extensive
invasion, hypoxia, necrosis, and infiltration of normal brain
tissue (16). Despite recent
advances in diagnostics and treatments, the prognosis of patients
with glioblastoma has not improved significantly over the past 20
years (17,18). Therefore, there is an urgent need to
gain deeper understanding of molecular mechanisms implicated in
glioblastoma progression and to develop improved conventional or
novel therapeutics.
Knowledge of genetic regulatory mechanisms
initiating and maintaining malignancy are essential for
understanding malignant cellular transformation, pathologic
attributes of cancer, and ultimately, for designing effective
strategies for cancer prevention and treatment (19). Genes encoding miRNAs are numerous
and an even greater number of predicted miRNAs targets have been
identified in the human genome. The breadth of genetic regulatory
effects potentially mediated by miRNAs and their central role in
diverse cellular and developmental processes has led to the
hypothesis that miRNAs might be a novel class of therapeutic
targets or an entirely new class of therapeutic agents for the
treatment of cancers (20–24). In the present report we detected the
miRNA-205 expression level in human glioma samples and found that
the decreased expression level of miRNA-205 was negatively
correlated with the increased malignancy of glioma. While miR-205
is down-regulated in glioma, breast (25) and esophageal cancer (26), it has been shown to be up-regulated
in various types of cancers, including lung cancer, bladder cancer,
ovarian cancer and head and neck cancer cell lines (6,27–30).
These findings may imply that miRNA-205 could play a dual role in
tumorigenicity, depending on tissue type and specific targets. Our
further investigation revealed that overexpression of miRNA-205
reduced cell proliferation, induced G0/G1 phase arrest, decreased
cell invasive capacity and increased apoptosis in glioma cells.
Taken together, the present study for the first time provides
evidence that miRNA-205 is a glioma-specific tumor suppressor.
Angiogenesis plays an essential role in tumor growth
and progression. A large body of research literature incriminates
vascular endothelial growth factor A (VEGF-A) as the most potential
mediator of tumor-induced angiogenesis in glioma (31,32).
Elevated VEGF-A expression is correlated with both increased tumor
microvessel density and increased risk for glioma recurrence and
poor prognosis (33,34). In the present study, we showed that
the expression of VEGF-A was significantly elevated with the
ascending order of glioma grade, accompanying the decrease of
miRNA-205. Our study is consistent with other studies that high
level expression of VEGF-A plays a critical role in glioma
malignancy. It has been reported that knockdown of VEGF-A in cancer
cells inhibited cell malignancy and invasion (33). Clinically, accumulating evidence
indicates that anti-VEGF-A therapeutic approaches have improved
glioma treatment (35). Therefore,
identification of VEGF-A as a direct target for miRNA-205 may imply
that miRNA-205 is a novel target for glioma therapy.
In the present study, the direct interaction between
miRNA-205 and VEGF-A mRNA is supported by several lines of
evidence: 1) the 3′UTR of human VEGF-A mRNAs contain a putative
binding site for miRNA-205 with significant seed match; 2)
miRNA-205 suppresses the activity of a luciferase reporter fused
with the 3′UTR of VEGF-A mRNA, 3) miRNA-205 represses the
endogenous expression of VEGF-A at both the mRNA and protein level.
This finding increases our understanding of VEGF-A regulation in
glioma cells.
In conclusion, we showed there is significant
low-expression of miRNA-205 in glioma cell lines and tissue
specimens. Moreover, we demonstrated that miRNA-205 plays a key
role in the malignancy of glioma cells by directly regulating
VEGF-A expression. This is the first study demonstrating that
miRNA-205 inhibits malignant properties of glioma cells indicating
the therapeutic potential of miRNA-205 in the treatment of
glioma.
Acknowledgements
This work was supported by the National Natural
Scientific Fund of China (No. 30973109), the Shandong Natural
Scientific Fund Project (ZR2009CM093, 2009ZRC03103, ZR2011HQ021),
the Shandong Medical and Health Project (2005HW135) and the
Shandong Scientific and Technological Project (2011GSF11820).
References
|
1
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010.PubMed/NCBI
|
|
2
|
Dykxhoorn DM: MicroRNAs and metastasis:
little RNAs go a long way. Cancer Res. 70:6401–6406. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ivey KN and Srivastava D: MicroRNAs as
regulators of differentiation and cell fate decisions. Cell Stem
Cell. 7:36–41. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kim YK, Heo I and Kim VN: Modifications of
small RNAs and their associated proteins. Cell. 143:703–709. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Volinia S, Calin GA, Liu CG, et al: A
microRNA expression signature of human solid tumors defines cancer
gene targets. Proc Natl Acad Sci USA. 103:2257–2261. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar
|
|
8
|
Lee YS and Dutta A: MicroRNAs in cancer.
Annu Rev Pathol. 4:199–227. 2009. View Article : Google Scholar
|
|
9
|
Ciafre SA, Galardi S, Mangiola A, et al:
Extensive modulation of a set of microRNAs in primary glioblastoma.
Biochem Biophys Res Commun. 334:1351–1358. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chan JA, Krichevsky AM and Kosik KS:
MicroRNA-21 is an antiapoptotic factor in human glioblastoma cells.
Cancer Res. 65:6029–6033. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhou X, Ren Y, Moore L, et al:
Downregulation of miR-21 inhibits EGFR pathway and suppresses the
growth of human glioblastoma cells independent of PTEN status. Lab
Invest. 90:144–155. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kefas B, Godlewski J, Comeau L, et al:
microRNA-7 inhibits the epidermal growth factor receptor and the
Akt pathway and is down-regulated in glioblastoma. Cancer Res.
68:3566–3572. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Godlewski J, Nowicki MO, Bronisz A, et al:
Targeting of the Bmi-1 oncogene/stem cell renewal factor by
microRNA-128 inhibits glioma proliferation and self-renewal. Cancer
Res. 68:9125–9130. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu H, Zhu S and Mo YY: Suppression of cell
growth and invasion by miR-205 in breast cancer. Cell Res.
19:439–448. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Majid S, Saini S, Dar AA, et al:
MicroRNA-205 inhibits Src-mediated oncogenic pathways in renal
cancer. Cancer Res. 71:2611–2621. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Furnari FB, Fenton T, Bachoo RM, et al:
Malignant astrocytic glioma: genetics, biology, and paths to
treatment. Genes Dev. 21:2683–2710. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wen PY and Kesari S: Malignant gliomas in
adults. N Engl J Med. 359:492–507. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Van Meir EG, Hadjipanayis CG, Norden AD,
Shu HK, Wen PY and Olson JJ: Exciting new advances in
neuro-oncology: the avenue to a cure for malignant glioma. CA
Cancer J Clin. 60:166–193. 2010.PubMed/NCBI
|
|
19
|
Gaur A, Jewell DA, Liang Y, et al:
Characterization of microRNA expression levels and their biological
correlates in human cancer cell lines. Cancer Res. 67:2456–2468.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kosik KS: MicroRNAs and cellular
phenotypy. Cell. 143:21–26. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bartel DP: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chiang HR, Schoenfeld LW, Ruby JG, et al:
Mammalian microRNAs: experimental evaluation of novel and
previously annotated genes. Genes Dev. 24:992–1009. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sempere LF, Christensen M, Silahtaroglu A,
et al: Altered MicroRNA expression confined to specific epithelial
cell subpopulations in breast cancer. Cancer Res. 67:11612–11620.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feber A, Xi L, Luketich JD, et al:
MicroRNA expression profiles of esophageal cancer. J Thorac
Cardiovasc Surg. 135:255–260. 2008. View Article : Google Scholar
|
|
27
|
Gottardo F, Liu CG, Ferracin M, et al:
Micro-RNA profiling in kidney and bladder cancers. Urol Oncol.
25:387–392. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tran N, McLean T, Zhang X, et al: MicroRNA
expression profiles in head and neck cancer cell lines. Biochem
Biophys Res Commun. 358:12–17. 2007. View Article : Google Scholar
|
|
29
|
Iorio MV, Visone R, Di Leva G, et al:
MicroRNA signatures in human ovarian cancer. Cancer Res.
67:8699–8707. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yanaihara N, Caplen N, Bowman E, et al:
Unique microRNA molecular profiles in lung cancer diagnosis and
prognosis. Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim KJ, Li B, Winer J, et al: Inhibition
of vascular endothelial growth factor-induced angiogenesis
suppresses tumour growth in vivo. Nature. 362:841–844. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Plate KH, Breier G, Weich HA and Risau W:
Vascular endothelial growth factor is a potential tumour
angiogenesis factor in human gliomas in vivo. Nature. 359:845–848.
1992. View
Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chaudhry IH, O'Donovan DG, Brenchley PE,
Reid H and Roberts IS: Vascular endothelial growth factor
expression correlates with tumour grade and vascularity in gliomas.
Histopathology. 39:409–415. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Varlet P, Guillamo JS, Nataf F, Koziak M,
Beuvon F and Daumas-Duport C: Vascular endothelial growth factor
expression in oligodendrogliomas: a correlative study with
Sainte-Anne malignancy grade, growth fraction and patient survival.
Neuropathol Appl Neurobiol. 26:379–389. 2000. View Article : Google Scholar
|
|
35
|
Jain RK, di Tomaso E, Duda DG, Loeffler
JS, Sorensen AG and Batchelor TT: Angiogenesis in brain tumours.
Nat Rev Neurosci. 8:610–622. 2007. View
Article : Google Scholar
|