Introduction
Liver cancer is one of the most malignant diseases.
The early diagnosis and treatment will greatly improve patient
prognosis. In recent years, there has been a growing awareness that
the cellular microenvironment during tumorigenesis plays a critical
role in cancer cell proliferation, survival, migration and invasion
(1–3). Substantial evidence shows that cancer
is the result of imbalance of endogenous angiogenesis inhibitor and
accelerator local concentrations in the tumor tissue. The most
important angiogenesis promoting agent is basic fibroblast growth
factor (bFGF) and vascular growth factor (VEGF) (4,5).
Several recent reports have also implicated bFGF as a major
survival factor, which has been shown to mediate this additional
function through the induction of Bcl-2 and the activation of the
PI3 kinase-Akt/protein kinase B (PKB) signaling pathway. bFGF can
also mediate the induction/upregulation of members of a newly
discovered family of antiapoptotic proteins, i.e., the inhibitors
of apoptosis (IAP) in vascular endothelial cells (6,7). PKB
is an enzyme that covalently attaches ATP-phosphate groups to the
serine/threonine on protein substrates to alter the activity of the
targeted protein (8,9). The activation of Akt is initiated with
the plasma membrane recruitment of phosphatidylinositol 3-kinase
(PI3K), an upstream enzyme stimulated by a variety of activated
growth factor receptors such as epidermal growth factor receptor
(EGFR), vascular growth factor receptor (VEGFR), bFGF, and
insulin-like growth factor receptor (IGFR) (10). The activation of Akt is completed
via the phosphorylation of Thr-308 and Ser-473, leading to
increased Akt activity toward a variety of downstream substrates,
such as the family of forkhead transcription factors for inhibiting
tumor proliferation, the mammalian target of rapamycin (mTOR) for
modulating protein synthesis, and the Bcl-xL/Bcl-2-associated death
promoter (BAD) for uncontrolled proliferation (9,11).
Since Akt acts as a signal hub in the regulation of cell survival,
proliferation and growth, the elevation in Akt activity is found to
be correlated with increased tumorigenicity (8). Thus, a variety of inhibitors targeting
Akt or its up- or downstream events are currently under clinical
trials (12). Wortmannin is a
potent and selective inhibitor of Akt, and molecular imaging of Akt
activity has become an important approach in monitoring Akt
activity in vivo. Numerous studies have confirmed that
PI3K/PKB plays a crucial role in promoting proliferation and
inhibiting apoptosis (13–16).
Survivin is the smallest protein of the known IAP
family (17). It is a unique
bifunctional protein that inhibits apoptosis by suppressing
caspase-3 and caspase-7 and modulates the G2/M phase of the cell
cycle through association with mitotic spindle microtubules
(18). Survivin contains a single
BIR domain and can bind caspases, preventing caspase-induced
apoptosis (19); it is the
least-expressed IAP family candidate in adult tissues (almost
absent in normal tissue), but can be recognized in developing fetal
tissues, with the exception of placenta and thymus (20–23).
Survivin is able to inhibit apoptosis and promote proliferation of
tumor cells. Recently, it was found that survivin protein
degradation is associated with the disorder of pantothenic acid -
proteasome pathway in a cell cycle-dependent manner (24). The potential value of survivin in
tumor diagnosis and treatment has gained considerable attention. It
is possible to inhibit growth and recurrence of liver cancer
through inhibiting the expression of survivin. Survivin is the
strongest inhibitor of apoptosis and its regulation and signaling
pathway has yet to be fully clarified. It was reported that bFGF
and its receptor FGFR1 highly expressed in hepatocarcinoma cells,
play an important role in the occurrence and development of liver
cancer and promote the growth of liver cancer cells by autocrine or
paracrine mechanisms (25).
Investigations into the FGF/FGFR signal pathway may reveal the
pathogenesis of numerous diseases, and it may become a new target
for the treatment of liver cancer and other diseases. In the
present study, we examined whether bFGF could regulate the
expression of survivin and affect the apoptosis of liver cancer
cells, and we elucidated its signal pathway so as to provide a
basis for bFGF in regulating apoptosis via the PI3K/PKB pathway in
the treatment of liver cancer.
Materials and methods
Cells lines and culture
Human hepatocarcinoma cell line Bel-7402 (Shanghai
Institute of Cell Biology, Chinese Academy of Sciences) was
maintained in DMEM medium supplemented with 10% fetal calf serum
(Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.,
China), 100 U/ml penicillin and 100 μg/ml streptomycin (Gibco) in
5% CO2 at 37°C. Bel-7402 cells were exposed to bFGF
(Beijing Shuanglu Pharmaceutical Co., Ltd., China) at different
concentrations (0, 12.5, 25, 50 and 75 ng/ml) for 10 min, or to 25
ng/ml bFGF at different times (0, 5, 10, 30 and 60 min)
respectively. Each group had 3 parallel wells. Wortmannin (Sigma)
was dissolved in dimethyl sulfoxide (DMSO) in serum-free medium
with a final concentration of 100, 200 and 400 nM.
Western blot analysis
Cultured cells were harvested, washed with
phosphate-buffered saline (PBS), and resuspended in lysis buffer
(20 mM Tris-HCl, pH 7.5, containing 1% SDS (sodium dodecyl
sulfate), 50 mM NaCl, 1 mM EDTA (ethylenediaminetetraacetic acid),
1 mM PMSF (phenylmethylsulfonyl fluoride), 10 mM sodium fluoride,
and 1 mM sodium orthovanadate. Cell samples were then agitated at
4°C for 1 h followed by centrifugation for 15 min. Equivalent
amounts of protein (40 μg) from each sample were separated on 10%
SDS-PAGE (polyacrylamide gel electrophoresis) and the separated
proteins were transferred to nitrocellulose membranes. Membranes
were routinely blocked in 5% nonfat milk in PBS with 0.1% Tween-20
for an hour with agitation and washed, and primary antibodies PKB
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA) or survivin
were added (1:500–1,000 dilution in 5% bovine serum albumin in PBS
buffer). Membranes were incubated overnight at 4°C with agitation,
washed, and then incubated with horseradish peroxidase-conjugated
secondary antibodies (1:5,000 dilutions in 5% bovine serum albumin
in PBS with 0.1% Tween-20) for 3 h at room temperature. Proteins
were detected using the Enhanced Chemiluminescence Western Blotting
Detection Reagent (Pierce, Rockford, IL, USA). The results were
described as a ratio of relative absorbance value of the protein
band of interest to β-actin.
RNA extraction and RT-PCR
Total RNA was extracted, reverse transcribed,
amplified and analyzed as previously described (26). Briefly, total RNA was extracted from
Bel-7402 cells using a total RNA isolation kit (TransGen) according
to the manufacturer’s protocol. An ultraviolet spectrophotometer
was used to determine the quantity and quality of total RNA.
β-actin was used to normalize cDNA quantities and was amplified
with the following primers: forward, 5′-ATCATGTTT GAGACCTTCAACA-3′
and reverse, 5′-CATCTCTTGCTC GAAGTCCA-3′. For survivin, the
following primers were used: forward, 5′-GCACTTTCTTCGCAGTTTCC-3′
and reverse, 5′-GGACCACCGCATCTCTACAT-3′ (Biosune). The PCR reaction
was carried out under the following conditions: 35 cycles of
denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and an
extension at 72°C for 2 min. A final extension was performed at
72°C for 1 min. PCR products were separated by 1% agarose gel
electrophoresis.
Cell-cycle and apoptosis assay
Bel-7402 cells were treated with different
concentrations of bFGF for different times in serum-free medium.
Following incubation, cells were harvested, washed in PBS, fixed in
cold 70% ethanol for 45 min, stained with propidium iodide solution
that contains RNase A for 30 min, and analyzed by flow cytometry
(BD Biosciences, San Jose, CA, USA). CellQuest analysis software
was used for DNA content analysis of the cell cycle.
Statistical analysis
All values were expressed as means ± SD. Statistical
analysis was processed using one-way ANOVA. P<0.05 was
considered to indicate a statistically significant difference.
Results
PKB activity increases in hepatocarcinoma
cells following bFGF treatment
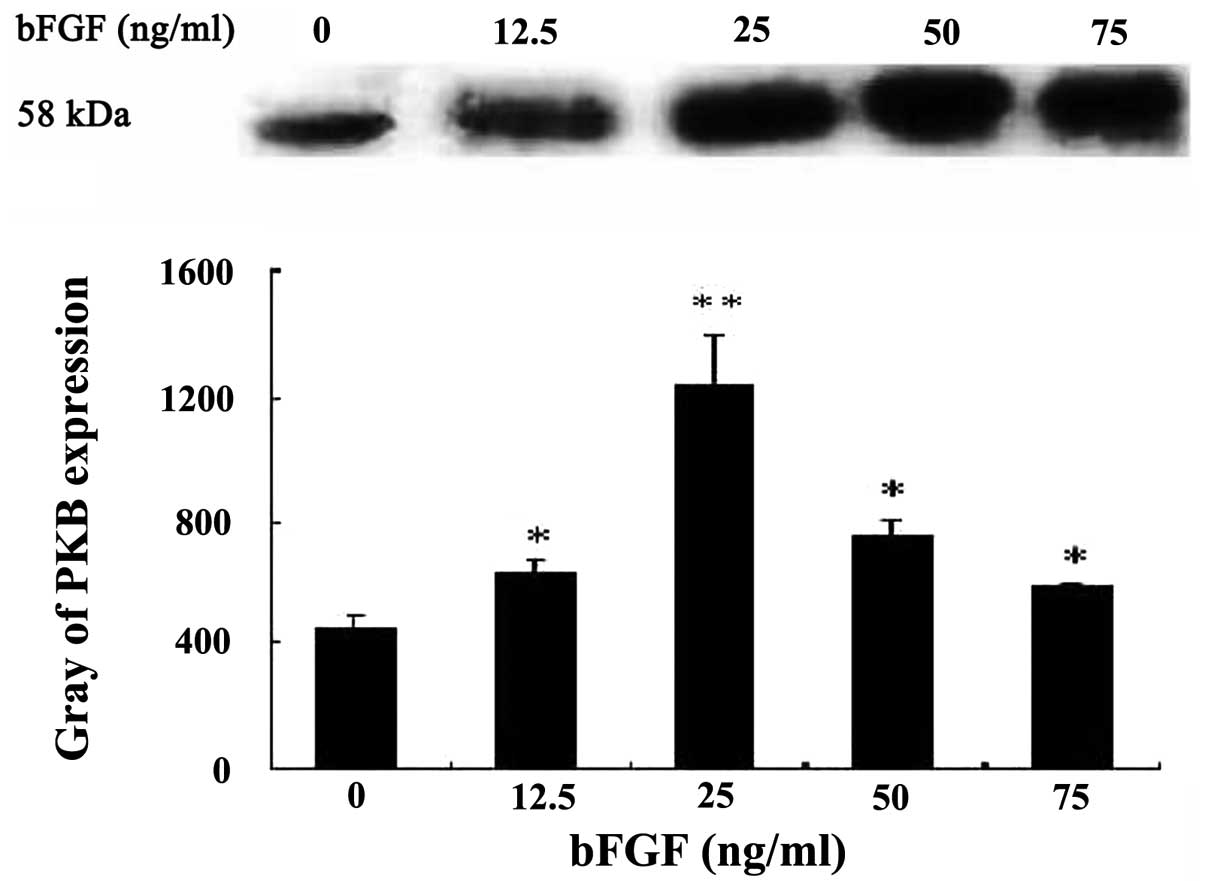
The cytosolic PKB activity gradually increased with
bFGF 0–25 ng/ml following different concentrations of bFGF
co-incubated with Bel-7402 cells for 10 min, and reached the
highest value at 25 ng/ml, 2.81-fold of control (P<0.01); then,
cytosol PKB activity declined. There was a significant difference
of PKB expression between the bFGF-treated and -untreated groups
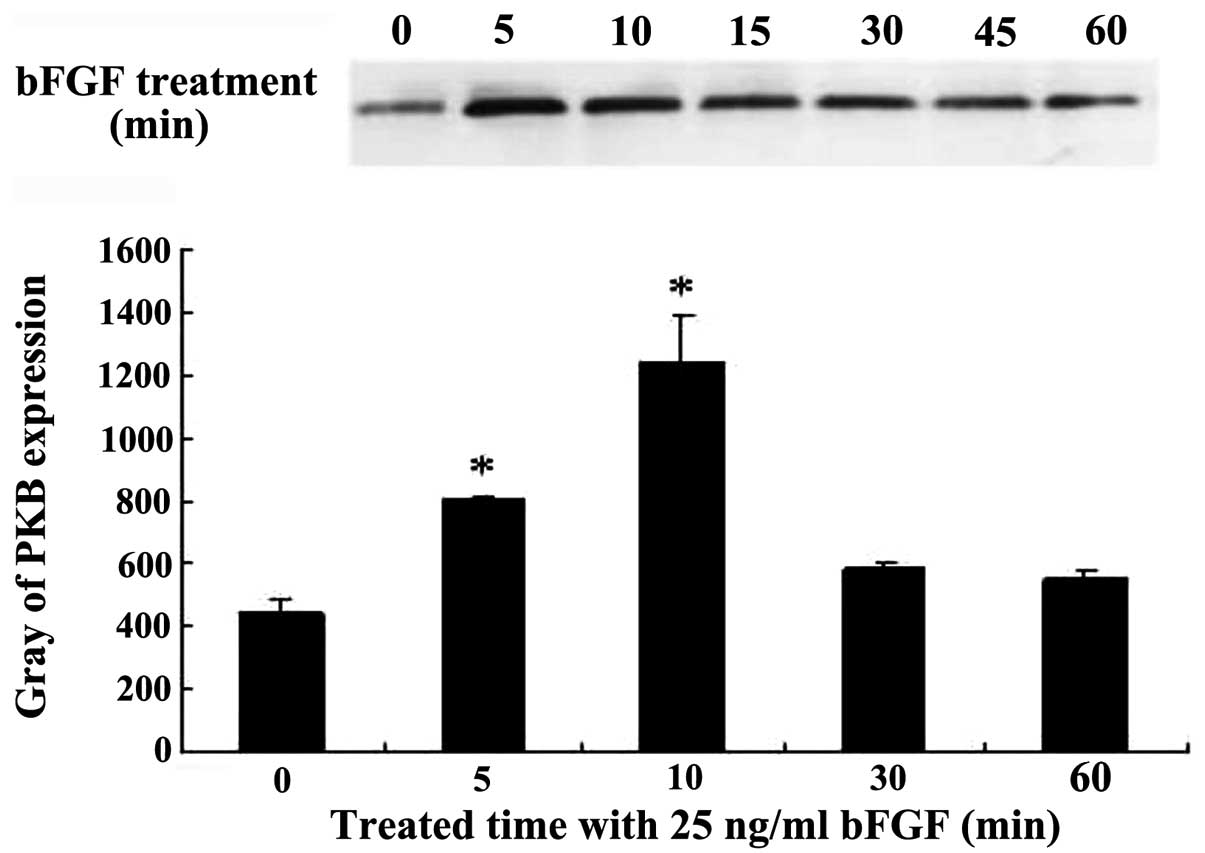
(P<0.05) (Fig. 1). Bel-7402
cells were co-incubated with 25 ng/ml bFGF for 5, 10, 30 and 60
min. The PKB cytosol activity began to increase at 5 min, 1.82-fold
for the control group, and it increased the most at 10 min, which
was 2.81-fold that of the control group, and subsequently PKB
activity began to decline at 60 min. There was a statistically
significant difference (P<0.05) (Fig. 2). Western blot analysis showed that
PKB expression was increased at 15, 30, 45 and 60 min with the
stimulation of 25 ng/ml bFGF, but PKB expression was clearly
inhibited at each time-point when wortmannin (200 nM) was added.
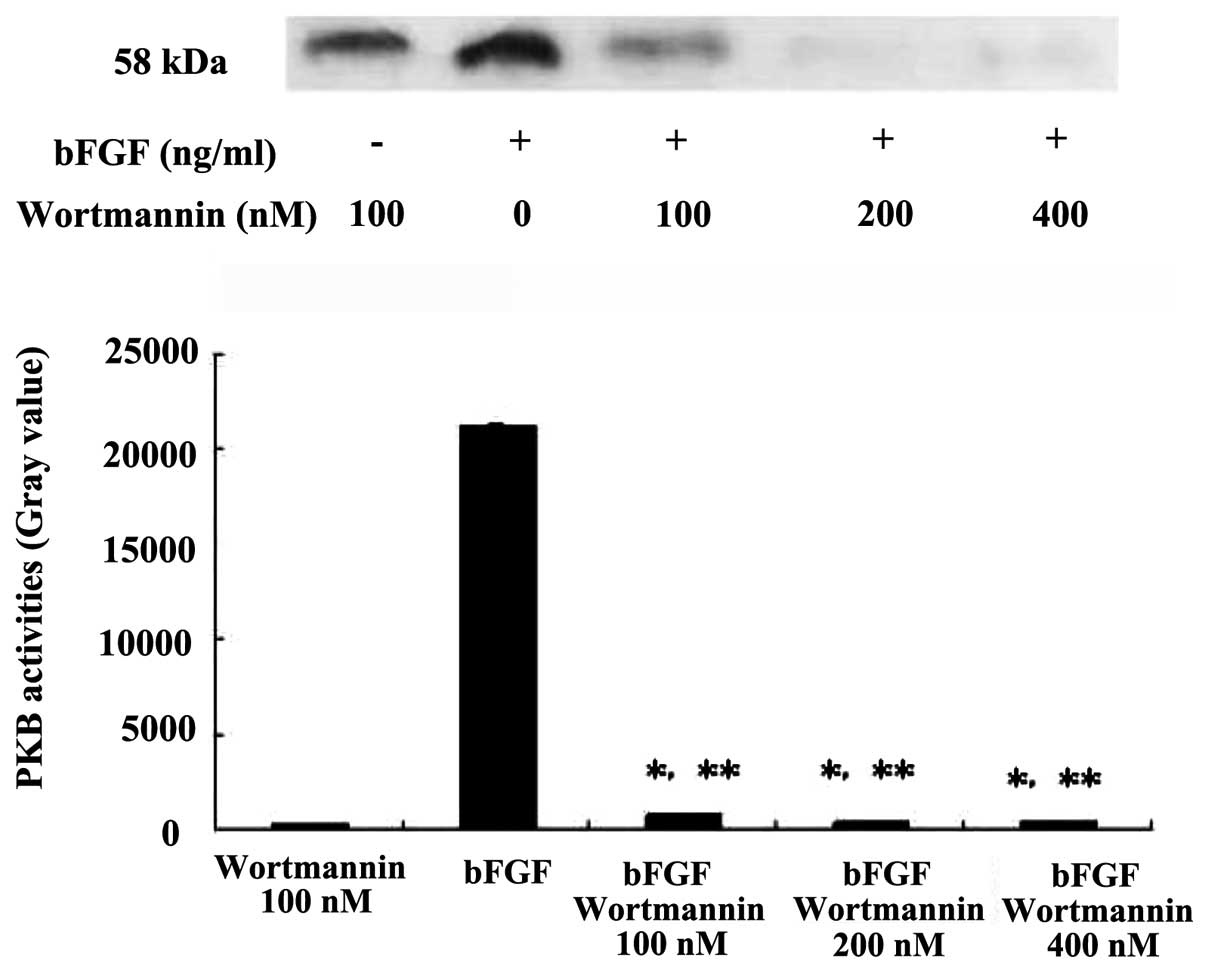
The inhibition occurred in a time-dependent manner (Fig. 3). When pretreated with different
concentrations of wortmannin under 25 ng/ml bFGF, the expression of
PKB was inhibited significantly, in a dose-dependent manner,
compared with the wortmannin control group P<0.05, compared with
the bFGF control group P<0.01 (Fig.
4).
Survivin mRNA is upregulated with bFGF
and downregulated with wortmannin
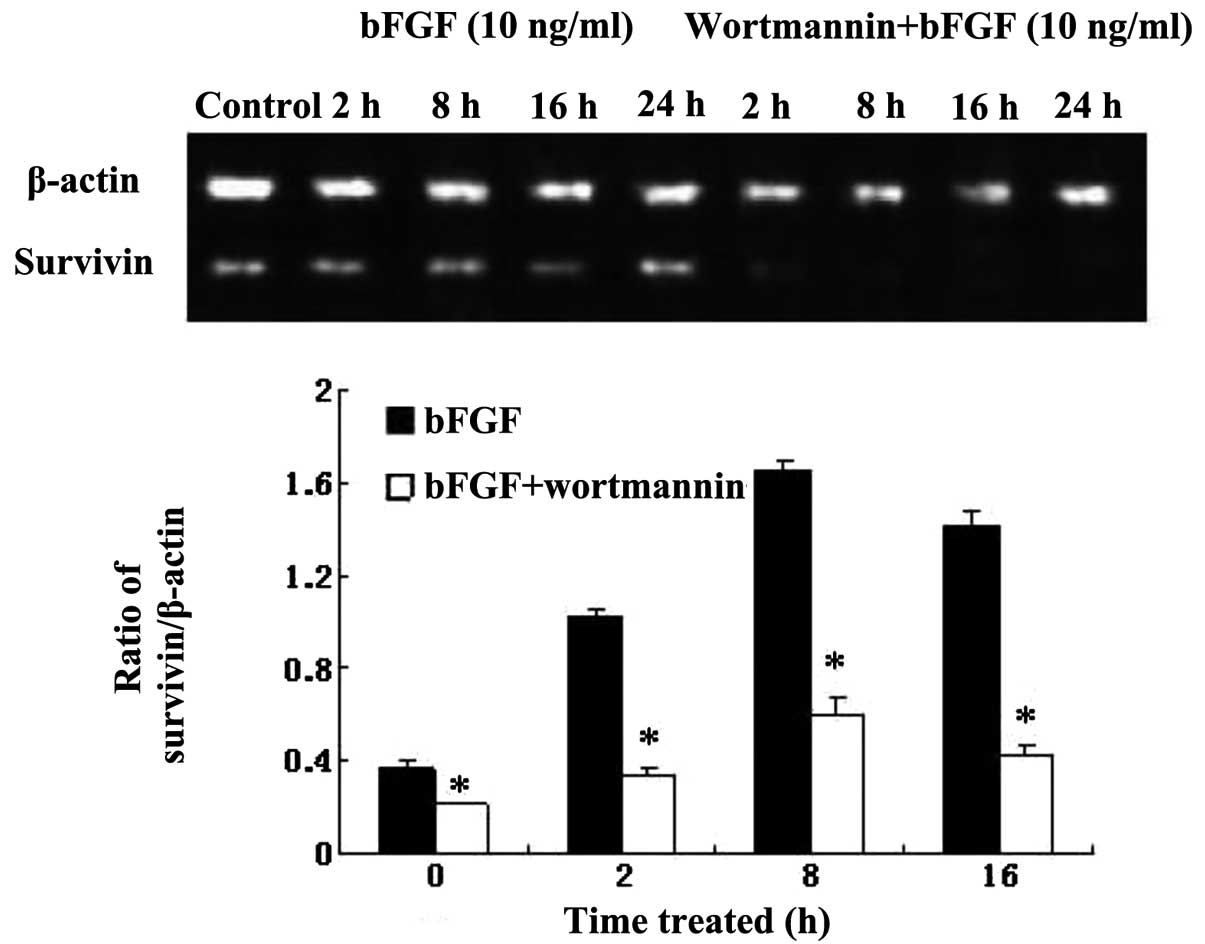
RT-PCR analysis showed that survivin mRNA expression
of Bel-7402 cells was increased, and reached a peak at 16 h,
7.86-fold upregulated compared with the control group after
treatment with 10 ng/ml bFGF (Fig.
5) (P<0.05), which suggested that bFGF could induce the
expression of survivin mRNA in liver cancer cells. While survivin
mRNA expression was significantly suppressed by pretreatment with
wortmannin (200 nM) for 1 h, P<0.05, the highest inhibition
occurred at 8 h (Fig. 5). The
results suggested that increasing survivin mRNA expression induced
by bFGF can be blocked by wortmannin in Bel-7402 cells.
bFGF and wortmannin are closely related
to cell proliferation and apoptosis
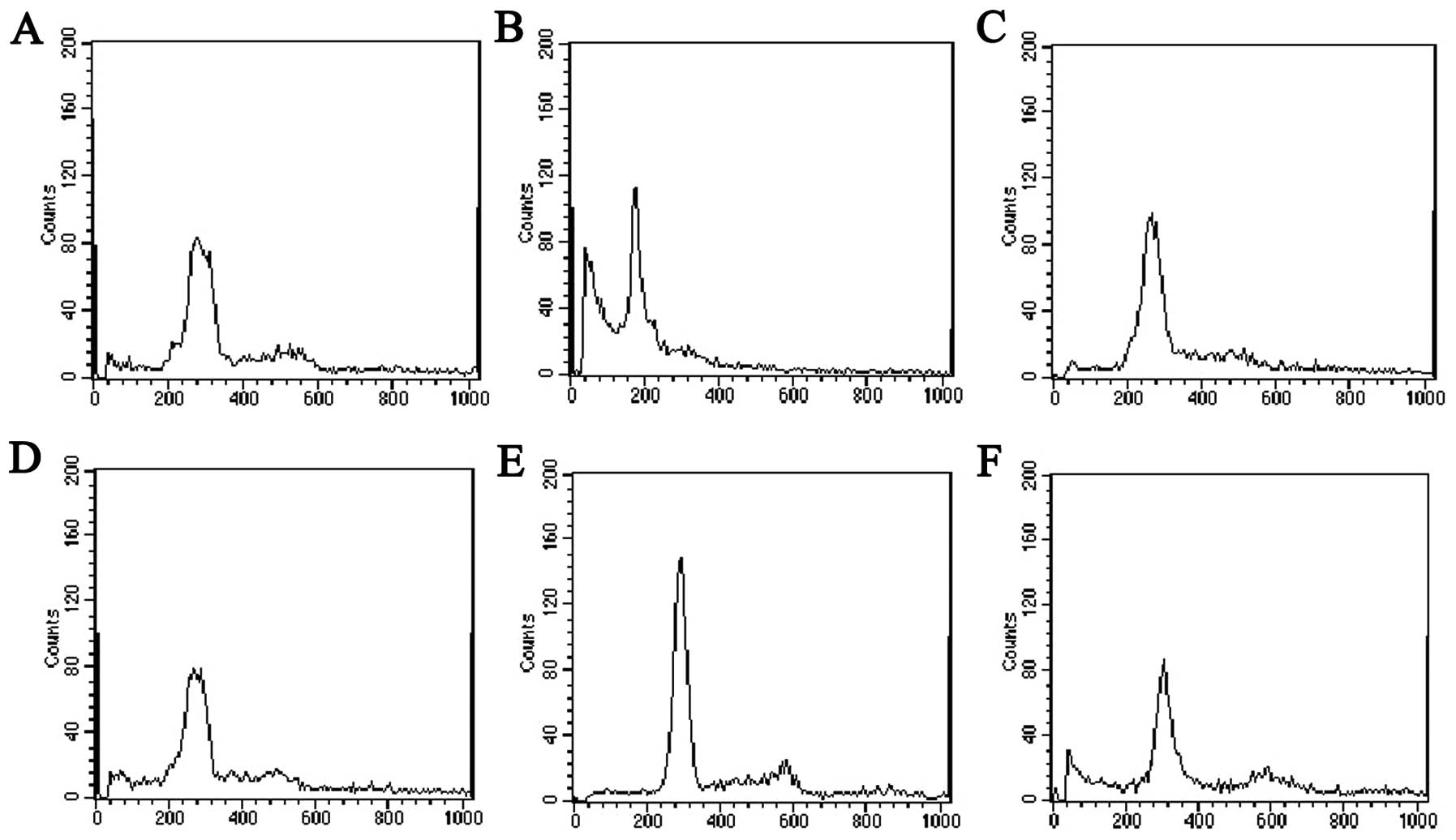
The results of flow cytometry showed lower diploid
peak and a small sub-G1 peak (apoptosis cells) in the Bel-7402 cell
control group (Fig. 6A). Bel-7402
cells pretreated with wortmannin (200 nM, 1 h) only showed
significant hypodiploid peak, accompanied by a significant decline
of S phase peak and apoptosis rate up to 50% compared with the cell
control group, P<0.05 (Fig. 6B).
Following incubation with 25 ng/ml bFGF for 10 min, the apoptosis
rate and M phase cells were apparently decreased, and S phase cells
increased compared with the wortmannin-treated group (Fig. 6C); however, no marked difference was
detected compared with control cells. When this group was
pretreated with wortmannin (200 nM) for 1 h, the apoptosis rate and
S phase increased significantly, M phase cells decreased (Fig. 6D). Following treatment with 10 ng/ml
bFGF for a longer time (16 h), Bel-7402 cells showed typical
diploid peak, G1 phase cells increased and the apoptosis rate
decreased significantly (Fig. 6E)
compared with the control cell group (P<0.05 vs. cell control
and wortmannin-treated group), S phase cells increased clearly
compared with the wortmannin-treated group. However, when this
group was pretreated with wortmannin (200 nM) for 1 h, G1 phase
cells decreased apparently, significant hypodiploid peak (sub-G1)
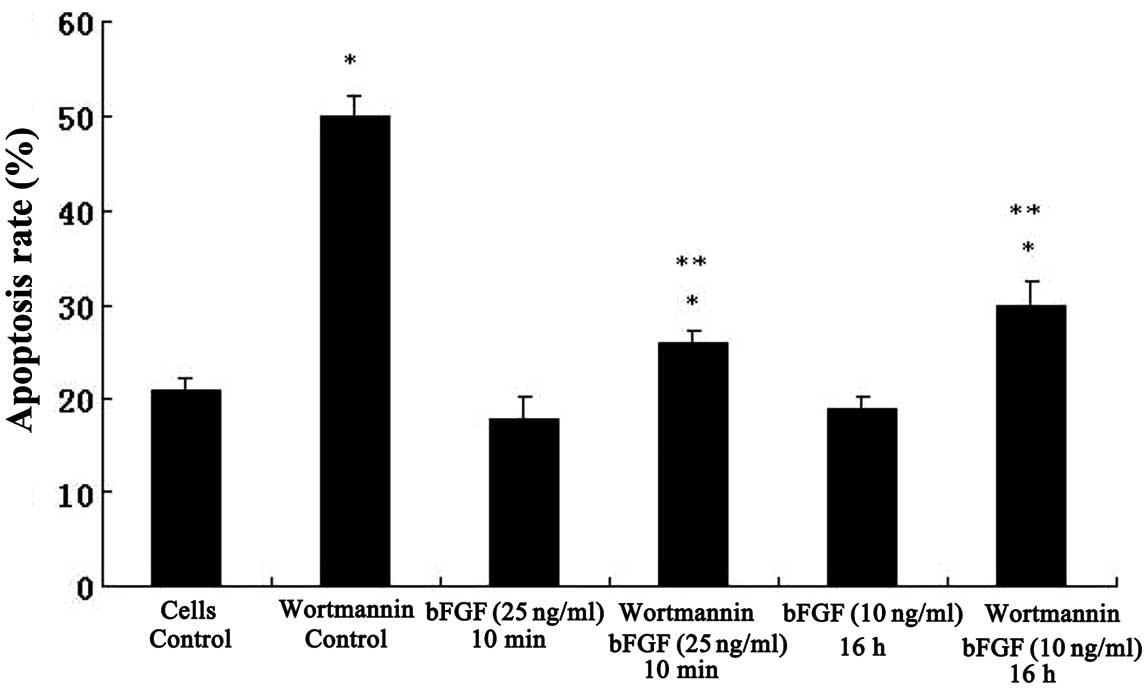
appeared and the apoptosis rate increased significantly (Fig. 6F). The effects of bFGF and
wortmannin on the apoptosis of Bel-7402 cells are also shown in
Fig. 7, demonstrating that
wortmannin induced high apoptosis rates and bFGF inhibited
apoptosis induced by wortmannin.
Discussion
The complex process of tumorigenesis in humans has
been revealed to be a series of stochastic events which occur in
almost all types of human cancer and involve the sequential
acquisition of a number of genetic, epigenetic or
somatic-alterations as a result of increasing genomic instability
(26,27). Cancer cells launch various signaling
pathways to respond to the altered external microenvironment and to
coordinate cell growth with stress responses (28). High levels of TGF-β and bFGF were
found in a high percentage of human lung cancer tissues and have
been associated with poor patient prognosis (29,30).
Although activation of the PBK signaling pathway by TGF-β and bFGF
is known to confer resistance to apoptosis in cancer cells, the
exact mechanism by which this pathway regulates cell survival has
yet to be elucidated.
bFGF in the intracellular transduction pathway has
previously been reported, but, to date, it is not fully understood.
On the one hand, it can play a regulatory role by different
intracellular transduction pathways, such as PLC/PKC, PI3K/PKB,
JAK/STAT (31–33). On the other hand, the role in
regulating is not consistent in different cells; there is a
negative regulatory role in certain cells (particularly tumor
cells), while in other cells is the proliferation of the positive
regulatory factors (34–36). PKB is the major downstream signal
transduction pathway of PI3K. The effect of FGF on PKB activity has
yet to be clarified. Although the point that PKB activation is
required for cell cycle progression remains to be confirmed,
studies show that PKB in some downstream target substances can
directly influence cell cycle progression.
It has been reported that survivin inhibits
apoptosis by suppressing caspase-3 and caspase-7 and modulates the
G2/M phase of the cell cycle through association with mitotic
spindle microtubules (18).
Cytokines and intracellular growth signal transduction can regulate
the expression of survivin. To examine whether bFGF can activate
PKB and survivin via the PI3K pathway in the hepatoma cell line
Bel-7402, we observed the changes of cytoplasmic PKB and survivin
mRNA expression in the Bel-7402 cells with bFGF stimulation and
PI3K pathway inhibitor wortmannin. The results showed that the
expression of PKB and survivin significantly increased with bFGF
(10–25 ng/ml) treatment. These effects were clearly inhibited by
pretreatment with wortmannin (200 nM). bFGF treatment inhibited the
apoptosis rate of Bel-7402 cells and decreased G2/M phase of the
cell cycle, while this effect was blocked by pretreatment with
wortmannin (200 nM) for 1 h. The results showed that wortmannin
could induce high apoptosis and bFGF could inhibit apoptosis
induced by wortmannin. The results revealed that bFGF could rapidly
activate the PKB activities, enhance the expression of survivin and
the proliferation of hepatocarcinoma cells via the PI3K pathway,
thus it may be a novel molecule for early targeting therapy of
hepatocarcinoma.
The ability of bFGF to inhibit apoptosis and promote
survival of cells could be explained by the following pathways:
first, upregulation of Bcl-2 (37–39);
secondly, P13 kinase-Akt/PKB signaling pathway which is further
confirmed by our results (37,40);
the third, upregulation of at least two members of the IAP gene
family, XIAP and survivin. For the third pathway, our results
showed approximately 8-fold upregulation of survivin induced by
bFGF, while PKB was upregulated approximately 3-fold under the same
conditions. Previous studies suggested that human survivin, while
absent in most adult differentiated tissues, could be detected in
almost all transformed cell lines and types of cancer, as well as
in fetal tissue (23). Thus,
survivin behaves as an ‘oncofetal’ protein. Our results therefore
suggest that therapeutically targeting survivin at the gene
expression or protein expression/function levels may result in
suppression of angiogenesis. It is also notable that the cell cycle
dependency of bFGF induced upregulation of survivin expression. Li
et al suggested that survivin may counteract a default
induction of apoptosis in the G2/M phase by associating with the
mitotic spindle at the beginning of mitosis (41). It could be hypothesized that the
bFGF functions in survival and mitogenesis of endothelial cells may
not be mutually exclusive. Indeed, survivin may be upregulated at
the G2/M interface in order to counteract an apoptotic signal,
thereby allowing liver cells to survive cell cycle progression and
to finally proceed to mitosis. This could therefore represent a
tighter association between the mitotic and survival functions.
Based on these results, it is also of interest to determine whether
agents which block bFGF, or bFGF receptor function, inhibit
angiogenesis, in part, by the simultaneous downregulation of
multiple effectors of cell survival such as survivin and XIAP.
Furthermore, certain other antiangiogenic agents or regulators
which are known to induce cell apoptosis, e.g., tubulin-binding
agents such as combretastatin-A4, or endogenous inhibitors of
angiogenesis, such as angiopoietin-2, may do so, at least in part,
by interfering with the survival functions of IAP proteins such as
survivin and XIAP expressed by such cells.
Promoting proliferation is one of the main functions
of bFGF. Survivin is currently the strongest known inhibitor of
apoptosis inhibitory proteins. Our results revealed the correlation
of both signaling pathways. The result is consistent with the
functionality of bFGF and survivin. Following pretreatment with
PI3K inhibitor wortmannin, and then bFGF treatment, cell
proliferation was inhibited, survivin mRNA expression levels
correspondingly decreased, suggesting that bFGF affects the
expression of survivin PKB signaling pathway, which regulates cell
proliferation and apoptosis.
Emerging data indicate a pivotal role for IAP family
members in maintaining cancer cell survival and inhibiting
apoptosis induced by anticancer drugs. Our data provide further
validation of IAP family members as potential drug discovery
targets for the improved treatment of liver cancer. The association
between bFGF, survivin and hepatoma cancer warrants further
investigation.
References
|
1
|
Hu M and Polyak K: Microenvironmental
regulation of cancer development. Curr Opin Genet Dev. 18:27–34.
2008. View Article : Google Scholar
|
|
2
|
Bissell MJ and Radisky D: Putting tumours
in context. Nat Rev Cancer. 1:46–54. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Radisky D, Hagios C and Bissell MJ: Tumors
are unique organs defined by abnormal signaling and context. Semin
Cancer Biol. 11:87–95. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Klagsbrun M and Soker S: VEGF/VPF: The
angiogenesis factor found? Curr Biol. 3:699–702. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Neufeld G, Cohen T, Gengrinovitch S and
Poltorak Z: Vascular endothelial growth factor (VEGF) and its
receptors. FASEB J. 13:9–22. 1999.PubMed/NCBI
|
|
6
|
Tran J, Rak J, Sheehan C, et al: Marked
induction of the IAP family antiapoptotic proteins survivin and
XIAP by VEGF in vascular endothelial cells. Biochem Biophys Res
Commun. 264:781–788. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ouyang G, Liu M, Ruan K, et al:
Upregulated expression of periostin by hypoxia in non-small-cell
lung cancer cells promotes cell survival via the Akt/PKB pathway.
Cancer Lett. 281:213–219. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang L, Bhojani MS, Ross BD and
Rehemtulla A: Molecular imaging of protein kinases. Cell Cycle.
7:314–317. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang L, Lee KC, Bhojani MS, et al:
Molecular imaging of Akt kinase activity. Nat Med. 13:1114–1119.
2007. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kunkel MT, Ni Q, Tsien RY, Zhang J and
Newton AC: Spatio-temporal dynamics of protein kinase B/Akt
signaling revealed by a genetically encoded fluorescent reporter. J
Biol Chem. 280:5581–5587. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Harvey RD and Lonial S: PI3 kinase/AKT
pathway as a therapeutic target in multiple myeloma. Future Oncol.
3:639–647. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang L, Dan HC, Sun M, et al: Akt/protein
kinase B signaling inhibitor-2, a selective small molecule
inhibitor of Akt signaling with antitumor activity in cancer cells
overexpressing Akt. Cancer Res. 64:4394–4399. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Skrzypski M, Kaczmarek P and Le TT:
Effects of orexin A on proliferation, survival, apoptosis and
differentiation of 3T3-L1 preadipocytes into mature adipocytes.
FEBS Lett. 586:4157–4164. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jin P, Wang YH, Peng YG, Hu S, Lu Q and
Yang LY: Effect of PI3K/AKT inhibitor on benign prostate
hyperplasia and its mechanism: an experimental study. Zhonghua Nan
Ke Xue. 16:1068–1075. 2010.(In Chinese).
|
|
15
|
Zhang HJ, Siu MK, Yeung MC, et al:
Overexpressed PAK4 promotes proliferation, migration and invasion
of choriocarcinoma. Carcinogenesis. 32:765–771. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yang Q, Liu HY, Zhang YW, et al:
Anandamide induces cell death through lipid rafts in hepatic
stellate cells. J Gastroenterol Hepatol. 25:991–1001. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Altieri DC and Marchisio C: Survivin
apoptosis: an interloper between cell death and cell proliferation
in cancer. Lab Invest. 79:1327–1333. 1999.PubMed/NCBI
|
|
18
|
Yamamoto T and Tanigawa N: The role of
survivin as a new target of diagnosis and treatment in human
cancer. Med Electron Microsc. 34:207–212. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tamm I, Want Y, Sausville E, et al:
IAP-family protein survivin inhibits caspase activity and apoptosis
induced by Fas (CD95), Bax, caspases, and anticancer drugs. Cancer
Res. 58:5315–5320. 1998.PubMed/NCBI
|
|
20
|
Altieri DC: Survivin versatile modulation
of cell division and apoptosis in cancer. Oncogene. 22:8581–8589.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kajiwara Y, Yamasaki F, Hama S, et al:
Exression of survivin in astrocytic tumors: correlation with
malignant grade and prognosis. Cancer. 97:1077–1083. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Islam A, Kageyama H, Takada N, et al: High
expression of survivin, mapped to 17q25, is significantly
associated with poor prognostic factors and promotes cell survival
in human neuroblastoma. Oncogene. 19:617–623. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ambrosini G, Adida C and Altieri D: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ngan CY, Yamamoto H, Takagi A, et al:
Oxaliplatin induces mitotic catastrophe and apoptosis in esophageal
cancer cells. Cancer Sci. 99:129–139. 2008.PubMed/NCBI
|
|
25
|
Huang CK, Lee SO, Lai KP, et al: Targeting
androgen receptor in bone marrow mesenchymal stem cells leads to
better transplantation therapy efficacy in liver cirrhosis.
Hepatology. 57:1550–1563. 2013. View Article : Google Scholar
|
|
26
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar
|
|
27
|
Hahn WC and Weinberg RA: Modelling the
molecular circuitry of cancer. Nat Rev Cancer. 2:331–341. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
López-Maury L, Marguerat S and Bähler J:
Tuning gene expression to changing environments: from rapid
responses to evolutionary adaptation. Nat Rev Genet. 9:583–593.
2008.PubMed/NCBI
|
|
29
|
Tateishi M, Ishida T, Mitsudomi T, Kaneko
S and Sugimachi K: Immunohistochemical evidence of autocrine growth
factors in adenocarcinoma of the human lung. Cancer Res.
50:7077–7080. 1990.PubMed/NCBI
|
|
30
|
Brattstrom D, Bergqvist M, Hesselius P,
Larsson A, Wagenius G and Brodin O: Serum VEGF and bFGF adds
prognostic information in patients with normal platelet counts when
sampled before, during and after treatment for locally advanced
non-small cell lung cancer. Lung Cancer. 43:55–62. 2004. View Article : Google Scholar
|
|
31
|
Sufen G, Xianghong Y, Yongxia C and Qian
P: bFGF and PDGF-BB have a synergistic effect on the proliferation,
migration and VEGF release of endothelial progenitor cells. Cell
Biol Int. 35:545–551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Drevs J, Zirrgiebel U and Schmidt-Gersbach
CI: Soluble markers for the assessment of biological activity with
PTK787/ZK 222584 (PTK/ZK), a vascular endothelial growth factor
receptor (VEGFR) tyrosine kinase inhibitor in patients with
advanced colorectal cancer from two phase I trials. Ann Oncol.
16:558–565. 2005. View Article : Google Scholar
|
|
33
|
Feng X, Zhang B, Wang J, et al:
Adenovirus-mediated transfer of siRNA against basic fibroblast
growth factor mRNA enhances the sensitivity of glioblastoma cells
to chemotherapy. Med Oncol. 28:24–30. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang H, Rubin M, Fenig E, et al: Basic
fibroblast growth factor causes growth arrest in MCF-7 human breast
cancer cells while inducing both mitogenic and inhibitory G1
events. Cancer Res. 57:1750–1757. 1997.PubMed/NCBI
|
|
35
|
Shankland SJ, Pippin J, Flanagan M, et al:
Mesangial cell proliferation mediated by PDGF and bFGF is
determined by levels of the cyclin kinase inhibitor p27Kip1. Kidney
Int. 51:1088–1099. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Johnson MR, Valentine C, Basilio C and
Mansukhani A: FGF signaling activates STAT1 and p21 and inhibits
the estrogen response and proliferation of MCF-7 cells. Oncogene.
16:2647–2656. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Carmeliet P, Lampugnani MG, Moons L, et
al: Targeted deficiency or cytosolic truncation of the VE-cadherin
gene in mice impairs VEGF-mediated endothelial survival and
angiogenesis. Cell. 98:147–157. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim JW, Ho WJ and Wu BM: The role of the
3D environment in hypoxia-induced drug and apoptosis resistance.
Anticancer Res. 31:3237–3245. 2011.PubMed/NCBI
|
|
39
|
Gerber HP, Dixit V and Ferrara N: Vascular
endothelial growth factor induces expression of the antiapoptotic
proteins Bcl-2 and A1 in vascular endothelial cells. J Biol Chem.
273:13313–13316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gerber HP, McMurtrey A, Kowalski J, Yan M,
Keyt BA, Dixit V and Ferrara N: Vascular endothelial growth factor
regulates endothelial cell survival through the
phosphatidylinositol 3′-kinase/Akt signal transduction pathway.
Requirement for Flk-1/KDR activation. J Biol Chem. 273:30336–30343.
1998.
|
|
41
|
Li F, Ambrosini G, Chu EY, Plescia J, et
al: Control of apoptosis and mitotic spindle checkpoint by
survivin. Nature. 396:580–584. 1998. View
Article : Google Scholar : PubMed/NCBI
|