Introduction
Prostate cancer is the second most frequently
diagnosed cancer and the sixth leading cause of cancer death in
males worldwide (1,2). Due to its high incidence and
mortality, early diagnosis, identification of highly aggressive
clinically silent forms and understanding of disease pathogenesis
with typical metabolic differences in order to develop specifically
targeted therapy are needed. To target these issues, biochemistry
of normal and tumour prostate cells is investigated. Based on these
investigations, several typical characteristics of prostate tissue
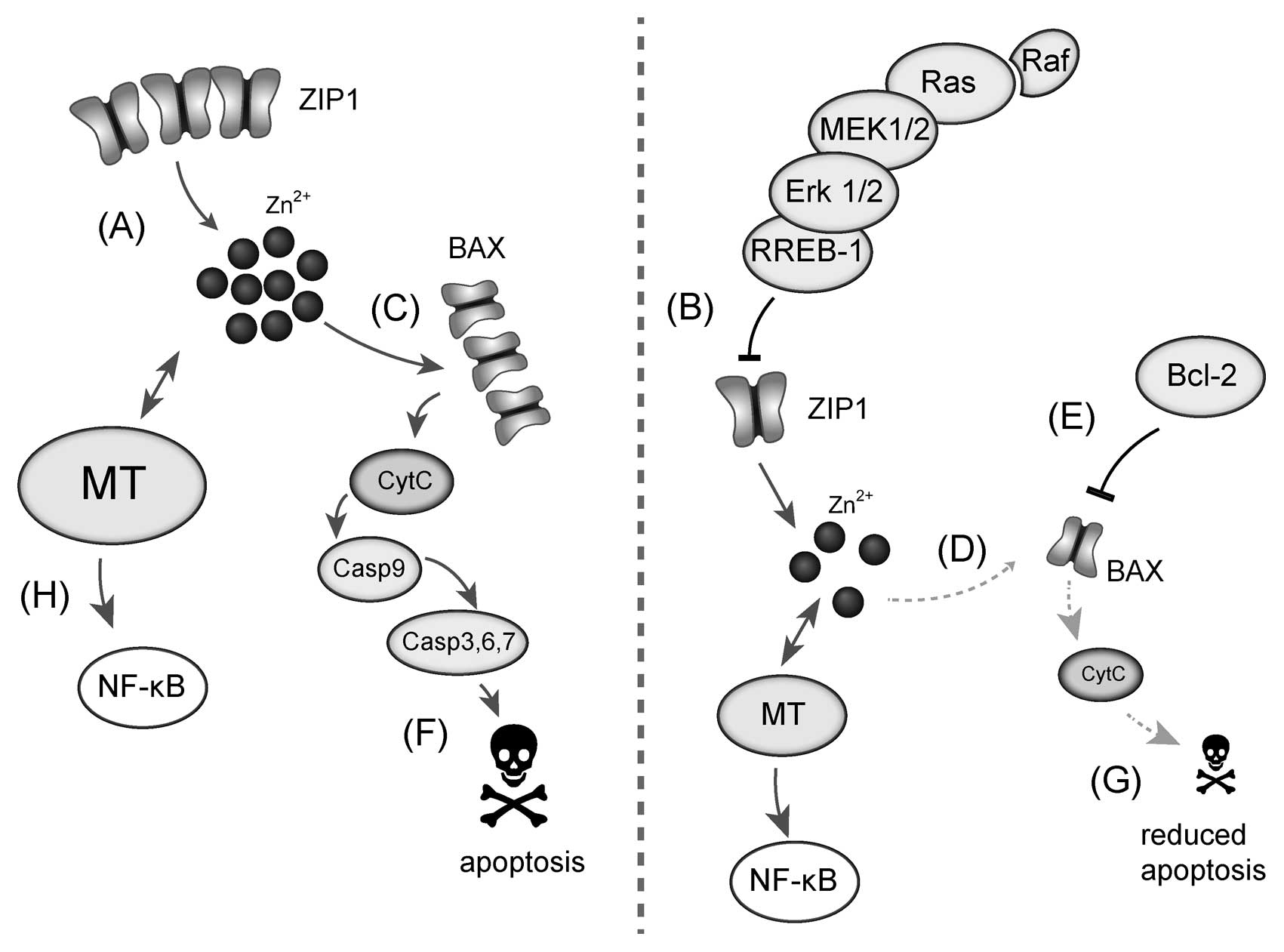
have been found including the ability to accumulate zinc(II) ions
(3-7), which are shown in Fig. 1. The intracellular concentration of
zinc(II) ions in prostate tissue exceeds up to ten times the
concentrations detected in other cell and tissue types. However,
this feature of prostate cell lines is lost during carcinogenesis
and, thus, prostate cells are unable to accumulate zinc(II) ions in
high levels. Therefore, we can expect that zinc(II) ions can
significantly contribute to the progression of tumour disease and
to the ability of prostate cell lines to metastasize (8).
As a result of numerous studies on cells as well as
on prostate cancer patients, several compounds have been found
connected with tumourigenesis in prostate cells including Bcl-2.
This intracellular protein belongs to a large group of proteins the
Bcl-2 family (9), and acts as an
inhibitor of apoptosis. Bcl-2 has been established to block
apoptotic death in various cell types such as lymphocytes and
motoric neurons. It prevents both apoptosis dependent on caspases
and oxidative necrosis. Under normal conditions, Bcl-2 is anchored
to the outer mitochondrial membrane and heads out into the cytosol,
which gives this protein the opportunity to interact with other
proteins. These interactions are important for maintaining
mitochondrial membrane integrity and function. By binding to the
pro-apoptotic family members, Bcl-2 prevents activation of
mitochondrial pathway of apoptosis based on the formation of pores
that disrupt the permeability of mitochondrial membrane (10-12).
It has been suggested that overexpression of the Bcl-2 oncoprotein
in human cancer cells contributes to their resistance to
chemotherapy- and radiotherapy-induced apoptosis and is connected
with unfavourable prognosis (13).
The majority of human prostate tumours overexpress Bcl-2, which is
responsible for tumour resistance to radiotherapy and chemotherapy
(14,15). This event is supported by the fact
that Bcl-2 knocked down by antisense oligodeoxynucleotides induces
radiosensitization in human PC-3 prostate tumour xenografts
(13). Moreover, it has been
reported that Bcl-2 expression is associated with tumour
progression and unfavourable prognosis in prostate cancer patients
(13,16,17)
and is associated with the development of androgen-independent
prostate cancer (18). Possible
associations with other proteins connected with tumour processes
such as c-Fos, c-Jun, Ki-67, NF-κB and p53 can be expected.
Therefore, we aimed our attention at determining of expression of
Bcl-2, c-Fos, c-Jun, Ki-67, NF-κB and p53 genes in two prostate
cell lines the 22Rv1 cell line, a model of aggressive partially
androgen-sensitive prostate cancer and the PNT1A cell line, a model
of healthy cell line. Moreover, we were interested in the issue how
exposure of these cell lines to zinc(II) ions could influence
expression of the above-mentioned genes. The expression levels were
correlated with the results obtained with a fluorescence microscopy
of treated cells, and zinc and -SH moieties content.
Materials and methods
Chemical and biochemical reagents
RPMI-1640 medium, Ham’s F12 medium, fetal bovine
serum (FBS) (mycoplasma-free), penicillin/streptomycin and trypsin
were purchased from PAA Laboratories GmbH (Pasching, Austria). PBS
was purchased from Invitrogen Corp. (Carlsbad, CA, USA).
Ethylenediaminetetraacetic acid (EDTA), zinc(II) sulphate
(BioReagent grade, suitable for cell cultures), RIPA buffer and all
other chemicals of ACS purity were purchased from Sigma-Aldrich Co.
(St. Louis, MO, USA), unless noted otherwise.
Cell cultures
Two human prostatic cell lines were used in this
study: a) PNT1A human cell line established by immortalisation of
normal adult prostatic epithelial cells by transfection with a
plasmid containing SV40 genome with a defective replication origin;
the primary culture was obtained from the normal prostatic tissue
of a 35-year old male at post mortem, and b) 22Rv1 human
cell line derived from a xenograft that was serially propagated in
mice after castration. Both cell lines used in this study were
purchased from HPA Culture Collections (Salisbury, UK).
Cell cultivation
PNT1A cells were cultured in RPMI-1640 medium
supplemented by 10% FBS. 22Rv1 cells were cultured in RPMI-1640
without phenol red (medium) with 10% FBS. The media were
supplemented with penicillin (100 U/ml) and streptomycin (0.1
mg/ml), and the cells were maintained at 37°C in a humidified (60%)
incubator with 5% CO2 (Sanyo, Japan). The passages of
PNT1A and 22Rv1 cell lines ranged from 10 to 35 h.
Zinc(II) treatments of cell cultures
Immediately the cells grew up to 50-60% confluence,
the cultivation media were replaced by a fresh medium to
synchronize cell growth. Cells were cultivated under these
conditions for 24 h, then cells were treated with zinc(II) sulphate
(0-100 μM for both cell lines) dissolved in fresh medium for 48
h.
RNA isolation, cDNA preparation
High pure total RNA isolation kit (Roche, Basel,
Switzerland) was used for RNA isolation. Briefly, cultivation
medium was removed and samples were washed twice with 5 ml of
ice-cold PBS. Cells were transferred to clean tubes and centrifuged
at 15,000 × g for 5 min at 4°C. After it, lysis buffer was added
and RNA isolation was carried out according to the manufacturer’s
instructions. Isolated RNA was used for cDNA construction. Total
RNA (600 ng) was transcribed using transcriptor first strand cDNA
synthesis kit (Roche). Prepared cDNA (20 μl) was diluted with
RNase-free water to 100 μl and directly analysed by real-time
polymerase chain reaction.
Real-time reverse-transcription
polymerase chain reaction (RT-PCR)
RT-PCR was performed in triplicates using the TaqMan
gene expression assay with 7500 real-time PCR system (Applied
Biosystems, Foster City, CA, USA). The amplified DNA was analysed
by the comparative Ct method using β-actin as an endogenous
control. The primer and probe sets for β-actin (assay ID:
Hs00185826_m1), Fos (assay ID: Hs00170630_m1), c-Jun (assay ID:
Hs00277190_m1), NF-κB (assay ID: Hs00765730_m1), p53 (assay ID:
Hs01034649_m1), Bcl-2 (assay ID: Hs99999018_m1) were selected from
TaqMan gene expression assay. Real-time PCR was performed under
following amplification conditions: total volume of 20 μl, initial
denaturation 95°C/10 min, then 40 cycles 95°C/15 sec, 60°C/1
min.
Cell content quantification
Total cell content was analysed using semi-automated
image-based cell analyser (Cedex XS, Innovatis, Roche, Basel,
Switzerland) according to the following protocol. Cultivation
medium was removed and samples were two times washed with 5 ml of
ice-cold PBS to maintain only viable cells. Cells were scraped and
transferred to clean tubes. Trypan blue solution (Innovatis) was
diluted to 0.2% prior to use and added to samples. Following
settings were used in operating software: cell type: standard
cells, dilution: none, process type: standard. All samples were
measured in duplicates.
Measurements of cell viability - MTT
test
MTT assay was used to determine cell viability. The
suspension of cells was diluted to the density of 5,000 cells/ml in
the cultivation medium. Volume of 200 μl was transferred to 2-11
wells of standard microtiter plates. Medium (200 μl) was added to
the first and to the last column (1 and 12, control). Plates were
incubated for 2 days at 37°C to ensure cell growth under the same
conditions described in Cell cultivation. Medium was removed from
columns 2 to 11. Columns 3-10 were filled with 200 μl of medium
containing increasing concentration of zinc(II) (0, 50, 100, 150,
200, 250, 300 and 500 μM). As control, columns 2 and 11 were not
filled with medium containing zinc(II) ions. Plates were incubated
for 24 h; then, media were removed and replaced by a fresh medium,
three times a day. Columns 1 to 11 were filled with 200 μl of
medium containing 50 μl of MTT (5 mg/ml in PBS) and incubated in a
humidified atmosphere for 4 h at 37°C, wrapped in aluminium foil.
After the incubation, MTT-containing medium was replaced by 200 μl
of 99.9% dimethyl sulphoxide (DMSO) to dissolve MTT-formazan
crystals. Then, 25 μl of glycine buffer was added to all wells and
absorbance was immediately determined at 570 nm (VersaMax
microplate reader, Molecular Devices, Sunnyvale, CA, USA).
Cell growth and proliferation assay using
impedance measurement with xCELLigence system
The xCELLigence system (Roche Applied Science and
ACEA Biosciences, San Diego, CA, USA) consists of four main
components: the RTCA analyser, the RTCA DP station, the RTCA
computer with integrated software and disposable E-plate 16.
Firstly, the optimal seeding concentration for proliferation and
cytotoxic assay was determined. After seeding the total number of
cells in 200 μl medium to each well in E-plate 16, the attachment,
proliferation and spreading of the cells was monitored every 15
min. All experiments were carried out for 250 h. The results are
expressed as relative impedance using the manufacturer’s software
(Roche Applied Science and ACEA Biosciences).
Densitometric and statistical
analysis
Software Statistica 10 (StatSoft Inc., Tulsa, OK,
USA) was used for statistical analysis. Student’s t-test for
independent values was used to evaluate differences between two
groups. Simple linear correlations were performed to reveal the
relationships between variables. Unless noted otherwise, a level of
statistical significance was set at p<0.05.
Fluorescence microscopy and cell
staining
For fluorescence microscopy, cells were cultivated
directly on microscope glass slides (75×25 mm, thickness 1 mm,
Fischer Scientific, Pardubice, Czech Republic) in Petri dishes in
above-described cultivation media (see Cultured cell conditions).
Cells were transferred directly onto slides, which were submerged
in cultivation media. After treatment, microscope glass slides with
monolayer of cells were removed from Petri dishes, rinsed with
cultivation medium without zinc(II) supplementation and PBS buffer
and directly used for staining and fluorescence microscopy.
For the staining of free thiols, respectively free
-SH groups, 5-(bromomethyl)fluorescein (5-BMF, Sigma-Aldrich) was
used. This probe reacts more slowly with thiols of peptides,
proteins and thiolated nucleic acids in comparison with other
fluorescent probes. However, it forms stronger thioether bonds that
are expected to remain stable under the conditions required for
fluorescence microscopy. Stock solution of 5-BMF (4 mM, anhydrous
DMSO) was prepared prior to staining because of 5-BMF stability.
Working solution was prepared immediately using stock solution by
diluting to final concentration of 20 μM (PBS buffer, pH 7.6).
Cells were incubated for 1 h at 37°C and in dark. Then, cells on
microscope glass slide were three times washed by PBS buffer (pH
7.6) and observed using fluorescence microscope (Axioskop 40, Carl
Zeiss AG, Oberkochen, Germany) equipped with wideband excitation
and set of filters (FITC, DAPI, Carl Zeiss). Photographs were taken
using digital camera (Olympus Camedia 750, Olympus, Tokyo, Japan).
Program NIS-elements was used for evaluation of intensity of
emission, all values were recalculated to control (100%). Ten
random fields from each variant and replicate were evaluated.
For free zinc(II) ion staining, fluorescent probe
N-(6-methoxy-8-quinolyl)-p-toluene sulphonamide (TSQ, Invitrogen)
was used. Working solution (10 μM, phosphate buffer pH 7.6) was
prepared by diluting of TSQ stock solution (10 mM, acetone). Cells
were carefully rinsed by PBS buffer to remove all cultivation
medium containing free zinc(II) ions, subsequently stained by
working TSQ solution (30 min, 37°C, dark), three times washed by
PBS buffer (pH 7.6) and observed under a fluorescence microscope
(Axioskop 40, Carl Zeiss) equipped by FITC and DAPI filters (Carl
Zeiss). Photographs were taken on digital camera (Olympus Camedia
750, Olympus). NIS-element program was used for evaluation of
intensity of emission, all values were recalculated to control
(=100%). Ten random fields from each variant and replicate were
evaluated.
Results
Growth and viability of treated PNT1A and
22Rv1 cells
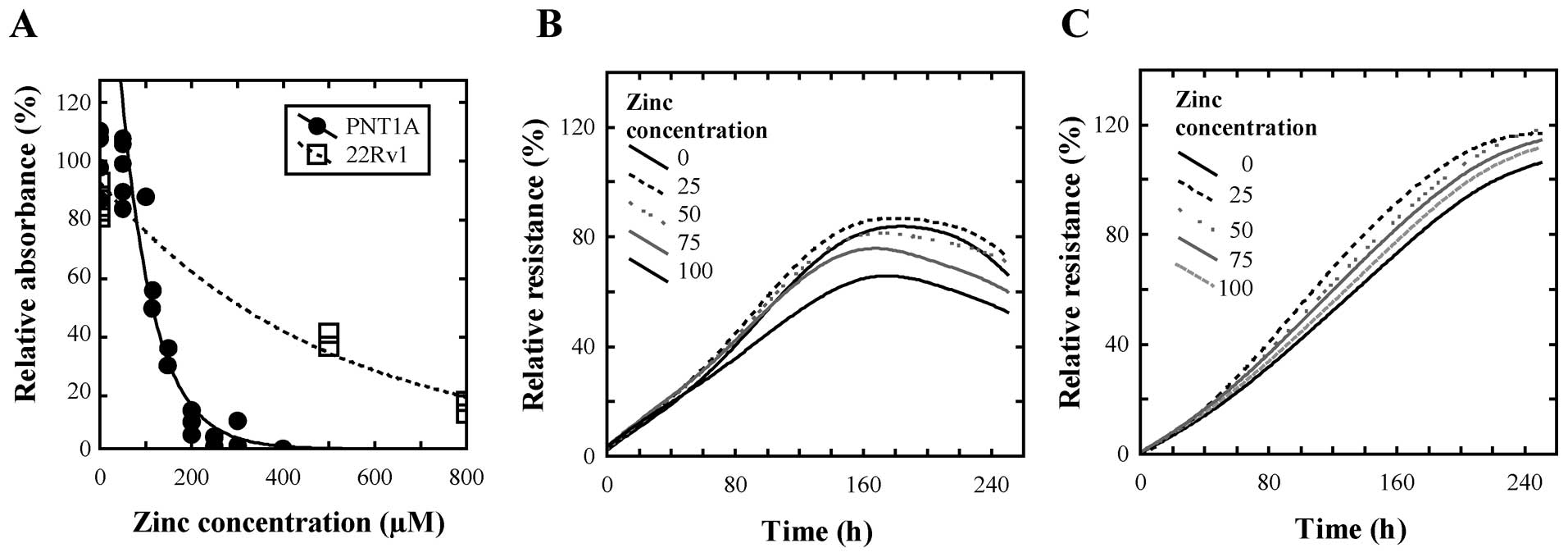
To select suitable concentration range of zinc(II)
ions for treatment of the cell lines, we determined IC50
values of zinc(II) for both cell lines using standard MTT
cytotoxicity assay with simple linear regression from the
descending part of the curve. We obtained IC50
corresponding to concentration of 197.9 μM for non-tumour PNT1A and
concentration of 369.1 μM for 22Rv1 tumour cells (Fig. 2A). Moreover, we found that PNT1A
cells reached stationary phase of growth after 160 h while 22Rv1
cells after 240 h. Thus, we decided to use zinc(II) ion
concentrations as follows: 0 (control), 25, 50, 75 and 100 μM for
both cell lines for the monitoring of changes in gene expression
for 240 h.
Further, we were interested to monitor growth of the
cell lines in real time. It is difficult to monitor on-line growth
of cell lines during experiment due to the destructive methods for
its determination. xCELLigence system offers good possibility to
monitor cell growth and proliferation in real time using impedance
measurement (19), therefore, we
used this system. Primarily, it was necessary to determine optimal
cell counts for real time monitoring. We obtained optimal signal
with 10,000 cells in each well. This signal corresponded to the
cell count. In wells with lower cell count, lower relative
impedance signal level was determined and, in some cases, higher
relative standard deviation was obtained. Higher cell counts gave
also well repeatable results with low relative standard deviation,
however, higher count of cells was not optimal from the point of
view of 240 h long experiment expecting increasing proliferation
and therefore cell count. Therefore, we decided to use 10,000 cells
per well to examine the effect of zinc(II) ions on prostatic cells
either derived from normal prostate epithelium (PNT1A) or cells
derived from primary prostate carcinoma (22Rv1). The
IC50 values of zinc(II) ions using the xCELLigence
system were also determined to evaluate the selected concentration
range. These IC50 values were as follows: 150.8 μM zinc
for PNT1A cells and 445.5 μM zinc for 22Rv1 cells. The changes
between these values and values determined using MTT assay can be
associated with the fact that both methods are based on measuring
of very different physical parameters, which are associated with
different physiological phenomena. Based on both measurements of
IC50 it may be concluded that zinc(II) ions are more
toxic to non-tumour cell line at 2.9-fold lower concentration.
Based on the obtained results 10,000 cells per well were treated
with the above-mentioned concentrations of zinc(II) ions for 240 h.
In the beginning of the treatment we observed negligible decrease
in relative impedance, i.e. in growth, of PNT1A cells after 15-h
long treatment (Fig. 2B) and no
changes in cell proliferation of 22Rv1 cells (Fig. 2C). These changes were compared with
cell lines treated with 0 μM of zinc(II) ions. Concentrations of
100 μM of zinc(II) ions induced 1.4-and 1.1-fold decrease of
relative resistance in PNT1A and 22Rv1 cell lines, respectively,
compared to non-treated samples. These results clearly show that
the selected concentration range is non-toxic for both cell lines
and we can observe the effects of physiological doses of zinc(II)
ions on expression of the selected markers as Bcl-2, c-Fos, c-Jun,
NF-κB, Ki-67 and p53.
Comparison of the expression of Bcl-2 and
other regulatory genes after zinc treatment
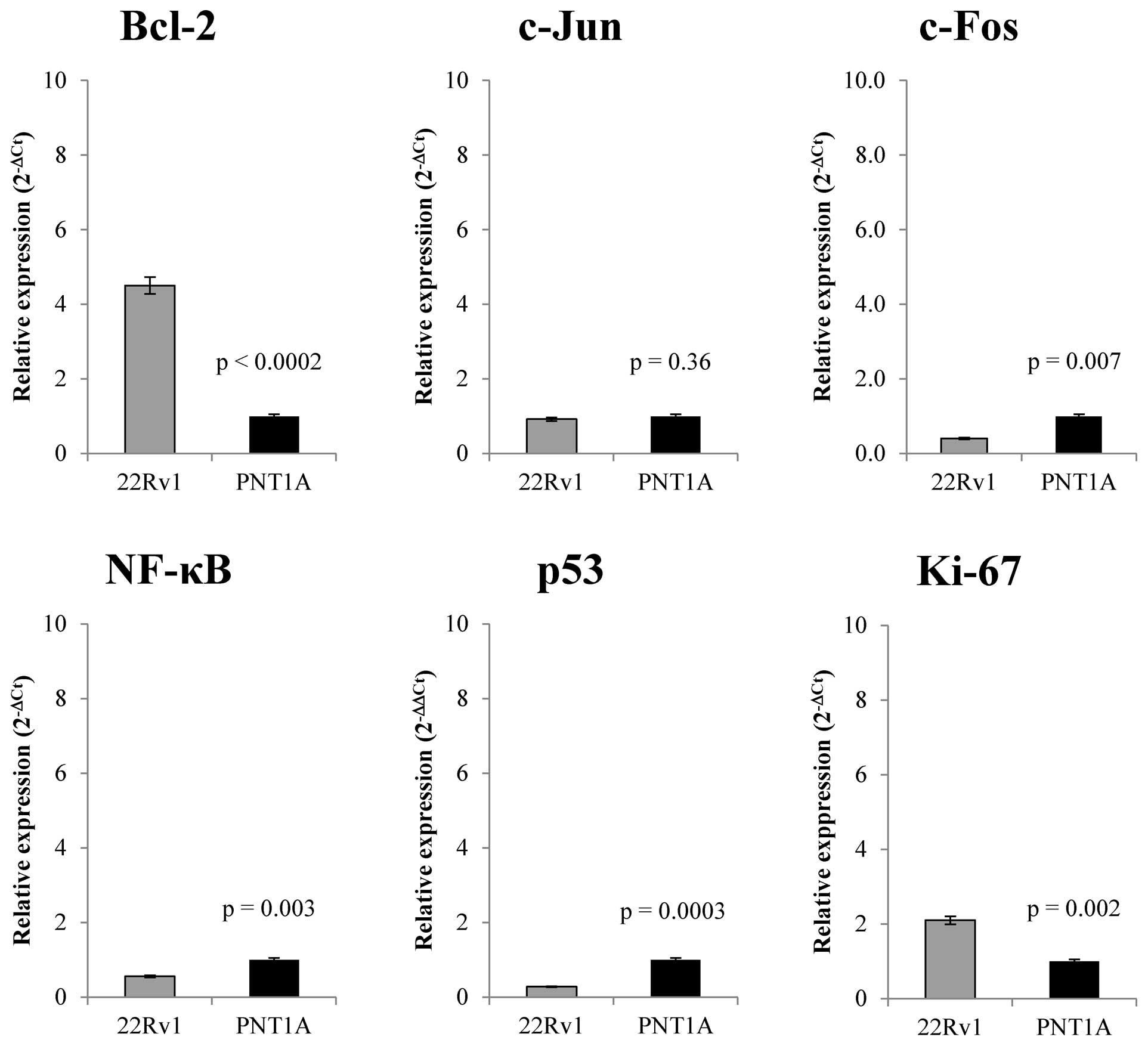
Further, we focused on comparison of the base line
expression and changes in transcription of Bcl-2, c-Fos, c-Jun,
NF-κB, Ki-67 and p53 genes on the RNA level in prostate cell lines
treated with zinc(II) ions. Baseline transcription level and
zinc(II) ions effect on transcription levels of selected genes was
performed by RT-PCR. Fig. 3 shows
that 22Rv1 cells demonstrate different expression patterns in
monitored genes compared to PNT1A cells. 22Rv1 cell line has
4.5-fold higher level of Bcl-2 anti-apoptotic gene expression
(n=5). We found no significant differences (p>0.05) in c-Jun
gene expression in either cell line. c-Fos gene that together with
c-Jun forms important part of AP-1 transcription factor shows
2.5-fold down-regulation in 22Rv1 cells. Ki-67, a nuclear protein
that is associated with cellular proliferation, is present in 22Rv1
cell line in 2-fold higher concentration compared to PNT1A.
Moreover, NF-κB is present in half concentration in 22Rv1cells
compared to PNT1A and p53, a key regulator of apoptosis, shows
3.3-fold decreased level compared with PNT1A cell line. After basic
characterization of the expression of selected markers in both
studied cell lines, we focused on zinc(II) ion treatment effect on
the expression changes of these selected regulatory genes (Fig. 4).
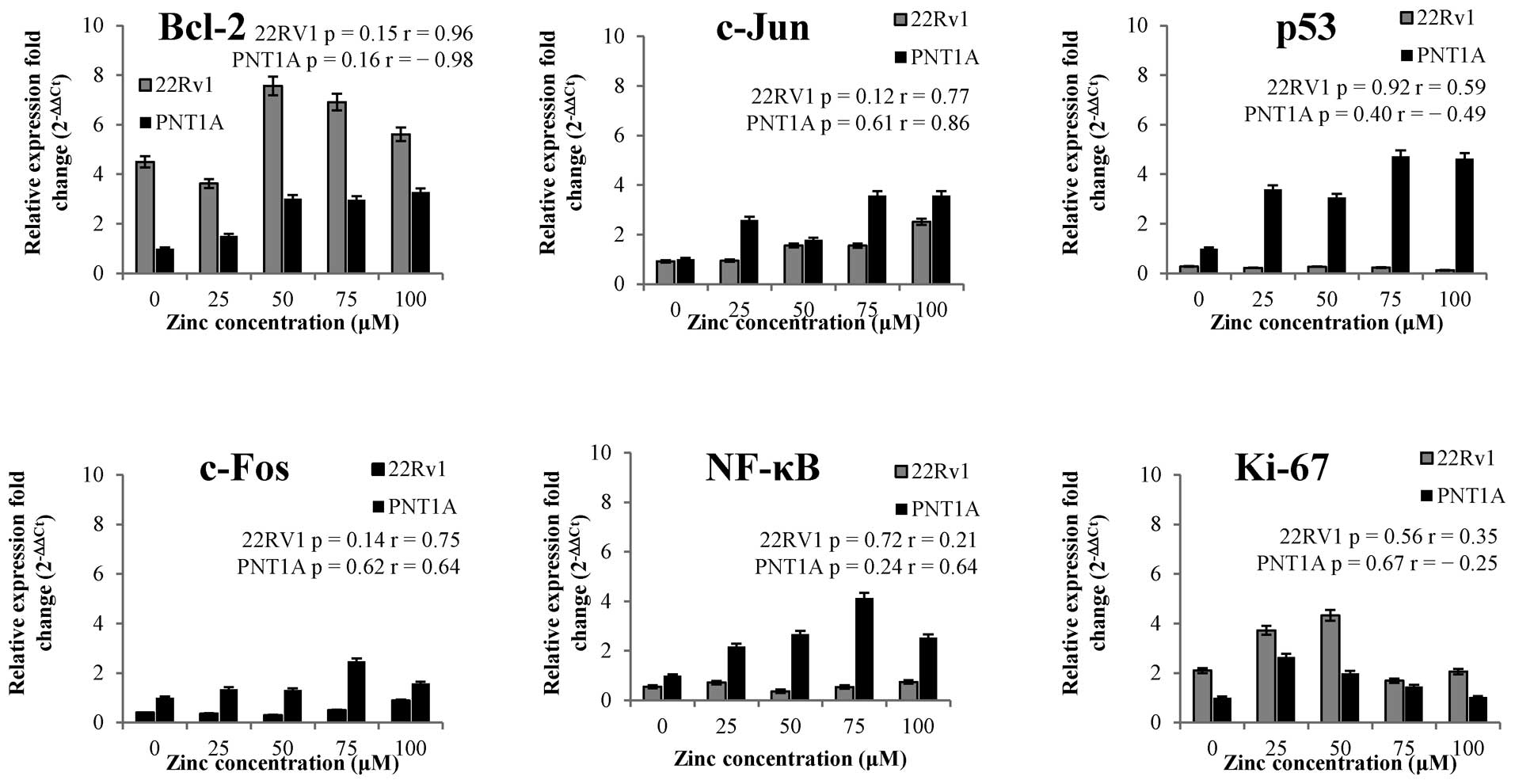
We found that zinc(II) ions influence positively
expression of Bcl-2 gene in both tested cell lines, however, more
in PNT1A cells [≤3.2-fold change in case of 100 μM zinc(II) ions
treatment]. In addition, we found no significant difference
(p>0.05) in p53 expression levels after zinc treatment in 22Rv1
cell line (Fig. 4). On the other
hand, zinc(II) ions treatment resulted in the increasing p53
expression from 3- up to 4.7-fold change in PNT1A. Ki-67 gene shows
similar pattern in both cell lines after zinc(II) treatment and it
is not surprising that 22Rv1 cell line, characterized by higher
proliferation rate (Fig. 2A and B),
demonstrated higher expression as a gene involved in the
proliferation process. Zinc(II) ions had up-regulative effect on
c-Jun gene in both cell lines. PNT1A demonstrated higher c-Jun
expression in all zinc treatments compared to 22Rv1. In addition,
we found no correlation between c-Jun and c-Fos gene expression
after zinc treatment, but expression profile of c-Fos showed
statistically significant positive correlation (data not shown)
with expression of NF-κB gene after the treatment (compare trends
in Fig. 4). Both genes are
expressed in higher concentration in PNT1A cell line on the base
expression level as well as after zinc treatment.
Correlation analysis of selected
genes
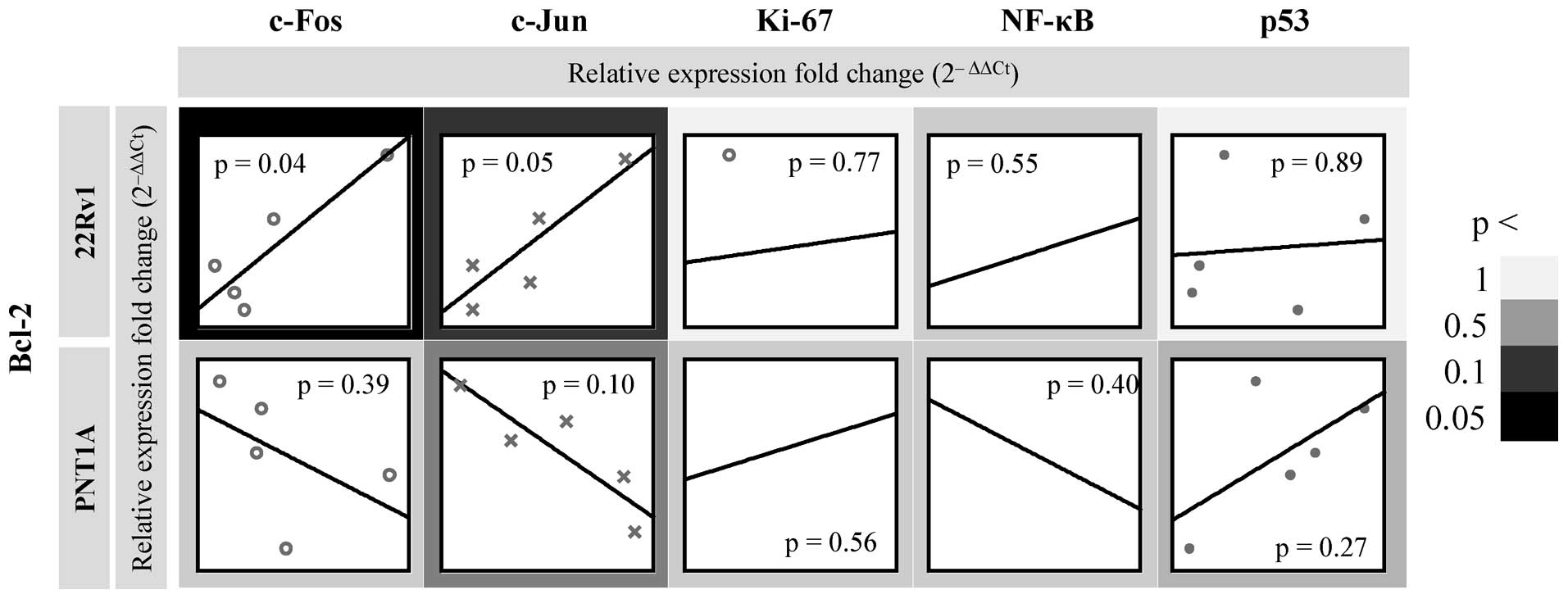
Subsequently, we focused on possible dependencies
between Bcl-2 mRNA and the mRNA levels of other regulatory genes.
After exposure to zinc(II) ions, we observed distinct trends
between Bcl-2 and c-Fos and c-Jun and no significant correlations
between Bcl-2 and Ki-67, NF-κB and p53 in either cell line
(Fig. 5). Bcl-2-p53 showed strong
positive correlation at r=0.61. In the case of c-Fos and c-Jun, we
found strong positive correlations at r>0.85 in 22Rv1 tumour
cell line and strong negative correlations at r<-0.50 in PNT1A
non-tumour cell line. In terms of statistical significance,
Bcl-2-c-Fos and - c-Jun showed correlations p<0.05 only in 22Rv1
cell line. In contrast, non-tumour cell line PNT1A showed an
inverse trend i.e. decrease of Bcl-2 in relation to regulatory
genes. In this cell line, negative significant correlation was
detected at Bcl-2-c-Jun at p=0.10 with r=-0.79 (Fig. 5).
Determination of free and bound zinc(II)
cellular levels after zinc(II) exposure
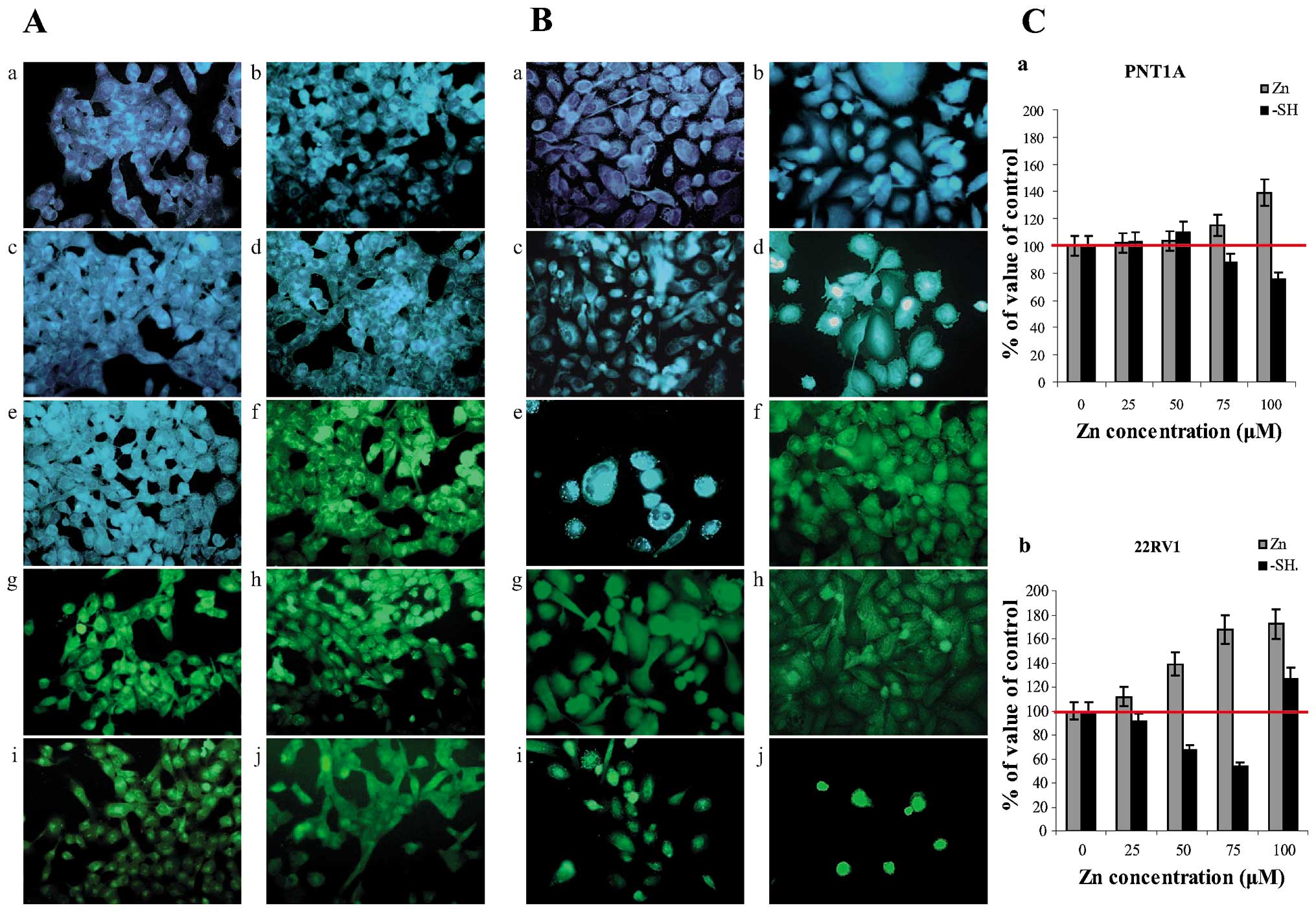
We characterized cell lines from the point of view
of pro- and anti-apoptotic factor expressions, free zinc(II) ions
were visualized using fluorescent probe
N-(6-methoxy-8-quinolyl)-p-toluene sulphonamide, probe
specific to these ions. Significant differences in zinc(II) ion
localization in both non-tumour PNT1A cells (Fig. 6Aa-e) and tumour 22Rv1 (Fig. 6Ba-e) cells were found. For
quantification of changes in free zinc(II) ion levels in cell
lines, program NIS-elements for image analysis was used. Detected
values (10 fields for each concentration and repetition) were
recalculated to control cells (100%) (Fig. 6C). In both PNT1A and 22Rv1 cells,
free zinc(II) ions levels were closely connected with zinc(II) ions
treatment in concentration-dependent manner. In the case of PNT1A
cell line, localization of zinc(II) ions around nuclei and
irregularly in nuclei was evident in cells treated with the highest
zinc(II) concentration (100 μM). Peripheral parts of cytoplasm
demonstrate only weak emission, representing only low free zinc(II)
levels in these localizations. In 22Rv1 cells, the intensity of
emission of fluorescence product significantly increases with the
increased supplementation of cultivation medium by zinc(II) ions
(Fig. 6Ba-e). In the lowest Zn(II)
supplementation, free zinc(II) ions were localized especially
around the nuclei. At the highest Zn(II) supplementation, free
zinc(II) ions were localized in nuclei and around the nuclei in the
form of spots with high emission. The origin of these spots, which
were visible only in the case of 22Rv1 cells in the highest Zn(II)
ions supplementation, is probably the zincosomes, compartments of
endoplasmic reticulum origin (20).
Determination of free thiols
Compounds rich in -SH moieties including low
molecular mass peptides and proteins as reduced glutathione and
metallothionein (21-25) are responsible for binging metal ions
in intracellular space. Therefore, monitoring of such compounds as
well as detection of free zinc(II) ions can answer questions on
metabolizing of metal ions in a cell. 5-(bromomethyl)fluorescein,
the probe that provides formation of fluorescent product after
reaction with -SH groups of thiols, was used for detection and
cellular compartmentation of free thiols. In the case of both PNT1A
(Fig. 6Af-j) and 22Rv1 cells
(Fig. 6Bf-j), amount of free thiols
continually decreases with the increasing zinc(II) ions
supplementation, however, selection of cells with high content of
free thiols was evident. This fact was evident in 22Rv1 cells
treated with the highest Zn(II) concentration. In both cell lines,
free thiols were localized around the nuclei and in nuclei in cells
treated with lower Zn(II) concentrations. On the other hand,
highest free thiols levels were detected in the nuclei in the case
of tumour 22Rv1 cell line. This fact is evidence on the possible
role of free thiols in transcriptional activity, which is regulated
not only by free thiols, but also by zinc(II) ions whose
concentration was high in nuclei in the same case.
Discussion
Zinc is involved in energetic metabolism (26), proliferation and apoptosis (27-29) in
prostate, therefore it is expectable that zinc may play an
important role in prostate cancer pathogenesis (4,30,31).
Our results revealed that the base line expression of the
anti-apoptic gene Bcl-2 is 4.5-fold higher in 22Rv1 than in PNT1A.
This result is in accordance with the previously published reports,
where elevated Bcl-2 expression in prostate cancer tumours has been
reported (14,16). Furthermore, this elevation was
associated with the development of androgen-independent prostate
cancer (18) and also with
radiotherapy and chemotherapy resistance (13,14,32).
Interestingly, we observed further increase in Bcl-2 expression in
both cell lines after zinc(II) treatment. However, in the case of
22Rv1 this enhancement in expression was higher, up to 7.5-fold,
compared to base line expression of Bcl-2 in PNT1A cell line.
Although the exact mechanism remains unclear, our findings suggest
that higher zinc(II) ion concentrations in prostate may contribute
to prostate tumour therapy resistance associated with elevated
Bcl-2 expression.
c-Jun forms homodimers and heterodimers with c-Fos
and other Jun-related proteins, which together comprise the AP-1
transcription factor that binds TPA response elements (TREs).
Therefore, c-Jun mediates transcriptional regulation in response to
a variety of stimuli, including cytokines, growth factors and
stress (33). Generally, AP-1
controls a number of cellular processes including differentiation,
proliferation, and apoptosis (34).
Although there is considerable evidence that c-Jun activation can
represent a positive step in the events leading towards apoptosis,
there are numerous contrary reports. Possible role of c-Jun in
inhibition of apoptosis and promoting of proliferation/cell
differentiation is reviewed in ref. 35. Growing amount of such evidence
implicates c-Jun in the protection of cells from stress-induced
apoptosis. It was reported that cells expressing the Ser63Ala,
Ser73Ala mutant of c-Jun are not protected against apoptosis
triggered by UV irradiation (36).
As 22Rv1 cells show higher viability and proliferation (Fig. 2) after zinc treatment, we can
conclude that the increasing expression of c-Jun does not have
negative impact on 22Rv1 cells.
NF-κB is a transcription factor, which effect cannot
be easily evaluated. Its expression strongly correlates with c-Fos
transcription in both cell lines used in our experiments. This
finding correlates with known fact that NF-κB enhances c-Fos
transcription via the direct binding to a response element situated
in the first intron (37).
Surprisingly, only PNT1A cells react on zinc(II) treatment by
increased expression of NF-κB. In the case of 22Rv1 we observed no
significant changes in NF-κB expression. However, this finding is
not in accordance with the previously reported studies that show
association of progression of prostate cells toward greater
tumourigenic potential with the increasing constitutive levels of
NF-κB activity (38,39).
NF-κB is generally viewed as anti-apoptotic and
oncogenic. However, recent reports suggest that NF-κB may promote
apoptosis and is not necessarily an anti-apoptotic factor in some
situations (40). For example,
Bohuslav et al have shown that p53 stimulates the ribosomal
S6 kinase, which in turn phosphorylates the p65/RELA subunit
(41). This phosphorylation of p65
was found to reduce its affinity to IκBγ, thereby preventing
IκBγ-mediated nuclear export of NF-κB (41). These results indicate a novel
non-classical mechanism of NF-κB activation via p53. However, it is
worth mentioning at this point that in contrast to these
co-operative efforts between p53 and NF-κB, there have been a few
reports which indicate an antagonistic relationship. For instance,
it was shown that p53 and NF-κB repress each other by competing for
a limiting pool of transcriptional co-activator proteins p300 and
CREB-binding protein (CBP) (42).
In our study, NF-κB expression after zinc(II) treatment correlates
with p53 expression in both cell lines, which supports the previous
findings. In our experiments, 22Rv1 did not increase expression of
p53 after zinc treatment. It has been reported that metals in
general are able to induce formation of reactive oxygen species
(ROS) and activate p53-dependend apoptotic pathway (43,44).
Moreover, it has been published that p53 activation by ROS is in
some cases of metal-based drug treatment necessary for induction of
apoptosis (45). Based on our
results and previously published studies, it can be expected that
low expression of p53 and no significant changes in its expression
after zinc treatment may prevent cells from undergoing
p53-dependent apoptosis e.g., caused by oxidative stress. However,
this expectation needs to be experimentally proven.
Ki-67 is a nuclear protein closely associated with
ribosomal RNA transcription and may be crucial for cellular
proliferation (46). Inactivation
of Ki-67 leads to inhibition of ribosomal RNA synthesis (47). Ki-67 has been established as a
promising marker of aggressive prostate cancer. There is plenty of
evidence on the overexpression of this protein in prostate cancer
(48-52). Our results indicate that 22Rv1 cells
express at base line two-fold more Ki-67 than PNT1A cells, in
accordance with previously published data. Zinc treatment increased
Ki-67 significantly in both tumour and non-tumour cell lines.
Collectively, elevated expression of Ki-67, a marker
of proliferation, extremely low expression of p53 and also no
changes in expression of p53 after zinc treatment, high expression
of Bcl-2, especially after zinc treatment and viability and
proliferation curves (Fig. 2)
indicate that zinc has significant positive effect on 22Rv1 cell
line proliferation and viability.
Acknowledgements
Financial support from CYTORES P301/10/0356, CEITEC
CZ.1.05/1.1.00/02.0068 and LPR 2011 and project for conceptual
development of research organization 00064203 was greatly
acknowledged.
References
|
1
|
Bray F, Lortet-Tieulent J, Ferlay J,
Forman D and Auvinen A: Prostate cancer incidence and mortality
trends in 37 European countries: an overview. Eur J Cancer.
46:3040–3052. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ghoneum M and Gollapudi S: Susceptibility
of the human LNCaP prostate cancer cells to the apoptotic effect of
marina crystal minerals (MCM) in vitro. Oncol Rep.
22:155–159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zaichick VY, Sviridova TV and Zaichick SV:
Zinc in human prostate gland: normal, hyperplastic and cancerous. J
Radioanal Nucl Chem. 217:157–161. 1997. View Article : Google Scholar
|
|
4
|
Costello LC and Franklin RB: Zinc is
decreased in prostate cancer: an established relationship of
prostate cancer! J Biol Inorg Chem. 16:3–8. 2011.
|
|
5
|
Krizkova S, Blahova P, Nakielna J, et al:
Comparison of metallothionein detection by using of Brdicka
reaction and enzyme-linked immunosorbent assay employing chicken
yolk antibodies. Electroanalysis. 21:2575–2583. 2009. View Article : Google Scholar
|
|
6
|
Krizkova S, Ryvolova M, Gumulec J, et al:
Electrophoretic fingerprint metallothionein analysis as a potential
prostate cancer biomarker. Electrophoresis. 32:1952–1961. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Krizkova S, Masarik M, Eckschlager T, Adam
V and Kizek R: Effects of redox conditions and zinc(II) ions on
metallothionein aggregation revealed by chip capillary
electrophoresis. J Chromatogr A. 1217:7966–7971. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bataineh ZM, Hani IHB and Al-Alami JR:
Zinc in normal and pathological human prostate gland. Saudi Med J.
23:218–220. 2002.PubMed/NCBI
|
|
9
|
Cory S, Huang DCS and Adams JM: The Bcl-2
family: roles in cell survival and oncogenesis. Oncogene.
22:8590–8607. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bruckheimer EM, Cho S, Brisbay S, et al:
The impact of bcl-2 expression and bax deficiency on prostate
homeostasis in vivo. Oncogene. 19:2404–2412. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gastman BR: Apoptosis and its clinical
impact. Head Neck-J Sci Spec Head Neck. 23:409–425. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gross A, McDonnell JM and Korsmeyer SJ:
Bcl-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Anai S, Goodison S, Shiverick K, Hirao Y,
Brown BD and Rosser CJ: Knock-down of Bcl-2 by antisense
oligodeoxynucleotides induces radiosensitization and inhibition of
angiogenesis in human PC-3 prostate tumor xenografts. Mol Cancer
Ther. 6:101–111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xu L, Yang DJ, Wang SM, et al:
(-)-gossypol enhances response to radiation therapy and results in
tumor regression of human prostate cancer. Mol Cancer Ther.
4:197–205. 2005.PubMed/NCBI
|
|
15
|
Nomura T, Yamasaki M, Nomura Y and Mimata
H: Expression of the inhibitors of apoptosis proteins in
cisplatin-resistant prostate cancer cells. Oncol Rep. 14:993–997.
2005.PubMed/NCBI
|
|
16
|
Concato J, Jain D, Uchio E, Risch H, Li WW
and Wells CK: Molecular markers and death from prostate cancer. Ann
Intern Med. 150:U595–U596. 2009. View Article : Google Scholar
|
|
17
|
Dachille G, Cai T, Ludovico GM, et al:
Prognostic role of cell apoptotic rate in prostate cancer: outcome
of a long-time follow-up study. Oncol Rep. 19:541–545.
2008.PubMed/NCBI
|
|
18
|
Catz SD and Johnson JL: Bcl-2 in prostate
cancer: a minireview. Apoptosis. 8:29–37. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Masarik M, Gumulec J, Sztalmachova M, et
al: Isolation of metallothionein from cells derived from aggressive
form of high-grade prostate carcinoma using paramagnetic
antibody-modified microbeads off-line coupled with electrochemical
and electrophoretic analysis. Electrophoresis. 32:3576–3588. 2011.
View Article : Google Scholar
|
|
20
|
Babula P, Kohoutkova V, Opatrilova R,
Dankova I, Masarik M and Kizek R: Pharmaceutical importance of zinc
and metallothionein in cell signalling. Chim Oggi-Chem Today.
28:18–21. 2010.
|
|
21
|
Krizkova S, Fabrik I, Adam V, Hrabeta J,
Eckschlager T and Kizek R: Metallothionein - a promising tool for
cancer diagnostics. Bratisl Med J-Bratisl Lek Listy. 110:93–97.
2009.PubMed/NCBI
|
|
22
|
Eckschlager T, Adam V, Hrabeta J, Figova K
and Kizek R: Metallothioneins and cancer. Curr Protein Pept Sci.
10:360–375. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Adam V, Fabrik I, Eckschlager T, Stiborova
M, Trnkova L and Kizek R: Vertebrate metallothioneins as target
molecules for analytical techniques. TRAC-Trends Anal Chem.
29:409–418. 2010. View Article : Google Scholar
|
|
24
|
Ryvolova M, Adam V and Kizek R: Analysis
of metallothionein by capillary electrophoresis (review). J
Chromatogr A. 1226:31–42. 2012. View Article : Google Scholar
|
|
25
|
Ryvolova M, Krizkova S, Adam V, et al:
Analytical methods for metallothionein detection. Curr Anal Chem.
7:243–261. 2011.
|
|
26
|
Costello LC, Liu YY, Franklin RB and
Kennedy MC: Zinc inhibition of mitochondrial aconitase and its
importance in citrate metabolism of prostate epithelial cells. J
Biol Chem. 272:28875–28881. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Beyersmann D and Haase H: Functions of
zinc in signaling, proliferation and differentiation of mammalian
cells. Biometals. 14:331–341. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Baranano DE, Ferris CD and Snyder SH:
Atypical neural messengers. Trends Neurosci. 24:99–106. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hogstrand C, Kille P, Nicholson RI and
Taylor KM: Zinc transporters and cancer: a potential role for ZIP7
as a hub for tyrosine kinase activation. Trends Mol Med.
15:101–111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Costello LC and Franklin RB: The
intermediary metabolism of the prostate: A key to understanding the
pathogenesis and progression of prostate malignancy. Oncology.
59:269–282. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Gumulec J, Masarik M, Krizkova S, et al:
Insight to physiology and pathology of zinc(II) ions and their
actions in breast and prostate carcinoma. Curr Med Chem.
18:5041–5051. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Scott SL, Higdon R, Beckett L, et al:
Bcl-2 antisense reduces prostate cancer cell survival following
irradiation. Cancer Biother Radiopharm. 17:647–656. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hess J, Angel P and Schorpp-Kistner M:
AP-1 subunits: quarrel and harmony among siblings. J Cell Sci.
117:5965–5973. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ameyar M, Wisniewska M and Weitzman JB: A
role for AP-1 in apoptosis: the case for and against. Biochimie.
85:747–752. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Leppa S and Bohmann D: Diverse functions
of JNK signaling and c-Jun in stress response and apoptosis.
Oncogene. 18:6158–6162. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wisdom R, Johnson RS and Moore C: c-Jun
regulates cell cycle progression and apoptosis by distinct
mechanisms. EMBO J. 18:188–197. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Charital YM, van Haasteren G, Massiha A,
Schlegel W and Fujita T: A functional NF-kappaB enhancer element in
the first intron contributes to the control of c-fos transcription.
Gene. 430:116–122. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Huang L, Kirschke CP and Zhang Y:
Decreased intracellular zinc in human tumorigenic prostate
epithelial cells: a possible role in prostate cancer progression.
Cancer Cell Int. 6:1–10. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Golovine K, Uzzo RG, Makhov P, Crispen PL,
Kunkle D and Kolenko VM: Depletion of intracellular zinc increases
expression of tumorigenic cytokines VEGF, IL-6 and IL-8 in prostate
cancer cells via NF-kappa B-dependent pathway. Prostate.
68:1443–1449. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Radhakrishnan SK and Kamalakaran S:
Pro-apoptotic role of NF-kappaB: implications for cancer therapy.
Biochim Biophys Acta. 1766:53–62. 2006.PubMed/NCBI
|
|
41
|
Bohuslav J, Chen LF, Kwon H, Mu YJ and
Greene WC: p53 induces NF-kappa B activation by an I kappa B
kinase-independent mechanism involving phosphorylation of p65 by
ribosomal S6 kinase 1. J Biol Chem. 279:26115–26125. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Webster GA and Perkins ND: Transcriptional
cross talk between NF-kappaB and p53. Mol Cell Biol. 19:3485–3495.
1999.PubMed/NCBI
|
|
43
|
Chowdhury R, Chowdhury S, Roychoudhury P,
Mandal C and Chaudhuri K: Arsenic induced apoptosis in malignant
melanoma cells is enhanced by menadione through ROS generation, p38
signaling and p53 activation. Apoptosis. 14:108–123. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Li ZS, Shi KJ, Guan LY, et al: ROS leads
to MnSOD upregulation through ERK2 translocation and p53 activation
in selenite-induced apoptosis of NB4 cells. FEBS Lett.
584:2291–2297. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Bragado P, Armesilla A, Silva A and Porras
A: Apoptosis by cisplatin requires p53 mediated p38 alpha MAPK
activation through ROS generation. Apoptosis. 12:1733–1742. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bullwinkel J, Baron-Luhr B, Ludemann A,
Wohlenberg C, Gerdes J and Scholzen T: Ki-67 protein is associated
with ribosomal RNA transcription in quiescent and proliferating
cells. J Cell Physiol. 206:624–635. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Rahmanzadeh R, Huttmann G, Gerdes J and
Scholzen T: Chromophore-assisted light inactivation of pKi-67 leads
to inhibition of ribosomal RNA synthesis. Cell Prolif. 40:422–430.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Nariculam J, Freeman A, Bott S, et al:
Utility of tissue microarrays for profiling prognostic biomarkers
in clinically localized prostate cancer: the expression of Bcl-2,
E-cadherin, Ki-67 and p53 as predictors of biochemical failure
after radical prostatectomy with nested control for clinical and
pathological risk factors. Asian J Androl. 11:109–118. 2009.
|
|
49
|
Mitra AV, Jameson C, Barbachano Y, et al:
Elevated expression of Ki-67 identifies aggressive prostate cancers
but does not distinguish BRCA1 or BRCA2 mutation carriers. Oncol
Rep. 23:299–305. 2010.PubMed/NCBI
|
|
50
|
Khatami A, Hugosson J, Wang WZ and Damber
JE: Ki-67 in screen-detected, low-grade, low-stage prostate cancer,
relation to prostate-specific antigen doubling time, Gleason score
and prostate-specific antigen relapse after radical prostatectomy.
Scand J Urol Nephrol. 43:12–18. 2009. View Article : Google Scholar
|
|
51
|
Jhavar S, Bartlett J, Kovacs G, et al:
Biopsy tissue microarray study of Ki-67 expression in untreated,
localized prostate cancer managed by active surveillance. Prostate
Cancer Prostatic Dis. 12:143–147. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li RL, Heydon K, Hammond ME, et al: Ki-67
staining index predicts distant metastasis and survival in locally
advanced prostate cancer treated with radiotherapy: an analysis of
patients in radiation therapy oncology group protocol 86-10. Clin
Cancer Res. 10:4118–4124. 2004. View Article : Google Scholar
|