Introduction
Despite recent advances in diagnosis and treatment,
lung cancer remains the leading cause of cancer death in males and
the second leading cause of cancer death in females in the world
(1). Non-small cell lung cancer
(NSCLC) is the most prevalent and accounts for 80% of lung cancers,
and patients usually present in the advanced stages with poor
prognosis and difficulty in management. As for primary
chemotherapy, advanced NSCLC is often treated with cisplatin or
carboplatin, in combination with gemcitabine, paclitaxel,
docetaxel, etoposide, or vinorelbine (2). The platinum-based chemotherapy was
also adopted as standard use of adjuvant chemotherapy for NSCLC.
However, the toxicity to normal cells largely reduced the success
of the platinum-based chemotherapy.
The epidermal growth factor receptor (EGFR) is a
promising target for anticancer therapy due to its expression or
overexpression in a variety of tumors, including NSCLC (3). High levels of EGFR expression and
dysregulation might promote tumor growth by increasing cell
proliferation, motility, invasive capacity or by evading apoptosis,
which were thus associated with poorer prognosis (4). Recently, gefitinib was indicated for
the treatment of adult patients with locally advanced or metastatic
NSCLC with activating mutations of the tyrosine kinase domain of
EGFR (5,6). EGFR mutation-positive patients have
better efficacy outcomes with first-line gefitinib when compared
with those who are EGFR mutation-negative. In addition to the EGFR
mutation status of the patients, several adverse drug reactions
largely limited the use of the EGFR-target therapy (7,8). Thus,
there is an urgent need to identify new therapeutic agents for
alternative treatments or combination therapies for lung
cancer.
Norcantharidin is the demethylated analog of
cantharidin isolated from blister beetles (Mylabris
phalerata Pall.). Norcantharidin was reported to possess
anticancer activity but less nephrotoxicity than cantharidin
(9). There is accumulating evidence
that norcantharidin inhibits the proliferation of a variety of
human tumor cell lines (10–12),
induces apoptosis (10,13,14),
suppresses the invasion and metastasis (15), inhibits angiogenesis (16), represses tumor growth in animals
(17,18). However, few studies have reported
the anticancer effect of norcantharidin against human lung cancer
cells.
In this study, we evaluated whether norcantharidin
exhibits anticancer effects against the human lung cancer cell
lines, A549 (EGFR mutation-negative) and PC9 (EGFR
mutation-positive), and determined the effects of combination
treatments with gefitinib and cisplatin, respectively. In addition,
since norcantharidin has been reported as a protein phosphatase 1
(PP1) and protein phosphatase 2A (PP2A) inhibitor (19–21),
the roles of the norcantharidin-activated signaling pathways will
be further discussed.
Materials and methods
Chemicals and antibodies
Norcantharidin and cisplatin were purchased from
Sigma-Aldrich (St. Louis, MO, USA). Epidermal growth factor (EGF)
was purchased from R&D Systems, Inc. (Minneapolis, MN, USA).
Gefitinib was provided by Astra Zeneca. Antibodies against the
cdc25C, the cyclin B1, the cyclin-dependent kinase 1 (cdk1), and
phospho-specific EGFR antibodies (pY1068, pS1046/1047, pY1148,
pY1173) were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Antibody against the EGFR (sc-03) was purchased
from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Antibody
against the α-tubulin was purchased from Sigma-Aldrich.
Cell culture
Human lung cancer cell lines A549 and PC9 were
cultured at 37°C in 5% CO2 in RPMI-1640 medium
supplemented with 10% FBS (Hyclone), 50 U/ml penicillin G, and 50
mg/ml streptomycin sulfate.
Trypan blue exclusion assay
Cells (1×105) were seeded in 6-well cell
culture cluster (Costar, Cambridge, MA, USA) overnight and then
treated with different concentrations of norcantharidin (0, 12.5,
25 and 100 μM), respectively. After treatment for 24 to 72 h, cells
were harvested by trypsin-EDTA and the cell pellet was resuspened
in culture medium containing 0.04% trypan blue and the viable cells
were counted by a hemocytometer.
MTT assay
Cells were seeded in a 24-well cell culture cluster
(Costar) at a density of 2×104 cells per ml and cultured
overnight prior to drug treatment. After norcantharidin treatment
for 48 h, the medium was discarded and replaced with an equal
volume (0.5 ml) of fresh medium containing 0.456 mg/ml
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl-tetrazolium bromide (MTT;
Sigma Chemical Co., St. Louis, MO, USA) and incubated for 1 h at
37°C in the dark. The medium was discarded, and cells were combined
with 100 μl dimethyl sulfoxide (DMSO) (Sigma Chemical Co.) to
dissolve the formazan produced. Cell viability was determined
according to the colorimetric comparison by reading optical density
(OD) values from a microplate reader (Spectra Max 250; Spectra
Diode Laboratories, Inc., San Jose, CA, USA) at an absorption
wavelength of 570 nm.
Western blotting
Cells were scraped from 10-cm dishes and suspended
in RIPA lysis buffer (980 μl RIPA, 5 μl aprotinin, 5 μl PMSF, 5 μl
EGTA and 5 μl Na3VO4) on ice. Collected cells
were fractured by sonication on ice and then centrifuged at 10,000
× g, at 4°C for 15 min. The protein concentration was determined
using Bradford reagent and then 30 μg of extracted protein in 4.5
μl of sample buffer (1.6 ml 1.25 M Tris-HCl, 3.2 ml glycerol, 0.64
g SDS, 1.6 ml β-mercaptoethanol, 0.8 ml 0.5% bromophenol blue and
0.8 ml dH2O) was denatured at 100°C for 10 min. Proteins
were separated by 8% SDS-PAGE and then electrophoretically
transferred to a nitrocellulose membrane. Subsequently, the
membranes were incubated in the presence of different primary
antibodies at 4°C overnight and then the membrane was incubated
with different secondary antibodies at 37°C for 1 h. Finally, ECL
solution was used for antibody binding and chemiluminescence of the
membrane. All results shown are representative of at least two
separate experiments.
Cell cycle analysis and determination of
apoptotic cells in sub-G1 phase
Procedures were carried out according to previously
reported methods (22). In brief,
after treatment with norcantharidin for 48 h, the cells were
trypsinized and resuspended in 70% ethanol, the cells were then
incubated on ice for at least 1 h and resuspended in 1 ml of cell
cycle assay buffer (0.38 mM sodium citrate, 0.5 mg/ml RNase A, and
14.9 μM propidium iodide) at a concentration of 5×105
cells/ml. Samples were stored in the dark at 4°C until cell cycle
analysis, which was carried out using a flow cytometer and ModFit
LT 2.0 software (Verity Software, Topsham, ME).
Flow cytometry analysis
A FACS Calibur flow cytometer (Becton Dickinson,
Bedford, MA) equipped with a 488-nm argon laser was used for the
flow cytometric analysis. Forward and side scatters were used to
establish size gates and exclude cellular debris from the analysis.
The excitation wavelength was set at 488 nm. In each measurement, a
minimum of 15,000 cells were analyzed. Data were acquired and
analyzed using the Cell Quest software (Becton Dickinson). Relative
change in the mean fluorescence intensity was calculated as the
ratio between mean fluorescence intensity in the channel of the
treated cells and that of the control cells.
Transwell migration assay
Transwell migration assay was carried out with a
24-well chamber (Costar 3422, Corning Inc., Corning, NY). The lower
and upper chambers were separated by a polycarbonate membrane (8 μm
pore size). Cells (1×105) were resuspended in RPMI
medium containing 1% FBS in the upper chamber. The RPMI medium
containing 20% FBS was added to the lower chamber. Cells were
allowed to migrate for 10 h (A549 cells) or 16 h (PC9 cells) at
37°C in a humidified atmosphere containing 5% CO2. The
membrane was fixed in methanol for 20 min at 4°C, and then stained
with Liu’s stain A for 5 min and Liu’s stain B for 30 min. Cells on
the upper side of the membrane were removed by PBS-rinsed cotton
swabs. Cells on the lower side of the membrane were counted under a
light microscope with the 10× objective lens. Two individuals
blinded to the treatment of the transwell filter counted cells from
four random fields in each of two wells per treatment; and the
results were pooled. Each experiment was performed in
triplicate.
Immunofluorescence and fluorescence
microscopic analysis
Cells were fixed using 3.7% formaldehyde for 20 min
at room temperature and then washed with PBS and wash buffer (0.1%
BSA in PBS). After incubation in blocking buffer (5% BSA and 0.3%
Triton X-100 in PBS) for 45 min at room temperature, the fixed
cells were stained for F-actin with 2 U/ml Oregon Green 488
phalloidin (Molecular Probes, Eugene, OR, USA) for 30 min and then
stained for DNA with 0.2 μg/ml 4′,6-Diamidino-2-phenylindole (DAPI)
for 10 min. The images were recorded by an Olympus IX70
fluorescence microscope (Olympus America Inc., Melville, NY, USA).
Cells from ten random fields in each treatment experiment were
counted and the ratio of the cells with bi-nucleus was calculated.
Each experiment was performed in triplicate.
Statistics
Data are shown as the mean ± SEM except where
indicated. Statistical comparison of data between groups were
performed using one-way analysis of variance (ANOVA), followed by
Student’s t-test. A p-value <0.05 is considered statistically
significant.
Results
Norcantharidin retards cell growth of
human lung cancer cells
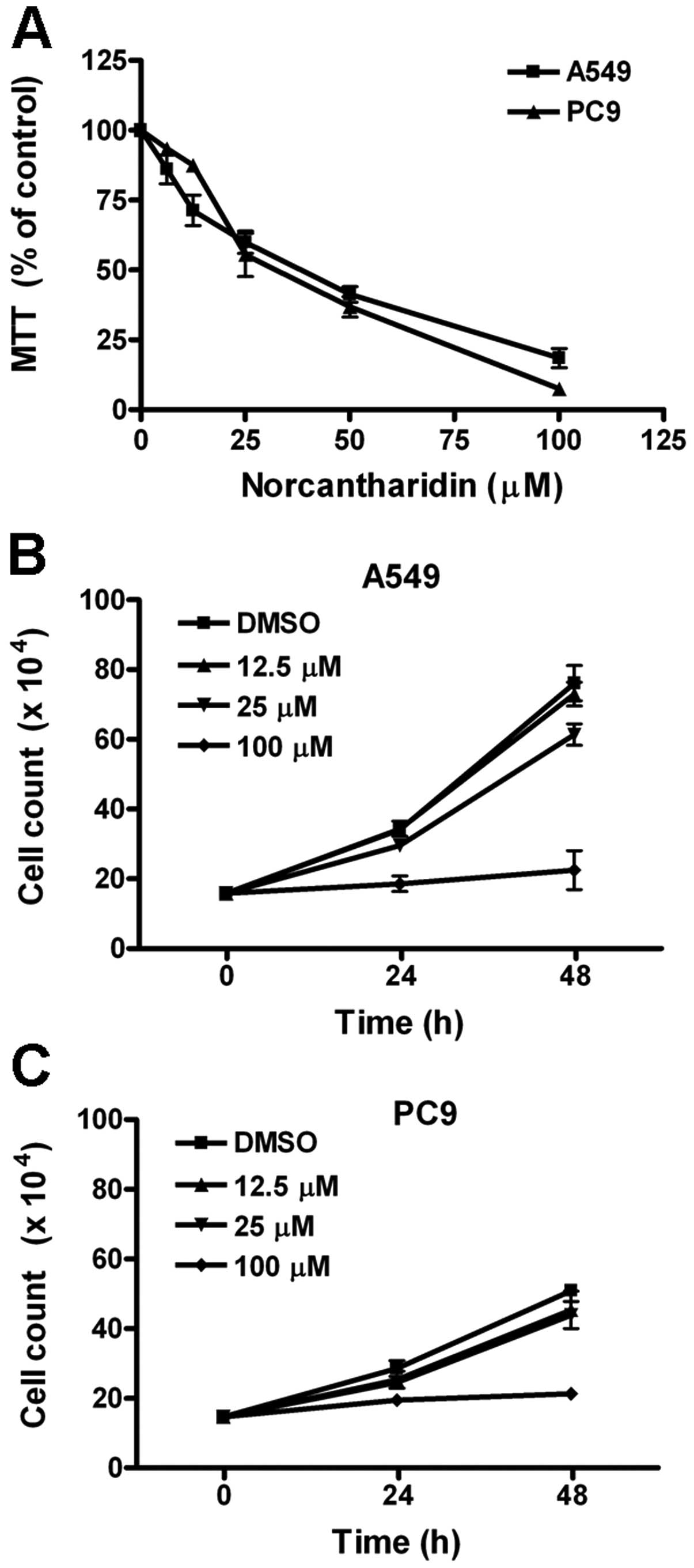
Using MTT assay, we first evaluated the effect of
norcantharidin on cell proliferation of two human lung cancer cell
lines A549 and PC9. After treatments with 6.25, 12.5, 25, 50 and
100 μM norcantharidin for 48 h, the MTT values were
dose-dependently decreased (Fig.
1A). The IC50 for A549 and PC9 cells were 29.3±2.9 μM and
31.0±1.8 μM, respectively. Using trypan blue exclusion assay, we
further counted the number of survived cells under treatments with
12.5, 25 and 100 μM norcantharidin for 24 and 48 h, respectively.
Both Fig. 1B and C show that
norcantharidin, especially at 100 μM, inhibited the increase in
survived cell count of A549 and PC9 cells. These results indicated
that norcantharidine suppressed cell growth of the two human lung
cancer cell lines studied.
Norcantharidin inhibits cell cycle and
induces cell death
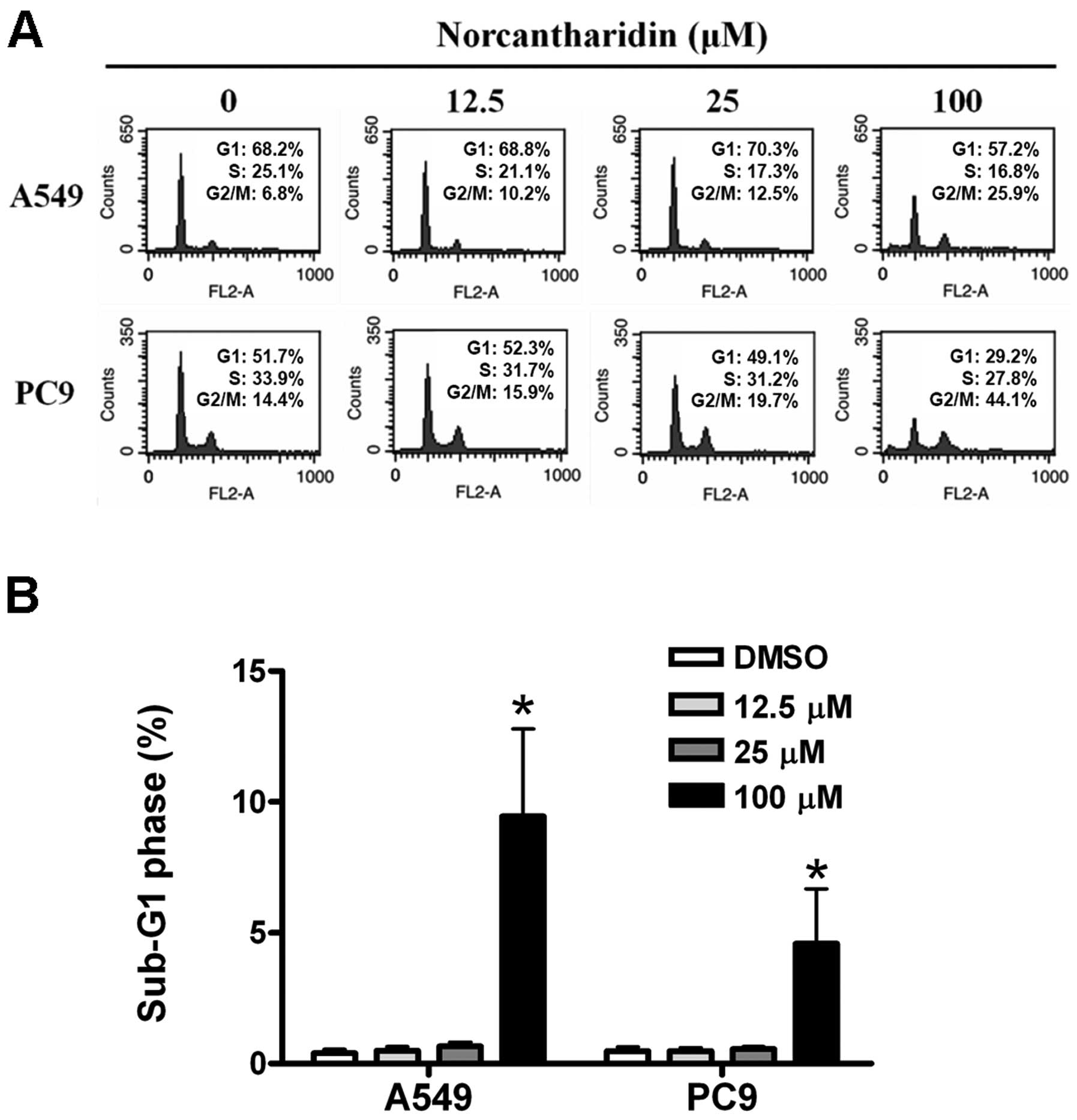
We analyzed the changes of cell cycle distribution
of the two lung cancer cell lines treated with 12.5, 25 and 100 μM
norcantharidin for 48 h, respectively. The results revealed that
the proportions of the treated cells at G2/M phase were
increased in a dose-dependent manner (Fig. 2A). Moreover, the proportions of
sub-G1 phase cells were increased in the two lung cancer cell lines
after 100 μM norcantharidin treatment for 48 h (Fig. 2B).
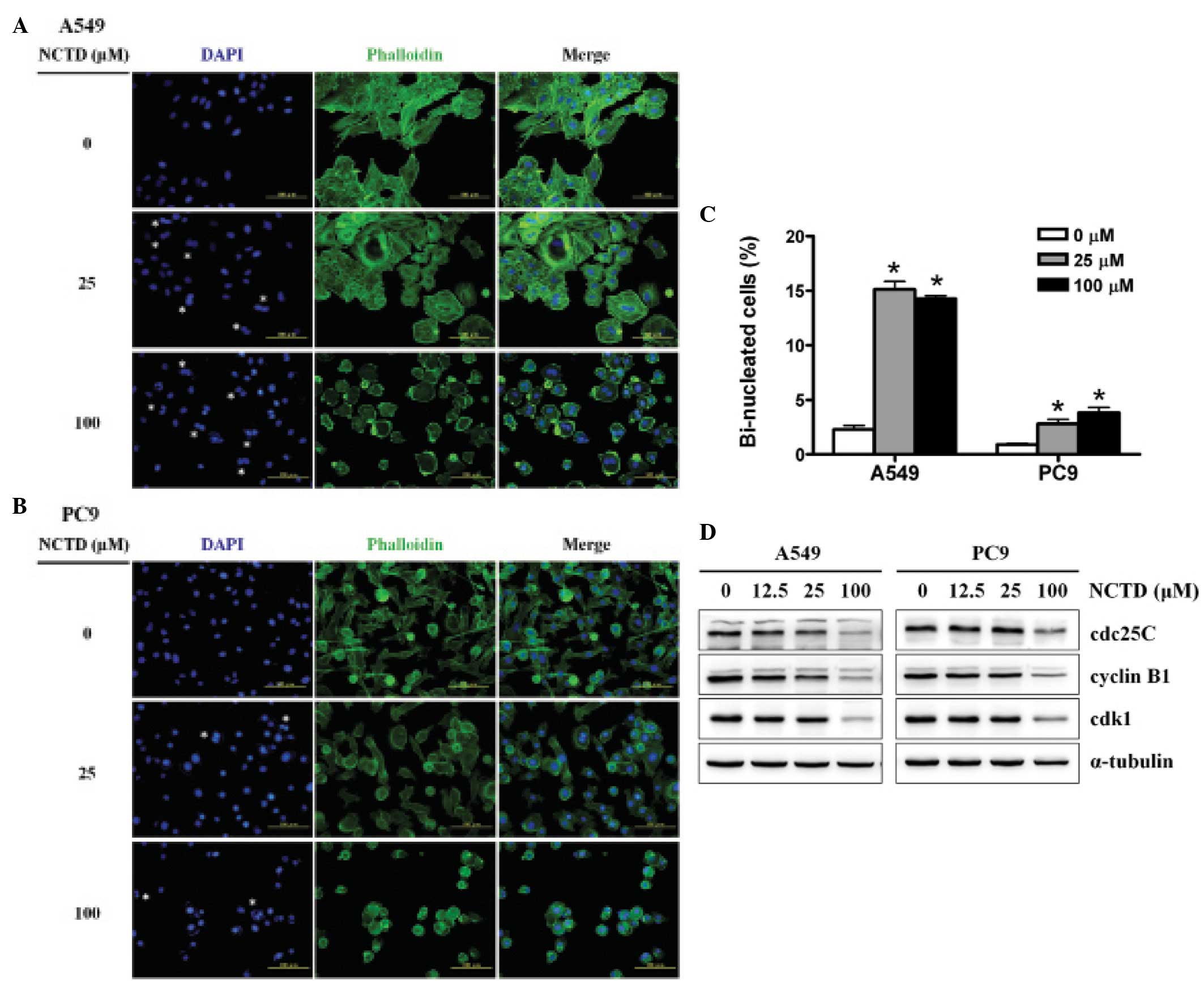
Using DAPI to stain the nucleus and phalloidin to
stain F-actin, we found that treatments with 25 and 100 μM
norcantharidin for 48 h significantly increased the proportions of
the bi-nucleated cells in the A549 (Fig. 3A) and PC9 cells (Fig. 3B) as compared with the untreated
cells. Fig. 3C shows that the
increased extents of the bi-nucleated proportion of the A549 cells
were higher than those of the PC9 cells. Using western blotting, we
further determined the protein contents of the cdc25, cyclin B1 and
cdk1, the important regulators at the G2/M check point,
and found that the three protein levels were obviously decreased in
the A549 cells after 12.5, 25 and 100 μM norcantharidin treatment
for 48 h; and the decrease in the PC9 cells was significant at the
norcantharidin concentration of 100 μM (Fig. 3D). These results suggested that
norcantharidin caused cell cycle arrest at G2/M phase
and high dose of norcantharidin induced cell death of the two human
lung cancer cell lines.
Norcantharidin represses cell
migration
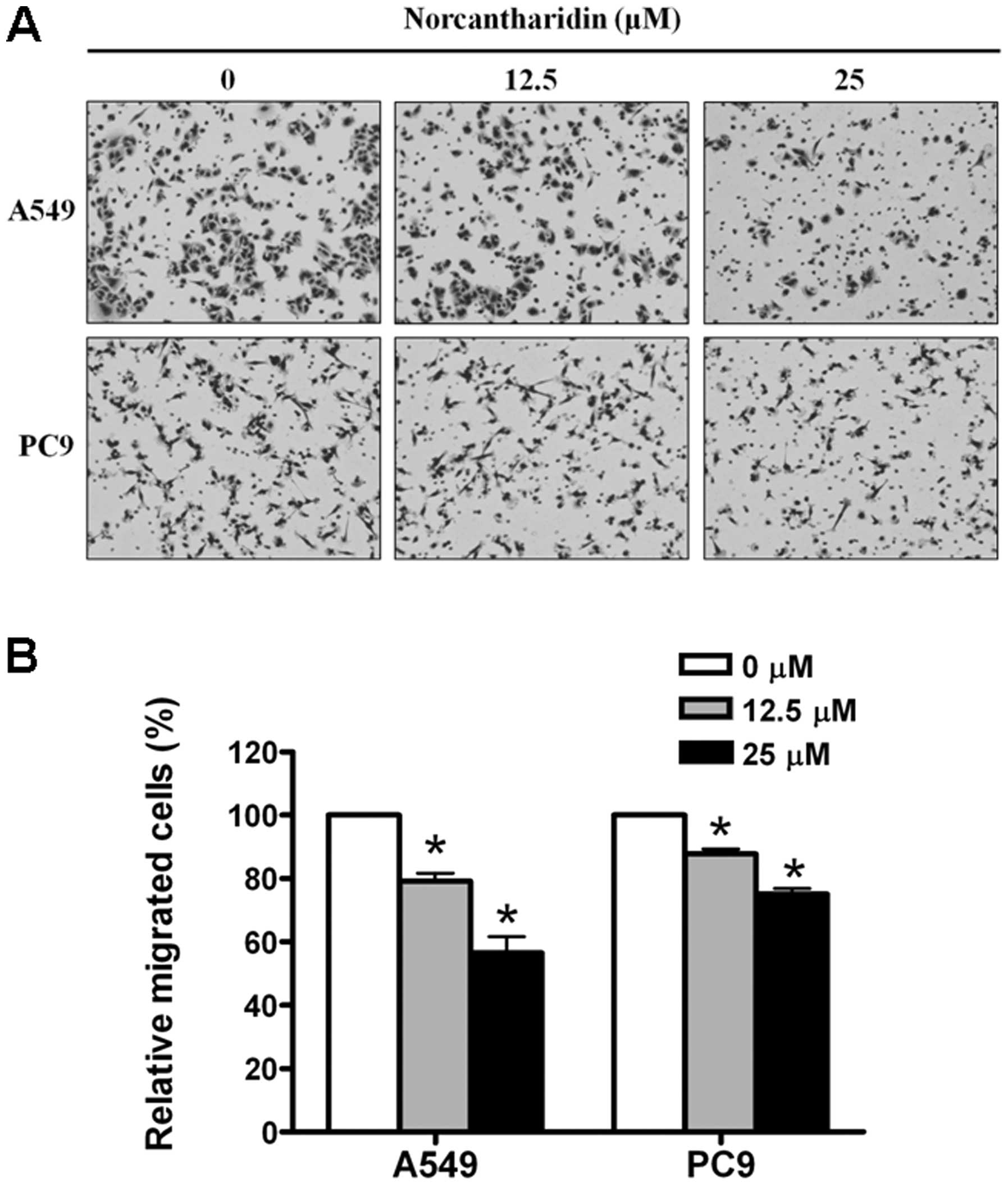
Using transwell cell migration assay, we evaluated
whether or not norcantharidin affects the migration ability of the
human lung cancer cells. The relatively low concentrations of
norcantharidin (12.5 and 25 μM, respectively) were used for the
experiments due to their minor effects on cell survival during the
first 10 h exposures for the A549 cells (Fig. 1B) and the first 16 h exposures for
the PC9 cells (Fig. 1C). We found
that norcantharidin dose-dependently reduced the migration ability
of the two cancer cell lines studied (Fig. 4).
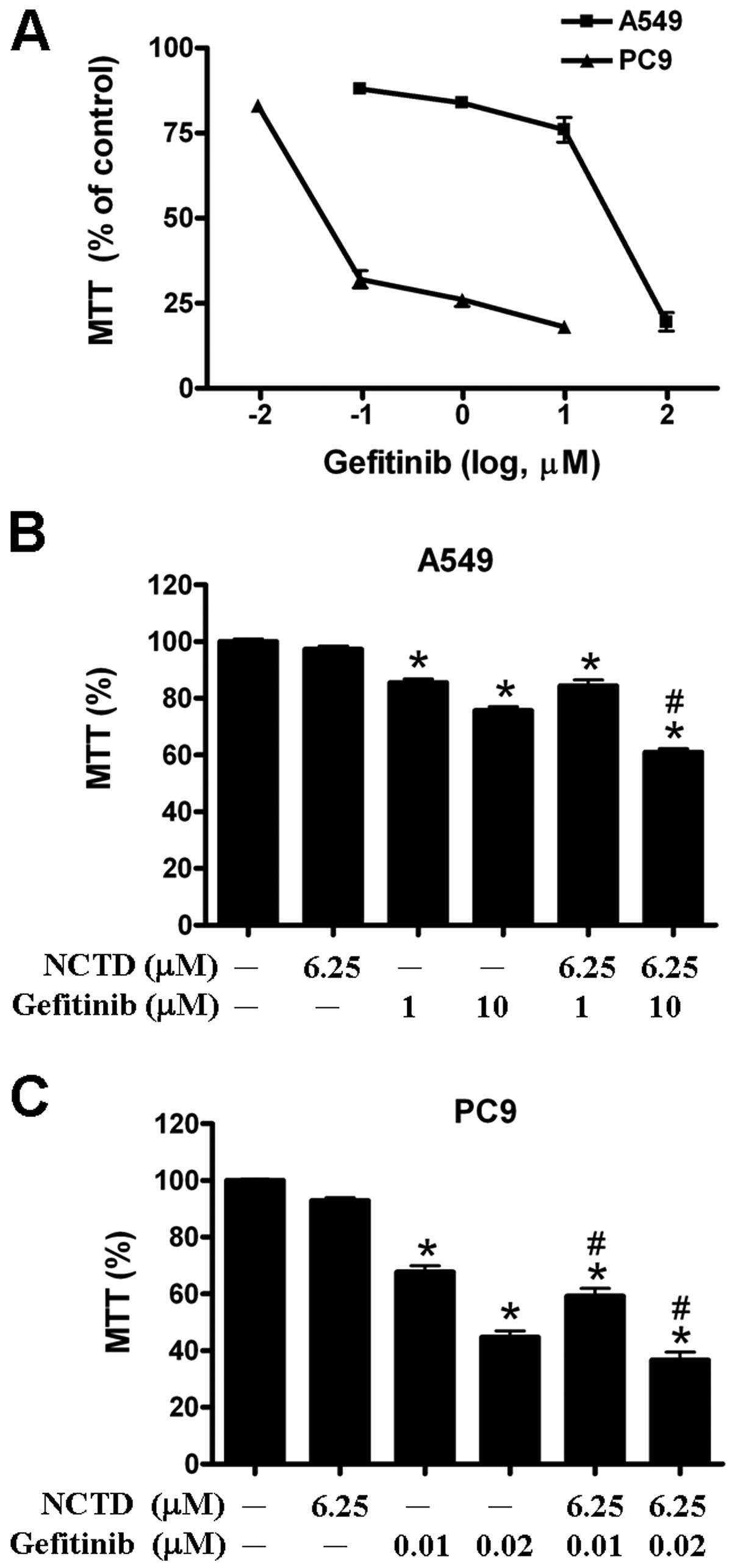
Norcantharidin enhanced anticancer
effects of gefitinib and cisplatin
We further examined whether norcantharidin can
enhance the cytotoxic effect of anticancer drugs against human lung
cancer cells. Gefitinib is one of tyrosine kinase inhibitors and
has been clinically used for lung cancer patients. For the two
human lung cancer cell lines, A549 cells were more resistant to
gefitinib than PC9 cells (Fig. 5A).
We found that combined treatment with 10 μM gefitinib, 6.25 μM
norcantharidin can significantly enhance the cytotoxic effect of
gefitinib against the A549 cells (Fig.
5B). Similarly, combined treatment with 0.02 μM gefitinib, 6.25
μM norcantharidin can significantly enhance the cytotoxic effect of
gefitinib against the gefitinib-sensitive PC9 cells (Fig. 5C).
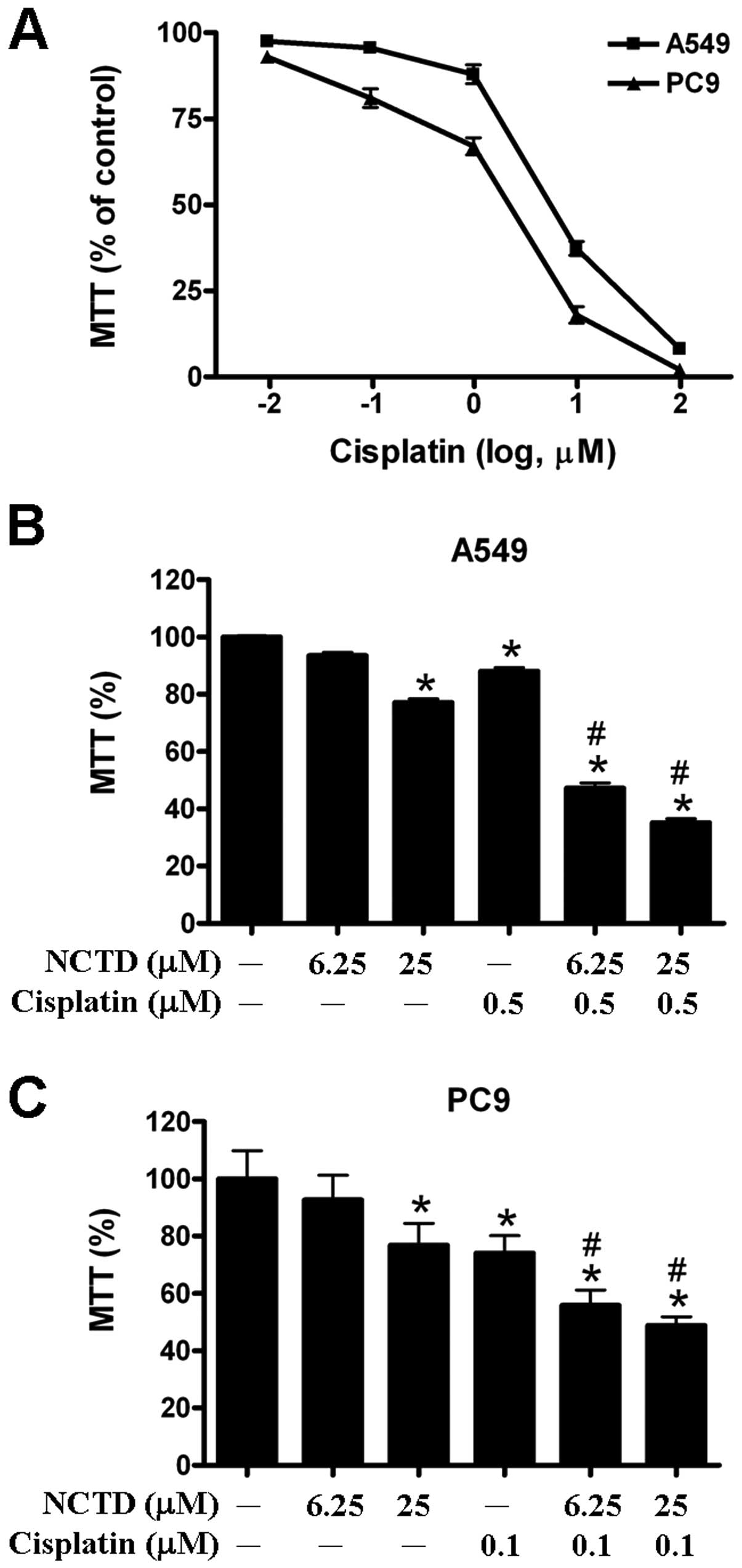
A549 and PC9 were shown to have similar sensitivity
to cisplatin (Fig. 6A). We found
that combined treatment with 0.5 μM cisplatin, 6.25 μM and 25 μM
norcantharidin significantly enhanced the cytotoxic effect of
cisplatin against the A549 cells (Fig.
6B). Moreover, when combined with 0.1 μM cisplatin,
norcantharidin significantly enhanced the cytotoxic effect of
cisplatin against the PC9 cells (Fig.
6C). These results indicated that norcantharidin enhanced the
anticancer effects of gefitinib and cisplatin against human lung
cancer cells.
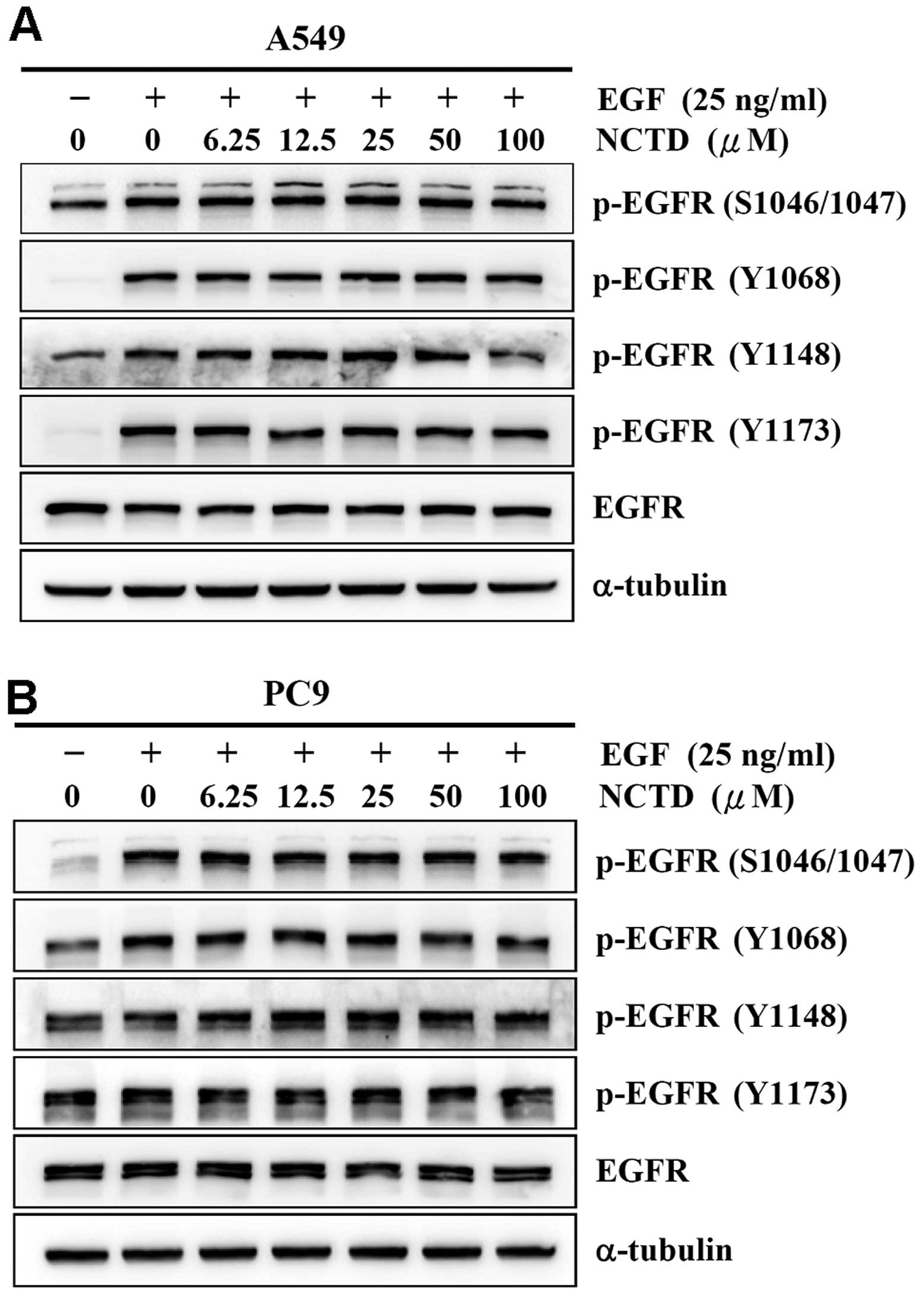
Norcantharidin does not alter
phosphorylation status of EGFR
Mutations that lead to EGFR upregulation or
overactivity are often associated with human lung cancer. Since
signaling through EGFR is a key regulator in proliferation and
migration of lung cancer cells, we hypothesized that EGFR could be
involved in the norcantharidin-induced cytotoxicity of human lung
cancer cells. To test this hypothesis, we examined the effect of
norcantharidin on the phosphorylation status of EGFR. After serum
starvation for 24 h, treatment with 25 ng/ml EGF for 1 h can
significantly increase phosphorylation of EGFR at pS1046/1047,
pY1068, pY1148, and pY1173 sites of the A549 (Fig. 7A) and PC9 cells (Fig. 7B). Of note, treatments with 6.25,
12.5, 25, 50, and 100 μM norcantharidin did not significantly
change the phosphorylation status of all examined phosphorylation
sites of EGFR in the two cell lines (Fig. 7). The results suggested that EGFR
could not be involved in the norcantharidin-induced cytotoxicity of
human lung cancer cells.
Discussion
In this study, we found that norcantharidin exhibits
anticancer effects against human lung cancer cell lines A549 (EGFR
mutation-negative) and PC9 (EGFR mutation-positive), including cell
growth inhibition, cell cycle arrest at G2/M phase, cell
migration reduction, and even apoptosis when the concentration is
high. Our findings are consistent with the previous studies in
various cancers (10–12,15,16,23).
Notable, we demonstrated that norcantharidin enhances the
anticancer effects of gefitinib and cisplatin, respectively. Our
results suggested the potential for norcantharidin as a
chemotherapeutic agent for treating lung cancer.
Inhibition of cell growth induced by norcantharidin
might be associated with disturbance of cell cycle progression of
the lung cancer cells. Norcantharidin increased the cell proportion
at the G2/M phase (Fig.
2A), the number of bi-nuclear cells (Fig. 3C), and reduced the protein contents
of the important regulators at the G2/M check point
(cdc25, cyclin B1 and cdk1, Fig.
3D), suggesting that norcantharidin might retard the cell cycle
at the G2/M phase. The results are consistent with
previous findings in human glioblastoma (24), hepatoma cells (10), leukemic Jurkat T cells (25) and breast cancer cells (26), though the proposed underlying
mechanisms are controversial. In addition, it is noteworthy that
apoptosis induced by norcantharidin was not obviously observed in
the lung cancer cells until high concentration (100 μM). These
findings suggested that norcantharidin mainly exhibits cytostatic
effects against lung cancer cells.
The signaling through EGFR is important in the
regulation of proliferation and migration of lung cancer cells
(4). This signaling pathway is
regulated by phosphorylation modification of the cytosolic domain
of EGFR through protein kinases and phosphatases (27). It has been reported that
norcantharidin is a protein phosphatase 1 (PP1) and protein
phosphatase 2A (PP2A) inhibitor (19,20).
Thus, we tested whether EGFR is involved in the
norcantharidin-induced cytotoxicity of human lung cancer cells. We
used calyculin A, an inhibitor of protein phosphatase PP1 and PP2A,
as a positive control for the effects of protein phosphatase
inhibitor on the phosphorylated pattern of EGFR, and found that 5
nM calyculin A treatments increased the phosporylated status of
EGFR at Ser1046/1047, but decreased the phosphorylation at Tyr1068,
Tyr1148 and Tyr1173, respectively (data not shown). However,
norcantharidin treatments did not obviously alter the
EGF-stimulated phosphorylation status of the EGFRs in the two cell
lines (Fig. 7). Moreover, the two
cell lines with different EGFR mutation status have similar
IC50 values for norcantharidin treatments (Fig. 1A), though the PC9 cells (EGFR
mutation-positive) are more sensitive to gefitinib than the A549
cells (EGFR mutation-negative). These results suggested that EGFR
might not be involved in the norcantharidin-induced anticancer
effects against these two kinds of lung cancer cells.
In conclusion, we provide in vitro evidence
in human lung cancer cell lines to suggest that norcantharidin
retards cell growth, disturbs cell cycle progression, and represses
cell migration, as well as enhances the anticancer effects of
gefitinib and cisplatin.
Acknowledgments
We thank Ms. Shu-Hui Li for her excellent technical
assistance. This work was partly supported by the grants V99E2-006
and V99E2-008 from Taipei Veterans General Hospital, a grant for
the Center of Excellence for Cancer Research at Taipei Veterans
General Hospital, the Department of Health (DOH99-TDC-111-007,
DOH100-TDC-111-007, and DOH101-TDC-111-007), Executive Yuan, and a
grant from Ministry of Education, Aim for the Top University Plan
and the grants NSC97-2320-B-010-022-MY3 and
NSC101-2320-B-010-068-MY3 from the National Science Council,
Taiwan, Republic of China.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: 2011 Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Ramalingam S and Belani C: Systemic
chemotherapy for advanced non-small cell lung cancer: recent
advances and future directions. Oncologist. 13:5–13. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sharma SV, Bell DW, Settleman J and Haber
DA: Epidermal growth factor receptor mutations in lung cancer. Nat
Rev Cancer. 7:169–181. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mendelsohn J: Blockade of receptors for
growth factors: an anticancer therapy - the fourth annual Joseph H.
Burchenal American Association of Cancer Research Clinical Research
Award Lecture. Clin Cancer Res. 6:747–753. 2000.
|
|
5
|
Cohen MH, Williams GA, Sridhara R, Chen G,
McGuinn WD Jr, Morse D, Abraham S, Rahman A, Liang C, Lostritto R,
Baird A and Pazdur R: United States Food and Drug Administration
Drug Approval summary: Gefitinib (ZD1839; Iressa) tablets. Clin
Cancer Res. 10:1212–1218. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wu JY, Shih JY, Chen KY, Yang CH, Yu CJ
and Yang PC: Gefitinib therapy in patients with advanced non-small
cell lung cancer with or without testing for epidermal growth
factor receptor (EGFR) mutations. Medicine. 90:159–167. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lacouture ME: Mechanisms of cutaneous
toxicities to EGFR inhibitors. Nat Rev Cancer. 6:803–812. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jiang H: Overview of gefitinib in
non-small cell lung cancer: an Asian perspective. Jpn J Clin Oncol.
39:137–150. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang GS: Medical uses of mylabris in
ancient China and recent studies. J Ethnopharmacol. 26:147–162.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen YN, Chen JC, Yin SC, Wang GS, Tsauer
W, Hsu SF and Hsu SL: Effector mechanisms of norcantharidin-induced
mitotic arrest and apoptosis in human hepatoma cells. Int J Cancer.
100:158–165. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hill TA, Stewart SG, Ackland SP, Gilbert
J, Sauer B, Sakoff JA and McCluskey A: Norcantharimides, synthesis
and anticancer activity: Synthesis of new norcantharidin analogues
and their anticancer evaluation. Bioorg Med Chem. 15:6126–6134.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fan YZ, Fu JY, Zhao ZM and Chen CQ:
Inhibitory effect of norcantharidin on the growth of human
gallbladder carcinoma GBC-SD cells in vitro. Hepatobiliary Pancreat
Dis Int. 6:72–80. 2007.PubMed/NCBI
|
|
13
|
Peng F, Wei YQ, Tian L, Yang L, Zhao X, Lu
Y, Mao YQ, Kan B, Lei S, Wang GS, Jiang Y, Wang QR, Luo F, Zou LQ
and Liu JY: Induction of apoptosis by norcantharidin in human
colorectal carcinoma cell lines: involvement of the CD95
receptor/ligand. J Cancer Res Clin Oncol. 128:223–230. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liao HF, Su SL, Chen YJ, Chou CH and Kuo
CD: Norcantharidin preferentially induces apoptosis in human
leukemic Jurkat cells without affecting viability of normal blood
mononuclear cells. Food Chem Toxicol. 45:1678–1687. 2007.
View Article : Google Scholar
|
|
15
|
Chen YJ, Shieh CJ, Tsai TH, Kuo CD, Ho LT,
Liu TY and Liao HF: Inhibitory effect of norcantharidin, a
derivative compound from blister beetles, on tumor invasion and
metastasis in CT26 colorectal adenocarcinoma cells. Anticancer
Drugs. 16:293–299. 2005. View Article : Google Scholar
|
|
16
|
Chen YJ, Tsai YM, Kuo CD, Ku KL, Shie HS
and Liao HF: Norcantharidin is a small-molecule synthetic compound
with anti-angiogenesis effect. Life Sci. 85:642–651. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang EB, Tang WY, Zhang K, Cheng LY and
Mack PO: Norcantharidin inhibits growth of human HepG2
cell-transplanted tumor in nude mice and prolongs host survival.
Cancer Lett. 117:93–98. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fan YZ, Zhao ZM, Fu JY, Chen CQ and Sun W:
Norcantharidin inhibits growth of human gallbladder carcinoma
xenografted tumors in nude mice by inducing apoptosis and blocking
the cell cycle in vivo. Hepatobiliary Pancreat Dis Int. 9:414–422.
2010.PubMed/NCBI
|
|
19
|
Liu XH, Blazsek I, Comisso M, Legras S,
Marion S, Quittet P, Anjo A, Wang GS and Misset JL: Effects of
norcantharidin, a protein phosphatase type-2A inhibitor, on the
growth of normal and malignant haemopoietic cells. Eur J Cancer.
31A:953–963. 1995.PubMed/NCBI
|
|
20
|
McCluskey A, Ackland SP, Gardiner E,
Walkom CC and Sakoff JA: The inhibition of protein phosphatases 1
and 2A: a new target for rational anti-cancer drug design?
Anticancer Drug Des. 16:291–303. 2001.PubMed/NCBI
|
|
21
|
Hill TA, Stewart SG, Sauer B, Gilbert J,
Ackland SP, Sakoff JA and McCluskey A: Heterocyclic substituted
cantharidin and norcantharidin analogues-synthesis, protein
phosphatase (1 and 2A) inhibition, and anti-cancer activity. Bioorg
Med Chem Lett. 17:3392–3397. 2007. View Article : Google Scholar
|
|
22
|
Wu CW, Ping YH, Yen JC, Chang CY, Wang SF,
Yeh CL, Chi CW and Lee HC: Enhanced oxidative stress and aberrant
mitochondrial biogenesis in human neuroblastoma SH-SY5Y cells
during methamphetamine induced apoptosis. Toxicol Applied
Pharmacol. 220:243–251. 2007. View Article : Google Scholar
|
|
23
|
Luan J, Duan H, Liu Q, Yagasaki K and
Zhang G: Inhibitory effects of norcantharidin against human lung
cancer cell growth and migration. Cytotechnology. 62:349–355. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hong CY, Huang SC, Lin SK, Lee JJ, Chueh
LL, Lee CH, Lin JH and Hsiao M: Norcantharidin-induced post-G(2)/M
apoptosis is dependent on wild-type p53 gene. Biochem Biophys Res
Commun. 276:278–285. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liao HF, Chen YJ, Chou CH, Wang FW and Kuo
CD: Norcantharidin induces cell cycle arrest and inhibits
progression of human leukemic Jurkat T cells through
mitogen-activated protein kinase-mediated regulation of
interleukin-2 production. Toxicol In Vitro. 25:206–212. 2011.
View Article : Google Scholar
|
|
26
|
Yang PY, Chen MF, Kao YH, Hu DN, Chang FR
and Wu YC: Norcantharidin induces apoptosis of breast cancer cells:
involvement of activities of mitogen activated protein kinases and
signal transducers and activators of transcription. Toxicol In
Vitro. 25:699–707. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Millward TA, Zolnierowicz S and Hemmings
BA: Regulation of protein kinase cascades by protein phosphatase
2A. Trends Biochem Sci. 24:186–191. 1999. View Article : Google Scholar : PubMed/NCBI
|