Introduction
Pancreatic cancer is a common malignancy, ranking
the thirteenth in the incidence, and the eighth as the cause of
cancer-related deaths worldwide (1). Because of the difficulty in the early
diagnosis of pancreatic cancer, most patients with this malignancy
have already reached an advanced stage when the first symptoms
appear. The standard treatment for advanced pancreatic cancer is
chemotherapy. Although gemcitabine is the first line drug for
pancreatic cancer, the median survival of patients treated with
gemcitabine is not satisfactory. Therefore, researchers expanded
their interests to the development of new treatments for inoperable
pancreatic cancer.
Cell invasion into adjacent tissues is a major
prognostic factor for advanced pancreatic cancer patients. Abnormal
cell migration leads to pathological states such as invasion and
metastasis of cancer. The cytoskeleton, which is composed of actin
filaments and a network of microtubules, controls cell motility
(2). Actin stress fibers generate
contractile forces by pulling against focal adhesions to induce
retraction of the rear cell membrane, suggesting that stress fibers
are important for cell migration (3). Cytoskeletal proteins such as vinculin,
actinin, and several non-receptor protein tyrosine kinases,
including members of the Src family and the focal adhesion kinase
(FAK), are involved in the organization of focal adhesion complexes
(4,5).
The platelet-derived growth factor (PDGF) family is
comprised of four different polypeptide chains encoded by different
genes, which have been identified as PDGF-A, PDGF-B, and the
recently discovered PDGF-C and PDGF-D. So far, four homodimers
(PDGF-AA, PDGF-BB, PDGF-CC and PDGF-DD) and one heterodimer
(PDGF-AB) have been described (6).
PDGF and its receptor are known to have a role in the pathogenesis,
invasion, and distant metastasis of human solid tumors and their
expressions are correlated with poor prognosis (7,8).
Ultraviolet (UV) radiation from sunlight is sorted
by wavelength regions: long-wavelength UVA (320–400 nm),
medium-wavelength UVB (280–320 nm) and short-wavelength UVC
(200–280 nm). UVA and UVB are recognized as the major carcinogenic
components of sunlight (9). On the
other hand, UVC does not actually reach the earth surface since it
is filtered out by the atmosphere, it is generally used for
studying DNA damage and cellular DNA repair process, and is
commonly applied for equipments such as water sterilization.
Recently, the application of UVC for the treatment of human cancer
has been suggested (10,11). Similarly, we recently reported that
UVC irradiation induces cell growth via downregulation of epidermal
growth factor receptor (EGFR) (12)
and also induces apoptosis (13) in
human pancreatic cancer cells. Moreover, our recent study showed
that UVC induced evasion of colon cancer cells from oncogenic
stimulation by EGF (14). Hence, we
believe that UVC could be applied for the clinical strategy against
human malignancies including pancreatic cancer. We herein
investigated the effect of UVC on pancreatic cancer cell migration
and showed that UVC strongly suppressed PDGF-BB-induced migration
in AsPC1 and BxPC3 cells.
Materials and methods
Materials
Recombinant human PDGF-BB was purchased from R&D
Systems, Inc. (Minneapolis, MN). The anti-GAPDH antibody was
purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA).
Antibodies against phospho-p44/p42 mitogen-activated protein kinase
(MAPK), phospho-p38 MAPK, phospho-stress-activated protein
kinase/c-Jun-N-terminal kinase (SAPK/JNK), phospho-Akt and
phospho-GSK-3β were purchased from Cell Signaling, Inc. (Beverly,
MA). The ECL Western blot detection system was purchased from GE
Healthcare (Buckinghamshire, UK). Other materials and chemicals
were obtained from commercial sources.
Cell culture
AsPC1 and BxPC3 pancreatic cancer cells were grown
in Roswell Park Memorial Institute RPMI-1640 (Invitrogen, San
Diego, CA) supplemented with 10% heat-inactivated fetal calf serum
(FCS), penicillin (100 U/ml) and streptomycin (100 μg/ml) (all from
Invitrogen), in a humidified 5% CO2 incubator at 37°C.
Unless indicated otherwise, they were incubated in serum-free
medium for 24 h before experiments as previously described
(13).
Cell migration assay
Cell migration was assessed using a Boyden chamber
(8 μm pores, Transwell®; Corning Costar Corp.,
Cambridge, MA). The cells (AsPC1 and BxPC3) were exposed to UVC
(0–100 J) and incubated for 6 h. These cells (5×104 per
well) were then seeded onto the upper chamber in RPMI containing
10% FCS. After a 16-h incubation at 37°C, the cells were treated
with PDGF-BB at the indicated concentrations for 36 h. The cells
were then fixed and stained with 1 ml of clonogenic reagent (50%
ethanol, 0.25% 1,9-dimethyl-methylene blue) for 30 min. The cells
on the upper surface of the membrane were then mechanically
removed, and the cells that had migrated to the lower surface of
the membrane were observed. The average number of migrated cells
from 5 randomly chosen fields on the lower surface of the membrane
was counted. All data were obtained from at least three independent
experiments.
Western blot analysis
Western blot analyses were performed as previously
described (15). In brief, the
protein lysates (5 μg) were fractionated and transferred onto an
Immune-Blot PVDF Membrane (Bio-Rad Laboratories, Hercules, CA).
Membranes were blocked with 5% fat-free dry milk in
phosphate-buffered saline containing 0.1% Tween-20 for 30 min
before incubation with the indicated primary antibodies.
Peroxidase-labeled antibodies were used as secondary antibodies.
The peroxidase activity on the membrane was visualized on X-ray
film by means of the ECL Western blot detection system.
Densitometric analysis
The densitometric analysis was performed using a
scanner and an image analysis software package (Image J ver. 1.32).
The background-subtracted signal intensity of each protein signal
was normalized to the respective control signal. All data were
obtained from at least three independent experiments.
Statistical analysis
The data were analyzed by ANOVA followed by the
Bonferroni method for multiple comparisons between the indicated
pairs, and p<0.05 was considered to be significant.
Results
PDGF-BB causes migration in AsPC1 and
BxPC3 pancreatic cancer cells
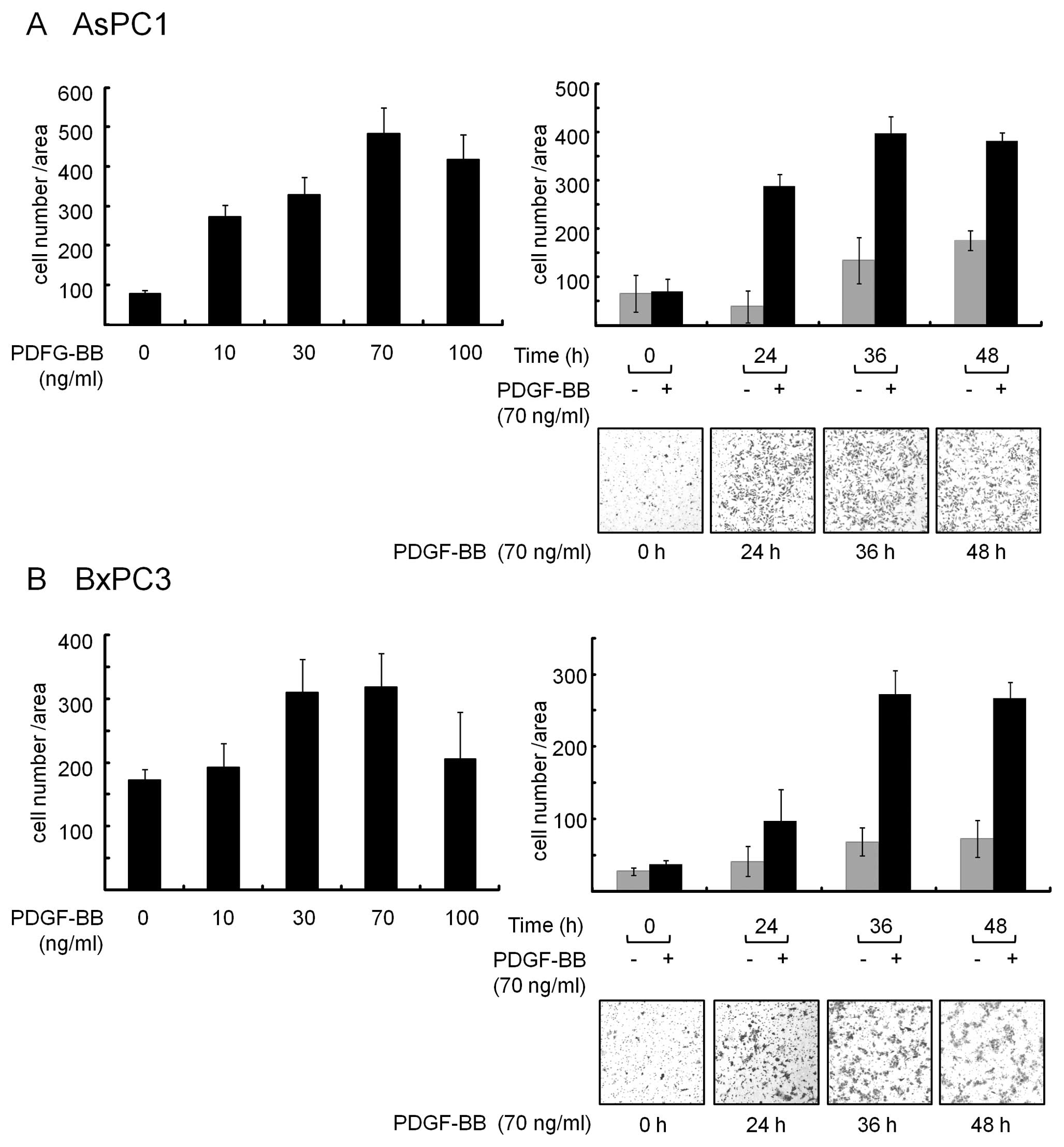
We first examined the effect of PDGF-BB on migration
in AsPC1 and BxPC3 cells. After incubation of the cells
(5×104/well) on the upper Boyden chamber Transwell, they
were treated with PDGF-BB at the indicated concentrations for 24 h.
As depicted in Fig. 1, PDGF-BB
increased the number of migrated cells which were observed on the
lower surface of the membrane. Maximum effects on the migration
were seen when they were exposed to PDGF-BB at a dose of 70 ng/ml
(Fig. 1, left bar graph,
respectively). In addition, time-course experiments showed that 36
h incubation with PDGF-BB exerted sufficient effects on migration
of these cells (Fig. 1, right bar
graph and panels, respectively). Therefore, we used 70 ng/ml of
PDGF-BB and incubated for 36 h after treatment in the following
experiments.
PDGF-BB caused phosphorylation of p44/p42
MAPK, SAPK/JNK, Akt and GSK-3β, but not p38 MAPK in AsPC1 and BxPC3
cells
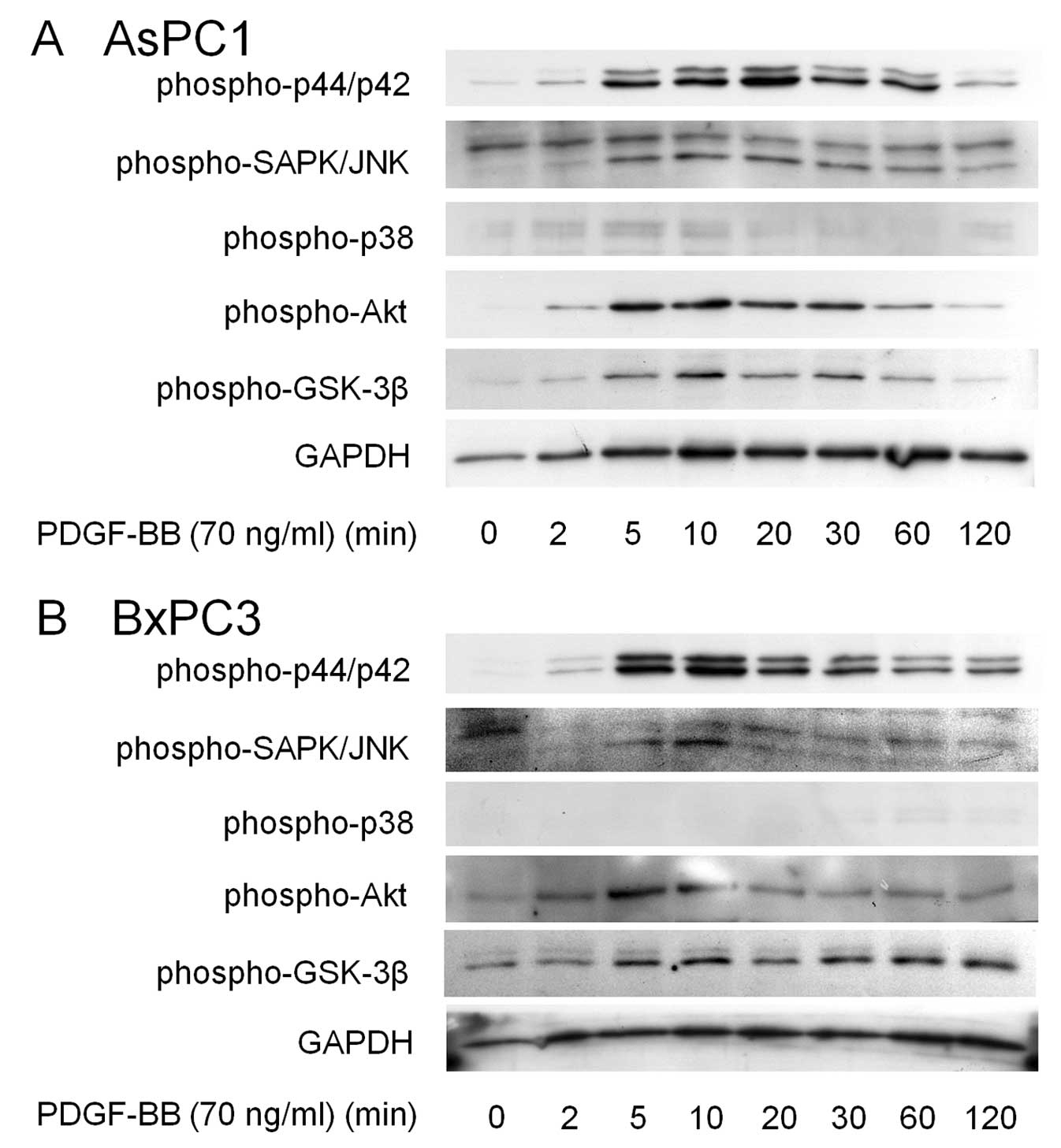
In order to elucidate how PDGF-BB causes pancreatic
cancer cell migration, we next examined the effects of PDGF-BB on
the activation of several kinase cascades. Western blotting
revealed that PDGF-BB induced activation of p44/p42 MAPK, SAPK/JNK
and Akt in AsPC1 cells (Fig. 2A).
GSK-3β is a critical downstream element of the PI3K/Akt cell
survival pathway, and its activity can be inhibited by Akt-mediated
phosphorylation (16) and we
observed that PDGF-BB induced phosphorylation of GSK-3β. However,
PDGF-BB had little effect on p38 MAPK in these cells (Fig. 2A). Since similar effects were also
observed in BxPC3 cells (Fig. 2B),
these results led us to further investigate which kinase plays a
critical role in migration induced by PDGF-BB in pancreatic cancer
cells.
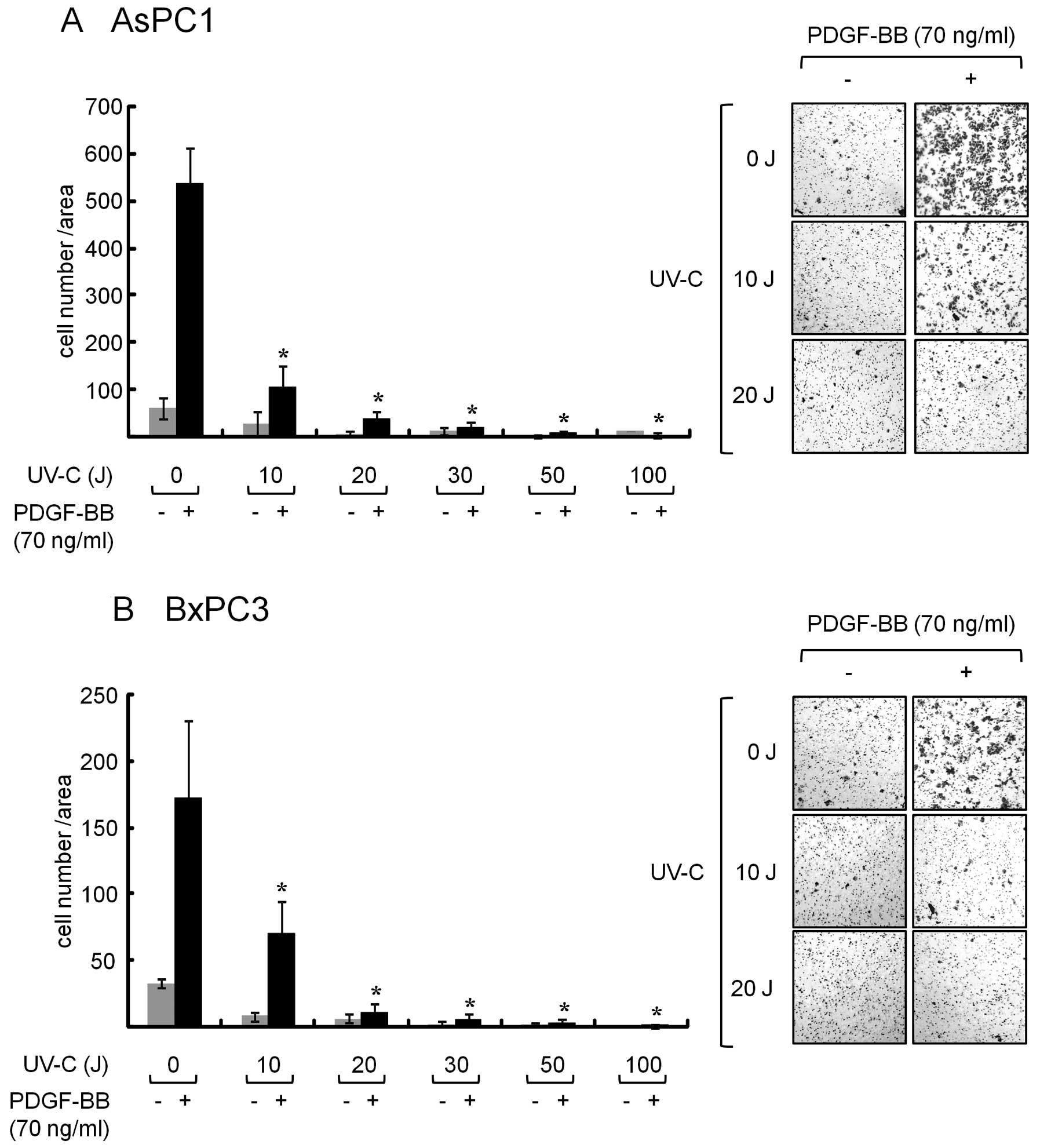
Pretreatement with UVC suppresses
PDGF-BB-induced migration in AsPC1 and BxPC3 cells
We previously reported that UVC induced
downregulation of the EGFR, which leads to cell growth inhibition
in pancreatic cancer cells (12).
Moreover, our recent study showed that UVC can induce apoptosis in
pancreatic cancer cells via the mitochondrial pathway (13). Therefore, we next examined whether
UVC affects migration induced by PDGF-BB in AsPC1 and BxPC3 cells.
As shown in Fig. 3, when the cells
were pretreated with increasing doses of UVC and then exposed to
PDGF-BB, the number of migrated cells was clearly decreased
(Fig. 3). The inhibitory effect of
UVC on migration was seen at a dose over 10 J in these cells. These
results strongly suggest that UVC has a suppressive effect on
pancreatic cancer cell migration.
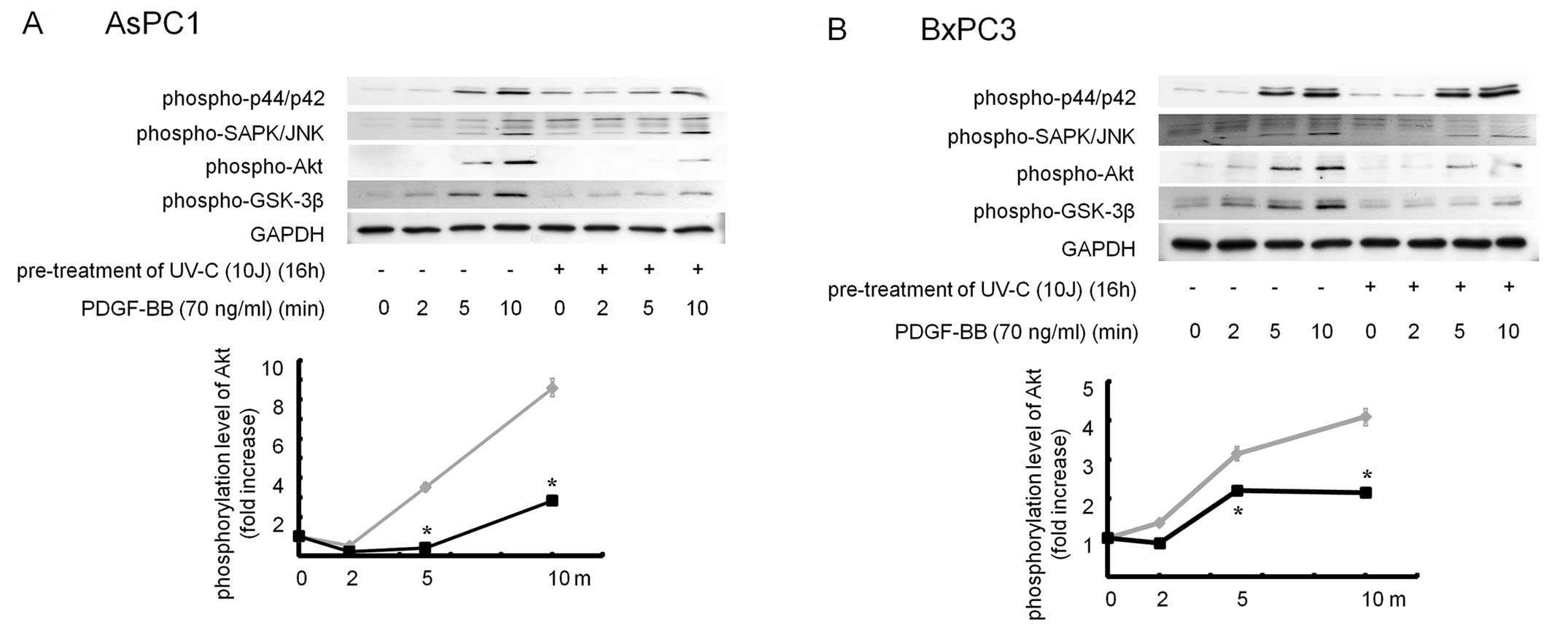
Pretreatment with UVC suppressed PDGF-BB-induced
phosphorylation of Akt and GSK-3β in AsPC1 and BxPC3 cells. In
order to elucidate how UVC suppressed PDGF-BB-induced migration in
pancreatic cancer cells, we next examined the effect of UVC on
several kinase cascade induced by PDGF-BB. While PDGF-BB activated
p44/p42 MAPK, SAPK/JNK, Akt and GSK-3β, but not p38 MAPK (Fig. 2), pretreatment with UVC
significantly inhibited PDGF-BB-induced phosphorylation of Akt and
GSK-3β in these cells. Since UVC failed to affect p44/p42 MAPK and
SAPK/JNK by PDGF-BB (Fig. 4), it is
likely that UVC inhibits PDGF-BB-induced migration by suppressing
Akt-GSK-3β pathway in pancreatic cancer cells.
Discussion
We recently demonstrated the potential applicability
of UVC treatment in patients with pancreatic cancer (12,13).
In these studies, we showed that UVC causes cell growth inhibition
as well as apoptosis in human pancreatic cancer cells. In our
present study, we provide novel evidence that UVC inhibits cancer
cell migration by suppressing the Akt-GSK-3β pathway. Using the
migration assay, we first showed that PDGF-BB, which is known to
play a critical role in invasion and metastasis in human cancers
(7,8), caused migration in AsPC1 and BxPC3
cells (Fig. 1). Moreover,
pretreatment with UVC significantly inhibited PDGF-BB-induced
migration of these cells (Fig. 3).
Since PDGF-BB induced phosphorylation of p44/p42 MAPK, SAPK/JNK,
Akt and GSK-3β in these cells (Fig.
2), we next examined the involvement of these kinases in
PDGF-BB-induced cell migration. Interestingly, UVC suppressed
PDGF-BB-induced phosphorylation of Akt and subsequent GSK3β, but
not p44/p42 MAPK and SAPK/JNK (Fig.
4). Therefore, our results suggest that UVC inhibits
PDGF-BB-induced migration by suppressing the Akt-GSK-3β pathway in
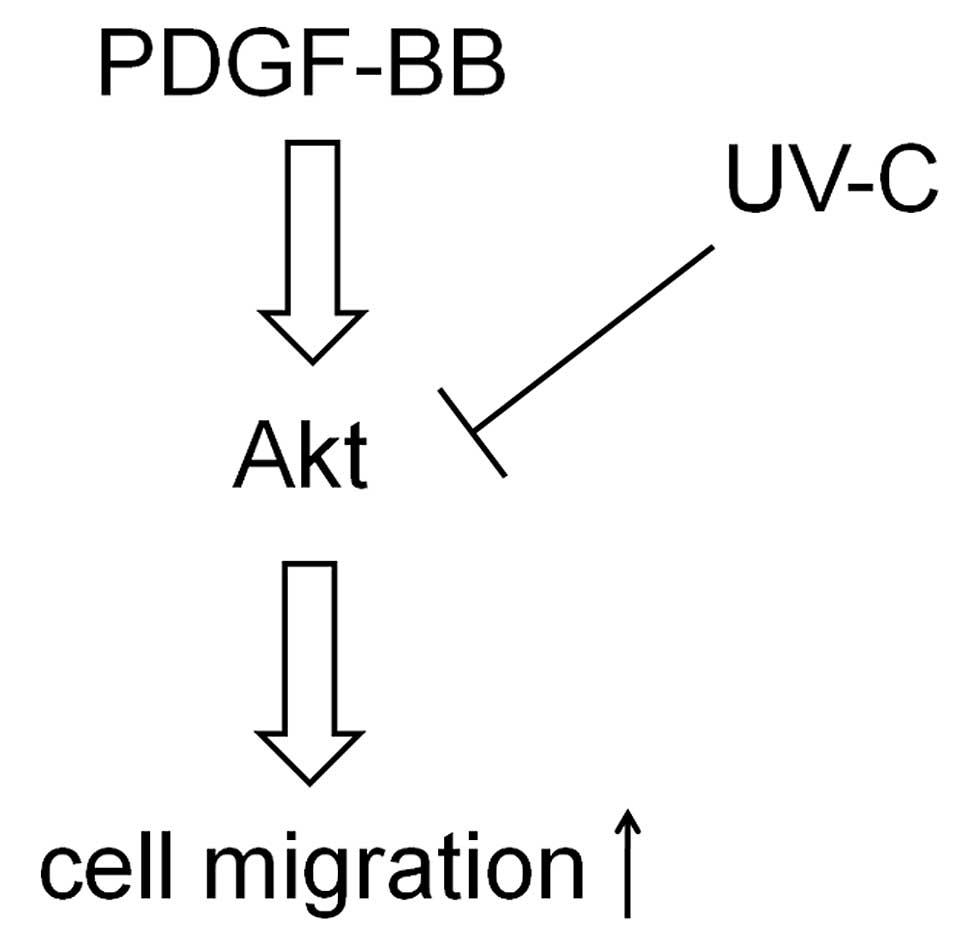
pancreatic cancer cells. A schematic representation of the
hypothetical mechanism by which UVC inhibits cell migration is
shown in Fig. 5.
We previously reported that Rho-kinase, which is a
downstream kinase of Rho, negatively regulates the migration of
colon cancer cells (17). In this
study, Rho-kinase inhibitor induced colon cancer cell migration by
disrupting focal adhesion formation via the Akt pathway. In the
present study, we showed that PDGF-BB-induced migration was
mediated through activation of Akt and that UVC inhibited cell
migration by suppressing the Akt pathway (Fig. 5). Our present findings could allow
us to consider the possibility of UVC as a therapeutic strategy for
pancreatic cancer, although further investigation is required on
the detailed mechanism of UVC suppression of the Akt pathway. In
addition, the development of devices that supply UVC radiation
efficiently is also required for future clinical application. For
example, delivery of UVC radiation to the pancreas through
endoscopic retrograde assisted cholangiopancreatography could be
attained. Moreover, accumulating evidence shows that extracorporeal
photochemotherapy, or photopheresis, which is a low-risk
therapeutical intervention, has been applied to a variety of
hematological malignancies, autoimmune conditions and
transplantation (18). As for
cancer therapy, the mode of action of photopheresis encompasses
apoptosis-induction and modifications of immunoregulatory
processes, leading to the elimination of malignant cells, as well
as the down-modulation of harmful immune responses (18). Therefore, the therapeutic action of
extracorporeal UV irradiation of circulating blood, where
metastatic cells exist, would be worthwhile to evaluate the
possibility of its application to patients with advanced cancer. In
conclusion, our results suggest that UVC irradiation suppresses
PDGF-BB-induced migration via the Akt pathway in human pancreatic
cancer cells.
Acknowledgements
We are very grateful to Ms. Yoko Kawamura for her
skillful technical assistance. This study was supported in part by
a Grant-in-Aid for Scientific Research (no. 22790639 to S.A.) from
the Ministry of Education, Science, Sports and Culture of
Japan.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Riento K and Ridley AJ: Rocks:
multifunctional kinases in cell behaviour. Nat Rev Mol Cell Biol.
4:446–456. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Burridge K: Are stress fibres contractile?
Nature. 294:691–692. 1981. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Humphries JD, Wang P, Streuli C, Geiger B,
Humphries MJ and Ballestrem C: Vinculin controls focal adhesion
formation by direct interactions with talin and actin. J Cell Biol.
179:1043–1057. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Burridge K and Chrzanowska-Wodnicka M:
Focal adhesions, contractility, and signaling. Annu Rev Cell Dev
Biol. 12:463–518. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang Z, Ahmad A, Li Y, et al: Emerging
roles of PDGF-D signaling pathway in tumor development and
progression. Biochim Biophys Acta. 1806:122–130. 2010.PubMed/NCBI
|
|
7
|
Henriksen R, Funa K, Wilander E, Backstrom
T, Ridderheim M and Oberg K: Expression and prognostic significance
of platelet-derived growth factor and its receptors in epithelial
ovarian neoplasms. Cancer Res. 53:4550–4554. 1993.PubMed/NCBI
|
|
8
|
Uren A, Merchant MS, Sun CJ, et al:
Beta-platelet-derived growth factor receptor mediates motility and
growth of Ewing’s sarcoma cells. Oncogene. 22:2334–2342.
2003.PubMed/NCBI
|
|
9
|
Latonen L and Laiho M: Cellular UV damage
responses-functions of tumor suppressor p53. Biochim Biophys Acta.
1755:71–89. 2005.PubMed/NCBI
|
|
10
|
Olsen BB, Neves-Petersen MT, Klitgaard S,
Issinger OG and Petersen SB: UV light blocks EGFR signalling in
human cancer cell lines. Int J Oncol. 30:181–185. 2007.PubMed/NCBI
|
|
11
|
Kim SC, Park SS and Lee YJ: Effect of UV
irradiation on colorectal cancer cells with acquired TRAIL
resistance. J Cell Biochem. 104:1172–1180. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamauchi T, Adachi S, Yasuda I, et al:
UV-C irradiation induces downregulation of EGF receptor via
phosphorylation at serine 1046/1047 in human pancreatic cancer
cells. Radiat Res. 176:565–574. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yamauchi T, Adachi S, Yasuda I, et al:
Ultra-violet irradiation induces apoptosis via mitochondrial
pathway in pancreatic cancer cells. Int J Oncol. 39:1375–1380.
2011.PubMed/NCBI
|
|
14
|
Adachi S, Yasuda I, Nakashima M, et al:
Ultraviolet irradiation can induce evasion of colon cancer cells
from stimulation of epidermal growth factor. J Biol Chem.
286:26178–26187. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Adachi S, Nagao T, Ingolfsson HI, et al:
The inhibitory effect of (−)-epigallocatechin gallate on activation
of the epidermal growth factor receptor is associated with altered
lipid order in HT29 colon cancer cells. Cancer Res. 67:6493–6501.
2007.
|
|
16
|
Cross DA, Alessi DR, Cohen P, Andjelkovich
M and Hemmings BA: Inhibition of glycogen synthase kinase-3 by
insulin mediated by protein kinase B. Nature. 378:785–789. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Adachi S, Yasuda I, Nakashima M, et al:
Rho-kinase inhibitor upregulates migration by altering focal
adhesion formation via the Akt pathway in colon cancer cells. Eur J
Pharmacol. 650:145–150. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Szodoray P, Papp G, Nakken B, Harangi M
and Zeher M: The molecular and clinical rationale of extracorporeal
photochemotherapy in autoimmune diseases, malignancies and
transplantation. Autoimmun Rev. 9:459–464. 2010. View Article : Google Scholar : PubMed/NCBI
|