Introduction
Many reports have linked oxidative damage to DNA and
the associated avoidance and repair processes to carcinogenesis,
ageing and neurodegeneration (1).
Cancer incidence increases with age and there is substantial
evidence that oxidative stress plays a role in human ageing and
neurodegeneration. Since base excision repair (BER) is the main
pathway for the repair of oxidative DNA lesions, the relationship
of BER to human ageing and carcinogenesis is of considerable
interest. Several reports have suggested that the accumulation of
unrepaired DNA lesions plays a causal role in mammalian ageing.
Cells from patients with Cockayne syndrome (CS), a rare inherited
photosensitive progeroid syndrome, are defective in the repair of
8-oxoguanine and 7,8-dihydro-8-oxoadenine (8-OxoA) resulting from
ROS, suggesting that disorders with defects in BER can result in
phenotypes displaying premature ageing (2). In mammalian cells, the oxidative DNA
lesions are removed by BER, which is initiated primarily by two
glycosylases, OGG1 and NTH1. Chen et al(3) reported an age-dependent decrease in
OGG1, and Szczesny et al(4)
reported an age-dependent deficiency in the import of OGG1 into
mitochondria. In additional, Cabelof et al(5) reported a lack of inducibility of DNA
polymerase β and AP endonuclease following exposure to ageing mice
as compared to young mice, suggesting an age-related loss in BER
after exposure to oxidative stress.
Several, but not all, studies have shown that cells
from individuals varying in age do have a different replicative
lifespan in vitro. Ksiazek et al(6) reported that human peritoneal
mesothelial cells (HPMCs) from aged donors (>75 years) have
lower proliferative capacity and increased 8-OxoG content compared
to cells from younger individuals (<25 years). However, even in
early passage HPMCs, repair of 8-OxoG was delayed in cells from
aged compared to young donors; a positive correlation was noted
between donor age and 8-OxoG level, and an inverse relationship was
noted between those 8-OxoG levels and subsequent cell lifespan.
Yuan et al(7)
demonstrated that primary human skin fibroblasts were more
susceptible to oxidative stress from hydrogen peroxide, the
superoxide radical, or linoleic acid hydroperoxide at late passage
compared to early passage, presumably due to decreases in cellular
reduced glutathione concentration and catalase activity.
Surprisingly, a more recent study by Matsuo et al(8) reported that primary human skin
fibroblasts derived from aged donors were more resistant to
oxidative stress from hydrogen peroxide, linoleic acid
hydroperoxide or UVB compared to fibroblasts from young donors. The
activity of glutathione peroxidase was higher in fibroblasts from
aged compared to young donors, suggesting that the fibroblasts from
aged donors are more resistant to oxidative stress due to an
increase in cellular glutathione peroxidase. Kaneko et
al(9) showed that skin
fibroblasts established from fetal tissue showed an increase in
8-oxo-2′-deoxyguanosine content and a similar decreased activity of
8-oxo-2′-deoxyguanosine endonucclease and DNA polymerases with
increasing cell passage during in vitro cellular ageing.
Increases in superoxide dismutase and glutathione peroxidase were
observed prior to increased 8-OxoG, while catalase activity
decreased with in vitro cellular ageing at late passage.
These results suggest that defense mechanisms against oxidative
stress remain sufficiently active in late passage fetal skin
fibroblasts, but that repair systems against oxidative damage
decrease at late passage. The 8-OxoG content in fibroblasts from
fetal skin changed little up to 60 population doubling levels (PDL)
and then increased significantly after 63 PDL (9). In contrast, the 8-OxoG content in
fibroblasts from a 36-year-old adult donor changed little up to 38
PDL but increased beyond 44 PDL and in fibroblasts from a
72-year-old donor the 8-OxoG content changed little up to 32 PDL,
but then increased after 36 PDL. These results suggest that repair
systems against oxidative damage may start to decrease at an
earlier cell passage number in fibroblasts from aged compared to
young donors.
There are numerous factors that could contribute to
ageing and lifespan, such as telomere shortening, hormone levels
and multiple targets of reactive oxygen species, making it
difficult to establish a direct role of BER in counteracting
ageing. It was, therefore, considered important to examine the
repair of oxidative DNA damage in skin fibroblasts from young and
elder donors.
We previously reported a host cell reactivation
(HCR) assay for examining BER of methylene blue plus visible light
(MB+VL)-induced 8-OxoG lesions in a number of different cell
strains including xeroderma pigmentosum fibroblasts from
complementation group C and CS fibroblasts (10–13).
The HCR assay utilizes a recombinant non-replicating adenovirus
(Ad5CMVlacZ) expressing the β-galactosidase (β-gal) reporter
gene under control of the cytomegalovirus immediate early (IE)
promoter (14) to examine the
ability of different cell types to remove damage and reactivate
reporter gene expression. Methylene blue (MB), a type II
photosensitizer, produces singlet oxygen
(1O2) upon exposure to visible light (VL) in
the presence of oxygen (15) which
through interaction with DNA predominantly leads to the formation
of 8-OxoG lesions with a small number of other single base
oxidative lesions occurring (16,17).
In the present study, we examined HCR of a
MB+VL-treated reporter gene in human skin fibroblasts from 10
donors of different ages and from fibroblasts from 2 donors as a
function of increasing passage number of cells grown in tissue
culture.
Materials and methods
Cell lines and virus
Skin fibroblasts from normal donors, GM9503, GM37G,
GM38A, GM969, GM8399, GM8400, GM01706A, AG02261B, GM1863 and
GM00288B were obtained from the National Institute of General
Medical Sciences, Human Genetic Cell Repository, Coriell Institute
for Medical Research (Camden, NJ, USA). Cell cultures were grown at
37°C in a humidified incubator in 5% CO2 and cultured in
Eagle’s α-minimal essential media (α-MEM) supplemented with 10%
fetal bovine serum and antimycotic-antibiotic 100 μg/ml penicillin,
100 μg/ml streptomycin and 250 ng/ml amphotericin B (Gibco-BRL,
Grand Island, NY, USA).
The recombinant adenovirus Ad5MCMVlacZ
(AdCA35) (14) was obtained from Dr
F.L. Graham, McMaster University. The virus was propagated,
collected and titered as previously described (18).
Treatment of the virus with methylene
blue plus visible light (MB+VL)
Preparation of MB was as previously described
(13). Treatment of the virus was
performed as previously described (10). Briefly, 80 μl of AdCMVLacZ
virus was added to 3.6 ml of phosphate-buffered saline (PBS)
containing 20 μg/ml MB in 35-mm Petri dishes on ice. With
continuous stirring, the virus suspensions were irradiated (or mock
irradiated) with VL. VL irradiation of virus employed a General
Electric 1000-watt halogen lamp (GE R1000) at a distance of 70 cm
from the bulb. After each time point, 400 μl of irradiated virus
was removed, diluted appropriately in unsupplemented α-MEM and used
to infect the cell monolayers.
HCR of MB+VL-treated Ad5CMVLacZ
Primary human fibroblasts were seeded at a density
of 1.5×104 to 1.8×104 cells/well in 96-well
plates (Falcon, Franklin Lakes, NJ, USA). After seeding, cells were
incubated for 18–24 h and subsequently infected with 40 μl of
untreated or MB+VL-treated virus for 90 min at a multiplicity of
infection (MOI) of 20–40 plaque forming units (pfu)/cell. Following
infection, cells were overlaid with 160 μl of complete α-MEM and
incubated for a further 12–40 h before harvesting. A single HCR
experiment consisted of triplicate wells for each treatment of the
virus, and triplicate wells of non-infected cells were used to
obtain background levels of β-gal activity. β-gal activity was
scored by measuring absorbance at 570 nm as previously described
(19).
Results
HCR of the MB+VL-treated reporter gene in
normal human skin fibroblasts from different passage numbers in
tissue culture
Normal skin fibroblasts from 2 donors of ages 10 and
19 years were examined for HCR of the MB+VL-treated reporter gene
in fibroblasts from different passage numbers in tissue culture.
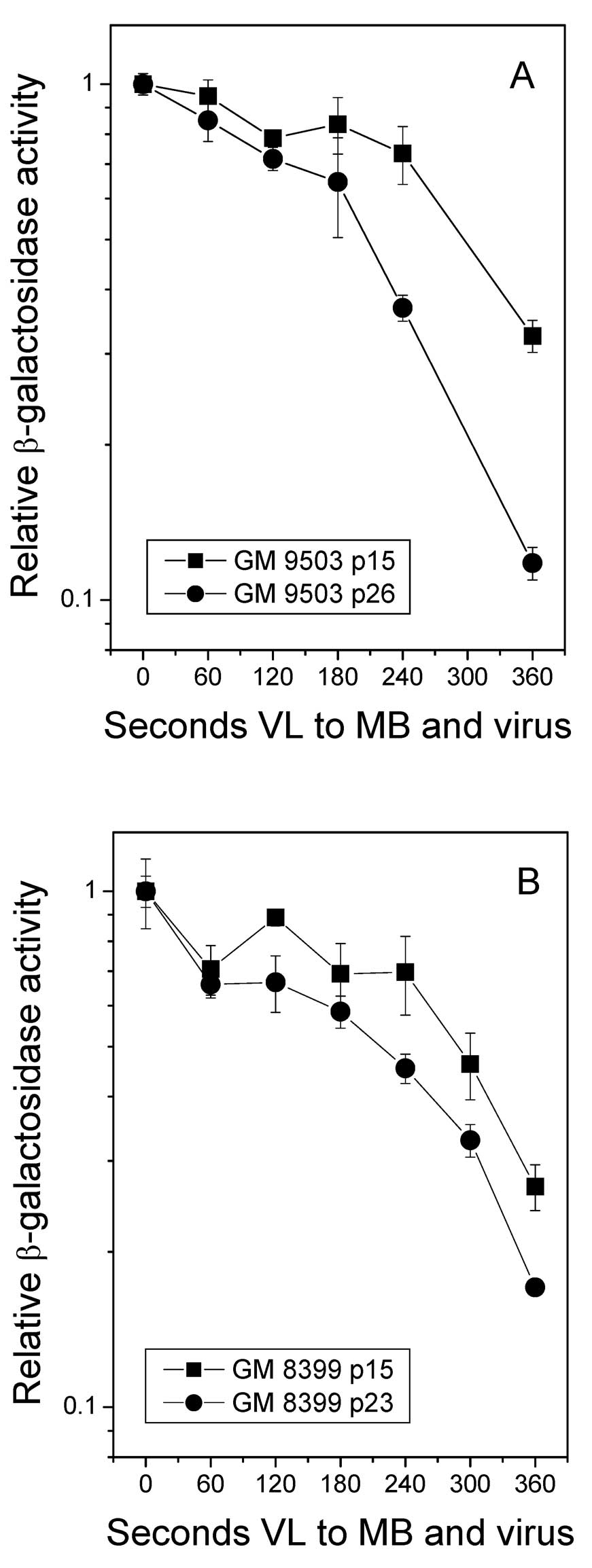
Representative survival curves for β-gal activity for MB+VL-treated
virus for the different fibroblast strains are shown in Fig. 1. From the β-gal survival curves, the
VL exposure required to reduce β-gal activity to 50% of that for
non-VL-exposed virus (D50) was used as a measure of HCR.
It was noted that for both GM9503 and GM8399 cells, HCR of the
MB+VL-treated reporter gene was greater in low passage compared to
high passage number cells. Similar results were obtained in
additional individual experiments where low passage number cells
ranged from 13 to 21 for GM9503 and 10 to 19 for GM8399 and high
passage numbers ranged from 22 to 31 for GM9503 and 19 to 29 for
GM8399. Differences between low and high passage number for
individual experiments ranged from 6 to 12 for GM9503 and 6 to 14
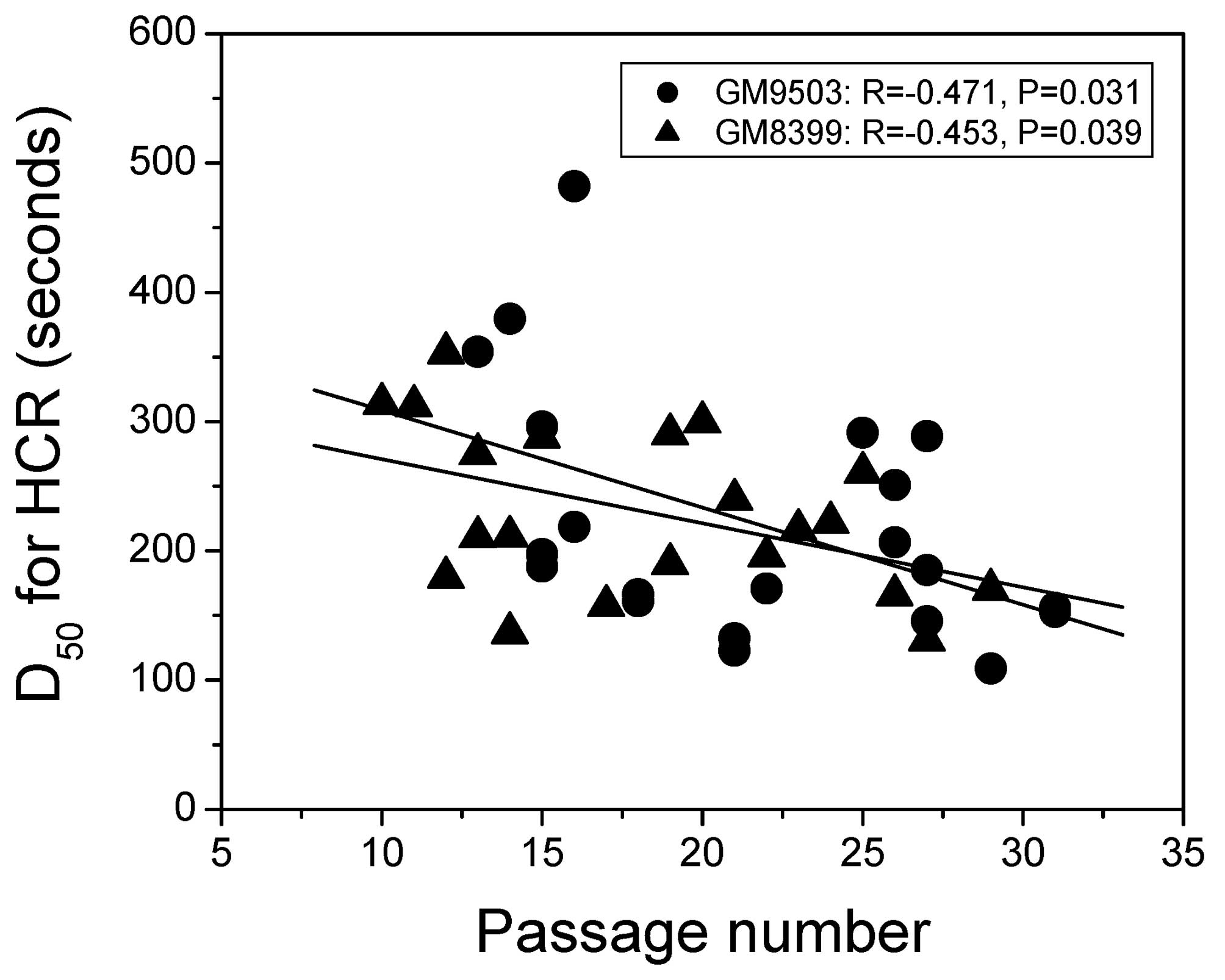
for GM8399. The D50 value for HCR of the MB+VL-treated
reporter gene is plotted against passage number for GM9503 and
GM8399 over a range of 10 to 32 cell passages in Fig. 2. A significant decrease in
D50 value with increasing passage number was found for
GM9503 (R=−0.471, P=0.031) and GM8399 (R=−0.453, P=0.039).
HCR of the MB+VL-treated reporter gene in
normal human skin fibroblasts of different ages
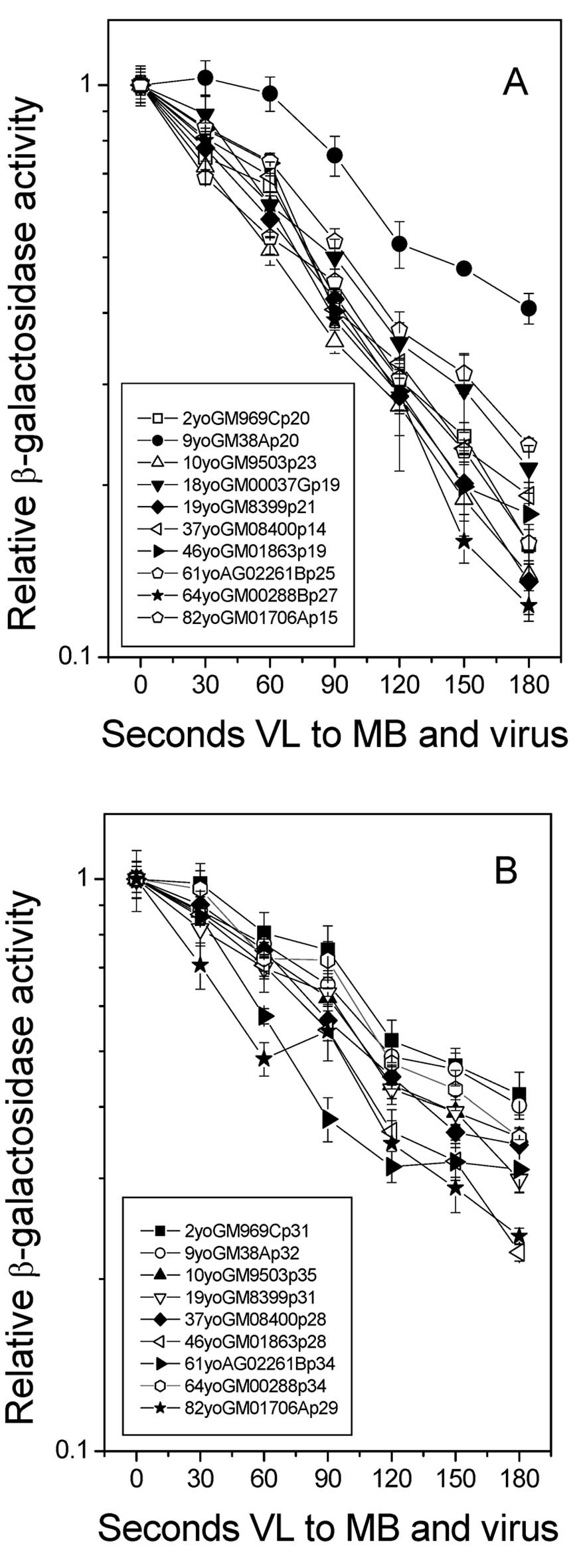
In a second series of experiments, normal human
fibroblasts from 10 donors of different ages ranging from 2 to 82
years at passage numbers 12 to 35 were examined for HCR of
MB+VL-treated Ad5CMVlacZ. Each experiment was performed
under a different condition of light output from the 1000-watt
halogen lamp. Survival curves of β-gal activity for the
MB+VL-treated virus in the various fibroblast strains are shown in
Fig. 3 for representative
experiments using low (A) and high (B) passage number. For each
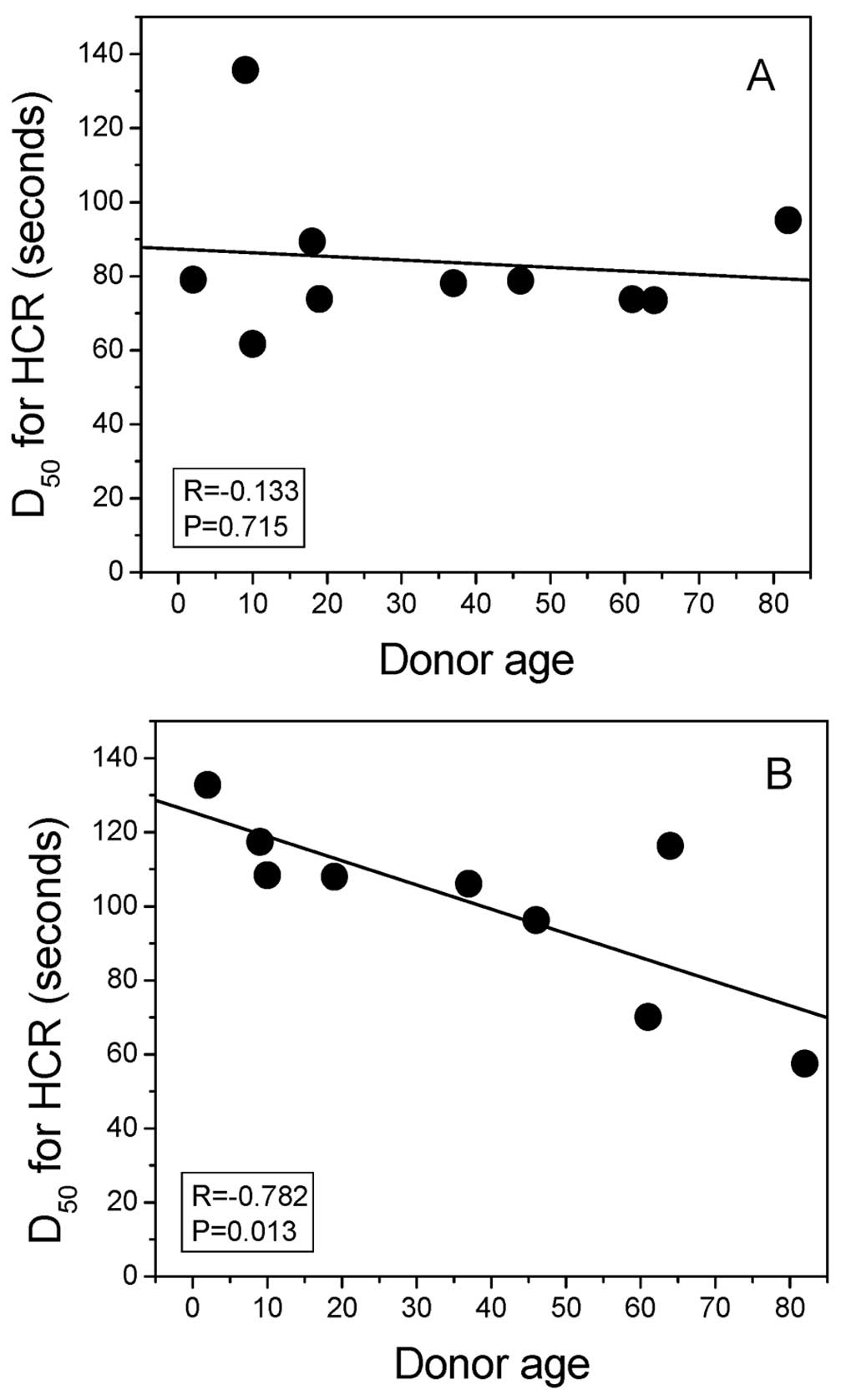
experiment, the D50 value for HCR of the MB+VL-treated
reporter gene was plotted against donor age in Fig. 4. For this sampling of normal skin
fibroblasts from donors of different ages, a significant
correlation of HCR was noted with donor age at high passage number
(28–35) (R=−0.782, P=0.013) (Fig.
4B) but not at low passage number (14–27) (R=−0.133, P=0.715)
(Fig. 4A).
Discussion
Reports on the repair of UV-induced DNA damage in
skin fibroblasts from young compared to elder donors are
conflicting. Using cells of a similar passage number of less than
13 PDL, Takahashi et al(20)
reported a significant decline in HCR of a transfected
UV-irradiated chloramphenicol acetyltransferase gene, but not in
the actual removal of cyclobutane pyrimidine dimers (CPDs) and 6–4
photoproducts as determined using an ELISA assay, in skin
fibroblasts from aged donors compared to fibroblasts from young
donors. These authors also found reduced expression of DNA repair
synthesis-related genes in skin fibroblasts from aged donors
compared to young donors and suggest that the reduced HCR reflects
a reduced post-UV repair capacity in cells from ageing donors that
results from an impairment in a late step of NER due to the
decrease in factors required for repair synthesis. In contrast,
Merkle et al(21) reported
no significant difference between normal skin from young and aged
donors for host cell reactivation of a transfected UV-irradiated
luciferase gene providing no indication that the higher incidence
of skin cancer observed with increasing age is due to an
age-related decrease in the ability to repair UV-induced DNA
damage. In their experiments they compared fibroblasts from young
and aged donors at cumulated PDL ranging from 7 to 15 in the
various fibroblast strains. In addition, they found that
fibroblasts from both young and aged donors showed a similar and
significant decrease in HCR with increasing PDL over the range of
10 to 40 PDL.
In the present study, we found a significant
reduction in HCR of the MB+VL-treated reporter gene with increasing
cell passage number, indicating that BER decreases with increasing
time of cells grown in tissue culture. We found also a significant
reduction in HCR of the MB+VL-treated reporter gene with increasing
donor age for high passage (passage 28–35), but not for low passage
primary human fibroblasts (passage 14–27). Kaneko et
al(9) reported that the 8-OxoG
content of fibroblasts from a 72-year-old donor changed little up
to 32 PDL and increased after 36 PDL, whereas the 8-OxoG content of
fibroblasts from fetal skin changed little up to 60 PDL and then
increased after 63 PDL. Although the study by Kaneko et al
did not directly measure DNA repair, their report is consistent
with a decrease in the repair of 8-OxoG at an earlier cell passage
number in fibroblasts from aged compared to young donors as
reported here.
Acknowledgements
The present study was supported by an operating
grant (no. 16066) to A.J.R. from the National Cancer Institute of
Canada with funds from the Canadian Cancer Society.
References
|
1
|
Maynard S, Schurman SH, Harboe C, de
Souza-Pinto NC and Bohr VA: Base excision repair of oxidative DNA
damage and association with cancer and aging. Carcinogenesis.
30:2–10. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tuo J, Jaruga P, Rodriguez H, Bohr VA and
Dizdaroglu M: Primary fibroblasts of Cockayne syndrome patients are
defective in cellular repair of 8-hydroxyguanine and
8-hydroxyadenine resulting from oxidative stress. FASEB J.
17:668–674. 2003. View Article : Google Scholar
|
|
3
|
Chen SK, Hsieh WA, Tsai MH, Chen CC, Hong
AI, Wei YH and Chang WP: Age-associated decrease of oxidative
repair enzymes, human 8-oxoguanine DNA glycosylases (hOgg1), in
human aging. J Radiat Res. 44:31–35. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Szczesny B, Hazra TK, Papaconstantinou J,
Mitra S and Boldogh I: Age-dependent deficiency in import of
mitochondrial DNA glycosylases required for repair of oxidatively
damaged bases. Proc Natl Acad Sci USA. 100:10670–10675. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cabelof DC, Raffoul JJ, Ge Y, Van Remmen
H, Matherly LH and Heydari AR: Age-related loss of the DNA repair
response following exposure to oxidative stress. J Gerontol A Biol
Sci Med Sci. 61:427–434. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ksiazek K, Piatek K and Witowski J:
Impaired response to oxidative stress in senescent cells may lead
to accumulation of DNA damage in mesothelial cells from aged
donors. Biochem Biophys Res Commun. 373:335–339. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan H, Kaneko T and Matsuo M: Increased
susceptibility of late passage human diploid fibroblasts to
oxidative stress. Exp Gerontol. 31:465–474. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matsuo M, Ikeda H, Sugihara T, Horiike S,
Okano Y and Masaki H: Resistance of cultured human skin fibroblasts
from old and young donors to oxidative stress and their glutathione
peroxidase activity. Gerontology. 50:193–199. 2004. View Article : Google Scholar
|
|
9
|
Kaneko T, Tahara S, Taguchi T and Kondo H:
Accumulation of oxidative DNA damage, 8-oxo-2′-deoxyguanosine, and
change of repair systems during in vitro cellular aging of cultured
human skin fibroblasts. Mutat Res. 487:19–30. 2001.
|
|
10
|
Kassam SN and Rainbow AJ: Deficient base
excision repair of oxidative DNA damage induced by methylene blue
plus visible light in xeroderma pigmentosum group C fibroblasts.
Biochem Biophys Res Commun. 359:1004–1009. 2007. View Article : Google Scholar
|
|
11
|
Kassam SN and Rainbow AJ: UV-inducible
base excision repair of oxidative damaged DNA in human cells.
Mutagenesis. 24:75–83. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leach DM and Rainbow AJ: Early host cell
reactivation of an oxidatively damaged adenovirus-encoded reporter
gene requires the Cockayne syndrome proteins CSA and CSB.
Mutagenesis. 26:315–321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pitsikas P, Lee D and Rainbow AJ: Reduced
host cell reactivation of oxidative DNA damage in human cells
deficient in the mismatch repair gene hMSH2. Mutagenesis.
22:235–243. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Addison CL, Hitt M, Kunsken D and Graham
FL: Comparison of the human versus murine cytomegalovirus immediate
early gene promoters for transgene expression by adenoviral
vectors. J Gen Virol. 78:1653–1661. 1997.PubMed/NCBI
|
|
15
|
Slamenova D, Kuboskova K, Horvathova and
Robichova S: Rosemary-stimulated reduction of DNA strand breaks and
FPG-sensitive sites in mammalian cells treated with
H2O2 or visible light-excited methylene blue.
Cancer Lett. 177:145–153. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Floyd RA, West MS, Eneff KL and Schneider
JE: Methylene blue plus light mediates 8-hydroxyguanine formation
in DNA. Arch Biochem Biophys. 273:106–111. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tuite EM and Kelly JM: Photochemical
interactions of methylene blue and analogues with DNA and other
biological substrates. J Photochem Photobiol B. 21:103–124.
1993.
|
|
18
|
Graham FL and Prevec L: Manipulation of
adenovirus vectors. Methods Mol Biol. 7:109–128. 1991.PubMed/NCBI
|
|
19
|
Pitsikas P, Francis MA and Rainbow AJ:
Enhanced host cell reactivation of a UV-damaged reporter gene in
pre-UV-treated cells is delayed in Cockayne syndrome cells. J
Photochem Photobiol B. 81:89–97. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takahashi Y, Moriwaki S, Sugiyama Y, Endo
Y, Yamazaki K, Mori T, Takigawa M and Inoue S: Decreased gene
expression responsible for post-ultraviolet DNA repair synthesis in
aging: a possible mechanism of age-related reduction in DNA repair
capacity. J Invest Dermatol. 124:435–442. 2005. View Article : Google Scholar
|
|
21
|
Merkle TJ, O’Brien K, Brooks PJ, Tarone RE
and Robbin JH: DNA repair in human fibroblasts, as reflected by
host-cell reactivation of a transfected UV-irradiated luciferase
gene, is not related to donor age. Mutat Res. 554:9–17. 2004.
View Article : Google Scholar : PubMed/NCBI
|