Introduction
Active immunotherapy represents a promising modality
for the treatment of malignant diseases. However, due to low
clinical response rates cancer vaccination with the use of
antigen-presenting cells (APCs) faced substantial skepticism some
years ago (1). Meanwhile, growing
body of knowledge on cancer immunosurveilance, and loss thereof,
led to a refinement of immunotherapeutic strategies (2). Particularly for cancer vaccinations
this has been further encouraged by recent progress in this field
with several successful trials which even resulted in the first
approval for a cellular vaccine by the US FDA (3). Nevertheless, further progress will
depend on the ability to circumvent the different tumor escape
mechanisms (4–6), specifically tumor-induced
immunosuppression which represents one of the major barriers to
successful tumor immunotherapy.
Prostaglandin E2 (PGE2)
belongs to the eicosanoid family of lipid mediators and is a potent
immunomodulator. PGE2 and its receptors play a role in a
broad range of physiologic processes and have been implied in a
number of pathologic conditions such as inflammatory disease,
infections and cancer (7–9). It has diverse and often opposing
effects on the immune system. In cancer PGE2 has been
identified as one of the major soluble tumor-derived factors
contributing to the immunosuppressive tumor environment (10). Many tumors exhibit increased
expression of cyclooxygenase-2 (COX-2) subsequently leading to an
increased production of PGE2. The suppressive effects
are mediated by inhibition of the production of pro-inflammatory
cytokines, by upregulation of the expression of immunosuppressive
cytokines and by inhibiting the function of important immune
effector cells such as T cells, natural killer cells and APCs
(11–14). Further evidence for the importance
of PGE2 in mediating immunosuppression stems from
experiments in which inhibition of COX-2 by non-steroidal
anti-inflammatory drugs resulted in an enhanced antitumor immune
response (15).
The complex functions of PGE2 are also
reflected in the debate about its use for the maturation of
monocyte derived DCs (moDCs). PGE2 is included in most
maturation cocktails as it has been shown to upregulate the
expression of CCR7 which is essential for the migration to
secondary lymph organs (16,17).
On the other hand PGE2 seems to inhibit the
differentiation and immunostimulatory function of moDCs by the
upregulation of IDO expression (18,19)
and by suppressing DC mediated attraction of naïve T cell (20).
In recent years B cells are increasingly recognized
as important APCs capable of inducing antigen-specific
CD4+ and CD8+ T cell responses under
physiologic and pathologic conditions (21,22).
CD40-activated B cells are currently being studied as an
alternative type of APC for cellular vaccines (23–26).
Most importantly, they can be easily expanded ex vivo from
peripheral blood of cancer patients (27). However, in contrast to DCs there is
little knowledge on the regulation of antigen presentation by B
cells. This is also true with regard to the influence of
PGE2. We therefore studied the effects of
PGE2 on key factors for the induction of an immune
response by APCs such as the expression of costimulatory molecules,
migratory potential to secondary lymphoid organs and finally the
induction of T cell activation and proliferation.
Materials and methods
Preparation of human CD40-activated B
cells
Human CD40-activated B cells were prepared as
described previously (28).
Briefly, whole PBMC were cultured on irradiated NIH3T3 cells
transfected with CD154 (tCD40L) in the presence of recombinant
human interleukin 4 (rhIL-4; 2 ng/ml; R&D Systems, Minneapolis,
MN, USA) and clinical-grade cyclosporin A (CsA; 5.5×10−7
M; Novartis, Basel, Switzerland) in Iscove’s modified Dulbecco’s
medium (IMDM; Invitrogen, Karlsruhe, Germany) supplemented with 10%
pooled human serum. The cells were recultured every 3–4 days. After
3 weeks CD40-activated B cells were used for experiments. The
CD40-activated B cells were cultured in the presence of
PGE2 (Sigma-Aldrich, St. Louis, MO, USA) or vehicle. Of
note, the inhibitory biological activity of PGE2 was
confirmed at different concentrations in T cell proliferation
assays of T cells activated by magnetic beads coated with
anti-CD3/anti-CD28 monoclonal antibodies as previously described
(29,30). After 3 days CD40-B cells were
harvested and used for flow cytometric analysis, and functional
assays. To obtain antigen-presenting cells free of PGE2
the cells were washed extensively prior to their use in functional
assays.
Flow cytometry
Immunophenotypic analyses were performed using
fluorescence-activated cell sorting (FACS). Cells were analyzed for
the expression of CD19, CD25, CD80, CD86, HLA-DR (BD Pharmingen,
Heidelberg, Germany), CCR7, CXCR4 (R&D Systems) and EP2 and EP4
(Cayman Chemical, Ann Arbor, MI, USA) using a FACSCanto flow
cytometer (Becton-Dickinson).
Chemotaxis assay
To assess B cell migration, 5×105
CD40-activated B cells were transferred into the upper chamber of
5-μm pore size transwell plates (Costar, Cambridge, MA, USA).
Varying amounts of the chemokines SDF-1α and SLC (R&D Systems)
were added to the lower chamber. After 2 h at 37°C, the number of
cells that had migrated into the lower chamber was determined using
a hemacytometer.
Allogeneic mixed lymphocyte reaction
CD4+ T cells were obtained from buffy
coats by negative selection using Rosette Sep® human
CD4+ T cell enrichment cocktail (StemCell Technologies,
Vancouver, Canada) according manufacturer’s instructions. Prior to
allogeneic mixed lymphocyte reaction (MLR) CD4+ T cells
were labeled with carboxyfluorescein succinimidyl ester (CFSE,
Molecular Probes, Eugene, OR, USA) according to standard protocols.
A total of 1×105 CFSE-labeled CD4+ T cells
were co-incubated with allogeneic CD40-activated B cells as
stimulators at various B to T-cell ratios ranging from 1:1 to 1:10.
After 5–7 days proliferation was assessed by flow cytometry.
Results
Phenotype of PGE2-treated
CD40-activated B cells
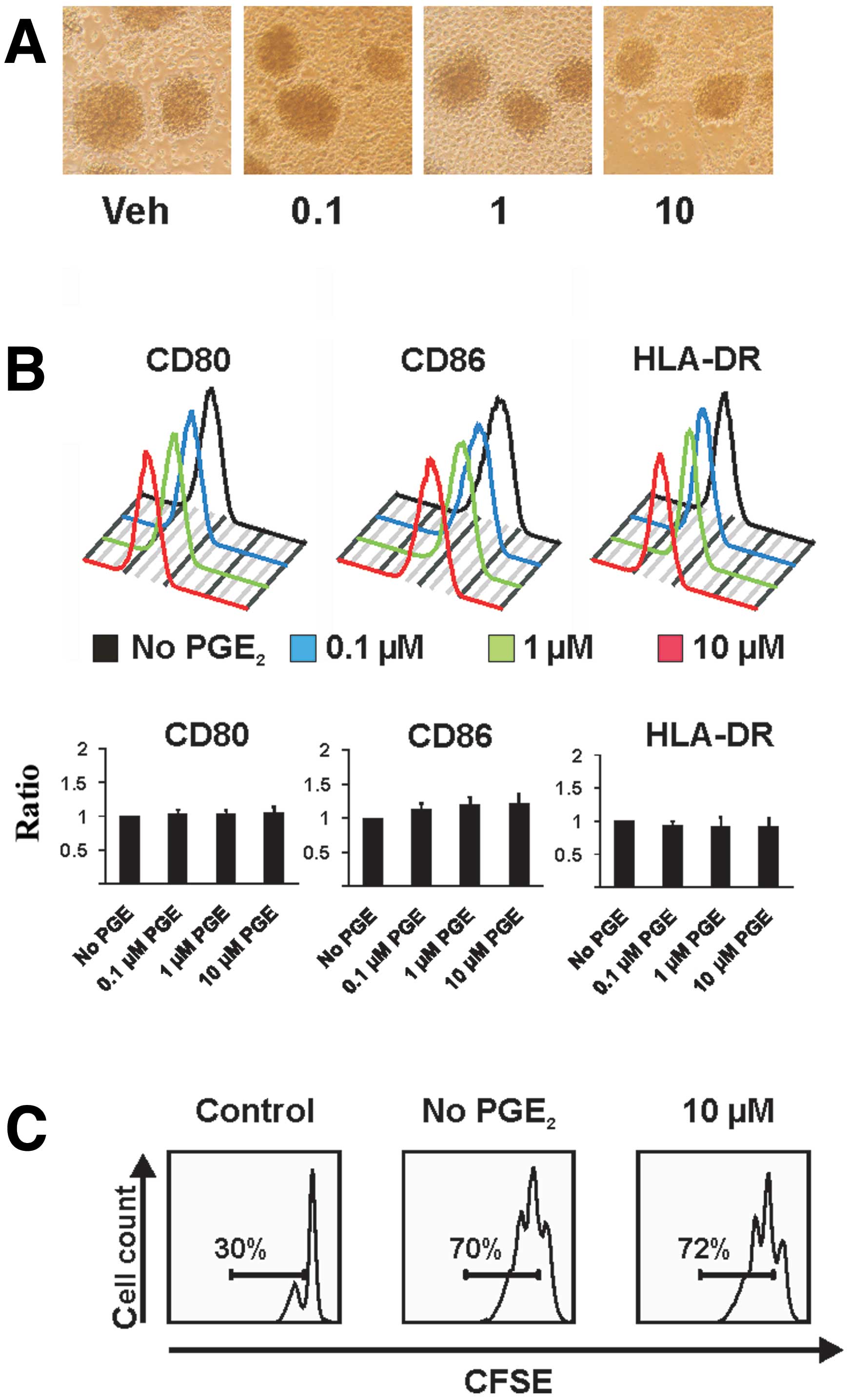
First, we assessed the effect of different
concentrations of PGE2 on the phenotype of
CD40-activated B cells. Morphology and cell surface expression of
costimulatory molecules CD80 and CD86 as well as major
histocompatibility complex (MHC) class II of treated cells were
compared to untreated CD40-activated B cells.
PGE2-treated cells were of normal morphology and formed
round clusters through homotypic adhesion (Fig. 1A). Also, the surface expression of
CD80, CD86 and HLA-DR was not significantly affected by the
exposure to PGE2 in a series of experiments (Fig. 1B). Next, we investigated the
potential influence of PGE2 on the proliferation of
CD40-activated B cells: the continuous proliferation of
CD40-activated B cells was likewise not affected by exposure to
PGE2 (Fig. 1C).
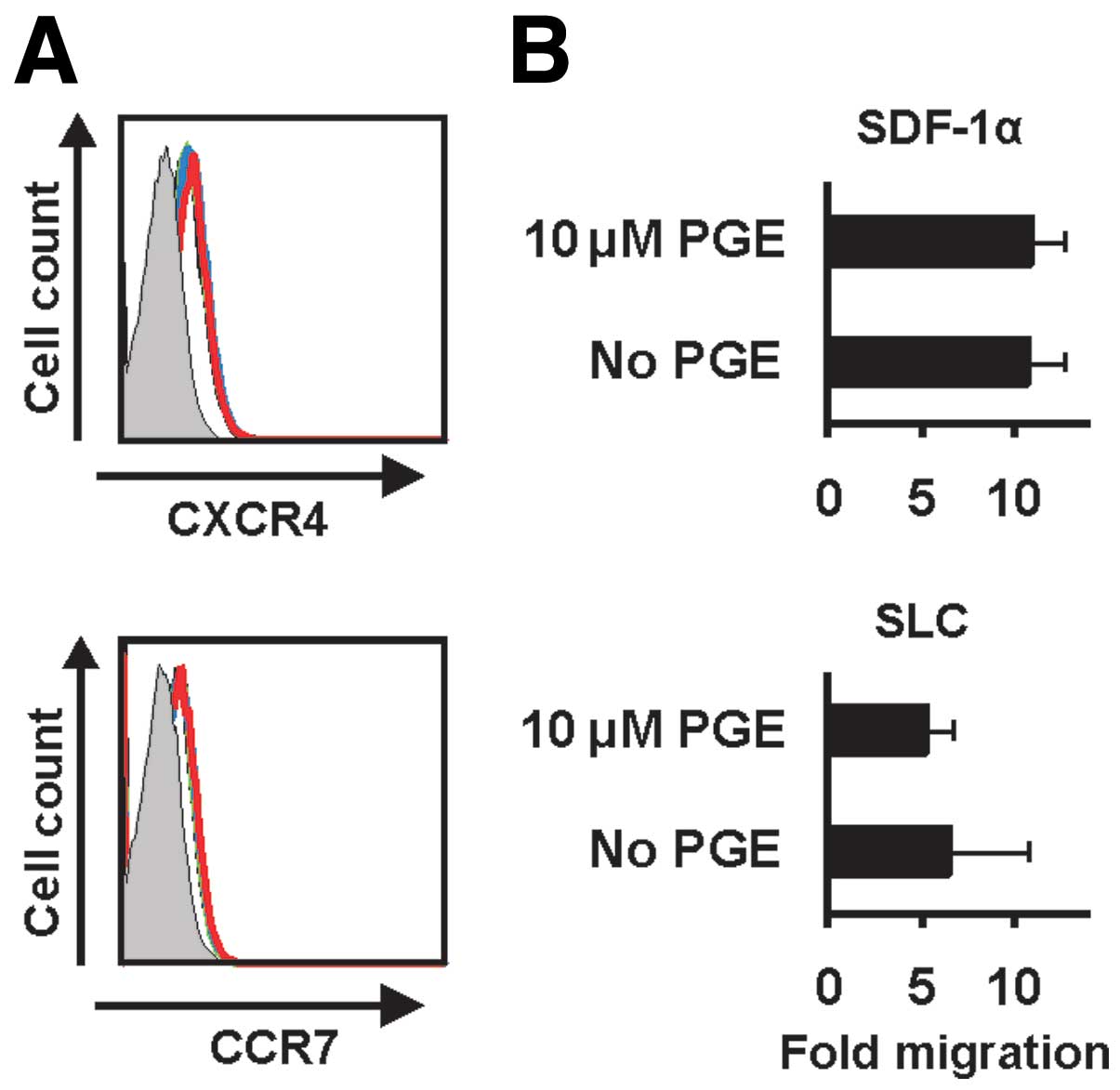
Expression of chemokine receptors CXCR4
and CCR7 as well as migration to their ligands are not affected by
PGE2
The migration of activated APCs to the secondary
lymphoid organs is a crucial step in the induction of an immune
response. The structure of the secondary lymphoid organs provides
the ideal microenvironment for the interaction of antigen-specific
T cells with APCs. Mainly, this is driven by chemokines and their
receptors. We therefore addressed the expression and function of
relevant chemokine receptors which are involved in the migration of
APCs to secondary lymph organs. Treatment with PGE2 did
not affect the expression of CXCR4 and CCR7, the receptors for
SDF-1α and SLC, respectively (Fig.
2A). We next, assessed the migration of CD40-activated B cells
with an in vitro migration assay to test whether the
function of these chemokine receptors are influenced by
PGE2. As demonstrated in Fig. 2B the migration of CD40-activated B
cells to SDF-1α and SLC was not affected by PGE2.
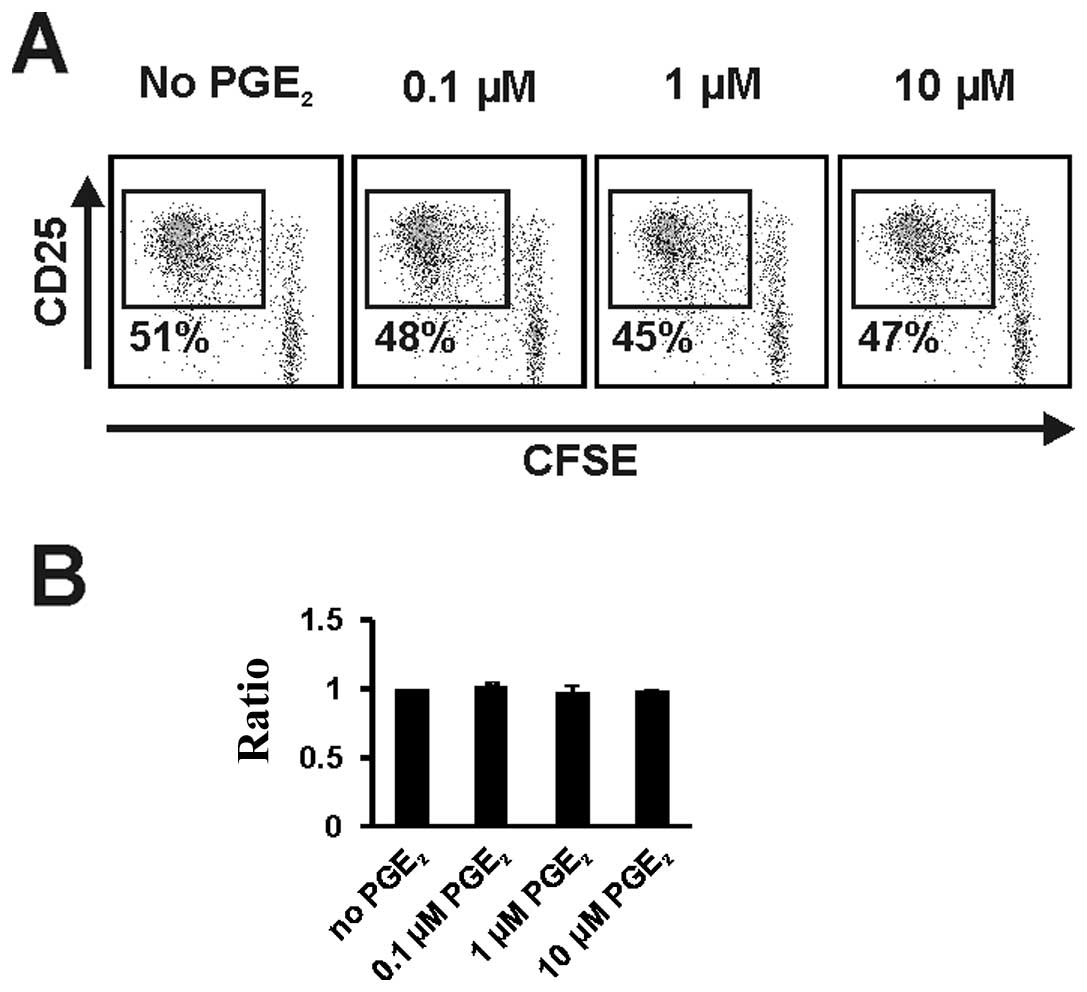
Immunostimulatory function of
CD40-activated B cells with exposure to PGE2
To examine the effects of PGE2 on the
immunostimulatory function of CD40-activated B cells we performed
allogeneic MLRs with purified CD4+ T cells. Changes in
activation and proliferation of T cells were assessed by tracing
expression of CD25 and proliferation of CFSE-labeled T-cells from
Day 5 to Day 7. No significant changes in T cell activation and
proliferation were detectable when PGE2-treated
CD40-activated B cells were used as stimulators (Fig. 3A). Proliferation difference of
T-cells were investigated in a series of experiments and also
displayed by the ratio of proliferating T cells after co-culture
with CD40-activated B cells untreated or treated with the indicated
concentration of PGE2. We observed no significant
changes in T cell proliferation between untreated CD40-activated B
cells and CD40-activated B cells exposed to different
concentrations of PGE2. Even at high doses
PGE2 had no effect on the stimulatory capacity of
CD40-activated B cells (Fig.
3B).
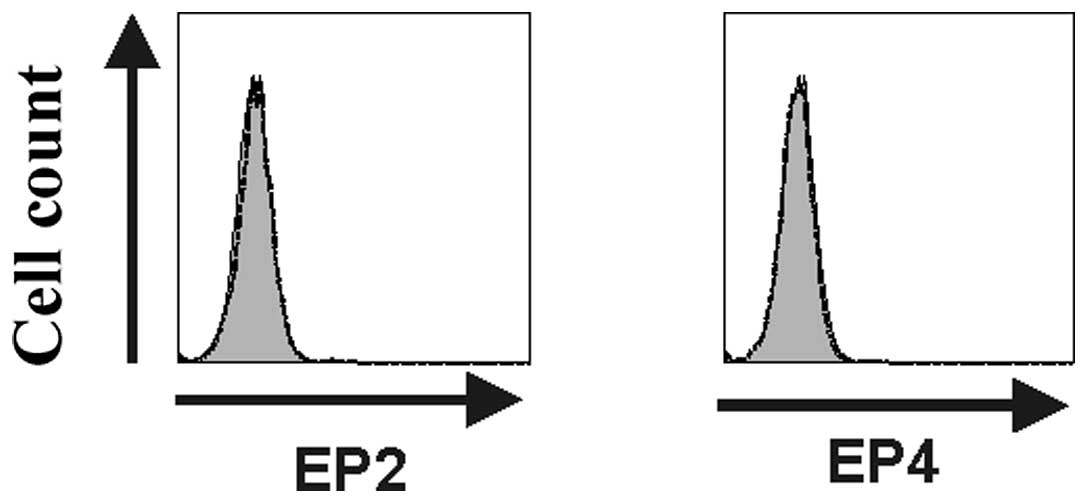
CD40-activated B cells do not express the
PGE2 receptors EP2 and EP4
It has previously been shown that the
immunosuppressive effects of PGE2 are mediated through
the EP2 and EP4 receptors. We thus studied the expression of these
two receptors in CD40-activated B cells by flow cytometry analyses.
Fig. 4 shows representative flow
cytometry plots (n=3) demonstrating the absence of EP2 and EP4
expression on the cell surface of CD40-activated B cells. The lack
of EP2 and EP4 receptor expression might explain the inherent
resistance of CD40-activated B cells to PGE2-mediated
immunosuppression.
Discussion
PGE2-mediated immunosuppression is a
major barrier to the induction of antitumor immune responses and
therefore has an important impact on the design of tumor
vaccination strategies. One important mechanism by which
tumor-derived PGE2 suppresses immune responses is the
induction of APC dysfunction in vivo resulting in an
inhibition of T cell responses against the tumor. This has been
demonstrated for human epithelial cancers such as head and neck and
cervical cancer (31,32). Several mechanisms by which
PGE2-induced tolerogenic DCs prevent tumoricidal immune
reactions have been discovered: they promote T helper type 2 (Th2)
instead of T helper type 1 (Th1) responses and they attract
regulatory T cells (33). Beside
these in vivo effects the use of PGE2 for DC
maturation in vitro is currently under debate. It has been
included as a major component in most moDC maturation cocktails
since it enhances DC maturation and the expression of CCR7 which is
crucial for the lymph node homing. However, it has been shown that
PGE2 induces the expression of indoleamine
2,3-dioxygenase (IDO) in moDCs which results in an inhibition of T
cells (18,19). Nevertheless, this drawback is still
discussed controversially as other authors stated that at least T
cell stimulation is not affected by PGE2 albeit an
increased IDO expression (34).
Scarce clinical trial data exist which addresses this question. At
least, a small DC-based cancer vaccine trial in melanoma raised the
concern that use of PGE2 for the maturation of DCs might
be detrimental. Following vaccination an accumulation of
IDO-expressing DCs and regulatory T cells was observed at the
injection site. All patients had a rapid progressive disease and a
short overall survival (35).
We and others have shown that B cells activated
in vitro by CD40 are potent APCs which could be used for
clinical cancer vaccination trials. Main advantages are their easy
and apparently unlimited proliferation capacities (26,28).
With regard to the growing knowledge on the adverse
immunoregulatory effects of PGE2 on other APCs,
especially DCs, it was the aim of this work to investigate the
influence of PGE2 on the phenotype and the functional
properties of CD40-activated B cells.
In order to increase their antigen-presenting
functions B cells have to be stimulated by inflammatory mediators,
such as pathogen associated molecular patterns. Following
activation they upregulate the expression of MHC and costimulatory
molecules and undergo clonal proliferation. Upon activation B cells
like DCs upregulate the expression of several chemokine receptors
(e.g., CCR7, CXCR4, CD62L), which enable them to enter secondary
lymphoid organs (25). This step is
essential to enable the complex interactions between immune cells
that are required for the induction of an effective immune response
(36). Among these chemokine
receptors CCR7 and CXCR4 have been identified to be crucial in
controlling the migration of DCs to lymph nodes (37,38).
We did not find any changes in the expression of
costimulatory and MHC-II molecules after exposure to
PGE2 at different concentrations. Furthermore, the
proliferative capacity of CD40-activated B cells was unaffected. In
addition, we could not find either a positive or a negative effect
of PGE2 on the expression of CCR7 and CXCR4 on
CD40-activated B cells. In line with these findings, the function
of these receptors, the migration of CD40-activated B cells to the
two important lymph node homing chemokines SLC (ligand for CCR7)
and SDF-1α (ligand for CXCR4) was not altered. CD40-activated B
cells generated from healthy individuals and tumor patients have
been shown to possess the capacity to stimulate T cells (27,39,40).
It is not known though whether these cells are susceptible to
tumor-induced immunosuppression. Importantly, we could exclude any
inhibitory effect of PGE2 on T cell activation and
proliferation induced by CD40-activated B cells suggesting that
CD40-activated B cells would not be affected by tumor-derived PGE2
in vivo. PGE2 exerts its effects through the four
PGE2 receptors EP1-4. At least in DCs the
immunosuppressive effects of PGE2 seem to be exclusively
mediated by the receptors EP2 and EP4 (41,42).
The finding that EP2 and EP4 are absent on CD40-activated B cells
provides a mechanistic explanation for their inherent resistance to
the inhibitory effects of PGE2. These data were further
substantiated by gene expression analyses showing no differential
EP2 and EP4 gene expression of CD40-activated B cells compared to
unstimulated B cells (data not shown). These findings are of
special interest as it has been shown both for mouse and human B
cells that under defined activation B cells express EP2 and EP4
receptors (43,44). Thus, it has to be addressed by
further investigations whether the utilized method to generate
CD40-activated B cells precludes the expression of EP2 and EP4.
Taken together, our data demonstrate that the
immunostimulatory function of CD40-activated B cells is not
affected by PGE2. These results have important
implications for the potentially clinical application of B cells
for immunotherapy. The use of B cells as APCs either by targeting
antigen to B cells in vivo or by ex vivo peptide
pulsing of activated B cells seems to be a promising strategy
especially in settings where PGE2 might prevent the
induction of an immune response.
Acknowledgements
This study was supported by grants from the Deutsche
Krebshilfe. We would like to thank Anne Fiedler for expert
technical assistance.
References
|
1
|
Rosenberg S, Yang J and Restifo N: Cancer
immunotherapy: moving beyond current vaccines. Nat Med. 10:909–915.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Finn OJ: Cancer immunology. N Engl J Med.
358:2704–2715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kantoff PW, Higano CS, Shore ND, et al:
Sipuleucel-T immunotherapy for castration-resistant prostate
cancer. N Engl J Med. 363:411–422. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Palucka K, Ueno H and Banchereau J: Recent
developments in cancer vaccines. J Immunol. 186:1325–1331. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Copier J, Dalgleish AG, Britten CM, et al:
Improving the efficacy of cancer immunotherapy. Eur J Cancer.
45:1424–1431. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zou W: Regulatory T cells, tumour immunity
and immunotherapy. Nat Rev Immunol. 6:295–307. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kalinski P: Regulation of immune responses
by prostaglandin E2. J Immunol. 188:21–28. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsuoka T and Narumiya S: The roles of
prostanoids in infection and sickness behaviors. J Infect
Chemother. 14:270–278. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sheng KC, Wright MD and Apostolopoulos V:
Inflammatory mediators hold the key to dendritic cell suppression
and tumor progression. Curr Med Chem. 18:5507–5518. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sombroek CC, Stam AG, Masterson AJ, et al:
Prostanoids play a major role in the primary tumor-induced
inhibition of dendritic cell differentiation. J Immunol.
168:4333–4343. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kolenko V, Rayman P, Roy B, et al:
Downregulation of JAK3 protein levels in T lymphocytes by
prostaglandin E2 and other cyclic adenosine monophosphate-elevating
agents: impact on interleukin-2 receptor signaling pathway. Blood.
93:2308–2318. 1999.
|
|
12
|
Walker W and Rotondo D: Prostaglandin
E2 is a potent regulator of interleukin-12- and
interleukin-18-induced natural killer cell interferon-γ synthesis.
Immunology. 111:298–305. 2004.
|
|
13
|
Obermajer N, Muthuswamy R, Lesnock J, et
al: Positive feedback between PGE2 and COX2 redirects
the differentiation of human dendritic cells toward stable
myeloid-derived suppressor cells. Blood. 118:5498–5505.
2011.PubMed/NCBI
|
|
14
|
Krishnamoorthy S and Honn KV: Eicosanoids
in tumor progression and metastasis. Subcell Biochem. 49:145–168.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Haas AR, Sun J, Vachani A, et al:
Cyclooxygenase-2 inhibition augments the efficacy of a cancer
vaccine. Clin Cancer Res. 12:214–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luft T, Jefford M, Luetjens P, et al:
Functionally distinct dendritic cell (DC) populations induced by
physiologic stimuli: prostaglandin E(2) regulates the migratory
capacity of specific DC subsets. Blood. 100:1362–1372. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Scandella E, Men Y, Gillessen S, et al:
Prostaglandin E2 is a key factor for CCR7 surface
expression and migration of monocyte-derived dendritic cells.
Blood. 100:1354–1361. 2002.
|
|
18
|
Braun D, Longman RS and Albert ML: A
two-step induction of indoleamine 2,3 dioxygenase (IDO) activity
during dendritic-cell maturation. Blood. 106:2375–2381. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Von Bergwelt-Baildon MS, Popov A, Saric T,
et al: CD25 and indoleamine 2,3-dioxygenase are up-regulated by
prostaglandin E2 and expressed by tumor-associated
dendritic cells in vivo: additional mechanisms of T-cell
inhibition. Blood. 108:228–237. 2006.PubMed/NCBI
|
|
20
|
Muthuswamy R, Mueller-Berghaus J,
Haberkorn U, et al: PGE(2) transiently enhances DC expression of
CCR7 but inhibits the ability of DCs to produce CCL19 and attract
naive T cells. Blood. 116:1454–1459. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Martin F and Chan AC: B cell immunobiology
in disease: evolving concepts from the clinic. Annu Rev Immunol.
24:467–496. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shimabukuro-Vornhagen A, Hallek MJ, Storb
RF, et al: The role of B cells in the pathogenesis of
graft-versus-host disease. Blood. 114:4919–4927. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mason NJ, Coughlin CM, Overley B, et al:
RNA-loaded CD40-activated B cells stimulate antigen-specific T-cell
responses in dogs with spontaneous lymphoma. Gene Ther. 15:955–965.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sorenmo KU, Krick E, Coughlin CM, et al:
CD40-activated B cell cancer vaccine improves second clinical
remission and survival in privately owned dogs with non-Hodgkin’s
lymphoma. PLoS One. 6:e241672011.PubMed/NCBI
|
|
25
|
Von Bergwelt-Baildon M,
Shimabukuro-Vornhagen A, Popov A, et al: CD40-activated B cells
express full lymph node homing triad and induce T-cell chemotaxis:
potential as cellular adjuvants. Blood. 107:2786–2789.
2006.PubMed/NCBI
|
|
26
|
Wiesner M, Zentz C, Mayr C, et al:
Conditional immortalization of human B cells by CD40 ligation. PLoS
One. 3:e14642008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kondo E, Gryschok L, Klein-Gonzalez N, et
al: CD40-activated B cells can be generated in high number and
purity in cancer patients: analysis of immunogenicity and homing
potential. Clin Exp Immunol. 155:249–256. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liebig TM, Fiedler A, Zoghi S, et al:
Generation of human CD40-activated B cells. J Vis Exp. pii.
13732009.PubMed/NCBI
|
|
29
|
Popov A, Driesen J, Abdullah Z, et al:
Infection of myeloid dendritic cells with Listeria
monocytogenes leads to the suppression of T cell function by
multiple inhibitory mechanisms. J Immunol. 181:4976–4988.
2008.PubMed/NCBI
|
|
30
|
Chemnitz JM, Driesen J, Classen S, et al:
Prostaglandin E2 impairs CD4+ T cell
activation by inhibition of lck: implications in Hodgkin’s
lymphoma. Cancer Res. 66:1114–1122. 2006.
|
|
31
|
Bekeredjian-Ding I, Schafer M, Hartmann E,
et al: Tumour-derived prostaglandin E2 and transforming
growth factor-β synergize to inhibit plasmacytoid dendritic
cell-derived interferon-α. Immunology. 128:439–450. 2009.
|
|
32
|
Herfs M, Herman L, Hubert P, et al: High
expression of PGE2 enzymatic pathways in cervical (pre)neoplastic
lesions and functional consequences for antigen-presenting cells.
Cancer Immunol Immunother. 58:603–614. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Muthuswamy R, Urban J, Lee JJ, et al:
Ability of mature dendritic cells to interact with regulatory T
cells is imprinted during maturation. Cancer Res. 68:5972–5978.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Krause P, Singer E, Darley PI, et al:
Prostaglandin E2 is a key factor for monocyte-derived
dendritic cell maturation: enhanced T cell stimulatory capacity
despite IDO. J Leukoc Biol. 82:1106–1114. 2007.
|
|
35
|
Wobser M, Voigt H, Houben R, et al:
Dendritic cell based antitumor vaccination: impact of functional
indoleamine 2,3-dioxygenase expression. Cancer Immunol Immunother.
56:1017–1024. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Forster R, Schubel A, Breitfeld D, et al:
CCR7 coordinates the primary immune response by establishing
functional microenvironments in secondary lymphoid organs. Cell.
99:23–33. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ohl L, Mohaupt M, Czeloth N, et al: CCR7
governs skin dendritic cell migration under inflammatory and
steady-state conditions. Immunity. 21:279–288. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kabashima K, Shiraishi N, Sugita K, et al:
CXCL12-CXCR4 engagement is required for migration of cutaneous
dendritic cells. Am J Pathol. 171:1249–1257. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Schultze JL, Grabbe S and von
Bergwelt-Baildon MS: DCs and CD40-activated B cells: current and
future avenues to cellular cancer immunotherapy. Trends Immunol.
25:659–664. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim SK, Nguyen Pham TN, Nguyen Hoang TM,
et al: Induction of myeloma-specific cytotoxic T lymphocytes ex
vivo by CD40-activated B cells loaded with myeloma tumor antigens.
Ann Hematol. 88:1113–1123. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Harizi H, Grosset C and Gualde N:
Prostaglandin E2 modulates dendritic cell function via
EP2 and EP4 receptor subtypes. J Leukoc Biol.
73:756–763. 2003.
|
|
42
|
Legler DF, Krause P, Scandella E, et al:
Prostaglandin E2 is generally required for human
dendritic cell migration and exerts its effect via EP2 and EP4
receptors. J Immunol. 176:966–973. 2006.PubMed/NCBI
|
|
43
|
Fedyk ER and Phipps RP: Prostaglandin
E2 receptors of the EP2 and EP4
subtypes regulate activation and differentiation of mouse B
lymphocytes to IgE-secreting cells. Proc Natl Acad Sci USA.
93:10978–10983. 1996.PubMed/NCBI
|
|
44
|
Lee H, Trott JS, Haque S, et al: A
cyclooxygenase-2/prostaglandin E2 pathway augments
activation-induced cytosine deaminase expression within replicating
human B cells. J Immunol. 185:5300–5314. 2010.PubMed/NCBI
|