Introduction
Lung cancer is one of the most common human cancers
and the leading cause of cancer death worldwide (1,2). At
present, the treatment of lung cancer mainly includes surgery,
radiotherapy and chemotherapy. Chemotherapy plays an important role
in the treatment of lung cancer, but it is limited to a significant
extent by its toxicities, side effects and drug resistance
(3,4). The mortality in lung cancer patients
remains high and the 5-year overall survival in many countries
generally is <15% (5).
Therefore, it is necessary to search for drugs capable of
preventing and treating lung cancer and other malignancies.
Recently, the use of natural compound as anticancer agents has
gained a great deal of attention for their possible therapeutic
qualities with minimal toxicity (6–9).
Some natural agents suppress the proliferation of
cancer cells by inducing apoptosis (10). The expression of several genes has
been demonstrated to be critical for the regulation of apoptosis
such as Bcl-2 family proteins. The members of the Bcl-2 family play
an important role in the regulation of the apoptotic pathway
(11). In the Bcl-2 family members,
Bcl-2 is the most important anti-apoptotic protein, whereas Bax is
a pro-apoptotic protein (12). The
ratio of Bax/Bcl-2 is critical for the induction of apoptosis and
this ratio determines whether cells will undergo apoptosis
(13).
MicroRNAs (miRNAs) are a kind of non-coding small
RNAs that can post-transcriptionally regulate the expression of
hundreds of their target genes (14,15).
miRNAs play an important role in diverse cellular processes, such
as development, proliferation, differentiation and apoptosis
(16,17), some miRNAs act as oncogenes or tumor
suppressors in tumorigenesis (18,19).
Recent evidence indicates that some anticancer agents suppress the
proliferation of malignant cells by regulation expression of miRNAs
(20–22).
Waltonitone (Fig.
1), an ursane-type pentacyclic triterpene isolated from Chinese
medical plant Gentiana waltonii Burkill, has been shown to
inhibit cell growth and induces cell apoptosis in human
hepatocellular carcinoma (23).
However, since the antitcancer effect of waltonitone in lung cancer
is unknown, we explored the effect of waltonitone treatment in lung
cancer A549 cells and investigated the mechanisms underlying the
inhibitory effect of waltonitone in lung cancers.
Materials and methods
Reagents and chemicals
RPMI-1640, penicillin, streptomycin, and trypsin
were purchased from Biological Industries (Kibbutz Beit Haemek,
Israel). Fetal bovine serum (FBS) was purchased from Solarbio
(Beijing Solarbio Science and Technology, China). 3-(4,5-Dimethyl
thiazol-2yl)-2,5-diphenyltetrazolium bromide (MTT), dimethyl
sulfoxide (DMSO) kit was purchased from Sigma-Aldrich (St. Louis,
MO, USA). Annexin V-FITC and PI double staining kit were purchased
from KeyGene (Nanjing, China). Waltonitone (HPLC purity ≥98%) was
purchased from Shanghai R&D Centre for Standardization of
Chinese Medicines (Shanghai, China). A 100-mM stock solution of
waltonitone was prepared in DMSO and stored at −20°C. Antibodies
against Bcl-2 and Bax were purchased from Santa Cruz Biotechnology
(Santa Cruz, CA). Horseradish peroxidase-conjugated secondary
antibody was obtained from Santa Cruz Biotechnology. TRIzol reagent
was from Invitrogen (CA, USA). All other chemicals were obtained
from Sinopharm Chemical Reagent Co., Ltd. (Shenyang, China).
Cell culture
A human lung cancer A549 cell line was obtained from
the China Center for Type Culture Collection (Wuhan, China). A549
cells were cultured in RPMI-1640 supplemented with 10% FBS, 100
U/ml penicillin, and 100 μg/ml streptomycin at 37°C in a humidified
atmosphere of 5% CO2. Culture medium was changed every
day. Cells for assays were detached by a solution of 0.125% trypsin
and 0.02% EDTA.
Cell viability assay
Viability was assessed by MTT assay. A549 cells were
plated at a density of 1×104 cells per well in 96-well
plates overnight. Cells were treated with different concentrations
of waltonitone (5, 10, 20, 30, 40 and 50 μmol/l). After exposure to
waltonitone for 24, 48, 72 h, 25 μl of MTT solution (2 mg/ml in
PBS) was added to each well and the plates were incubated for an
additional 4 h at 37°C. Then the medium was totally removed and 150
μl of DMSO were added to each well to solubilize the formazan
crystals formed in viable cells. Finally, the plates were shaken
and the optical density was determined at 570 nm (OD570) using
ELISA plate reader (Model 550, Bio-Rad, Laboratories, Hercules, CA,
USA). This procedure was replicated three times.
Fluorescent microscopy
The morphology of apoptotic cells was detected by
nuclear staining with Hoechst 33342. A549 cells (5×105)
were grown on coverslips placed into 6-well plates overnight. Cells
were treated with different concentrations of waltonitone (0 and 20
μmol/l). After 24 h, the cells were washed twice with cold PBS,
then fixed with cold methanol and acetic acid (3/1, v/v) at 4°C
overnight and stained with Hoechst 33342 for 30 min in the dark,
washed again in PBS and finally mounted in mounting medium (80%
glycerol in PBS). Processed cells were observed with a fluorescence
microscope (Nikon, Japan).
Annexin V/PI flow cytometric
analysis
The number of apoptotic cell death induced by
waltonitone was measured by flow cytometry using Annexin V-FITC
Apoptosis kit. Briefly, following treatment with waltonitone (0,
10, 20 and 30 μmol/l) for 24 h, 1×106 cells were
harvested by centrifugation (1,000 rpm/min), washed twice with cold
PBS. The cell pellet was re-suspended in 1X binding buffer at a
concentration of 1×106 cells/ml. Cell suspension (100
μl) was transferred into FCM tube. Annexin V-FITC (5 μl) and 10 μl
of PI were added into the cell suspension, followed by gentle
vortexing. The cells were incubated at room temperature for 15 min
in the dark. An additional 400 μl of 1X binding buffer was added to
each tube. The cells were analyzed using a FACScan flow cytometer
and analyzed using CellQuest software (Becton-Dickinson, Redlands,
CA).
Western blot analysis
The expression of apoptosis-related proteins was
evaluated by western blot analysis. In brief, 3×107
cells were incubated with the designated doses of waltonitone (0,
10, 20 and 30 μmol/l) for 24 h. The cells were then washed in PBS
and suspended in five volumes of lysis buffer (20 mM HEPES, pH 7.9,
20% glycerol, 200 mM KCl, 0.5 mM EDTA, 0.5% NP40, 0.5 mM DTT, 1%
protease inhibitor cocktail). Lysates were then collected and
stored at −20°C until further use. Supernatant protein
concentration was determined by the Bradford method. Supernatant
samples containing 40 μg of total protein were resolved by SDS-PAGE
gel depending on the target protein sizes, transferred to PVDF
membranes by electroblotting, and probed with anti-Bax, anti-Bcl-2
and anti-actin. Membranes were incubated with horseradish
peroxidase-conjugated secondary antibodies. Blots were developed
using an ECL kit.
Microarray analysis of miRNA
profiles
miRNA microarray was used to investigate the
differentially expressed miRNAs in A549 cells after waltonitone
treated. A549 cells were treated with waltonitone (0 and 30
μmol/l). Total RNA was extracted from cells using TRIzol reagent 24
h after waltonitone treatment. Total RNA samples were analyzed by
CapitalBio Corp. for miRNAs microarray experiments. The GeneChip
miRNA Array containing probes complementary to 847 human miRNAs
registered in miRBase 11.0. Experimental procedures were performed
as described in detail on the website of CapitalBio (http://www.capitalbio.com). A miRNA was determined as
differentially expressed if its expression change was >1.5-fold,
and it was identified as significantly changed using the
Significance Analysis of Microarrays method with FDR <0.05.
Classification and target prediction of
miRNAs
The candidate target genes of miRNAs are predicted
and analyzed by miRanda (http://www.ebi.ac.uk/enright-srv/microcosm/htdocs/targets/v5/),
PicTar (http://pictar.mdc-berlin.de/) and
TargetScan (http://www.targetscan.org/), and we chose the genes
found by all the three software as the miRNA targets. Then, human
genes related to cell proliferation and apoptosis were selected
from the Gene Ontology database (http://www.geneontology.org/), and the candidate genes
predicted as targets of the miRNAs and those selected based on gene
ontology were aligned by their gene names. Genes appearing in both
lists were chosen and listed.
Statistical analysis
All experiments were conducted three times. Data
were expressed as mean ± SD. Statistical correlation of data was
checked for significance by ANOVA and Student's t-test. The
statistical significance was defined as P<0.05. These analyses
were performed using SPSS 11.0 software.
Results
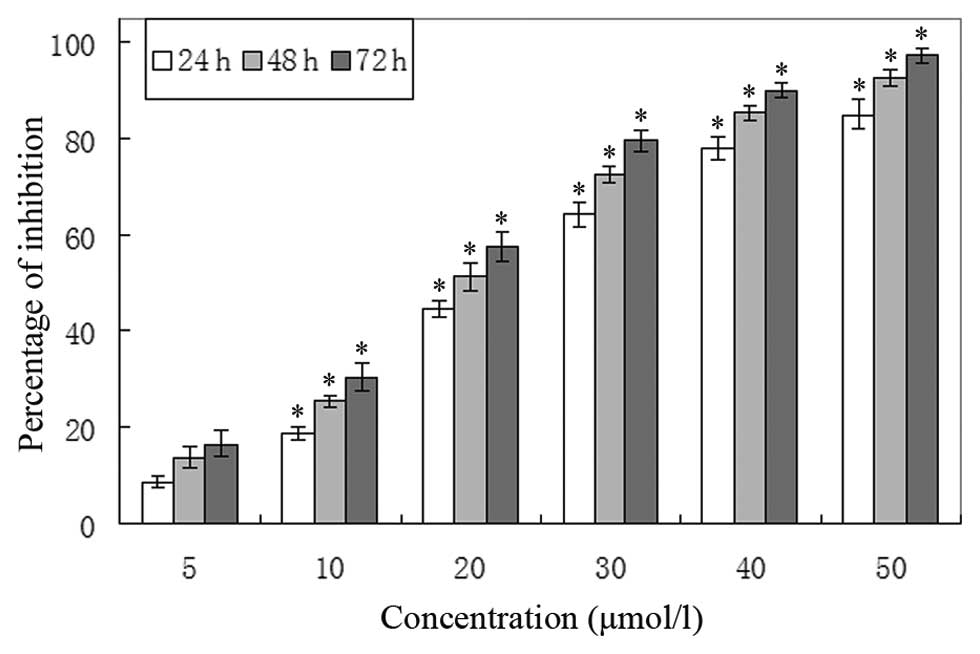
Waltonitone inhibits A549 cell
proliferation
In order to evaluate the growth inhibition effects
of waltonitone on A549 cells, the cells were treated with various
concentrations of waltonitone for 24, 48 and 72 h. As shown in
Fig. 2, waltonitone had significant
growth inhibitory effects on A549 cells in a concentration- and
time-dependent manner.
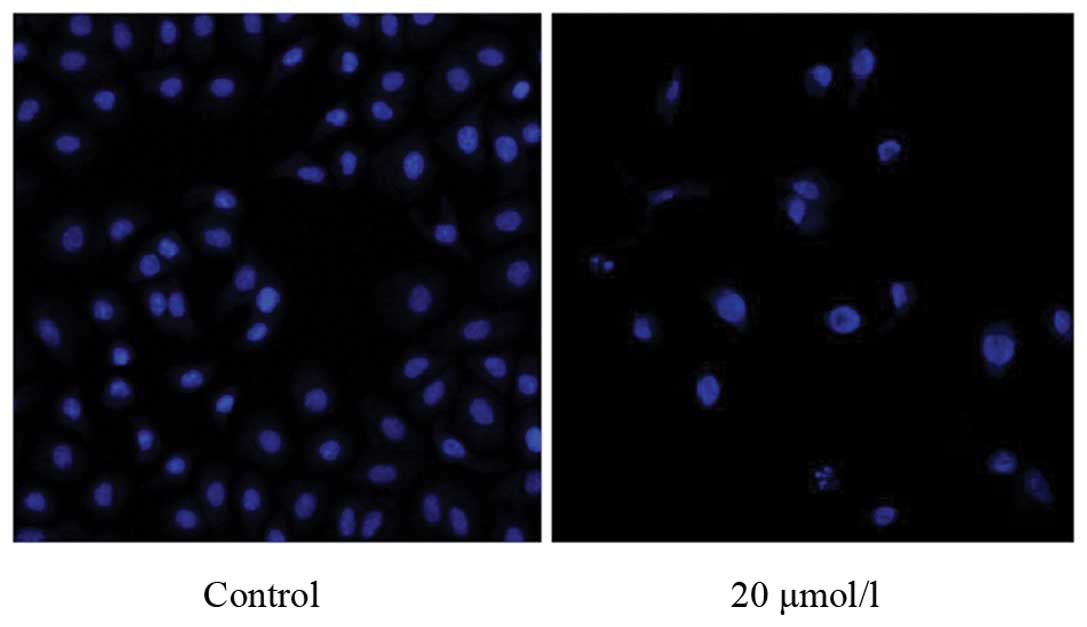
Waltonitone induces A549 apoptosis
Fluorescent microscopy analysis exhibited different
morphological alterations in A549 cells after treatment with
waltonitone (0 and 20 μmol/l) for 24 h. As showed in Fig. 3, condensation of chromatin, nuclear
fragmentations and apoptotic bodies were found clearly in treated
A549 cells using Hoechst 33324 staining. The results showed that
when exposed to waltonitone (20 μmol/l), A549 cells underwent the
typical morphologic changes of apoptosis.
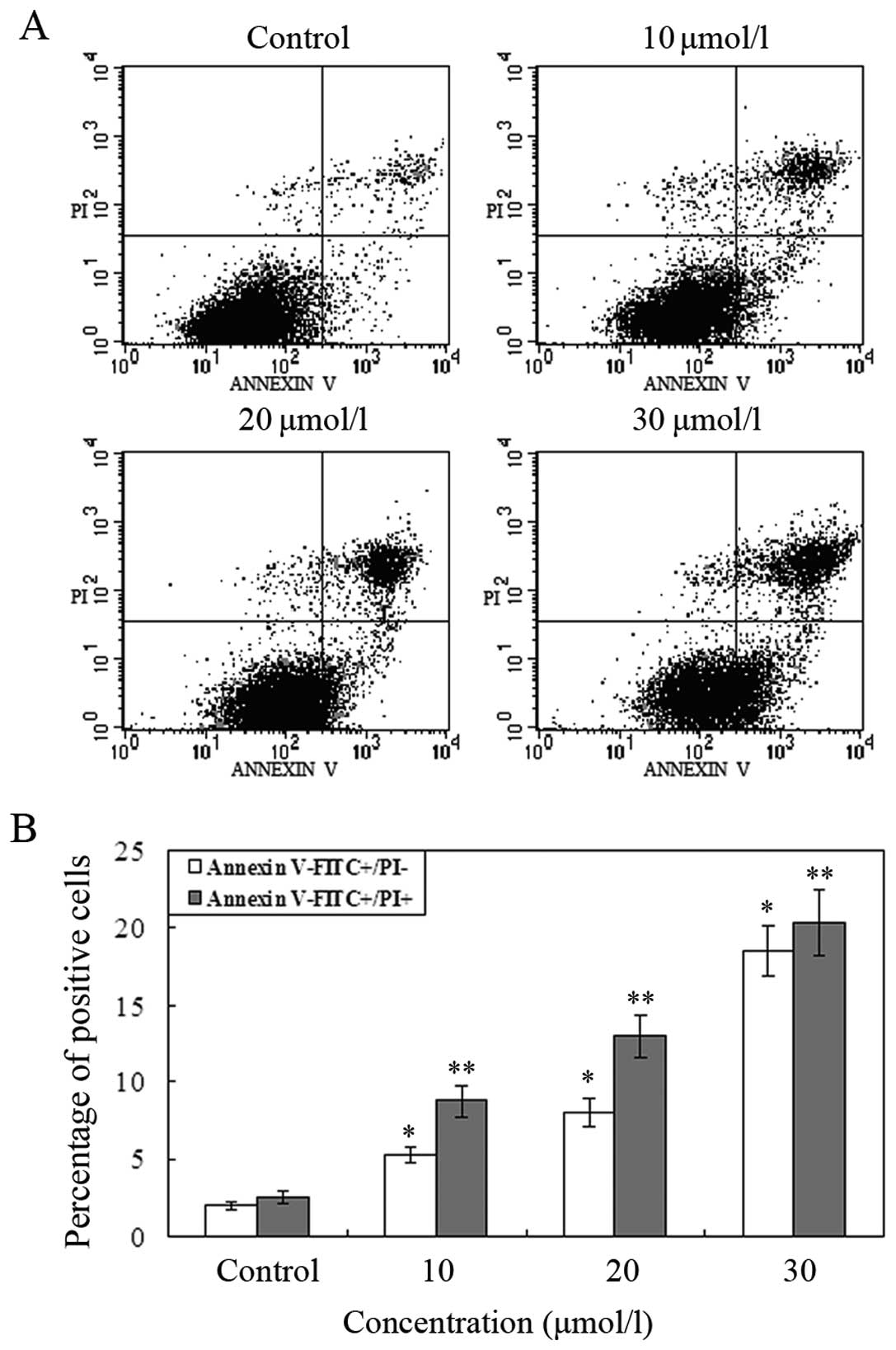
The number of apoptotic cell death induced by
waltonitone was measured by Annexin V/PI staining. A549 cells were
treated with different concentrations of waltonitone (0, 10, 20,
and 30 μmol/l) for 24 h and were analyzed by flow cytometry. As
displayed in Fig. 4, the numbers of
early and late apoptotic cells were significantly increased
compared to control group. The results show that when treated with
waltonitone for 24 h, the ratio of apoptotic cells significantly
increased in a concentration-dependent manner.
Waltonitone induced expression of
apoptosis-related Bcl-2 family proteins
In order to understand the molecular basis of
waltonitone-induced apoptosis, we investigated the expression
levels of Bcl-2 and Bax by western blot analysis. The western blot
analysis showed that waltonitone treatment leads to decrease in
Bcl-2 levels; increase in Bax levels as compared to control cells
(Fig. 5). The ratios of Bax/Bcl-2
increased as the concentration of waltonitone increased.
Waltonitone affects the expression of
miRNAs
We investigated whether the treatment of cells with
waltonitone could affect the expression of miRNAs. Using miRNA
microarray analysis, we found 27 miRNAs with >1.5-fold
expression changes in waltonitone-treated A549 cells (Fig. 6), of which 15 were upregulated, and
12 downregulated. A total of 27 miRNAs with expression levels
regulated by waltonitone treatment (Table I) were selected to identify
potential target genes related to cell proliferation and
apoptosis.
 | Table IThe 27 miRNAs showing 1.5-fold
expression changes after waltonitone exposure. |
Table I
The 27 miRNAs showing 1.5-fold
expression changes after waltonitone exposure.
| miRNA name | Change-fold |
|---|
| Upregulation |
| hsa-miR-1246 | 3.461 |
| hsa-miR-663 | 2.513 |
| hsa-miR-1308 | 2.504 |
| hsa-miR-1225 | 1.844 |
| hsa-miR-21 | 1.764 |
| hsa-miR-346 | 1.739 |
| hsa-miR-29a | 1.728 |
| hsa-miR-720 | 1.624 |
| hsa-miR-1228 | 1.602 |
| hsa-miR-126 | 1.562 |
| hsa-miR-149 | 1.554 |
| hsa-miR-500 | 1.542 |
| hsa-miR-221 | 1.532 |
| hsa-miR-638 | 1.519 |
| hsa-miR-22 | 1.51 |
| Downregulation |
|
hsa-miR-455-3p | 1.501 |
|
hsa-miR-371-5p | 1.524 |
| hsa-miR-10a | 1.526 |
|
hsa-miR-324-3p | 1.536 |
| hsa-miR-18b | 1.538 |
| hsa-miR-186 | 1.543 |
| hsa-let-7c | 1.689 |
|
hsa-miR-886-3p | 1.697 |
|
hsa-miR-516a-5p | 1.745 |
| hsa-miR-411 | 1.754 |
| hsa-miR-18a | 1.919 |
| hsa-miR-324 | 2.481 |
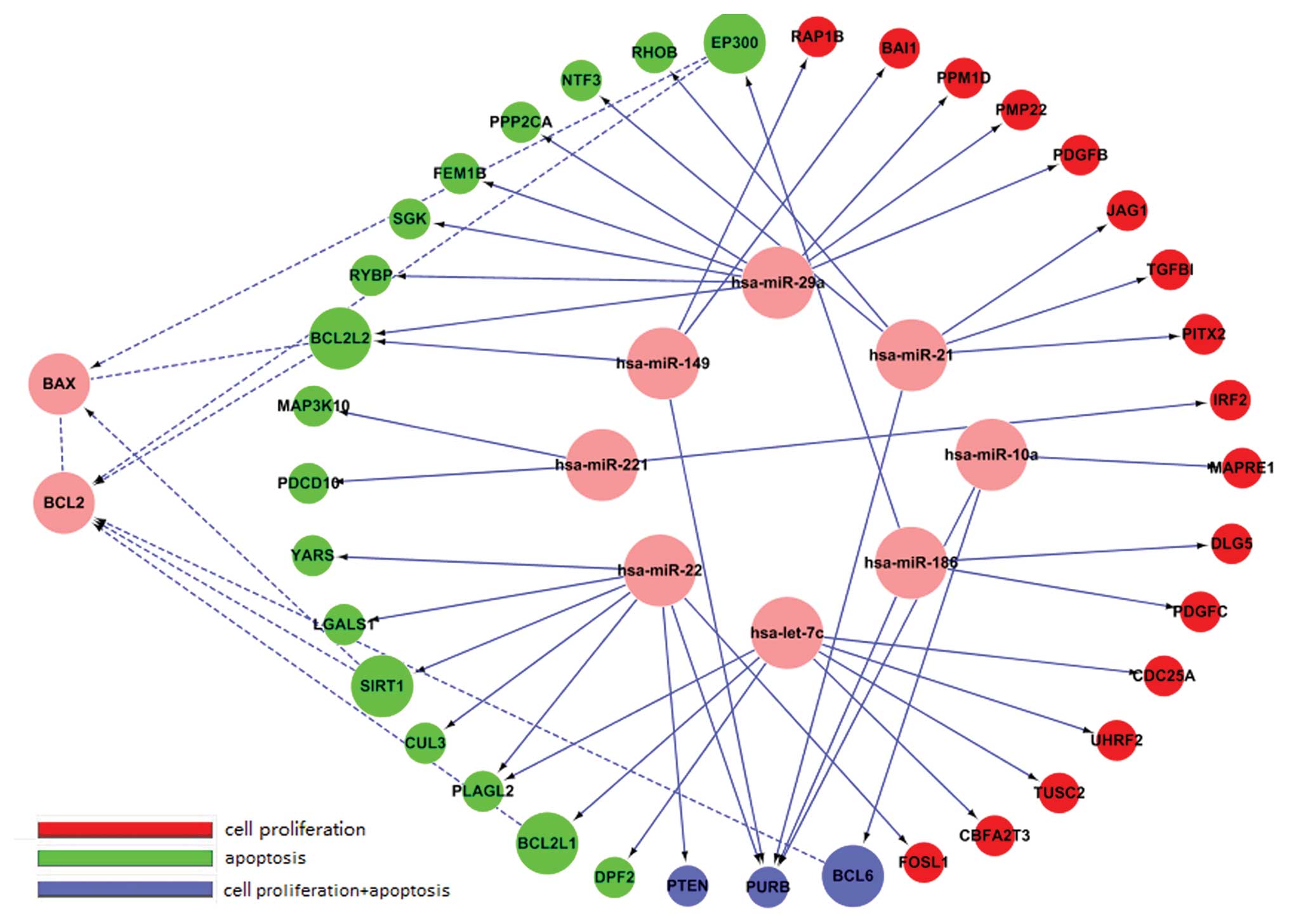
Latent targets of each miRNA were
predicted using miRanda, PicTar and TargetScan
We investigated related targets from the Gene
Ontology database that were separated into two groups: cell
proliferation and apoptosis. We found 26 genes associated with cell
proliferation, and 26 genes associated with apoptosis (Table II) (Fig. 7).
| Table IIClassification and target prediction
of miRNAs. |
Table II
Classification and target prediction
of miRNAs.
| Functions of target
genes |
|---|
|
|
|---|
| miRNA name | Cell
proliferation | Apoptosis |
|---|
| hsa-let-7c | UHRF2, TUSC2,
CDC25A, CBFA2T3 | DPF2, PLAGL2,
BCL2L1 |
| hsa-miR-10a | MAPRE1, BCL6,
PURB | BCL6, PURB |
| hsa-miR-126 | - | - |
| hsa-miR-149 | PURB, BAI1,
RAP1B | PURB, BCL2L2 |
| hsa-miR-186 | PURB, DLG5,
PDGFC | EP300, PURB |
| hsa-miR-21 | JAG1, PURB, TGFBI,
PITX2 | PURB, NTF3,
RHOB |
| hsa-miR-22 | PURB, CUL3, PTEN,
FOSL1 | LGALS1, PURB,
SIRT1, PLAGL2, CUL3, YARS, PTEN |
| hsa-miR-221 | IRF2 | MAP3K10,
PDCD10 |
| hsa-miR-29a | PPM1D, PDGFB,
PMP22, LAMC1 | FEM1B, RYBP,
BCL2L2, SGK, PPP2CA |
Discussion
Lung cancer is the leading cause of cancer-related
deaths in the world. It accounts for 17.6% of cancer death, and its
5-year survival rate is only 8.9–15% (1,2,5). Lung
cancer is generally classified into two histological types, small
cell lung cancer and non-small cell lung cancer. Non-small cell
lung cancer accounts for approximately 85% of the cases and it is
further divided into squamous cell carcinoma, adenocarcinoma, large
cell carcinoma, and others (24,25).
Despite advances in early detection and standard treatment,
non-small cell lung cancer is often diagnosed at an advanced stage
and has a poor prognosis (26).
Chemotherapy plays an important role in the treatment of non-small
cell lung cancer, but it is limited to a significant extent by its
toxicities, drug resistance and significant side effects, including
myelosuppression, neutropenia and thrombocytopenia (3,4). One
possible way to increase the efficacy of anticancer drugs and to
decrease toxicities or side effects is to develop natural
compounds, especially from medicinal plants.
Waltonitone, an ursane-type pentacyclic triterpene
isolated from Chinese medical plant Gentiana waltonii
Burkill, has been used for centuries in treatment of patients
with various autoimmune diseases in China. Study has shown that it
inhibits growth of cultured cells and induces cell apoptosis in
cancer cells (23). In this study,
we first demonstrated that A549 cells treated with waltonitone
showed a concentration- and time-dependent inhibition of the
proliferation. Nuclear fragmentation, chromosome condensation and
formation of apoptotic bodies were also observed by fluorescent
microscopy. Flow cytometric analysis revealed that waltonitone
treatment results in an increase of apoptotic cells. We observed
the molecular basis of waltonitone-induced apoptosis by
downregulation of Bcl-2 protein levels and upregulation of Bax
protein expression. Furthermore, the mechanism of
waltonitone-inhibited proliferation involves the regulation of
miRNAs.
The Bcl-2 family regulates the apoptotic pathway and
can be divided into two types: anti-apoptotic proteins and
pro-apoptotic proteins (11–13).
Many anticancer agents extracted from Chinese herbs induce cancer
cell apoptosis targeting the balance of pro- and anti-apoptotic
proteins (27–29). Bcl-2 is an anti-apoptotic protein of
the Bcl-2 family, and Bax is an pro- apoptotic protein. The ratio
of Bax/Bcl-2 is a decisive factor and plays an important role in
determining whether cells will undergo apoptosis. Our results
showed that expression of Bcl-2 was downregulated by waltonitone,
whereas Bax expression was upregulated, leading to upregulation of
the ratio between Bax and Bcl-2. This might contribute to the
apoptosis promotion activity of waltonitone.
miRNAs are small non-coding RNA molecules that
modulate the post-transcriptional regulation of gene expression in
multi-cellular organisms by complementary interaction with the
3′-untranslated regions (3′-UTR) of target mRNA (14,30,31).
miRNA interaction influences protein expression by promoting the
degradation or suppressing mRNA translation (15). The expression of miRNAs has been
recognized as integral components of many biologic processes
including cell proliferation, development, apoptosis and
differentiation (16,17). Recently, many studies have shown
that miRNAs play an important role in the development and
progression of cancers (18,19).
The miRNAs expression patterns could be potential biomarkers used
for diagnosis, prognosis, and personalized therapy in cancer
include non-small cell lung cancer (32–35).
It is likely, therefore, that some anticancer agents suppress the
proliferation of malignant cells by regulation expression of miRNAs
(20–22). While altered expression of miRNAs
has been the important application for discovering effective
targets and mechanisms in treatment of cancer with natural
compound.
To further understand the mechanisms of action of
waltonitone against the growth of human lung cancer cells, we
investigated the effects of waltonitone on the expression of
miRNAs. Using miRNA microarray analysis, we found 27 miRNAs with
>1.5-fold expression changes in waltonitone-treated A549 cells,
15 upregulated and 12 downregulated miRNAs. Furthermore, we
identified potential target genes of these miRNAs related to cell
proliferation and apoptosis. We found 26 genes associated with cell
proliferation and 26 genes associated with apoptosis. Thus,
waltonitone may inhibit proliferation and induce apoptosis of human
lung cancer A549 cells through regulation of expression of
miRNAs.
In conclusion, we demonstrated that waltonitone
inhibited proliferation and promoted apoptosis in lung cancer A549
cells. This apoptotic response is associated with the upregulation
of the ratio of Bax/Bcl-2. Moreover, modulation of miRNA expression
may be an important mechanism underlying the biological roles of
waltonitone. Based on the outcome of this study, we suggest that
waltonitone, at least in part, inhibited proliferation of human
lung cancer A549 cells through regulating expression of miRNAs.
Waltonitone shows potential for development as an agent for the
management of lung cancer.
Acknowledgements
The authors thank Dr Ting-Shu Jiang (Department of
Respiratory Medicine, Shengjing Hospital of China Medical
University) for experimental guidance.
References
|
1
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
2
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar
|
|
3
|
Chau GY, Lui WY, Tsay SH, Chao Y, King KL
and Wu CW: Postresectional adjuvant intraportal chemotherapy in
patients with hepatocellular carcinoma: a case-control study. Ann
Surg Oncol. 13:1329–1337. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ono T, Yamanoi A, Nazmy El Assal O, Kohno
H and Nagasue N: Adjuvant chemotherapy after resection of
hepatocellular carcinoma causes deterioration of long-term
prognosis in cirrhotic patients: metaanalysis of three randomized
controlled trials. Cancer. 91:2378–2385. 2001. View Article : Google Scholar
|
|
5
|
Erridge SC, Moller H, Price A and Brewster
D: International comparisons of survival from lung cancer: pitfalls
and warnings. Nat Clin Pract Oncol. 4:570–577. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Reyes-Zurita FJ, Rufino-Palomares EE,
Lupianez JA and Cascante M: Maslinic acid, a natural triterpene
from Olea europaea L, induces apoptosis in HT29 human
colon-cancer cells via the mitochondrial apoptotic pathway. Cancer
Lett. 273:44–54. 2009.PubMed/NCBI
|
|
7
|
Moghaddam SJ, Barta P, Mirabolfathinejad
SG, Ammar-Aouchiche Z, Garza NT, Vo TT, Newman RA, Aggarwal BB,
Evans CM, Tuvim MJ, et al: Curcumin inhibits COPD-like airway
inflammation and lung cancer progression in mice. Carcinogenesis.
30:1949–1956. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Harikumar KB, Kunnumakkara AB, Sethi G,
Diagaradjane P, Anand P, Pandey MK, Gelovani J, Krishnan S, Guha S
and Aggarwal BB: Resveratrol, a multitargeted agent, can enhance
antitumor activity of gemcitabine in vitro and in orthotopic mouse
model of human pancreatic cancer. Int J Cancer. 127:257–268.
2010.PubMed/NCBI
|
|
9
|
Sukhthankar M, Yamaguchi K, Lee SH,
McEntee MF, Eling TE, Hara Y and Baek SJ: A green tea component
suppresses posttranslational expression of basic fibroblast growth
factor in colorectal cancer. Gastroenterology. 134:1972–1980. 2008.
View Article : Google Scholar
|
|
10
|
Xavier CP, Lima CF, Preto A, Seruca R,
Fernandes-Ferreira M and Pereira-Wilson C: Luteolin, quercetin and
ursolic acid are potent inhibitors of proliferation and inducers of
apoptosis in both KRAS and BRAF mutated human colorectal cancer
cells. Cancer Lett. 281:162–170. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Burlacu A: Regulation of apoptosis by
Bcl-2 family proteins. J Cell Mol Med. 7:249–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Metrailler-Ruchonnet I, Pagano A,
Carnesecchi S, Ody C, Donati Y and Barazzone Argiroffo C: Bcl-2
protects against hyperoxia-induced apoptosis through inhibition of
the mitochondria-dependent pathway. Free Radic Biol Med.
42:1062–1074. 2007. View Article : Google Scholar
|
|
13
|
Yao Y, Huang C, Li ZF, Wang AY, Liu LY,
Zhao XG, Luo Y, Ni L, Zhang WG and Song TS: Exogenous
phosphatidylethanolamine induces apoptosis of human hepatoma HepG2
cells via the bcl-2/Bax pathway. World J Gastroenterol.
15:1751–1758. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pillai RS, Bhattacharyya SN and Filipowicz
W: Repression of protein synthesis by miRNAs: how many mechanisms?
Trends Cell Biol. 17:118–126. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ambros V and Chen X: The regulation of
genes and genomes by small RNAs. Development. 134:1635–1641. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ambros V: MicroRNA pathways in flies and
worms: growth, death, fat, stress, and timing. Cell. 113:673–676.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Calin GA, Dumitru CD, Shimizu M, Bichi R,
Zupo S, Noch E, Aldler H, Rattan S, Keating M, Rai K, et al:
Frequent deletions and down-regulation of micro-RNA genes miR15 and
miR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci
USA. 99:15524–15529. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen CZ: MicroRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang D, Qiu C, Zhang H, Wang J, Cui Q and
Yin Y: Human microRNA oncogenes and tumor suppressors show
significantly different biological patterns: from functions to
targets. PLoS One. 5:e130672010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Rossi L, Bonmassar E and Faraoni I:
Modification of miR gene expression pattern in human colon cancer
cells following exposure to 5-fluorouracil in vitro. Pharmacol Res.
56:248–253. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, VandenBoom TG II, Kong D, Wang Z,
Ali S, Philip PA and Sarkar FH: Up-regulation of miR-200 and let-7
by natural agents leads to the reversal of
epithelial-to-mesenchymal transition in gemcitabine-resistant
pancreatic cancer cells. Cancer Res. 69:6704–6712. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tili E, Michaille JJ, Adair B, Alder H,
Limagne E, Taccioli C, Ferracin M, Delmas D, Latruffe N and Croce
CM: Resveratrol decreases the levels of miR-155 by upregulating
miR-663, a microRNA targeting JunB and JunD. Carcinogenesis.
31:1561–1566. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Z, Wang S, Qiu H, Duan C, Ding K and
Wang Z: Waltonitone induces human hepatocellular carcinoma cells
apoptosis in vitro and in vivo. Cancer Lett. 286:223–231. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Brambilla E, Travis WD, Colby TV, Corrin B
and Shimosato Y: The new World Health Organization classification
of lung tumours. Eur Respir J. 18:1059–1068. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wistuba II and Gazdar AF: Lung cancer
preneoplasia. Annu Rev Pathol. 1:331–348. 2006. View Article : Google Scholar
|
|
26
|
Herbst RS, Heymach JV and Lippman SM: Lung
cancer. N Engl J Med. 359:1367–1380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pongrakhananon V, Nimmannit U, Luanpitpong
S, Rojanasakul Y and Chanvorachote P: Curcumin sensitizes non-small
cell lung cancer cell anoikis through reactive oxygen
species-mediated Bcl-2 downregulation. Apoptosis. 15:574–585. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jiang T, Zhou L, Zhang W, Qu D, Xu X, Yang
Y and Li S: Effects of sinomenine on proliferation and apoptosis in
human lung cancer cell line NCI-H460 in vitro. Mol Med Rep.
3:51–56. 2010.PubMed/NCBI
|
|
29
|
Ren G, Zhao YP, Yang L and Fu CX:
Anti-proliferative effect of clitocine from the mushroom
Leucopaxillus giganteus on human cervical cancer HeLa cells
by inducing apoptosis. Cancer Lett. 262:190–200. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luthra R, Singh RR, Luthra MG, Li YX,
Hannah C, Romans AM, Barkoh BA, Chen SS, Ensor J, Maru DM, et al:
MicroRNA-196a targets annexin A1: a microRNA-mediated mechanism of
annexin A1 downregulation in cancers. Oncogene. 27:6667–6678. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Raver-Shapira N, Marciano E, Meiri E,
Spector Y, Rosenfeld N, Moskovits N, Bentwich Z and Oren M:
Transcriptional activation of miR-34a contributes to p53-mediated
apoptosis. Mol Cell. 26:731–743. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yanaihara N, Caplen N, Bowman E, Seike M,
Kumamoto K, Yi M, Stephens RM, Okamoto A, Yokota J, et al: Unique
microRNA molecular profiles in lung cancer diagnosis and prognosis.
Cancer Cell. 9:189–198. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu SL, Chen HY, Chang GC, Chen CY, Chen
HW, Singh S, Cheng CL, Yu CJ, Lee YC, Chen HS, et al: MicroRNA
signature predicts survival and relapse in lung cancer. Cancer
Cell. 13:48–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ventura A and Jacks T: MicroRNAs and
cancer: short RNAs go a long way. Cell. 136:586–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Raponi M, Dossey L, Jatkoe T, Wu X, Chen
G, Fan H and Beer DG: MicroRNA classifiers for predicting prognosis
of squamous cell lung cancer. Cancer Res. 69:5776–5783. 2009.
View Article : Google Scholar : PubMed/NCBI
|