Introduction
Dendritic cells (DCs) are the most potent
antigen-presenting cells, critical for the induction of specific
immune responses (1,2). Despite some differences between the
murine and human immune system, DC-based cellular vaccines in mouse
tumor models may provide significant information on how to optimize
therapeutic antitumor approaches in humans (3). DCs have been proved effective as both
prophylactic and therapeutic vaccines in a number of experimental
models (4). One of the possible
ways to augment DC efficacy in presentation of tumor antigens to
effector cells is pulsing them with tumor cell lysates (5). However, DCs pulsed with tumor antigens
are not usually able to reach final maturity, which affects their
properties to induce antitumor response. Prior to in vivo
application, incubation of DCs with certain immunostimulators is
therefore necessary. The change in their phenotype greatly enhances
their antigen-presenting properties (6,7). It
appears that DCs stimulated in such a manner are able to further
differentiate to respond to factors secreted by the host immune
system.
As the immune response of the host may be suppressed
by the tumor, injections of adjuvants/cytokines (such as IL-12) in
combination with cellular vaccines may have a crucial effect on
overcoming tumor evasion mechanisms (8,9).
Systemic injections of IL-12 lead to enhancement of natural killer
(NK) and cytotoxic T-cell activity and result in the increase of
IFN-γ production, as well as in affecting differentiation of
antigen-specific Th lymphocytes (10,11).
IL-12 was found to effectively inhibit the growth of several
experimental tumors in mice, via IFN-γ and T cell-dependent
pathways, as well as due to anti-angiogenic effects (12,13).
JAWSII cells, immortalized C57BL/6 murine bone
marrow-derived DCs, seem to be a promising candidate for use in
cancer therapy in a cellular vaccine approach (14,15).
In a previous study, we analyzed phenotype and functional
properties of JAWSII cells and optimized conditions of their
stimulation (16). The purpose of
this study was to determine the in vivo applicability of
JAWSII cells as a component of DC-based cellular vaccine used in
combination with IL-12 in a murine B78-H1 melanoma model. Three
antitumor approaches of this combination have been tested:
prophylactic, local (intratumoral), and systemic therapy.
Materials and methods
Cells
JAWSII cells were purchased from the American Type
Culture Collection (CRL-11904). The cells were grown in RPMI-1640
medium (Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10%
non-inactivated FCS (Gibco-Invitrogen, Paisley, Scotland, UK),
antibiotics (penicillin + streptomycin + amphotericin,
Sigma-Aldrich), and 5 ng/ml murine GM-CSF (PeproTech, London, UK).
JAWSII cells were maintained in a humidified atmosphere at 37°C and
5% CO2 and passaged twice a week.
B78-H1 cells, an amelanotic clone of the murine
melanoma B16 cell line, were primarily provided by Dr L.H. Graf
(Chicago, IL, USA) and were found to be suitable in our previous
immunotherapeutic models (17,18).
The cells were cultured in Dulbecco’s modified Eagle’s medium
(DMEM, Sigma-Aldrich) supplemented with 10% inactivated FCS and
antibiotics. Other culturing conditions were similar to those used
for JAWSII cells.
For in vivo experiments, B78-H1 cells were
trypsinized, washed twice in ice-cold PBS and resuspended in PBS at
the concentration of 2×105/20 μl per mice (or in some
experiments 5×105/20 μl per mice). Cell viability was
determined prior to the inoculation using trypan blue exclusion
test and estimated to be >90–95%.
In vitro JAWSII stimulatory
conditions
In the present study, we used a protocol of
stimulation of JAWSII cells that was previously described (16). In brief, JAWSII cells were pulsed
with tumor cell lysates (JAWSII cells to B78-H1 melanoma cells
ratio = 1:1) for 3 h followed by a 48-h incubation with
polyriboinosinic polyribocytidylic acid (poly I:C sodium salt,
Sigma-Aldrich, concentration in cultures, 100 μg/ml) and interferon
γ (recombinant mouse IFN-γ, BD Pharmingen™, 10 ng/ml in
cultures).
Mice
For in vivo experiments, C57BL/6 mice
(8-week-old, bred in a local animal facility and kept in
conventional conditions) were used. All the experiments were
approved by the Local Ethics Committee.
Flow cytometric analysis of stimulated
JAWSII cell phenotype
The following monoclonal antibodies, previously
described (16), were used to study
surface markers of JAWSII cells: anti-MHC class I, anti-MHC class
II, anti-CD11c, anti-CD40, anti-CD80, anti-CD86, anti-CCR7, and
anti-CD8α; relevant isotype controls were used. Anti-CD80 and
anti-CD86 double staining was performed so as to determine
simultaneous expression of both CD80 and CD86. Surface markers were
analyzed after pulsing JAWSII cells with tumor-cell lysates and
incubation with appropriate immunostimulators at concentrations
described above. The cells were then collected, resuspended in PBS
with 0.5% BSA and 0.05% sodium azide (0.5 million cells in 50 μl),
incubated for 30 min at 4°C with appropriate mAbs, and analyzed in
FACS Scan (Becton-Dickinson). The level of surface marker
expression, estimated by the mean fluorescence intensity (MFI), was
analyzed using CellQuest software.
Phagocytosis assay
JAWSII cells were stained with 1 mM DiI for 10 min
at 37°C and washed two times with ice-cold PBS and seeded in
12-well plates. The following day, JAWSII cells were stimulated
with 10 ng/ml IFN-γ or/and 100 μg/ml poly I:C. Twenty-four hours
later, B78-H1 melanoma cells were stained with 5 μM CFSE for 10 min
at 37°C, washed two times with ice-cold PBS, and placed in a 25
cm2 culture flask. After 24 h, B78-H1 cells were lysed
by the freeze/thaw method, added in a volume of 100 μl and
cocultured with JAWSII cells for an additional 3 h at 37°C and 5%
CO2 or at 4°C. Then, the cells were collected, washed
and resuspended in 300 μl of PBS. A total of 10,000 cells were
analyzed on a FACS Scan (Becton-Dickinson) using CellQuest Pro
Software Version 5.2. Phagocytosis level was measured as a
percentage of double positive cells.
IL-12
Recombinant mouse IL-12 (specific activity
4.6/106 U/mg protein) was a generous gift from the
Genetics Institute (Cambridge, MA, USA) (18). For in vivo experiments, the
cytokine was diluted with 0.1% BSA (Sigma Chemicals, St. Louis, MO,
USA).
In vivo protocols used in the B78-H1
melanoma model
Three variants of experiments with JAWSII cells were
performed: a) prophylactic, b) therapeutic - intratumoral, and c)
therapeutic - systemic schemes.
In the prophylactic scheme, randomly selected mice
were injected into the footpad of the left hind limb with
stimulated JAWSII on Day −7. On days: −6, −5 and −4 mice were
administered with IL-12 or diluent (0.1% BSA in PBS). On Day 0, all
the mice were inoculated with B78-H1 melanoma cells in the right
hind limb.
In the intratumoral therapeutic scheme, all the mice
were inoculated with B78-H1 melanoma cells into the footpad of the
right hind limb. Next, on Day +3, mice were injected into the same
site with stimulated JAWSII cells. On Days +4, +5 and +6, mice were
injected with IL-12 or diluent (0.1% BSA in PBS).
In the systemic therapeutic scheme, mice were
inoculated with B78-H1 melanoma cells into the footpad of the right
hind limb. Next, on Day +3, mice were injected into the
contralateral hind limb with stimulated JAWSII or PBS. On Days: +4,
+5, and +6 mice were treated with IL-12 or diluent (0.1% BSA in
PBS).
Statistical analysis
Results of experiments on FACS are presented on
histograms together with MFI or percentage values. Statistical
analysis of the in vivo experiments was performed with the
Mann-Whitney-Wilcoxon test (tumor diameter in mm measured seven
weeks after tumor cell inoculation) and the log-rank test
(percentage of mice without tumor) using Statistica®
software. For clarity, 3 levels of statistical significance were
used: p<0.05; p<0.01 and p<0.001 that were related to the
control group (unless otherwise indicated).
Results
Expression of surface markers in the
stimulated JAWSII cells
Prior to the in vivo experiments in a murine
melanoma model, JAWSII cells were pulsed with B78-H1 lysates
followed by 48 h of incubation with poly I:C + IFN-γ. Then, the
influence of this treatment on the expression of surface markers of
the cells was determined. In some experiments, FACS analysis of the
stimulated JAWSII cells that were additionally incubated with IL-12
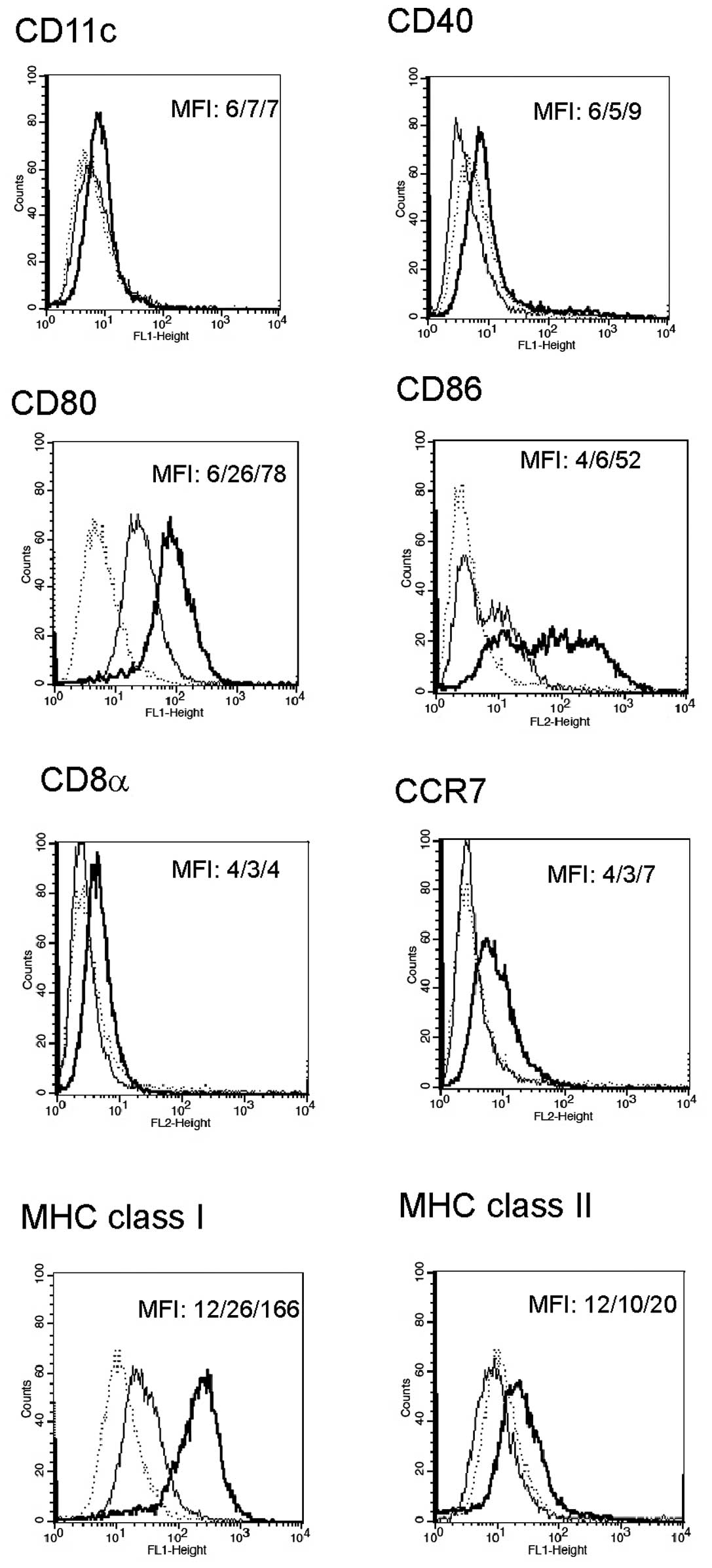
for the following 24 h was performed. As presented in Fig. 1, the cells pulsed with tumor cell
lysates and incubated with poly I:C + IFN-γ were characterized by
high expression of CD80, CD86, and MHC class I molecules. Moreover,
there was a moderate increase in the expression of CD40, CCR7, and
MHC class II molecules in the cells.
A supplementary incubation with IL-12 (for 24 h),
after pulsing with tumor lysates and a 48-h incubation with poly
I:C + IFN-γ, did not result in a change of surface markers when
compared to the cells analyzed without IL-12 (data not shown).
Phagocytosis of the frozen-thawed B78-H1
melanoma cells by JAWSII cells
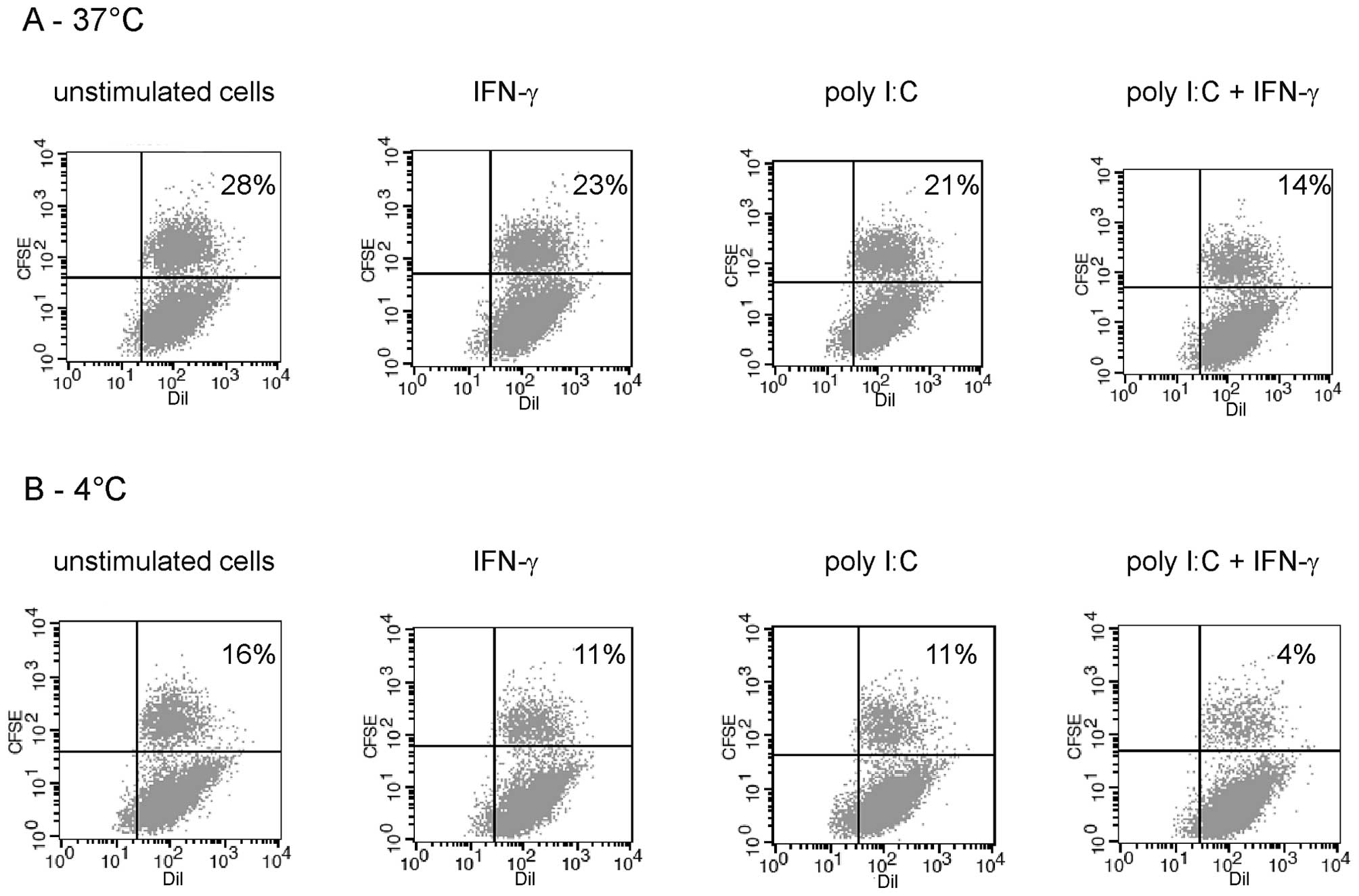
The ability of JAWSII cells to phagocytose B78-H1
lysates was determined in the experiment on FACS with CFSE dye. It
was found that JAWSII cells phagocytosed frozen-thawed B78-H1
melanoma cells effectively. As presented in Fig. 2, the phagocytic capacity of JAWSII
cells was decreased after stimulation with poly I:C (100 μg/ml) and
IFN-γ (10 ng/ml), particularly in combination of these two
agents.
Production and secretion of IL-12 by
JAWSII cells
IL-12 was not present in supernatants from
unstimulated JAWSII cultures and JAWSII lysates, using an ELISA kit
(Mouse IL-12p70 ELISA MAX™ Set Deluxe, Biolegend, Inc., San Diego,
CA, USA) (data not shown). It was also revealed that neither poly
I:C nor IFN-γ, alone or in combination, stimulated secretion of
IL-12 in 24-h JAWSII cultures.
In vivo experiments
Prophylactic scheme
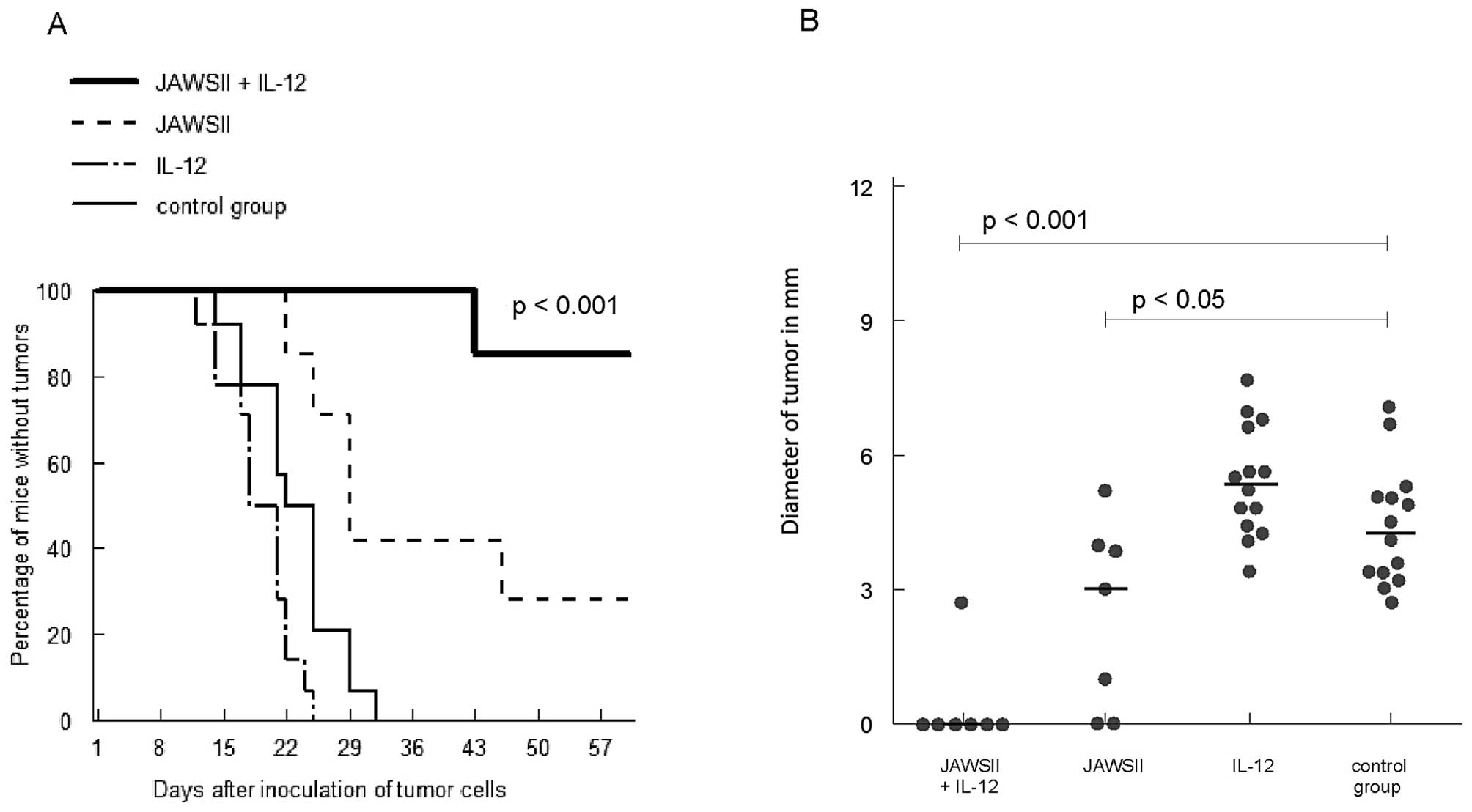
As shown in Fig. 3,
injection of JAWSII cells pulsed with tumor cell lysates and
incubated with poly I:C + IFN-γ followed by the administration of
IL-12 induced effective immunization; no tumor was observed in 86%
of mice at the end of the experiment (p<0.001). Injection with
JAWSII cells alone resulted in some preventive effect; no tumor
development in 29% of cases (p<0.01 vs. control), which was
statistically inferior when compared with the group treated with
JAWSII + IL-12 (p<0.05). Furthermore, smaller diameters of
tumors were observed in that group (p<0.05 vs. control). No
prophylactic effect was demonstrated in the group of mice treated
with IL-12 alone.
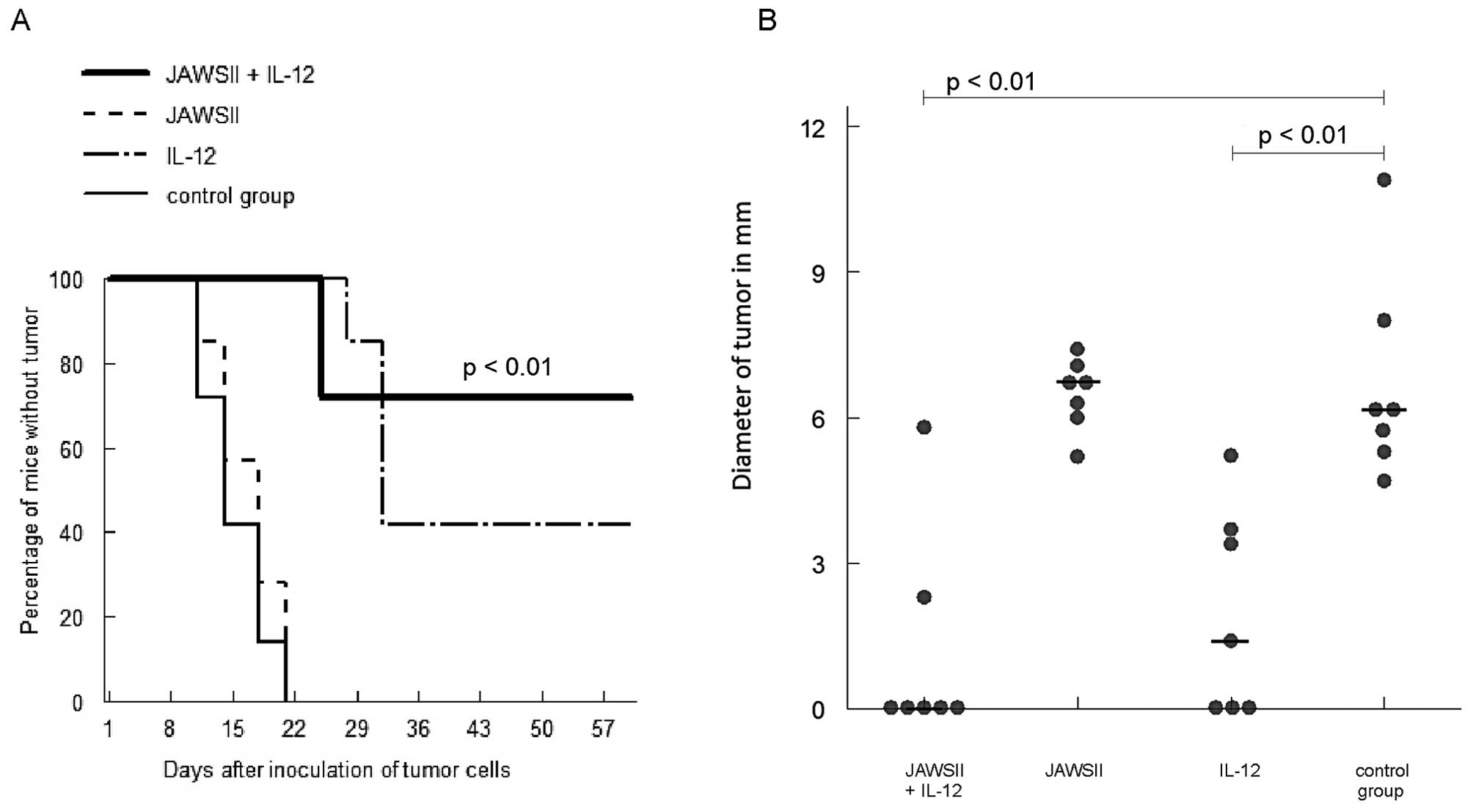
Intratumoral therapeutic scheme
In the experiment, combination of JAWSII cells
pulsed with B78-H1 cell lysates + incubated with poly I:C + IFN-γ
and IL-12, when injected intratumorally, led to tumor eradication
in 71% of cases (p<0.01 vs. control) and significantly smaller
diameters of tumors at the end of the observation period (median =
0, p<0.01 vs. control) (Fig. 4).
On the contrary, in the group of mice treated with JAWSII cells
alone, all the mice developed neoplasms. Injection with IL-12 alone
led to some curative effect (eradication of tumors in 43% of mice)
(p<0.01 vs. control) and a decrease of tumor diameter (p<0.01
vs. control). However, no statistical difference was found between
the group treated with JAWSII cells + IL-12 and the group injected
with IL-12 alone, with regard to the rate of tumor development and
diameter of tumors.
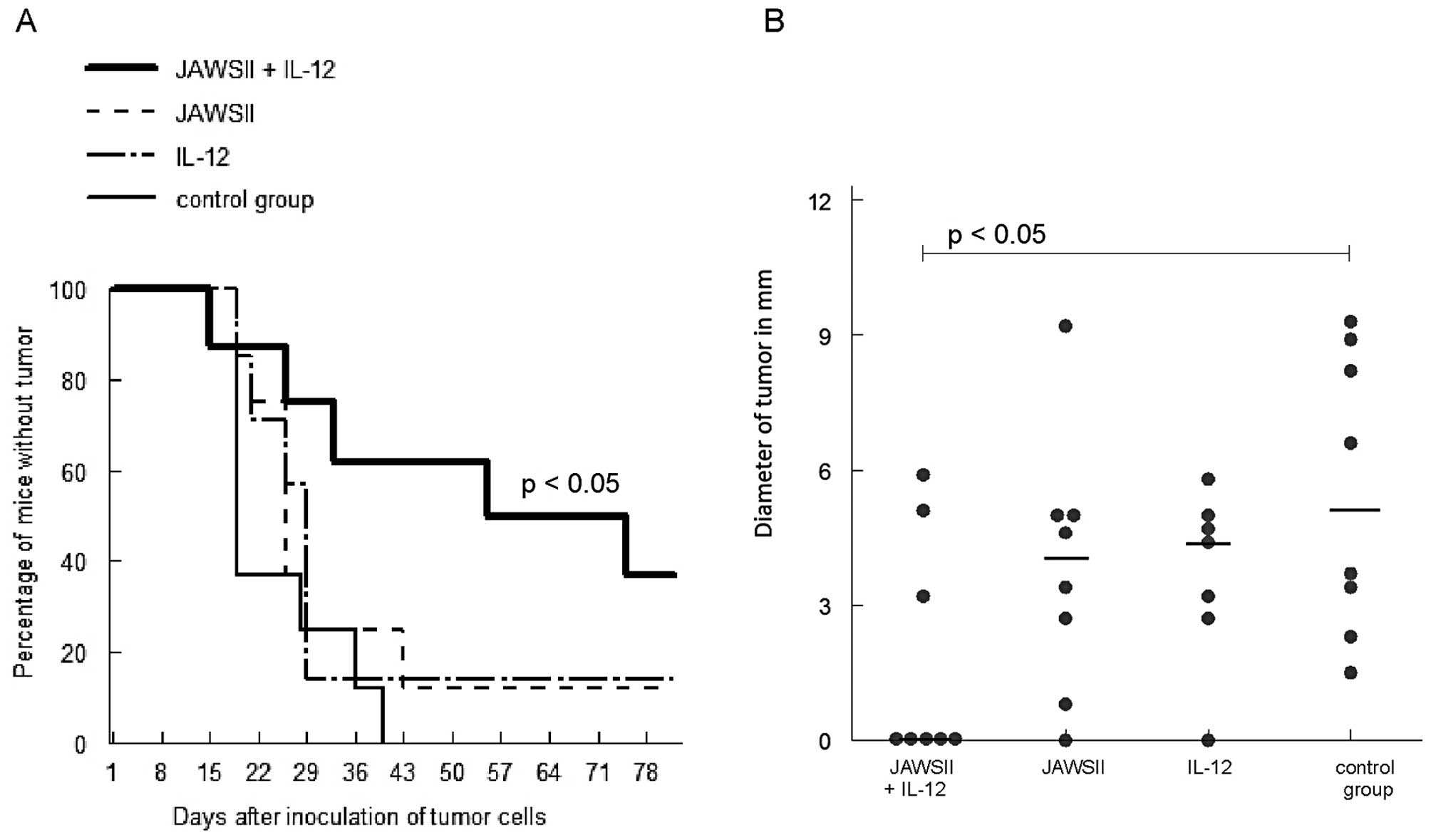
Systemic therapeutic scheme
Results of the experiment on the efficacy of
systemic antitumor therapy consisting of stimulated JAWSII ± IL-12
are presented in Fig. 5. The
combination of JAWSII cells and IL-12 resulted in a significant
rate of tumor eradication; 38% of mice did not develop tumors at
the end of observation period (p<0.05 vs. control), and smaller
diameters of tumors were observed 7 weeks after tumor cell
inoculation (p<0.05 vs. control). Injections with either JAWSII
cells or IL-12 alone did not lead to a significant therapeutic
effect; no statistically significant differences were noted in
comparison with the control group.
Discussion
Among different methods of introduction of antigenic
material into DCs (tumor lysates, peptides, RNA, DNA), the use of
lysates of whole tumor cells is the most frequently described
protocol (5,19,20).
However, it was claimed that DCs pulsed with tumor antigens are
unable to reach maturity and produce insufficient amounts of IL-12
to induce efficacious antitumor response (7). As JAWSII cells did not produce IL-12,
neither unstimulated nor stimulated (poly I:C, IFN-γ alone and in
combination), we decided to introduce injections of IL-12 to in
vivo protocols in order to improve the activity of cellular
vaccine by modification of tumor environment and eliciting
additional antitumoral defense mechanisms. It was found that JAWSII
cells pulsed with tumor lysates and incubated with the combination
of immunostimulators (poly I:C + IFN-γ) revealed high expression of
CD80 and CD86 molecules, and also increased levels of CCR7, and MHC
molecules when compared with the control group (Fig. 1). Both in the previous (16) and in the present study, we found
that incubation of JAWSII cells with poly I:C and IFN-γ increases
expression of CD40, although some authors argue against the
presence of the CD40 molecule in this cell line (21,22).
Markedly, further incubation of the cells with IL-12 did not affect
the expression of the studied surface markers. In the study of
Rossowska et al, pulsing bone marrow-derived DCs with tumor
cell lysates did not alter the levels of the surface markers
(6). However, a slight decrease in
expression of CD86, MHC class I and II molecules was observed when
compared with untreated cells (6).
In our previous study (16), the
decreased ability of endocytosis of JAWSII cells (reflecting more
mature stage of these cells) in cultures incubated with LPS or poly
I:C was described, while in the present study we demonstrated
maturation-inducing potential of combination of poly I:C and IFN-γ.
JAWSII cells are able to phagocytose lysed B78-H1 melanoma cells
efficiently and the combination of immunostimulators diminishes
that ability (Fig. 2). The impact
of appropriate immunostimulators, mostly microbial elements, is
crucial for the ability of DCs to promote anticancer response in
vivo(6,7). They induce changes in the phenotype of
DCs, which start to express high levels of costimulatory and MHC
molecules (6). Furthermore,
stimulated in that manner, semi-mature DCs retain properties for
further maturation in vivo after the contact with the
factors secreted by the host’s immune system, mainly cytokines. As
the immune response is usually suppressed by the tumor,
coadministration of IL-12 with the cellular vaccine may induce
stronger antitumor mechanisms (9).
In the prophylactic scheme of therapy in our murine
melanoma model, JAWSII cells pulsed with tumor lysates and
incubated with poly I:C + IFN-γ demonstrated significant antitumor
effect, which was augmented in the group of mice treated
additionally with IL-12 (Fig. 3).
It should be noted that the cytokine alone was found to be
ineffective. By contrast, in the intratumoral therapeutic scheme,
injections of stimulated JAWSII cells alone showed no antitumor
effects while administration of IL-12 induced eradication of tumors
in some mice (Fig. 4). The best
treatment option was, as in the prophylactic protocol, application
of JAWSII cells in combination with IL-12; most mice were cured.
Despite induction of similar antitumor effects, it is possible that
different mechanisms operated in the prophylactic versus the
intratumoral therapeutic protocol. As we have shown in our previous
studies, injection of activated and tumor lysate-fed JAWSII cells
was associated with specific induction of effector cytotoxic T
lymphocytes in regional lymph nodes (16). It is quite probable that
supplementation of IL-12 enhanced this specific response in the
prophylactic scheme (9,23). In the intratumoral therapeutic
schedule, non-specific, NK cell-dependent mechanisms were able to
contribute to the overall antitumor effect in the combination
treatment, since IL-12 is the strong activator of these cells
(24,25). The antiangiogenic effect of IL-12
may also be helpful in the eradication of tumors (26,27).
Activation of both specific and non-specific mechanisms of immunity
in the combination therapy (JAWSII cell vaccine + IL-12) was strong
enough to induce, as shown in Fig.
5, antitumor responses in the most challenging systemic
therapeutic scheme, mimicking clinical situation.
IL-12 was used in immunotherapy as a single agent in
a number of models of murine cancers (28). However, it should be stressed that
although some authors described a decrease in the rate of tumor
growth after injection with IL-12, superior effects were noted when
the cytokine was used in combination with other forms of therapy
(12,29,30).
IL-12 may improve stimulating properties of bone marrow-derived DCs
in vitro(31). In the study
of Tatsumi et al, the use of IL-12 in combination with DCs
pulsed with tumor lysates was more effective in comparison with DCs
alone in a murine liver cancer model, leading to an induction of
specific antitumor responses (8).
Furthermore, Fallarino et al observed specific response of
cytotoxic lymphocytes after administration of IL-12 and pulsed DCs
in nearly 100% of mice (9). In one
clinical study, peripheral blood mononuclear cells (PBMCs) loaded
with tumor antigenic peptides and IL-12 induced CD8+
T-cell response (23). However,
methodological differences (using PBMCs in selected
HLA-A2+ patients, lack of IL-12 alone-treated controls)
makes comparison of this study with our model unreliable.
In conclusion, in the present study we presented
antitumor effects of the JAWSII cell-based cellular vaccine used in
combination with IL-12 in a murine model of B78-H1 melanoma. The
findings of this study may aid in the planning optimal DC-based
therapeutic protocols in cancer patients.
Acknowledgements
This study was supported by the Medical University
of Warsaw, Poland (grant nos. 1M19/W2, 1M19/NM5/07) and by the
Ministry of Science and Higher Education, Poland (grant no. N N401
011536).
References
|
1
|
Ueno H, Klechevsky E, Morita R, et al:
Dendritic cell subsets in health and disease. Immunol Rev.
219:118–142. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Palucka K, Ueno H, Roberts L, Fay J and
Banchereau J: Dendritic cell subsets as vectors and targets for
improved cancer therapy. Curr Top Microbiol Immunol. 344:173–192.
2012.PubMed/NCBI
|
|
3
|
Gilboa E: DC-based cancer vaccines. J Clin
Invest. 117:1195–1203. 2007. View
Article : Google Scholar
|
|
4
|
Pajtasz-Piasecka E and Indrova M:
Dendritic cell-based vaccines for the therapy of experimental
tumors. Immunotherapy. 2:257–268. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Palucka K, Ueno H and Banchereau J: Recent
developments in cancer vaccines. J Immunol. 186:1325–1331. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rossowska J, Pajtasz-Piasecka E, Szyda A,
Krawczenko A, Zietara N and Dus D: Tumour antigen-loaded mouse
dendritic cells maturing in the presence of inflammatory cytokines
are potent activators of immune response in vitro but not
in vivo. Oncol Rep. 21:1539–1549. 2009.PubMed/NCBI
|
|
7
|
Vegh Z and Mazumder A: Generation of tumor
cell lysate-loaded dendritic cells preprogrammed for IL-12
production and augmented T cell response. Cancer Immunol
Immunother. 52:67–79. 2003.PubMed/NCBI
|
|
8
|
Tatsumi T, Takehara T, Kanto T, et al:
Administration of interleukin-12 enhances the therapeutic efficacy
of dendritic cell-based tumor vaccines in mouse hepatocellular
carcinoma. Cancer Res. 61:7563–7567. 2001.PubMed/NCBI
|
|
9
|
Fallarino F, Uyttenhove C, Boon T and
Gajewski TF: Improved efficacy of dendritic cell vaccines and
successful immunization with tumor antigen peptide-pulsed
peripheral blood mononuclear cells by coadministration of
recombinant murine interleukin-12. Int J Cancer. 80:324–333. 1999.
View Article : Google Scholar
|
|
10
|
Gately MK, Warrier RR, Honasoge S, et al:
Administration of recombinant IL-12 to normal mice enhances
cytolytic lymphocyte activity and induces production of IFN-gamma
in vivo. Int Immunol. 6:157–167. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
McKnight AJ, Zimmer GJ, Fogelman I, Wolf
SF and Abbas AK: Effects of IL-12 on helper T cell-dependent immune
responses in vivo. J Immunol. 152:2172–2179. 1994.PubMed/NCBI
|
|
12
|
Brunda MJ, Luistro L, Rumennik L, et al:
Antitumor activity of interleukin 12 in preclinical models. Cancer
Chemother Pharmacol. 38(Suppl): S16–S21. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nastala CL, Edington HD, McKinney TG, et
al: Recombinant IL-12 administration induces tumor regression in
association with IFN-gamma production. J Immunol. 153:1697–1706.
1994.PubMed/NCBI
|
|
14
|
Pajtasz-Piasecka E, Rossowska J, Szyda A,
Krawczenko A and Dus D: Generation of anti-tumor response by JAWS
II mouse dendritic cells transduced with murine interleukin 12
genes. Oncol Rep. 17:1249–1257. 2007.PubMed/NCBI
|
|
15
|
Xu Y, Darcy PK and Kershaw MH:
Tumor-specific dendritic cells generated by genetic redirection of
Toll-like receptor signaling against the tumor-associated antigen,
erbB2. Cancer Gene Ther. 14:773–780. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zapala L, Drela N, Bil J, Nowis D, Basak
GW and Lasek W: Optimization of activation requirements of immature
mouse dendritic JAWSII cells for in vivo application. Oncol
Rep. 25:831–840. 2011.PubMed/NCBI
|
|
17
|
Basak GW, Zapala L, Wysocki PJ, Mackiewicz
A, Jakobisiak M and Lasek W: Interleukin 15 augments antitumor
activity of cytokine gene-modified melanoma cell vaccines in a
murine model. Oncol Rep. 19:1173–1179. 2008.PubMed/NCBI
|
|
18
|
Switaj T, Jalili A, Jakubowska AB, et al:
CpG immunostimulatory oligodeoxynucleotide 1826 enhances antitumor
effect of interleukin 12 gene-modified tumor vaccine in a melanoma
model in mice. Clin Cancer Res. 10:4165–4175. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ballestrero A, Boy D, Moran E, Cirmena G,
Brossart P and Nencioni A: Immunotherapy with dendritic cells for
cancer. Adv Drug Deliv Rev. 60:173–183. 2008. View Article : Google Scholar
|
|
20
|
Palucka K and Banchereau J: Cancer
immunotherapy via dendritic cells. Nat Rev Cancer. 12:265–277.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Haase C, Michelsen BK and Jorgensen TN:
CD40 is necessary for activation of naive T cells by a dendritic
cell line in vivo but not in vitro. Scand J Immunol. 59:237–245.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jorgensen TN, Haase C and Michelsen BK:
Treatment of an immortalized APC cell line with both cytokines and
LPS ensures effective T-cell activation in vitro. Scand J Immunol.
56:492–503. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Peterson AC, Harlin H and Gajewski TF:
Immunization with Melan-A peptide-pulsed peripheral blood
mononuclear cells plus recombinant human interleukin-12 induces
clinical activity and T-cell responses in advanced melanoma. J Clin
Oncol. 21:2342–2348. 2003. View Article : Google Scholar
|
|
24
|
Wehner R, Dietze K, Bachmann M and Schmitz
M: The bidirectional crosstalk between human dendritic cells and
natural killer cells. J Innate Immun. 3:258–263. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Manetti R, Parronchi P, Giudizi MG, et al:
Natural killer cell stimulatory factor [interleukin 12 (IL-12)]
induces T helper type 1 (Th1)-specific immune responses and
inhibits the development of IL-4-producing Th cells. J Exp Med.
177:1199–1204. 1993.
|
|
26
|
Bielawska-Pohl A, Blesson S, Benlalam H,
et al: The anti-angiogenic activity of IL-12 is increased in
iNOS−/− mice and involves NK cells. J Mol Med.
88:775–784. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Airoldi I, Di Carlo E, Cocco C, et al:
Endogenous IL-12 triggers an antiangiogenic program in melanoma
cells. Proc Natl Acad Sci USA. 104:3996–4001. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hill HC, Conway TF Jr, Sabel MS, et al:
Cancer immunotherapy with interleukin 12 and granulocyte-macrophage
colony-stimulating factor-encapsulated microspheres: coinduction of
innate and adaptive antitumor immunity and cure of disseminated
disease. Cancer Res. 62:7254–7263. 2002.
|
|
29
|
Kozar K, Kaminski R, Switaj T, et al:
Interleukin 12-based immunotherapy improves the antitumor
effectiveness of a low-dose 5-Aza-2′-deoxycitidine treatment in
L1210 leukemia and B16F10 melanoma models in mice. Clin Cancer Res.
9:3124–3133. 2003.PubMed/NCBI
|
|
30
|
Grohmann U, Bianchi R, Ayroldi E, et al: A
tumor-associated and self antigen peptide presented by dendritic
cells may induce T cell anergy in vivo, but IL-12 can prevent or
revert the anergic state. J Immunol. 158:3593–3602. 1997.PubMed/NCBI
|
|
31
|
Kelleher P and Knight SC: IL-12 increases
CD80 expression and the stimulatory capacity of bone marrow-derived
dendritic cells. Int Immunol. 10:749–755. 1998. View Article : Google Scholar : PubMed/NCBI
|