Introduction
The presentation of tumor-associated antigen (TAA)
by professional antigen presenting cells (APC), especially
dendritic cells (DCs) play an essential role in antitumor effects
in vitro and in vivo (1). DCs are believed to be the most potent
professional antigen-presenting cells (2,3). They
can take up antigen efficiently, and present the antigen on their
surface in association with major histocompatibility complex (MHC)
molecules stimulating naive T cells to proliferate and
differentiate (4–6). Therefore, the investigation of
DCs-based vaccines in cancer therapy has recently received much
attention. Different strategies have been developed to load DCs
with TAA, including synthetic peptides derived from the known
antigens (7), tumor lysates
(8), tumor RNA (9) and dying tumor cells (10) to induce antigen-specific immune
responses. It has been reported that the endogenous processing and
presentation of TAA peptides may be more efficient for cell surface
presentation than the exogenous loading of synthetic TAA peptides
(11).
Telomerase is a unique ribonucleoprotein that
mediates RNA-dependent synthesis of telomeric DNA, the distal ends
of eukaryotic chromosomes that stabilize the chromosomes during
replication (12). Telomerase is
active in more than 85% of human cancers and some stem cells but
repressed in most normal human somatic tissue (13,14).
Human telomerase reverse transcriptase (hTERT) is the rate-limiting
component of telomerase (15). In
cells where telomerase is activated, hTERT synthesizes a TTAGGG
sequence from the RNA template that is then added to the end of the
shortening chromosome (16), thus
saving the cells from death. The above mechanism is exploited by
tumour cells to maintain their immortality (14,17).
The widespread expression of telomerase in cancer, coupled with the
critical role of hTERT in the telomerase complex, suggests that
hTERT maybe used as a universal TAA. Furthermore, there is
increasing evidence that peptides derived from the protein of hTERT
could been specifically recognized by CD8+ and
CD4+ T lymphocytes (18).
hTERTC27 (C27) is an artificially derived 27 kDa
C-terminal polypeptide fragment of human TERT. It has previously
been demonstrated that overexpression of hTERTC27 in HeLa cells
could reduce the tumorigenicity and suppress the growth of
xenografted glioblastoma in nude mice (19). C27 can also upregulate genes that
are involved in apoptosis, the cell cycle, and the immune response
(20). The rAAV-/rAdv-hTERTC27
viral cocktail can also activate NK cells, but not T cells, against
melanoma (21). Since hTERT was
identified as a universal tumor-associated antigen, we hypothesize
that hTERTC27 could suppress tumor growth through the specific CTL
response. In the present study, we explored whether DCs-transfected
with rAd-hTERTC27-EGFP (rAd-C27 DCs) would elicit potent adaptive
immunity against gliomas. Recombinant adenoviral vectors were
selected in this study since others have found the adenovirus to be
a highly efficient and reproducible method of gene transfer into
DCs (22). We found that DCs
transduced with rAd-C27 effectively induce specific cytotoxic T
lymphocytes (CTL) against gliomas cells in vitro and in
vivo.
Materials and methods
Cell culture
The glioblastoma cell line GL261 was a gift from
Professor Wang (Academy of Military Medical Sciences). Cells were
cultured in DMEM (Gibco, Hangzhou, China), with 10% fetal bovine
serum, 100 U/ml penicillin and 100 mg/ml streptomycin (Gibco). The
cells were maintained at 37°C in 5% CO2 cultured and
passaged at weekly intervals.
DC generation from mouse bone marrow
DCs from mouse bone marrow were generated as
previously described (23,24). In brief, bone marrow was flushed
from the tibias and femurs of C57BL/6 mice (Laboratory Animal
Center, Sun Yat-sen University, China) and depleted of erythrocytes
with commercial lysis buffer (Sigma, St. Louis, MO, USA). The cells
were washed twice in serum-free RPMI-1640 medium and cultured in a
6-well plate at 5×106 cells/well with RPMI-1640 medium
containing 10 ng/ml recombinant mouse GM-CSF (R&D Systems,
Inc., USA) and 10 ng/ml recombinant mouse IL-4 (R&D Systems).
On Days 3 and 5, half of the media were refreshed and fresh
cytokine-containing mGM-CSF and mIL-4 media were added. On Day 6,
200 ng/ml LPS was added to the media. On Day 7, non-adherent cells
obtained from these cultures were considered mature bone
marrow-derived DCs.
Flow cytometric analysis
DCs were collected and resuspended in PBS. Cells
were immunostained with fluorescein isothiocyanate
(FITC)-conjugated anti-mouse MHC-II, and phycoerythrin
(PE)-conjugated anti-mouse CD80, CD86 antibodies (eBioscience,
USA). Corresponding FITC immunoglobulin G (IgG) isotype control
antibody (eBioscience) was used. A total of 1×106 cells
were incubated half an hour at 4°C with antibodies. The cells were
then washed twice with PBS resuspended, and analyzed on a FACScan
(Becton-Dickinson, USA).
Recombinant adenovirus-mediated gene
transfer
Transduction of mouse mature DCs with rAd vector was
conducted in 6-well plates with 1×106 DCs per well in a
2-ml volume of RPMI-1640 medium containing 10% FBS. Viruses were
added to the wells at a multiplicity of infection (MOI) of 200 and
the DCs were harvested after 24 h of incubation.
Western blot analysis
For western blot analysis, proteins in the cell
extracts were separated by 10% sodium dodecyl sulphate
polyacrylamide gel electrophoresis (SDS-PAGE) and then transferred
onto a nitrocellulose membrane. The membrane was incubated with 5%
non-fat milk in PBS and then with anti-hTERT antibody (Abcam, Hong
Kong, China) overnight at 4°C. After washing, the membranes were
incubated with an alkaline phosphatase-conjugated goat anti-rabbit
IgG antibody (DingGuo, Beijing, China) for 1 h at room temperature.
Immunoreactive bands were detected using the ECL western blot
analysis system.
In vitro experimentation
Mixed lymphocyte reaction (MLR)
Briefly, for preparation of T lymphocytes, spleens
of C57BL/6 mice were removed aseptically, passed over nylon wool
with their purity determined by FACS and prepared for the following
experiments. The allogeneic T cells were mixed with
1×105 DCs transduced with rAd-C27 (rAd-C27 DCs), DCs
transduced with rAd-EGFP (rAd-EGFP DCs) and normal control DCs in a
well of flat-bottomed 96-well plates in 200 μl of RPMI-1640
containing 10% FCS, and cultured at 37°C for 3 days. DCs were used
as the stimulator (S) and T cells were used as the responder cells
(R) with the S/R ratio varying from 1:5 to 1:40. CCK8 (20 μl)
(Dojindo, Japan) was added into each well at 6 h before termination
of the incubation. Subsequently, the absorbance values (at 450 nm)
were recorded on the culture medium of each sample using a Bio-Rad
microplate reader (Bio-Rad Laboratories, Hercules, CA, USA).
Cytotoxicity assays
T cells were co-cultured with rAd-C27 DCs, rAd-EGFP
DCs and normal control DCs in a 24-well tissue culture plate in a
1-ml complete RPMI-1640 medium at 37°C in 5% CO2 for 72
h for the cytotoxic T lymphocytes (CTL). Then the CTLs were
collected and used as the effector cells in CTL assays against U87
cells. The U87 cells, as the target cells, were placed in 96-well
tissue culture plates at 1×104 cells/well and
co-cultured with the effector cells (CTL) at the (effect/target)
ratio of 5:1, 20:1 and 40:1, at 37°C in 5% CO2. The
cytotoxic activities were determined by CCK8.
ELISA
cells were co-cultured with Ad-C27 DCs, rAd-EGFP DCs
and normal control DCs (S/R ratio, 1:10) in a 24-well tissue
culture plate in 1 ml complete RPMI-1640 medium at 37°C in 5%
CO2 for 3 days. The cell culture supernatant was
harvested. IL-2 and IFN-γ in the supernatant was determined with
ELISA using anti-human IL-2 and IFN-γ purified monoclonal antibody
and biotin-labeled anti-human IL-2 and IFN-γ monoclonal
antibody.
In vivo experimentation
Murine brain tumor model and
intracerebral injection of DCs
Female C57BL/6 mice were anesthetized with an
intraperitoneal injection of 4% chloral hydrate and fixed in a
stereotactic head frame (Huaibei Zhenghua Instruments Co., Anhei,
China). A midline scalp incision was made and the bregma was
identified. Stereotactic coordinates were measured (2.0 mm lateral,
and 1.2 mm anterior to the bregma) for implantation of cells into
the deep frontal white matter. A burr hole was drilled at this
point and 1×105 GL261 cells suspended in 2.5 μl of PBS
were injected through a Hamilton syringe (Shanghai Libao
Instruments Co., Shanghai, China) with a fixed, 25-gauge needle at
a depth of 3.0 mm relative to the dura mater. Injections were
performed at 1 μl/min. The needle was withdrawn and the incision
sutured. On Day 3 and Day 10 after the tumor implantation, the mice
were injected with 1×105 rAd-C27 DCs, rAd-EGFP DCs or
DCs suspended in 2.5 μl of PBS in the same location. The animals
were monitored daily after treatment and the survival time of each
mouse was recorded.
CTL activity assay
Splenocytes were harvested as previously described
and pooled from two mice per group on Day 7 after the second
intracerebra injection of rAd-C27 DCs, rAd-EGFP DCs or DCs. These T
cells (2×107) were restimulated in vitro with
1×106 GL26 cells which were treated with 150 μg/ml
mitomycin C at 37°C for 1 h beforehand. Then the mixed cells were
co-cultured for 5 days in the presence of 20 IU/ml recombinant
human IL-2. GL261 cells (3×104) as target cells were
incubated in a 96-well plate at 37°C for 12 h. The above T cells
used as effector cells were co-cultured with GL26 cells at the
effector/target ratios of 5:1, 20:1 and 40:1, at 37°C in 5%
CO2. The cytotoxic activities were determined by
CCK8.
Histology
On Day 21 after tumor implantation, two mice from
each group were euthanized to obtain brain tissues. These tissues
were stained with hematoxylin and eosin (H&E) in order to
clearly display the tumor outline. The tumor volume
(mm3) was calculated using the formula of π/6xa2xb where
a is width and b is length.
Statistical analysis
Data were analyzed using χ2 analysis. The
in vivo anticancer effect of different treatments was
assessed by plotting survival curves according to the Kaplan-Meier
method, and groups were compared using the log-rank test.
Differences were considered statistically significant when the
P-value was <0.05. All statistical analyses were carried out
with SPSS 13.0 software.
Results
Morphological and phenotypic
characteristics of mouse bone marrow-derived DCs
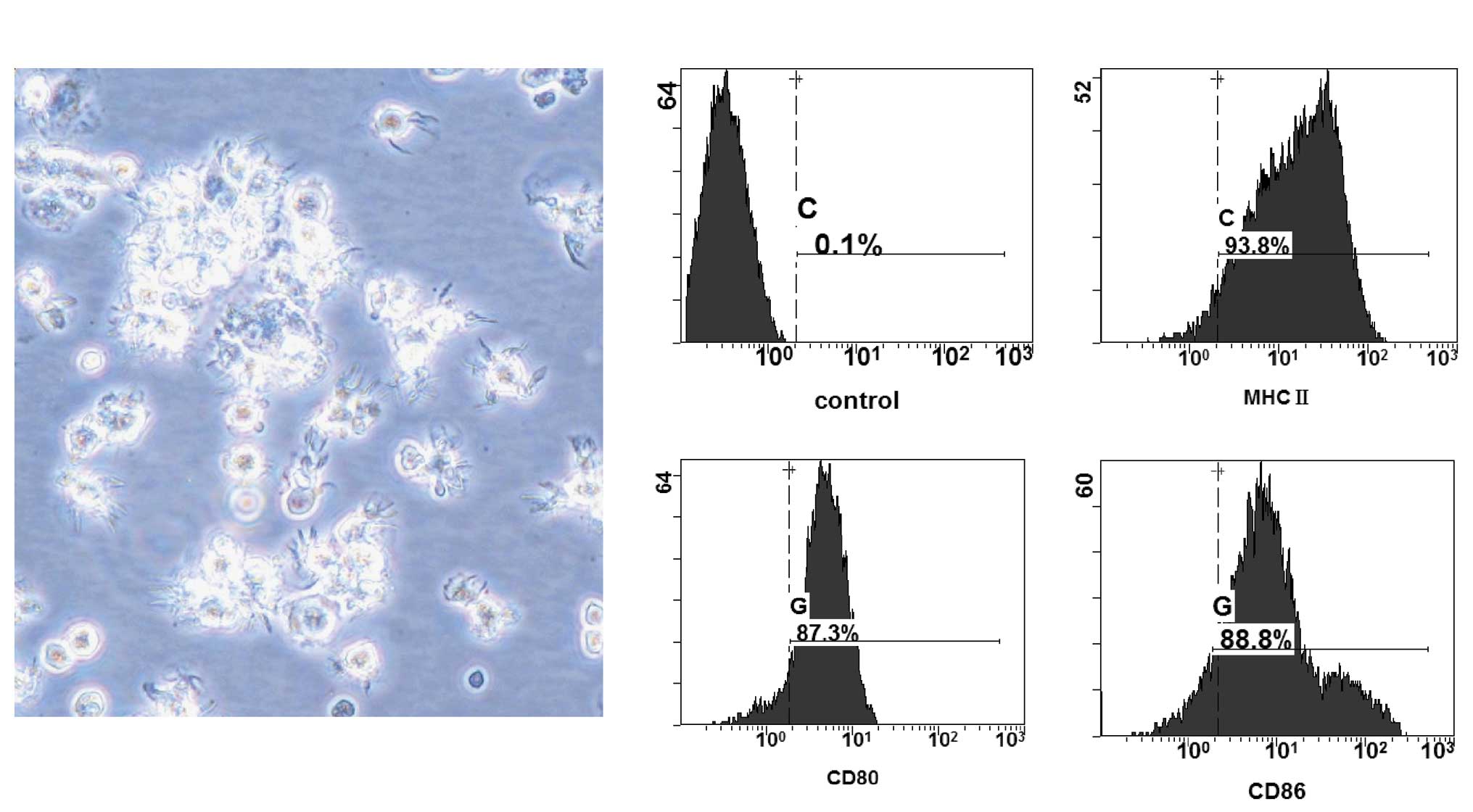
On Day 7 of cell culture, mature DCs displaying
typical morphological characteristics were harvested from monocytes
cultured in medium containing mGM-CSF, mIL-4 and LPS. When viewed
by phase contrast microscopy, these mature cells were suspended
together, exhibited an irregular cell shape, and displayed the
pricking and dendritic eminences on their surfaces (Fig. 1A). The phenotype of the mature DCs
was analyzed using FACS. The results showed that these mature DCs
expressed high levels of CD80 (87.3%), CD86 (88.8%) and MHC-II
(93.8%) (Fig. 1B). The results
demonstrated the successful preparation of DCs from the bone marrow
of mice to be used for subsequent experiments.
Infection of DCs with adenovirus and the
identification of hTERTC27 protein expression
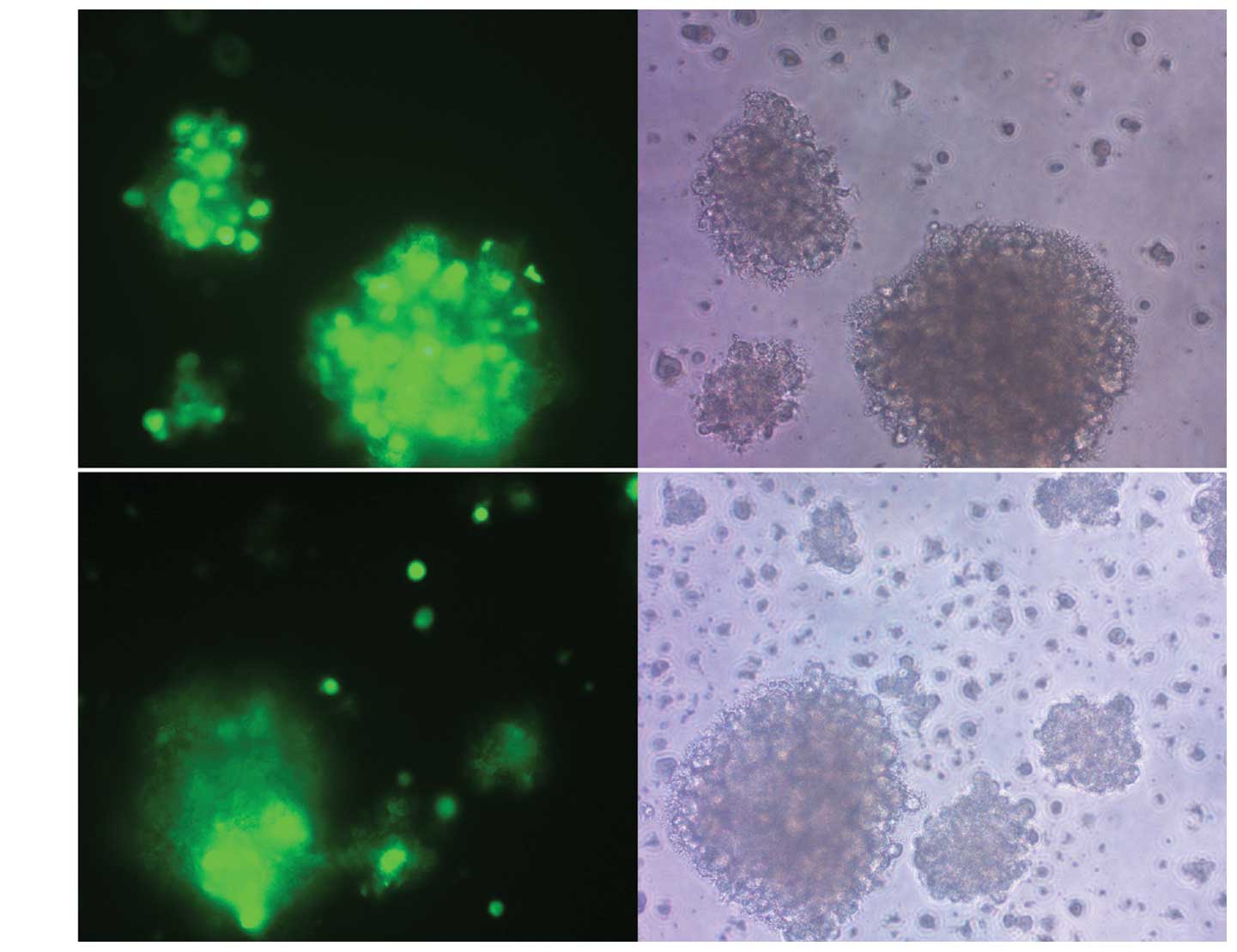
DCs derived from mouse bone marrow were cultured in
the previously-defined medium containing mGM-CSF and mIL-4. rAd-C27
was constructed and used to infect the DCs at an MOI of 200 by the
centrifugal method, which was optimized for suitable transfection
efficiency and toxicity. A control vector rAd-EGFP was used to
infect the DCs in parallel, which resulted in EGFP expression in
about 80% of the DCs (Fig. 2A),
indicating a high transduction efficiency of the adenovirus vector
(Fig. 2B). The expression of the
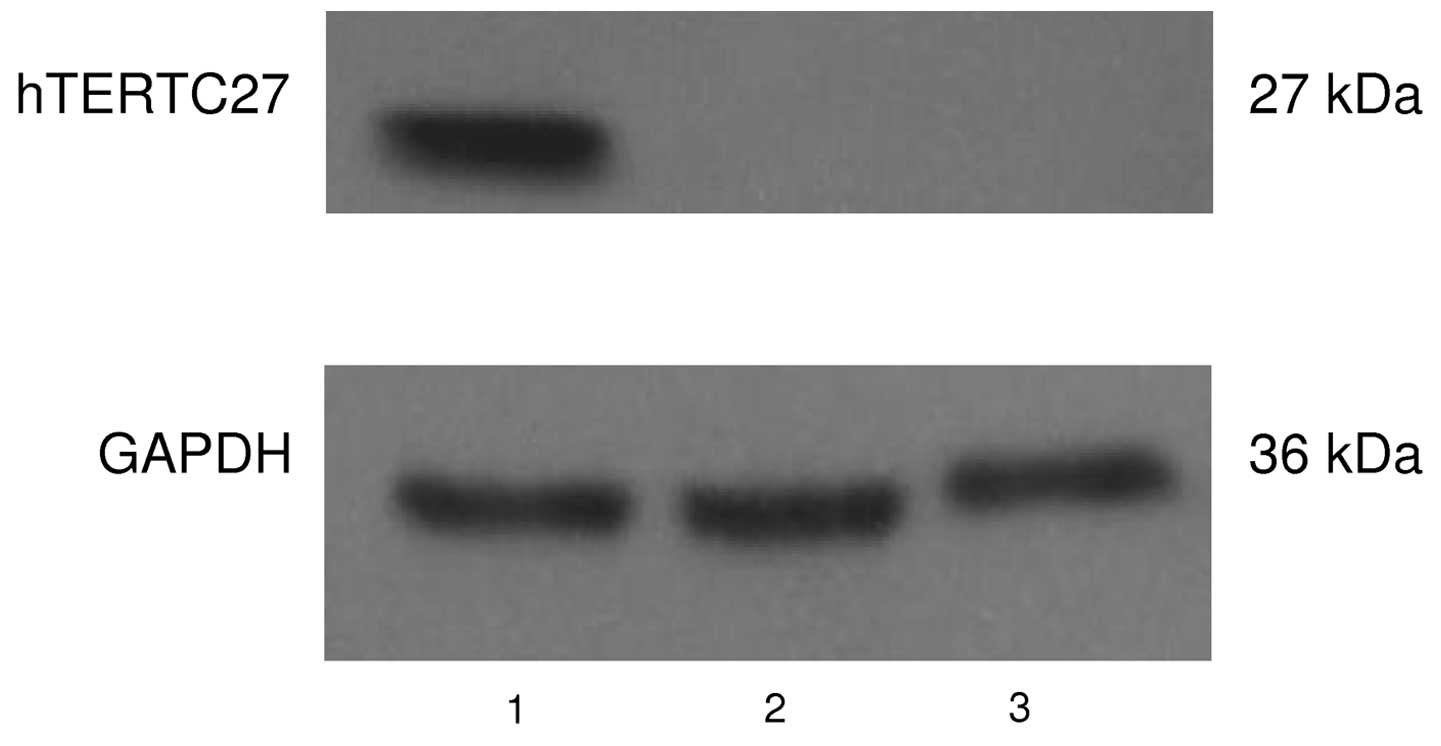
hTERTC27 protein was readily detectable in the DCs 48 h after the
adenoviral infection by western blot analysis using an anti-hTERT
antibody. As shown in Fig. 3, only
the DCs transduced with rAd-C27 expressed hTERTC27 protein. These
results suggest that hTERTC27 gene was successfully transfected
into DCs and the adenovirus-mediated gene transfer was highly
efficient.
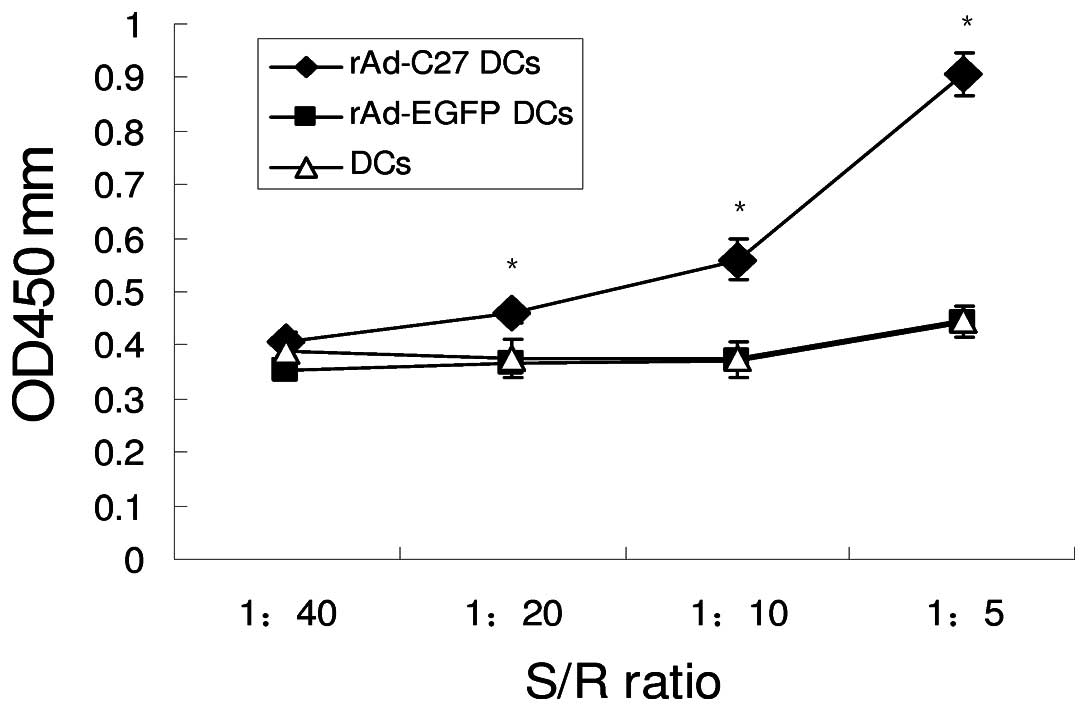
Improved proliferation of T cells
rAd-C27 and rAd-EGFP transduced DCs respectively on
Day 3 after transduction were used to induce T cell proliferation.
Allogeneic T cells were mixed with 1×105 rAd-27 DCs,
rAd-EGFP DCs and normal control DCs, at stimulator/responder ratios
of 1:40, 1:20, 1:10, 1:5, and incubated for 3 days (Fig. 4). rAd-27 DCs demonstrated identical
levels of stimulation which was much higher than the other two
groups at S/R ratios from 1:20 to 1:5 (P<0.05), while the levels
of rAd-EGFP DCs and DCs were nearly identical. These results
indicate that the enhancement was related to the transgene
expression and not due to adenoviral infection. Therefore, rAd-27
DCs could induce stronger proliferation of T cells.
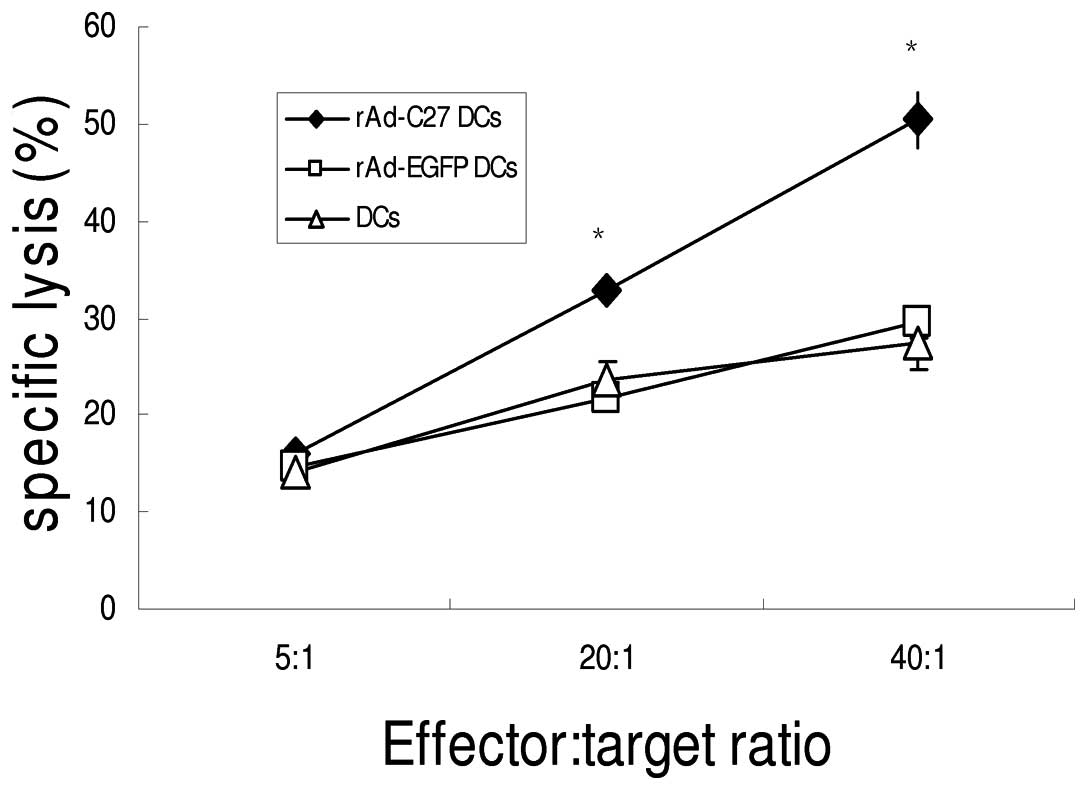
Induced CTL responses in vitro
The functional capability of the CTL responding to
adenovirus-infected DCs was examined by determining whether they
could specifically lyse tumor cells. The T cell lines were
generated from autologous mononuclear cells and were plated in
96-well plates in medium containing IL-2 (2 ng/ml). The DCs were
added at varying T cell-to-DC ratios and co-cultured at 37°C in 5%
CO2. As a result, rAd-C27 DCs could induce specific CTL
activity against hTERT positive GL261 cells (Fig. 5). The cytotoxic activity of rAd-C27
DCs was 50.38±2.95% at a 40:1 effector/target ratio (E/T), while no
obvious lysis by rAd-EGFP DCs or DCs was detected, even at the
highest E/T ratio (29.53±1.49%, 27.53±2.71%). These results clearly
demonstrate that the CTL were mainly induced by hTERTC27
peptides.
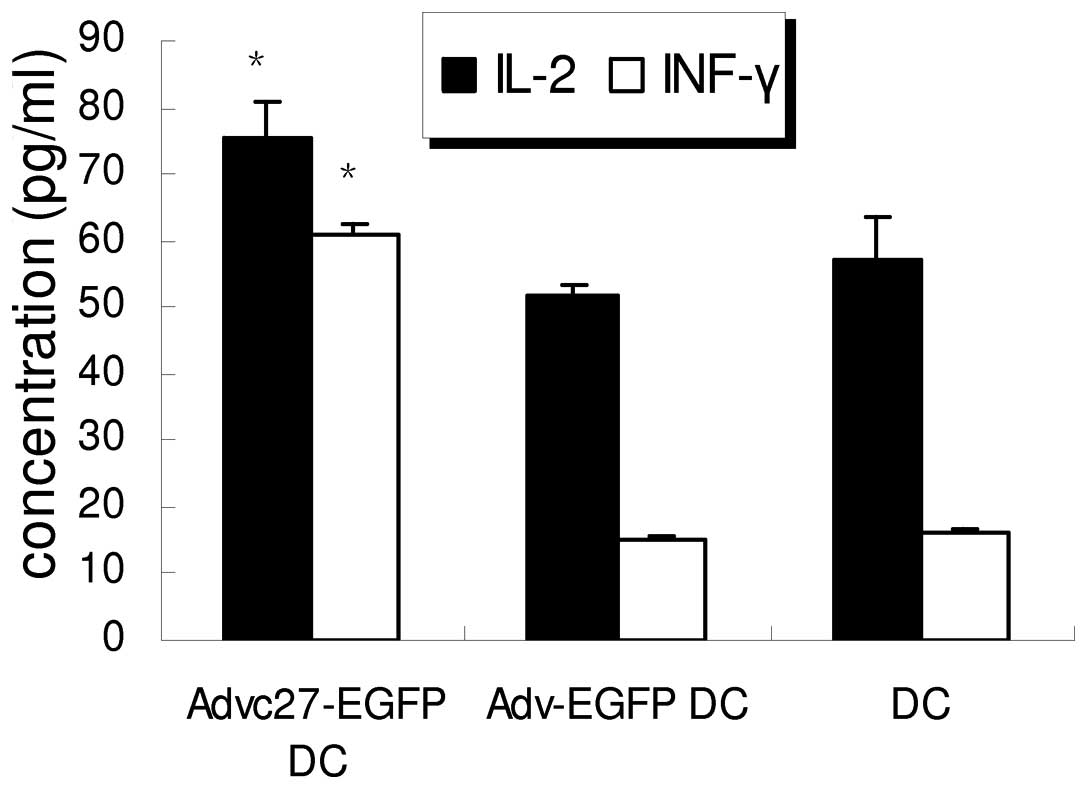
Augmentation of the concentration of IL-2
and IFN-γ in the supernatants of T cells
The T cells were co-cultured with the rAd-C27 DCs,
rAd-EGFP DCs and normal control DCs for 3 days. IL-2 and IFN-γ in
the supernatant was determined with ELISA (Fig. 6). The T cells co-cultured with
rAd-C27 DCs produced 75.54±5.32 pg/ml of IL-2, while IL-2 produced
by rAd-EGFP DCs group and DCs group was 51.83±1.39 and 57.23±6.30
pg/ml, which both were lower than rAd-C27 DCs group (P=0.001).
Also, the concentration of IFN-γ in rAd-C27 DCs group was higher
than other groups. These results suggest that an hTERTC27 vaccine
may increase IL-2 and IFN-γ secretion to enhance the immune
response.
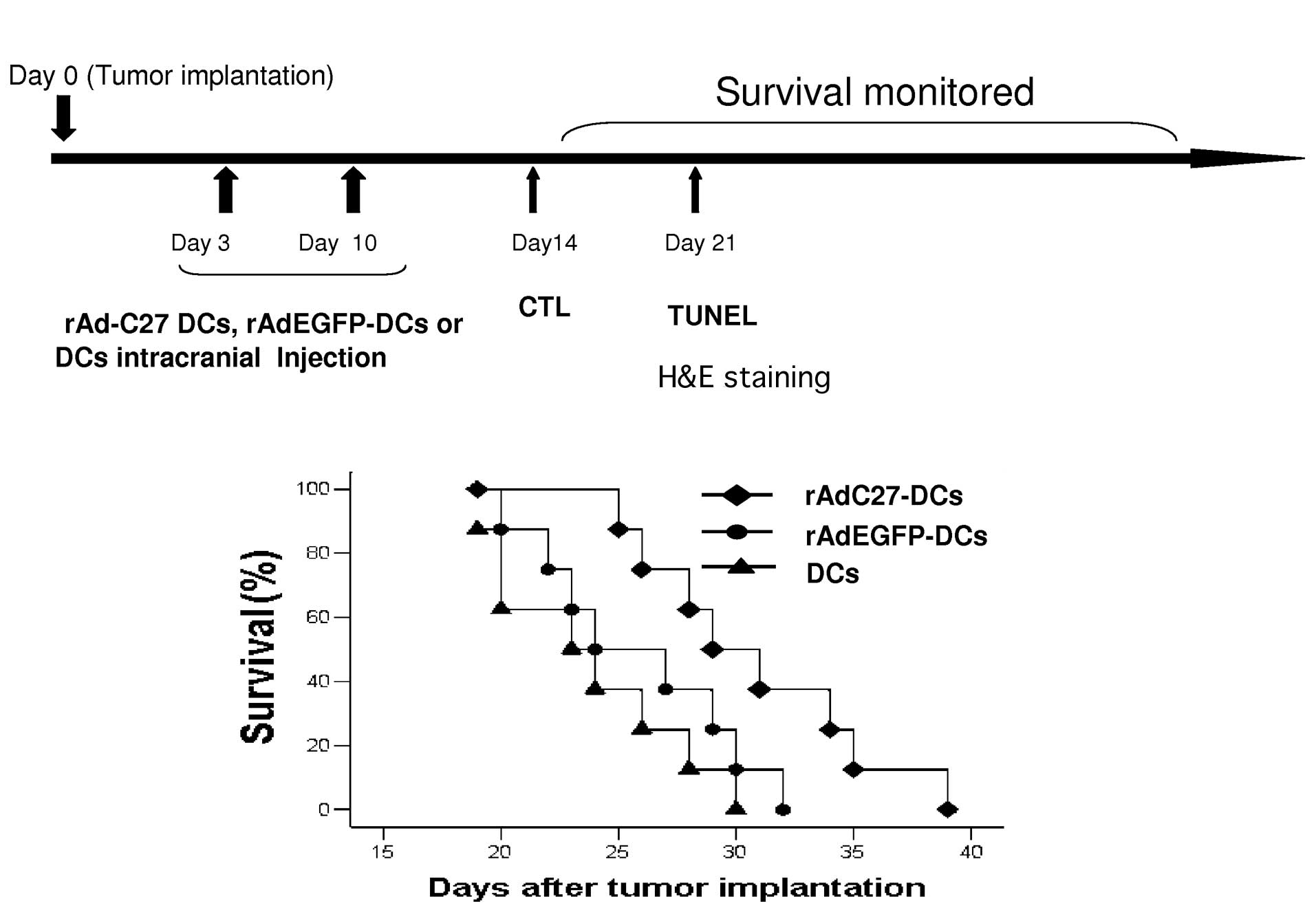
Prolongation of the survival time of
tumor bearing mice
To examine whether the rAd-C27 DCs provided a
therapeutic benefit for brain tumor, we implemented DCs
immunotherapy in an established mouse glioma model. rAd-C27 DCs,
rAd-EGFP DCs or DCs (1×105 each) were intracerebrally
injected on Day 3 and Day 10 after the tumor implantation (Fig. 7A). The administration of Ad-C27 DCs
resulted in a significantly prolonged survival compared with
rAd-EGFP DCs or DCs (Fig. 7B).
Approximately 50% of the rAd-C27 DCs-treated mice survived beyond
an observation period of 30 days, but almost all mice treated with
rAd-EGFP-DCs and DCs survived for 30 days. Statistical analysis
revealed that the effect of rAd-C27 DCs was significantly different
from that of rAd-EGFP DCs or DCs (P=0.005). In addition, there was
no significant difference between the DC-empty and non-transduced
DC treatments (P=0.307).
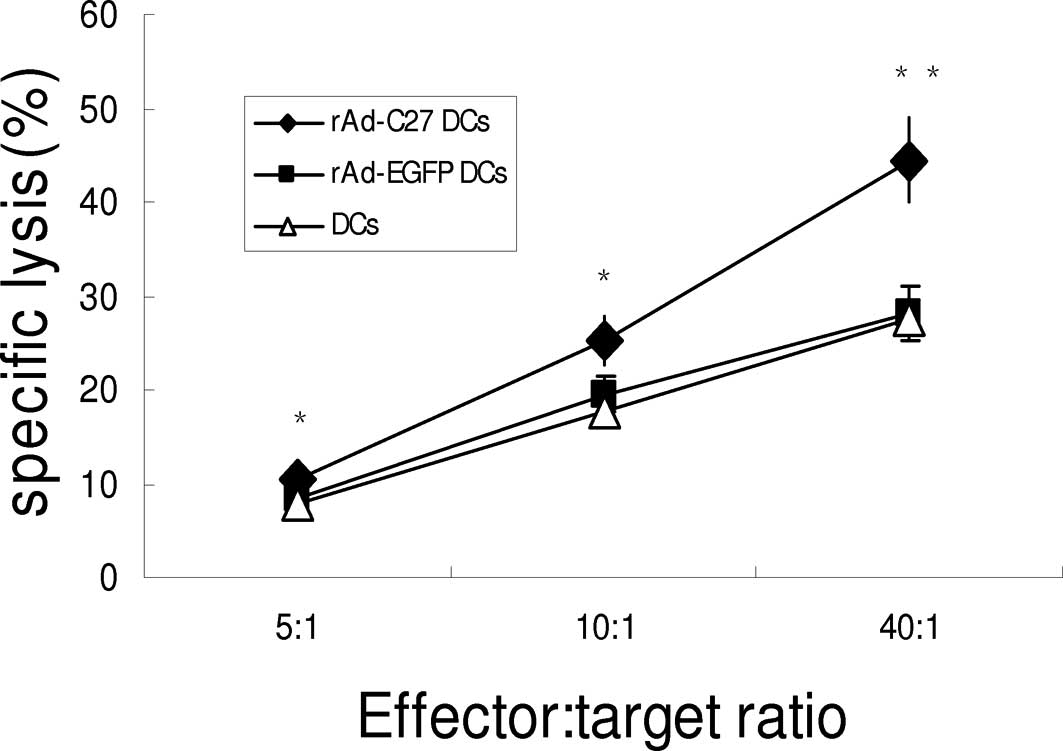
Induced cytotoxicity against murine
gliomas in vivo
To further evaluate whether intratumoral injections
with rAd-C27 DCs influence the induction of tumor-specific T cell
responses in tumor-bearing mice, C57BL/6 mice were immunized twice,
with a 7-day interval, with rAd-C27 DCs, rAd-EGFP-DCs or DCs. Mice
that had received rAd-C27 DCs therapy exhibited remarkable
GL261-specific CTL responses (Fig.
8). The cytotoxicity elicited by rAd-C27 DCs was much higher
than the other groups at the E/T ratios of 5:1, 10:1 and to 40:1
(P<0.05). Therefore, rAd-C27 DCs could induce antigen-specific
cytotoxic T lymphocytes against gliomas in vitro and in
vivo.
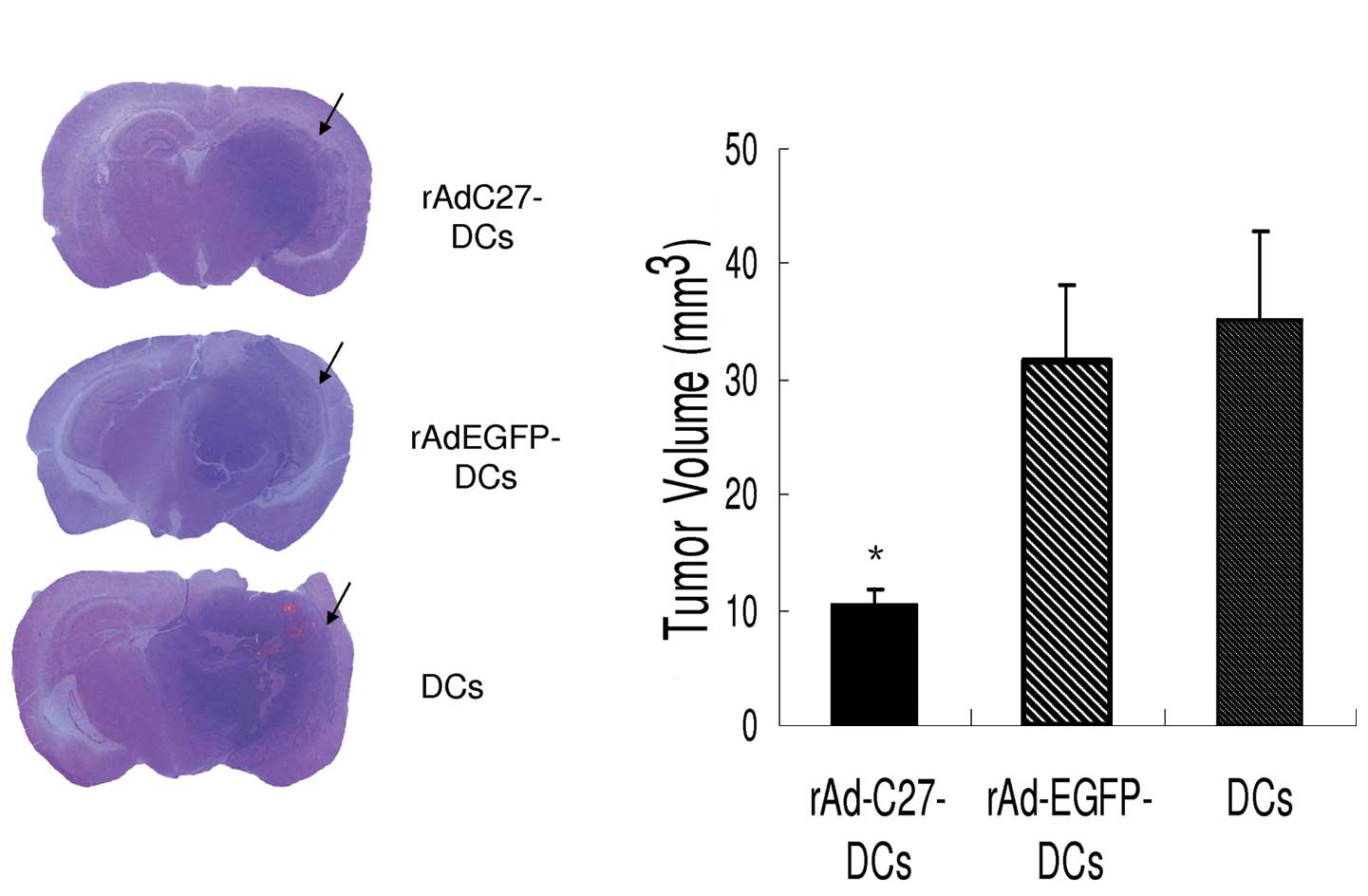
Inhibition of the tumor growth in C57BL
mice
On Day 21 after tumor implantation, four mice from
each group were euthanized to obtain brain tissues and compare the
tumor volume. These tissues stained with H&E clearly demarcated
the tumor outline. Thus, it was verified that the tumor
transplantation had been successfully performed in all tissues
(Fig. 9). The average tumor size
for the DC groups was 35.21±7.70 mm3, while it was
31.50±6.79 mm3 for the rAd-EGFP-DCs group. For the
groups treated with rAd-C27 DCs, the average tumor sizes were
10.53±1.24 mm3. According to our results, the rAd-C27
DCs treatment could significantly diminish the glioma tumor size
when compared with the other groups (P<0.001).
Discussion
In this study, we have demonstrated that hTERTC27
could increase T cell proliferation, augment the concentration of
IL-2 and IFN-γ in the supernatants of T cells. Furthermore, it
demonstrated the ability to mount a strong and specific CTL
response against the glioblastoma cells in vitro and in
vivo. It could prolong the survival time and inhibit the growth
of gliomas-bearing mice.
Cancer vaccine targeting tumor antigens has
attracted much attention in recent years because of its higher
specificity and lower toxicity than non-surgical approaches
(25,26). Cancer vaccines can be categorized
into peptide/protein vaccines, tumor cell vaccine, DNA vaccines,
recombinant virus vaccine as well as DC vaccines (25). The ascendancy of using DC vaccines
in cancer treatment is that tumor antigen(s) or DNA encoding
antigen(s) will be taken up and processed easily. Here, we found
that rAd-C27 DCs not only promoted T-cell proliferation, but also
triggered a specific CTL response to lyse cancer cells. However,
Huo et al (21) reported
that the rAAV-/rAdv-hTERTC27 viral cocktail only revoked the NK
cell response and could not induce a T lymphocyte response in the
mice, which was not in accordance with our study. We presumed that
the antigens contain in the rAAV-/rAdv-hTERTC27 were not sufficient
to stimulate T lymphocytes, and DCs could present the antigens and
augment the antigens' influence. Our presumption is supported by
the following evidence. Nestle et al reported that peptide-
or tumor lysate-loaded DCs can lead to a systemic response
effective in inducing tumor regressions in the clinical study. By
contrast, administration of melanoma peptides, without DCs, failed
to elicit clinically significant responses (27). Therefore, DC vaccines may minimize
the immune escape because of loss of antigen expression and were
considered as a promising approach to tumour immunotherapy.
As a universal tumor-associated antigen, TERT is an
ideal target for cancer therapy. Several immunogenic hTERT peptides
have been discovered, (28) p973
and p988 were definitively contained in hTERTC27 (29,30).
p988 was restricted to the MHC class I allele HLA-A*0201
(HLA-A2), the most frequently expressed HLA allele found among
nearly 50% of Caucasians, Asians, and Hispanics and 33% of
African-Americans. This evidence suggests that hTERTC27 may be
utilized in tumor patients from different areas. Both p973 and p988
were demonstrated to induce specific CD8+ T cell
responses in vitro or in vivo to lyse
hTERT+ tumor cells. More importantly, hTERT-specific CTL
lysed neither telomerase-negative normal cell nor
telomerase-positive CD34+ hematopoietic progenitor cells
(31). Activated B-cells are
susceptible to hTERT-specific lysis and notably represent the only
cells other than tumor cells that to date have been demonstrated to
undergo hTERT-specific lysis in vitro. In fact, we did not
find the growth difference between DCs transfected by AdvC27-EGFP
and that transfected by Adv-EGFP in our study (data not shown).
Therefore, the immunotherapy targeting hTERTC27 was possibly a safe
and effective strategy against carcinoma.
IL-2 acts as an antitumor agent by increasing the
cytolytic activity of antigen-specific cytotoxic T lymphocytes and
NK cells and by increasing the gene expression responsible for
encoding the lytic component of cytotoxic granules, ie, perforin
and granzymes (32,33). IFN-γ, the product of Th1, CTL, and
NK cells, is one of the major effector molecules in cell-mediated
immunity. It plays important roles in the induction of CTL and
differentiation of Th1 cells. It also leads to an increase in MHC
class I and II expression and contributes to efficient antigen
presentation to lymphocytes (34,35).
In the present study, T cell lines co-cultured with
AdvC27-EGFP-transduced DCs produced a high level of IL-2 and IFN-γ.
It was possible that the high level of cytokine was one of the main
reasons why AdvC27-EGFP-transduced DCs could induce a strong CTL
response against the cancer cells. All in all, in this study, we
have demonstrated that DCs transduced with AdvC27-EGFP are able to
induce a potent antitumor immune response against hTERT-positive
tumor cells, which will be instrumental for the clinical
application of hTERTC27 in cancer therapy.
Acknowledgements
This research was supported by the ‘863’ Programs of
China (2007AA021101 to Y.P.), National Natural Science Foundation
of China (NSFC) (30672411 and 30973479 to Y.P.) and the Science and
Technology Planning Project of Guangdong Province, China
(2009B060700040 to Y.P.).
Abbreviations:
|
DCs
|
dendritic cells
|
|
IL-2
|
interleukin-2
|
|
IFN-γ
|
interferon-γ
|
|
CTL
|
cytotoxic T lymphocytes
|
|
TAA
|
tumor-associated antigen
|
|
APC
|
antigen presenting cells
|
|
hTERT
|
human telomerase reverse
transcriptase
|
|
C27
|
hTERTC27
|
|
rAd
|
recombinant adenoviral
|
|
MLR
|
mixed lymphocyte reaction
|
|
NK
|
natural killer
|
References
|
1
|
Tanaka H, Shinto O, Yashiro M, et al:
Transforming growth factor beta signaling inhibitor, SB-431542,
induces maturation of dendritic cells and enhances anti-tumor
activity. Oncol Rep. 24:1637–1643. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kushwah R and Hu J: Complexity of
dendritic cell subsets and their function in the host immune
system. Immunology. 133:409–419. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Phuc PV, Lam DH, Ngoc VB, Thu DT, Nguyet
NT and Ngoc PK: Production of functional dendritic cells from
menstrual blood-a new dendritic cell source for immune therapy. In
Vitro Cell Dev Biol Anim. 47:368–375. 2011.PubMed/NCBI
|
|
4
|
Guermonprez P, Valladeau J, Zitvogel L,
Thery C and Amigorena S: Antigen presentation and T cell
stimulation by dendritic cells. Annu Rev Immunol. 20:621–667. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pawlowska AB, Hashino S, McKenna H, Weigel
BJ, Taylor PA and Blazar BR: In vitro tumor-pulsed or in vivo Flt3
ligand-generated dendritic cells provide protection against acute
myelogenous leukemia in non-transplanted or syngeneic bone
marrow-transplanted mice. Blood. 97:1474–1482. 2001. View Article : Google Scholar
|
|
6
|
Onaitis M, Kalady MF, Pruitt S and Tyler
DS: Dendritic cell gene therapy. Surg Oncol Clin N Am. 11:645–660.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Trivedi D, Williams RY, O'Reilly RJ and
Koehne G: Generation of CMV-specific T lymphocytes using
protein-spanning pools of pp65-derived overlapping
pentadecapeptides for adoptive immunotherapy. Blood. 105:2793–2801.
2005. View Article : Google Scholar
|
|
8
|
Bachleitner-Hofmann T, Friedl J, Hassler
M, et al: Pilot trial of autologous dendritic cells loaded with
tumor lysate(s) from allogeneic tumor cell lines in patients with
metastatic medullary thyroid carcinoma. Oncol Rep. 21:1585–1592.
2009. View Article : Google Scholar
|
|
9
|
Xiao ZY, Tang H, Xu ZM, et al: An
experimental study of dendritic cells transfected with cancer
stem-like cells RNA against 9L brain tumors. Cancer Biol Ther.
11:974–980. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Palucka AK, Ueno H, Connolly J, et al:
Dendritic cells loaded with killed allogeneic melanoma cells can
induce objective clinical responses and MART-1 specific
CD8+ T-cell immunity. J Immunother. 29:545–557. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun L, Liu J, Cao X, et al: Enhancement of
antigen presenting function of dendritic cells by IL-2 gene
modification and its mechanism. Zhonghua Xue Ye Xue Za Zhi.
23:247–250. 2002.(In Chinese).
|
|
12
|
Harrington L, Zhou W, McPhail T, et al:
Human telomerase contains evolutionarily conserved catalytic and
structural subunits. Genes Dev. 11:3109–3115. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Collins K and Mitchell JR: Telomerase in
the human organism. Oncogene. 21:564–579. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shay JW, Zou Y, Hiyama E and Wright WE:
Telomerase and cancer. Hum Mol Genet. 10:677–685. 2001. View Article : Google Scholar
|
|
15
|
Zhang P, Chen Y, Jiang X, Tu Z and Zou L:
Tumor-targeted efficiency of shRNA vector harboring chimera
hTERT/U6 promoter. Oncol Rep. 23:1309–1316. 2010.PubMed/NCBI
|
|
16
|
Li H, Wang J, Zhou T, Zhang Y and Zhang Z:
An investigation of the effects of nanosize delivery system for
antisense oligonucleotide on esophageal squamous cancer cells.
Cancer Biol Ther. 7:1852–1859. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Keith WN, Bilsland A, Evans TR and
Glasspool RM: Telomerase-directed molecular therapeutics. Expert
Rev Mol Med. 4:1–25. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vonderheide RH, Hahn WC, Schultze JL and
Nadler LM: The telomerase catalytic subunit is a widely expressed
tumor-associated antigen recognized by cytotoxic T lymphocytes.
Immunity. 10:673–679. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang JJ, Lin MC, Bai YX, et al: Ectopic
expression of a COOH-terminal fragment of the human telomerase
reverse transcriptase leads to telomere dysfunction and reduction
of growth and tumorigenicity in HeLa cells. Cancer Res.
62:3226–3232. 2002.
|
|
20
|
Ng SS, Gao Y, Chau DH, et al: A novel
glioblastoma cancer gene therapy using AAV-mediated long-term
expression of human TERT C-terminal polypeptide. Cancer Gene Ther.
14:561–572. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huo L, Yao H, Wang X, Wong GW, Kung HF and
Lin MC: Inhibition of melanoma growth by subcutaneous
administration of hTERTC27 viral cocktail in C57BL/6 mice. PLoS
One. 5:e127052010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang JY, Cao DY, Liu WC, Zhang HM, Teng ZH
and Ren J: Dendritic cell generated from CD34+
hematopoietic progenitors can be transfected with adenovirus
containing gene of HBsAg and induce antigen-specific cytotoxic T
cell responses. Cell Immunol. 240:14–21. 2006.PubMed/NCBI
|
|
23
|
Chen T, Tang XD, Wan Y, et al:
HLA-A2-restricted cytotoxic T lymphocyte epitopes from human
heparanase as novel targets for broad-spectrum tumor immunotherapy.
Neoplasia. 10:977–986. 2008.PubMed/NCBI
|
|
24
|
Inaba K, Inaba M, Romani N, et al:
Generation of large numbers of dendritic cells from mouse bone
marrow cultures supplemented with granulocyte/macrophage
colony-stimulating factor. J Exp Med. 176:1693–1702. 1992.
View Article : Google Scholar
|
|
25
|
Berzofsky JA, Terabe M, Oh S, et al:
Progress on new vaccine strategies for the immunotherapy and
prevention of cancer. J Clin Invest. 113:1515–1525. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vonderheide RH, Domchek SM, Schultze JL,
et al: Vaccination of cancer patients against telomerase induces
functional antitumor CD8+ T lymphocytes. Clin Cancer
Res. 10:828–839. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nestle FO, Alijagic S, Gilliet M, et al:
Vaccination of melanoma patients with peptide- or tumor
lysate-pulsed dendritic cells. Nat Med. 4:328–332. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huo LF, Tang JW, Huang JJ, et al: Cancer
immunotherapy targeting the telomerase reverse transcriptase. Cell
Mol Immunol. 3:1–11. 2006.PubMed/NCBI
|
|
29
|
Vonderheide RH, Anderson KS, Hahn WC,
Butler MO, Schultze JL and Nadler LM: Characterization of
HLA-A3-restricted cytotoxic T lymphocytes reactive against the
widely expressed tumor antigen telomerase. Clin Cancer Res.
7:3343–3348. 2001.PubMed/NCBI
|
|
30
|
Scardino A, Gross DA, Alves P, et al:
HER-2/neu and hTERT cryptic epitopes as novel targets for broad
spectrum tumor immunotherapy. J Immunol. 168:5900–5906. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Minev B, Hipp J, Firat H, Schmidt JD,
Langlade-Demoyen P and Zanetti M: Cytotoxic T cell immunity against
telomerase reverse transcriptase in humans. Proc Natl Acad Sci USA.
97:4796–4801. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rabinovich PM, Komarovskaya ME, Wrzesinski
SH, et al: Chimeric receptor mRNA transfection as a tool to
generate antineoplastic lymphocytes. Hum Gene Ther. 20:51–61. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Iuchi T, Teitz-Tennenbaum S, Huang J, et
al: Interleukin-21 augments the efficacy of T-cell therapy by
eliciting concurrent cellular and humoral responses. Cancer Res.
68:4431–4441. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dredge K, Marriott JB, Todryk SM, et al:
Protective antitumor immunity induced by a costimulatory
thalidomide analog in conjunction with whole tumor cell vaccination
is mediated by increased Th1-type immunity. J Immunol.
168:4914–4919. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Pletneva M, Fan H, Park JJ, et al:
IFN-producing killer dendritic cells are antigen-presenting cells
endowed with T-cell cross-priming capacity. Cancer Res.
69:6607–6614. 2009. View Article : Google Scholar : PubMed/NCBI
|