Introduction
Lung cancer ranks highly among the leading causes of
cancer-related mortality worldwide, with non-small cell lung cancer
(NSCLC) accounting for ~75–85% of all lung malignancies (1). Despite significant advances in
oncology research, 5-year survival rate of NSCLC does not exceed
16% (2). Gaining insight into the
complex molecular mechanisms underlying the pathogenesis of NSCLC
is essential for the identification of prognostic and predictive
biological markers that may be targeted to improve clinical
outcome.
The phosphoinositide 3-kinase (PI3K)/v-akt murine
thymoma viral oncogene homolog (AKT)/mammalian target of rapamycin
(mTOR) signalling pathway is emerging as a common theme in human
carcinogenesis. mTOR coordinates basic cellular functions including
cellular growth and proliferation (3). The aberrant activation of the pathway
presents as either gain of growth-promoting function or loss of
inhibitory function, facilitating neoplastic transformation. The
engagement of receptor tyrosine kinases (RTKs) [epidermal growth
factor receptor (EGFR), fibroblast growth factor receptor (FGFR),
insulin-like growth factor-I receptor (IGF-IR) and platelet-derived
growth factor receptor (PDGFR)], as well as Ras activate PI3K, thus
leading to the generation of phosphatidylinositol triphosphate
(PIP3) and in turn to the recruitment of AKT into the cell membrane
for phosphorylation at threonine (Thr)308 and serine (Ser)473
(4). Once activated, AKT moves into
the cytoplasm and the nucleus, where it phosphorylates, activates,
or inhibits a number of downstream targets, regulating various
cellular functions including cellular metabolism, protein
synthesis, cell survival/inhibition of apoptosis and cell cycle
progression. Among the various signalling cascades emanating from
AKT, mTOR functions prominently as a critical downstream effector.
mTOR is a serine/threonine kinase existing in 2 functionally
distinct complexes-mTORC1 and mTORC2 (5). mTORC1 is sensitive to rapamycin and
constitutes a major regulator of ribosomal biogenesis and protein
synthesis, through the activation of the promoter of mRNA
translation, S6K, and inactivation of the repressor of mRNA
translation, 4E-BP1 (6). The
phosphorylation of 4E-BP1 disrupts its interaction with eIF4E,
allowing it to interact with eIF4G and form the eIF4F complex that
enhances the overall translational machinery of the cell (7). mTORC2 is insensitive to rapamycin,
phosphorylates AKT and regulates cell survival, migration and the
actin cytoskeleton. Hence, mTORC1 and mTORC2 have different
physiological functions (8). mTORC1
can be phosphorylated and activated by AKT directly or indirectly
through the phosphorylation and subsequent inactivation of tuberous
sclerosis complex 2 (TSC2) (9).
PI3K proteins, of which PI3Ka is the most relevant
in the context of cancer, are heterodimers, consisting of a p110
catalytic subunit and a p85 regulatory subunit, which in the basal
state, restricts the activity of the catalytic subunit (9). Oncogenic mutations of PIK3CA
disrupt the inhibitory function of the p85 subunit, resulting in
the unrestricted constitutive activity of p110 (10) with the consequent upregulation of
PI3K/AKT signalling and are commonly encountered in human cancers.
In NSCLC however, such mutations are uncommon (11,12).
The aberrant activation of PI3K may also be secondary to the
loss-of-function gene alterations of phosphatase and tensin homolog
(PTEN)(13). PTEN
antagonises the ability of PI3K to phosphorylate AKT by removing
phosphates at position 3 of PIP3, thereby serving as a tumour
suppressor (14).
The PI3K/AKT/mTOR pathway is constitutively active
in NSCLC (15,16), not only due to PTEN inactivation but
also to copy number gains of PIK3CA(12). In contrast to the low frequency of
mutations, PIK3CA copy number gains have been reported in
33.1% of squamous cell lung cancer and in 6.2% of lung
adenocarcinomas (11). In
vitro data indicates that PI3K signalling mediates
bronchioalveolar stem cell expansion initiated by oncogenic
K-RAS, whereas oncogenic PIK3CA mutations enhance the
anchorage-independent growth and migration activity of immortalised
respiratory epithelial cells (17).
More importantly, the inhibition of PI3K/AKT/mTOR signalling
through pharmacologic and genetic approaches induces the
anti-proliferative effects on certain NSCLC cell lines, as well as
in mouse models of lung cancer (12).
On the basis of this experimental evidence, it is
not surprising that much effort has been devoted to the
investigation of the expression profiles of various components of
the PI3K/AKT/mTOR pathway in NSCLC (16,18–32),
sometimes yielding inconsistent results. In particular, a
comprehensive analysis of all components of this pathway, along
with the PIK3CA, K-RAS, AKT1 and PTEN mutational
status has not been performed thus far. Therefore, the aims of the
present study were as follows: first, to search for any correlation
between the mutational status of these 4 genes and the expression
levels of the p85α and p110γ subunits of PI3K, phosphorylated AKT
(p-AKT), phosphorylated mTOR (p-mTOR), PTEN, phosphorylated p79S6K
(p-p79S6K) and phosphorylated 4E-BP1 (p-4E-BP1); second, to examine
the associations of these molecules with clinicopathological
characteristics; and third, to assess their potential prognostic
utility in a series of NSCLC patients.
Materials and methods
Cases and classification
This is a study of 102 adult Caucasian patients of
Greek ethnicity with NSCLC (36)
diagnosed between 2004 and 2010 at the First Department of
Pathology, University of Athens Medical School, for whom archival
primary tumour material at diagnosis, prior to chemo/radiotherapy,
was available. The study was approved by the Ethics Committee of
the University of Athens Medical School and informed consent was
obtained from each patient prior to enrolment in the study. The
demographic data and follow-up information of our patients are
shown in Table I. The total sample
set consisted of 71 surgical specimens and 31 biopsies and was
composed of 14 females and 88 males with a median age of 71 years
(range, 46–89). The pathological stage of each tumour was assigned
following the guidelines from the 7th edition of the TNM
classification (1) and was known
for 71 patients: 23 patients had stage I disease and 48 had stage
II-III disease. The medical records of the patients were collected
to obtain information regarding patient survival status and cause
of death was obtained from the medical records and/or interview
with the family. Follow-up information was available for 75
patients.
 | Table IClinicopathological characteristics
of the 102 patients enrolled in the present study. |
Table I
Clinicopathological characteristics
of the 102 patients enrolled in the present study.
| Characteristic | No. of patients
(%) |
|---|
| Gender |
| Female | 14 (13.73) |
| Male | 88 (86.27) |
| Histological
type |
|
Adenocarcinoma | 39 (38.24) |
| Squamous cell
carcinoma | 48 (47.06) |
| Adenosquamous
carcinoma | 9 (8.82) |
| Large cell
carcinoma | 3 (2.94) |
| Sarcomatoid
carcinoma | 3 (2.94) |
| Histological
grade |
| I | 3 (2.94) |
| II | 47 (46.08) |
| III | 52 (50.98) |
| Stage |
| I | 23 (32.39) |
| II | 35 (49.30) |
| III | 13 (18.31) |
| Tumour status |
| T1 | 10 (14.08) |
| T2 | 41 (57.75) |
| T3 | 20 (28.17) |
| Nodal status |
| N0 | 50 (70.42) |
| N1 | 19 (26.76) |
| N2 | 2 (2.82) |
| Recurrence
rate |
| Absence | 27 (49.09) |
| Presence | 28 (50.91) |
| Follow-up |
| Alive | 68
(66.67)
Follow-up: 33 (2–108) months |
| Dead from
disease | 34
(33.34)
Follow-up: 12 (1–96 months) |
| Age |
| Median
(range) | 71 (46–89) |
PIK3CA, AKT1, PTEN and K-RAS mutational
analysis DNA extraction from paraffin-embedded tissues
Sections (10-μm-thick) were cut from
paraffin-embedded tissue blocks following tumour enrichment under a
light microscope. DNA was extracted from the selected tissue areas
following a standard DNA extraction kit protocol (NucleoSpin
Tissue; Macherey-Nagel, Duren, Germany). The extracted DNA was
quantified on a Picodrop Microliter spectrophotometer.
High resolution melting analysis
(HRMA)
PIK3CA exons 9 and 20, AKT1 exon 4,
PTEN exons 7 and 8, as well as K-RAS codons 12 and 13
were screened for mutations in 61 tumour specimens using HRMA on a
LightCycler 480 (obtained from Roche Diagnostics, GmbH, Mannheim,
Germany). Each reaction consisted of 20 ng DNA, 200 nmol/l of each
primer, 10 μl LightCycler 480 HRM Master Mix (Roche) and 3.5 mM
MgCl2, in a total volume of 20 μl. The profile used in
the Light Cycler was: 95ºC for 10 min, followed by 50 cycles of
95ºC for 10 sec, 58–60ºC for 15 sec and 72ºC for 7 sec. DNA samples
from colorectal cancers or cell lines harbouring K-RAS codon
12 and 13 mutations or exon 9 and 20 PIK3CA mutations were
used as the positive controls for HRM analysis. In detail, for
PIK3CA exon 20 mutational analysis we examined DNA extracted
from the HCT-116 colon cancer cell line (PIK3CA mutation,
p.H1047R). For the other genes, previously identified mutant
samples from colorectal cancers were used. The sequences of the
primers for the K-RAS, PIK3CA and AKT1 genes have
been published previously (33).
The primers used for PTEN gene analysis were as follows:
PTEN exon 7 forward, 5′-TCGTTTTTGACAGTTT GACAGTT-3′ and
reverse, 5′-GGATATTTCTCCCAATGAAA GTAAA-3′; and PTEN exon 8
forward, 5′-GTCATTTCATTTC TTTTTCTTTTCTTT-3′ and reverse,
5′-CAACAACCCCCA CAAAT-3′.
Sequencing, pyrosequencing and
restriction fragment length polymorphism (RFLP) analysis
PIK3CA (exons 9 and 20) and PTEN
(exons 7 and 8) PCR products shown to be positive by HRM, were
sequenced using the BigDye Terminator Cycle Sequencing kit (Applied
Biosystems, Foster City, CA, USA). The sequencing products were
analysed on an ABI PRISM 310 Genetic Analyzer (Applied Biosystems).
PCR primers were also used for sequencing analysis to confirm the
presence of mutations. The results were verified by the sequencing
of at least 2 independent PCR products. K-RAS mutations at
codon 12 were verified by RFLP analysis as described previously
(34). AKT1 mutations were
identified using pyrosequencing with the Q24 pyrosequencer
according to the manufacturer’s instructions (Qiagen GmbH, Hilden,
Germany).
Immunohistochemistry
Immunostaining was performed on paraffin-embedded
4-μm-thick sections of formalin-fixed tumour tissue using the
two-step peroxidase conjugated polymer technique (Dako Envision
kit; Dako, Carpinteria, CA, USA). The primary antibodies used are
listed in Table II. In the
negative controls primary antibodies were substituted with
non-immune serum.
| Table IICharacteristics of primary antibodies
used in immunohistochemical analysis. |
Table II
Characteristics of primary antibodies
used in immunohistochemical analysis.
| Protein | No. of cases | Clone | Company | Catalog no. | Raised in | Positive
controls | Antigen retrieval
method | Dilution and
incubation time for immunohistochemistry |
|---|
p-mTOR
(Ser2448) | 102 | Monoclonal | Cell Signaling
Technology, USA | 2976 | Rabbit | Human breast
cancer | Citrate buffer, pH
9.0 | 1:50,
overnight |
p-p70S6K
(Thr421/Ser424)
(specific for p70 subunit) | 100 | Polyclonal | Santa Cruz
Biotechnology, USA | sc-7984-R | Rabbit | Human colon
carcinoma | Citrate buffer, pH
6.0 | 1:250,
overnight |
p-4E-BP1
(Thr37/46) 236B4 | 102 | Monoclonal | Cell Signaling
Technology, USA | 2855 | Rabbit | Human breast
cancer | Citrate buffer, pH
6.0 | 1:800,
overnight |
AKT-pS473
(phosphorylation site-specific) | 102 | Polyclonal, clone
14–5 | Dako,
Dakopatts | M3628 | Rabbit | Mouse kidney | Citrate buffer, pH
9.0 | 1:20, 1 h |
p85a
(B-9)
(specific for α subunit of p85) | 102 | Monoclonal | Santa Cruz
Biotechnology, USA | sc-1637 | Mouse | Normal tonsillar
tissue | Citrate buffer, pH
9.0 | 1:50,
overnight |
p110γ
(specificity for γ subunit of p110) | 102 | Polyclonal | Santa Cruz
Biotechnology, USA | Sc-7177 | Mouse | Normal bone
marrow | Citrate buffer, pH
6.0 | 1:50,
overnight |
PTEN
(FL-403) | 102 | Polyclonal | Santa Cruz
Biotechnology, USA | sc-9145 | Rabbit | Normal tonsillar
tissue | Citrate buffer, pH
9.0 | 1:80,
overnight |
Two pathologists (Eleni Trigka and Penelope
Korko-lopoulou) viewed the first 20 cases for each antibody
together to obtain the consensus regarding the evaluation, without
knowledge of the clinical data. The remaining cases were then
examined by a pathologist (Eleni Trigka) and another 20 cases were
examined for reproducibility by a second pathologist (Penelope
Korkolopoulou). Nuclear and cytoplasmic immunoreactivity was
recorded separately. For statistical analysis only the predominant
pattern i.e., cytoplasmic for p-mTOR and PTEN and nuclear for
p-p70S6K, p85αPI3K and p-4E-BP1 was taken into account. For p-AKT
and p110γPI3K both nuclear and cytoplasmic expression patterns were
analysed separately. A Histo-score (H-score) based on the
percentage of stained neoplastic cells multiplied by the staining
intensity was calculated, as previously described (35).
Statistical analysis
Statistical analysis was performed by a
biostatistician (Georgia Levidou). In the basic statistical
analysis, p-mTOR, p-p70S6K, PTEN, p-AKT, p4E-BP1, p110γPI3K and
p85αPI3K H-scores were treated as continuous variables.
Correlations among the immunohistochemical expression of the
investigated proteins as well as with the clinicopathological
parameters were examined using non-parametric tests with correction
for multiple comparisons where necessary (Kruskal-Wallis ANOVA,
Mann-Whitney U test, Spearman’s rank correlation co-efficient,
Fisher’s exact test and χ2 as appropriate).
Survival analysis was performed using death by
disease as the endpoint for cancer-specific survival (CSS). The
effect of various parameters on clinical outcome was assessed by
comparing the groups using the log-rank test. Multivariate analysis
was performed using Cox’s model. For statistical calculations the
statistical package STATA 11.0 for Windows was used. All results
with a two-sided p-value ≤0.05 were considered to indicate
statistically significant differences.
Results
PIK3CA, AKT1, PTEN and K-RAS mutations
(Fig. 1)
Overall, 61 cases were analysed for the presence of
PIK3CA, AKT1, PTEN and K-RAS mutations. Molecular
analysis was successful as regards the presence of activating
mutations at exons 9 and 20 of the PIK3CA gene in 60 and 59
cases, respectively, whereas mutations at codons 12 and 13 of the
K-RAS gene were observed in 54 cases, at exon 4 of
AKT1 gene in 57 cases and at exons 7 and 8 of PTEN
gene in 50 and 49 cases, respectively, by high resolution melting
analysis. PI3KCA and PTEN mutations were identified
by sequencing analysis, AKT1 by pyrosequencing, whereas
K-RAS mutations at codon 12 were verified by RFLP analysis.
PIK3CA mutations were observed in 5% of the examined samples
(3/59). In particular, 1 sample displayed a G>A transition at
nucleotide 3052 (c.3052G>A) leading to an aspartic acid to
asparagine substitution (p.D1018N), which is rare and has been
previously identified in thyroid cancer [Sanger Institute Catalogue
Of Somatic Mutations In Cancer (COSMIC) database] (Fig. 1A). In addition, 2 samples displayed
a common PIK3CA mutation at exon 9, identified as a G>A
transition at nucleotide 1633 (c.1633G>A) leading to a glutamic
acid to lysine substitution (p.E545K) (Fig. 1B). Two PIK3CA mutations (in
exons 9 and 20) were observed in squamous cell lung carcinomas and
1 in an adenocarcinoma. As regards AKT1 mutations, the most
frequent alteration, namely a G>A transition at nucleotide 49
(c.49G>A), causing glutamic acid to lysine substitution at
hotspot codon 17 (p.E17K) was found in 1 adenocarcinoma case (1/57
of the examined cases, 1.75%) (Fig.
1C). Furthermore, 10 out of the 54 examined cases (18.5%) were
positive for K-RAS mutations at codon 12. PTEN
mutations were also encountered in 5 cases, 1 in exon 7 (1/50, 2%)
and 4 in exon 8 (4/49, 8.2%). In detail, a PTEN exon 7
C>T transition (c.654 C>T) leading to a silent mutation
(p.C218C) was found in 1 squamous carcinoma (Fig. 1D). Accordingly, as far as
PTEN exon 8 is concerned, a C>T transition (c.1016C>T)
leading to a proline to leucine substitution at codon 339 (p.P339L)
was found in 1 adenocarcinoma case (Fig. 1E). Furthermore, a G>A
substitution (c.881G>A) at codon 881 leading to p.S294N mutation
and a C>T (c.883 C>T) silent mutation (p.L295L) were found in
2 squamous cell carcinoma cases. Finally, an adenosquamous
carcinoma case showed a C>T silent mutation at codon 850
(c.850C>T), p.T286T). Notably, 4 out of 5 PTEN mutations
observed were C>T substitutions. In our cohort, PIK3CA
mutations were not found to co-exist with K-RAS, AKT1 or
PTEN mutations. The single AKT1 mutant case displayed
one of the highest nuclear p110γPI3K H-scores (i.e., 20). Moreover,
all cases with a PTEN mutation displayed p110γPI3K
cytoplasmic immunoreactivity (p=0.0506). Finally, the presence of
PTEN mutations correlated with the presence of nodal
metastasis (Fisher’s exact test, p=0.047).
PI3K/AKT/mTOR pathway in NSCLCs (Fig. 2)
p-mTOR immunoreactivity was cytoplasmic and/or
membranous in 58/102 cases (56.86%) and nuclear in 7/102 cases
(6.8%). Nuclear p-p70S6K and p-4E-BP1 immunoexpression was observed
in 96/100 cases (96%) and 71/102 cases (69.61%).
Tumour-infiltrating lymphocytes and endothelial cells expressed
both proteins and served as the internal positive controls in each
case. Sixty-eight cases (68/100, 68%) co-expressed p-p70S6K and
p-mTOR, with 28 p-p70S6K positive cases being negative for p-mTOR
(28/96, 21.16%). All these cases but one, however, displayed
cytoplasmic p-AKT (27/28, 96.42%). Fifty-three cases co-expressed
p-mTOR and p-4E-BP1 (53/102, 51.96%), whereas 18 cases positive for
p-4E-BP1 were negative for p-mTOR (18/71, 25.35%). The percentage
of co-expression of all 3 components of the mTOR cascade rose to
51% (51/100 cases). Two cases (2/100, 2%) were triple-negative for
the p-mTOR pathway components, whereas all the cases that were
positive for p-mTOR but double negative for p-p70S6K and p-4E-BP1
expressed p-AKT.
p-AKT immunoreactivity was mainly cytoplasmic
(95/102, 93.14%) with 85 cases (85/102, 83.33%) showing nuclear
immunoexpression. Cytoplasmic p-AKT was co-expressed with all 3
mTOR pathway components in 49/100 cases (49%) and nuclear in 45/100
(45%) cases. p85αPI3K nuclear expression was recorded in 58/102
cases (56.86%). p85αPI3K was co-expressed with cytoplasmic p-AKT in
56/102 (54.9%) cases and with nuclear p-AKT in 48/102 (47.05%)
cases. p85αPI3K-positive cases displayed higher levels of p-AKT
cytoplasmic immunoreactivity (Mann-Whitney U test, p=0.0442). The
co-expression of p85αPI3K with cytoplasmic p-AKT and all 3 members
of the mTOR pathway was observed in 33 cases (33/100, 33%).
p110γPI3K cytoplasmic immunoreactivity was observed in 42/102
(41.185) cases, whereas only 19.61% (20/102) of cases displayed
nuclear immunoexpression. All p110γPI3K-positive cases displayed
p-AKT cytoplasmic immunoexpression (Fisher’s exact test, p=0.039).
The co-expression of cytoplasmic or nuclear p110γPI3K with
cytoplasmic p-AKT and all 3 members of the mTOR pathway was
observed in 22 (22/100, 22%) and 15 (15/100, 15%) cases,
respectively. PTEN cytoplasmic immunoreactivity was observed in
79/102 cases (77.45%) with 6 cases showing nuclear immunoreactivity
(6/102, 5.8%). Twenty-one of the cases that displayed cytoplasmic
p-AKT immunoreactivity (21/95, 22.1%) were negative for PTEN.
The correlations among these proteins are shown in
Table III. A significant positive
correlation was established between cytoplasmic p-AKT and p-mTOR,
p-p70S6K or p-4E-BP1, between p85αPI3K and p-mTOR, p-p70S6K or
p-4E-BP1 as well as between p-mTOR and p-p70S6K, p-p70S6K and
p-4E-BP1. p110γPI3K nuclear and cytoplasmic immunoreactivity both
positively correlated with p-mTOR, p-p70S6K or p-4E-BP1, whereas a
positive correlation was observed between nuclear p110γPI3K and
p85αPI3K immunoexpression.
| Table IIIAssociations among the molecules
under study in the entire cohort. |
Table III
Associations among the molecules
under study in the entire cohort.
| p85aPI3K
H-score | p110γPI3K
cytoplasmic H-score | p110γPI3K nuclear
H-score | PTEN H-score | p-AKT cytoplasmic
H-score | p-mTOR H-score | p-p70S6K
H-score | p-4E-BP1
H-score |
|---|
| p110γPI3K
cytoplasmic H-score | R=0.2988 | | | | | | | |
| p=0.0023 | | | | | | | |
| p110γPI3K nuclear
H-score |
R=0.3620 | R=0.1406 | | | | | | |
|
p=0.0002 | p>0.10 | | | | | | |
| PTEN H-score | R=−0.0452 | R=0.1172 | R=0.0739 | | | | | |
| p>0.10 | p>0.10 | p>0.10 | | | | | |
| pAKT cytoplasmic
H-score |
R=0.2204 | R=0.1195 | R=0.0752 | R=0.1259 | | | | |
|
p=0.0276 | p>0.10 | p>0.10 | p>0.10 | | | | |
| p-mTOR H-score | R=0.0634 |
R=0.1771 |
R=0.1861 |
R=0.2620 |
R=0.2450 | | | |
| p>0.10 |
p=0.0750 |
p=0.0611 |
p=0.0085 |
p=0.0140 | | | |
| p-p70S6K
H-score |
R=0.4323 |
R=0.2159 |
R=0.3329 | R=0.0954 |
R=0.3283 |
R=0.2884 | | |
|
p<0.0001 |
p=0.0310 |
p=0.0007 | p>0.10 |
p=0.0009 |
p=0.0036 | | |
| p-4E-BP1
H-score |
R=0.4256 |
R=0.1605 |
R=0.1777 | R=−0.0403 |
R=0.3016 | R=0.1199 |
R=0.5443 | |
|
p<0.0001 | P=0.1005a |
p=0.0740 | p>0.10 |
p=0.0023 | p>0.10 |
p<0.0001 | |
| p-AKT nuclear
H-score | R=0.1327 | R=0.0530 | R=0.0127 | R=−0.0782 |
R=0.4384 | R=0.1051 |
R=0.1791 | R=0.125 |
| p>0.10 | p>0.10 | p>0.10 | p>0.10 |
p<0.0001 | p>0.10 |
p=0.0746 | p>0.10 |
p-mTOR, p-p70S6K, p85αPI3K and p-AKT were negative
in the alveolar epithelium but weakly positive in seromucous glands
and normal bronchi. p110γPI3K was also negative in the alveolar
epithelium, whereas a nuclear immunoreactivity was observed in the
normal bronchial epithelium. PTEN was present in the alveolar and
normal bronchial epithelium, as well as in seromucous glands.
p-4E-BP1 was absent in the normal lung epithelium.
Association between PI3K/AKT/mTOR pathway
components and clinicopathological features (Table IV and Fig. 3)
p-mTOR immunoreactivity prevailed in adenocarcinomas
(Kruskal Wallis, ANOVA, p=0.0001), whereas p-4E-BP1 H-score was
lower in this histological subtype (Kruskal Wallis, ANOVA,
p=0.0271) when compared to the remaining tumour types. In
particular, p-mTOR expression was significantly higher in
adenocarcinomas when compared to squamous cell carcinomas
(Mann-Whitney U test, Bonferoni correction, p=0.0416), whereas
p-4E-BP1 was significantly higher in squamous cell carcinomas when
compared to adenocarcinomas (Mann-Whitney U test, Bonferoni
correction, p=0.0032). Moreover, p110γPI3K nuclear positivity was
more frequently encountered in adenocarcinomas (12/39, 30.77%)
compared to the remaining tumour histological types (8/63, 12.7%),
(χ2 test, p=0.025).
Furthermore a positive correlation was observed
between p-4E-BP1 or p-AKT nuclear immunoexpression and tumour
status (Kruskal Wallis, ANOVA, T1 vs. T2 vs. T3, p=0.0033) or stage
(Mann-Whitney U test, p=0.0482), respectively, whereas the
respective correlation between p-p70S6K immunoreactivity and tumour
status was an inverse correlation and of marginal significance
(Kruskal Wallis, ANOVA, T1 vs. T2 vs. T3, p=0.0787). The former
correlation remained in adenocarcinomas. p85αPI3K also correlated
with tumour status, with the T2 tumours exhibiting higher levels of
expression (Kruskal Wallis, ANOVA, T1 vs. T2 vs. T3, p=0.0383) and
marginally correlated with nodal status in squamous cell carcinomas
(p=0.0887). p110γPI3K immunoreactivity also positively correlated
with the presence of nodal metastasis (Mann-Whitney U test,
p=0.0951).
Inverse correlations, although of borderline
significance were also obtained between histological grade and
PTEN, p-mTOR and p-p70S6K immunoreactivity (Mann Whitney U test,
I/II vs. III, p=0.0798, p=0.0987 and 0.0816, respectively), the
latter being significant in squamous cell carcinomas
(p=0.0298).
Survival analysis
Univariate survival analysis was performed for the
entire cohort and separately for adenocarcinomas and squamous cell
carcinomas. The results are shown in Table V.
| Table VResults of univariate survival
analysis for cancer-specific survival (log-rank test). |
Table V
Results of univariate survival
analysis for cancer-specific survival (log-rank test).
| Variable | Entire cohort
p-value | Adenocarcinoma
p-value | SCC p-value |
|---|
| Age | 0.9243 | 0.1468 | 0.8511 |
| Gender (Fenale vs.
male) | 0.7871 | 0.5917 | 0.4468 |
| Histological
type | 0.4559 | - | - |
| Histological
grade | 0.8496 | 0.4102 | 0.8404 |
| Stage (I vs.
II/III) | 0.4824 | 0.6171 | 0.2716 |
| Tumour status (T1
vs. T2 vs. T3) | 0.8289 | 0.5127 | 0.9425 |
| Nodal status (N0
vs. N1/N2) | 0.4559 | 0.4142 | 0.4059 |
| PTEN
mutations (absence vs. presence) | 0.4669 | 0.5892 | 0.1284 |
| K-RAS
mutations (absence vs. presence) | 0.5892 | 0.5596 | 0.7389 |
| PIK3CA
mutations (absence vs. presence) | 0.7434 | 0.7228 | 0.8348 |
| AKT1
mutations (absence vs. presence) | 0.6305 | 0.6821 | a |
| p85aPI3K H-score
(negative vs. positive) | 0.6983 | 0.2360 | 0.4510 |
| p110γPI3K
cytoplasmic H-score (negative vs. positive) | 0.2964 | 0.2964 | 0.4113 |
| p110γPI3K nuclear
H-score (negative vs. positive) | 0.9352 | 0.9540 | 0.5390 |
| PTEN H-score
(<10 vs. ≥10) | 0.8559 | 0.7343 | 0.7916 |
| p-AKT cytoplasmic
H-score (<90 vs. ≥90) | 0.7398 | 0.0657 | 0.3678 |
| p-AKT nuclear
H-score (<2.125 vs. ≥2.125) | 0.2781 | 0.5245 | 0.2774 |
| p-mTOR H-score
(<2 vs. ≥2) | 0.2752 | 0.7156 | 0.2890 |
| p-p70S6K H-score
(<77.5vs. ≥77.5) | 0.7273 | 0.1715 | 0.4448 |
| p-4E-BP1 H-score
(negative vs. positive) | 0.3202 | 0.0306 | 0.7362 |
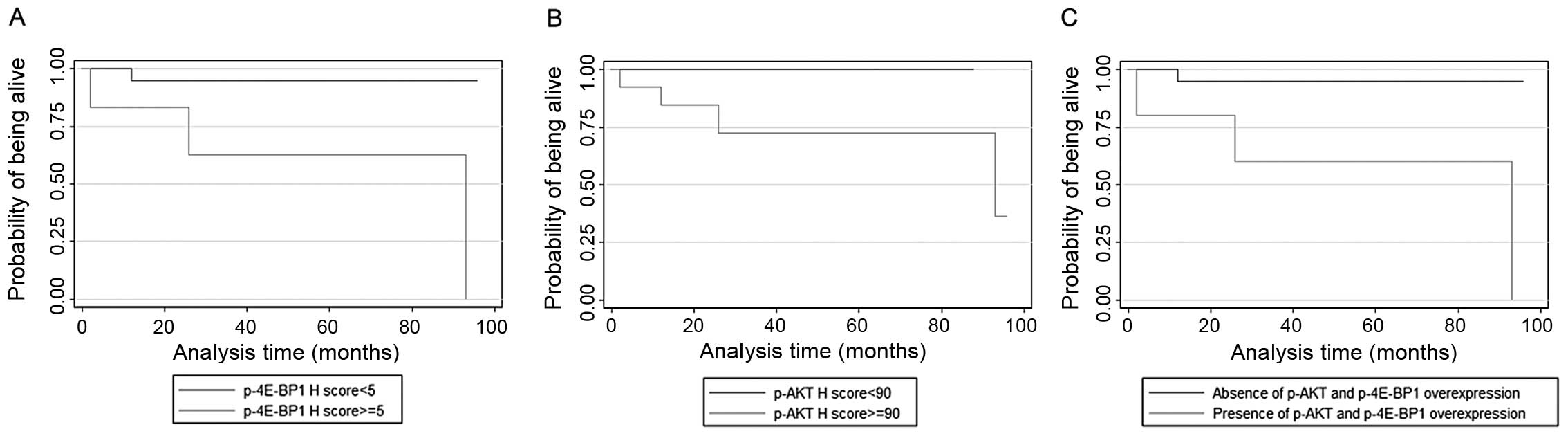
The only parameter that adversely affected survival
in adenocarcinomas was p-4E-BP1 immunoreactivity (p=0.0306,
Fig. 4A), with cytoplasmic p-AKT
expression attaining a marginal significance in this regard
(p=0.0657, Fig. 4B). In addition,
we evaluated the prognostic effect of the combination of these 2
differentially activated proteins using Kaplan-Meier analysis.
Subjects with adenocarcinomas showing p-AKT and p-4E-BP1
overexpression had significantly worse prognosis than the other 2
combinations (p=0.0252, Fig.
4C).
Multivariate survival analysis was performed for the
entire cohort. In order to improve the stability and power, we
adjusted a multivariate model including the combination of p-AKT
and p-4E-BP1 overexpression along with histological grade and stage
and an interaction term between concurrent p-AKT and p-4E-BP1
overexpression and histological type. This model is presented in
Table VI. In particular, the
combination of p-AKT and p-4E-BP1 overexpression was proven to be
an independent predictor of adverse prognosis, with the interaction
term being significant, validating the assumption of a significant
interaction between the effect of these molecules on survival and
histological type.
| Table VICox proportional hazards estimation
model for the entire cohort. |
Table VI
Cox proportional hazards estimation
model for the entire cohort.
| Hazard ratio
(HR) | p-value | 95% confidence
interval of HR |
|---|
| Concurrent
overexpression of p-AKT-p-4E-BP1 | 13.419 | 0.036 | 1.184 | 152.121 |
| Histological type
(adenocarcinoma vs. other type) | 5.160 | 0.133 | 0.608 | 43.810 |
| Interaction
terma | 0.035 | 0.024 | 0.002 | 0.641 |
| Histological
grade | 1.052 | 0.932 | 0.333 | 3.322 |
| Stage | 1.009 | 0.992 | 1.156 | 6.526 |
Discussion
The PI3K/AKT/mTOR pathway has attracted broad
scientific and clinical interest in NSCLC, not only due to the fact
that its aberrant activation promotes resistance to
chemo/radiotherapy in vitro(15), but also since its components can be
targeted for therapeutic purposes. Given the relatively common
occurrence of PTEN and EGFR gene alterations, NSCLC
has logically been considered a suitable candidate for the
therapeutic use of molecules (e.g., rapamycin) inhibiting the
PI3K/AKT/mTOR cascade (36).
However, the results of previous clinical trials have tempered
initial optimism showing that mTOR inhibitors have a limited
efficacy as single agents in NSCLC (37). The insufficiency of therapeutic
response may be somewhat related to the non-specific selection of
patients for m-TOR inhibition (21). It is, therefore, hypothesised that
delineating protein expression profiles of a wide array of
molecules participating in PI3K/AKT/mTOR signalling may aid in the
more accurate selection of NSCLC patients with a greater likelihood
to respond to the inhibition of this pathway. Another reason
accounting for the modest antitumour activities of rapamycin is the
abrogation of the feedback inhibition of EGFR signalling and the
promotion of AKT activation (22).
The incidence of PIK3CA mutations recorded in
the present series is in accordance with that recorded in the
literature, which ranges between 1.6 (11) and 5% (38). A total of 2 out of 3 mutations were
observed in the squamous cell carcinoma group and were mutually
exclusive with AKT1 mutations in agreement with the majority
of published studies (12).
However, in our subset, combined K-RAS, PIK3CA, AKT1 or
PTEN mutations were not observed.
In our cohort, using immunohistochemistry with
phosphorylation-specific antibodies, half (51%) of the NSCLC cases
were triple-positive for mTOR cascade components and essentially
all (49%) co-expressed cytoplasmic p-AKT, underlining that the
activation of this pathway is common in this tumour. Of note, the
percentage of cases expressing p-p70S6K (96%) far exceeded that of
p-mTOR expression (56.86%), fitting well with the proposed direct
phosphorylation of p70S6K independent of mTOR, by mitogen-activated
protein kinases (MAPKs) through P90RSK, PDK1 or directly by AKT
through TSC2 (39,40). The latter is verified by our finding
that all but one p-mTOR-negative/p-p70S6K-positive cases displayed
cytoplasmic p-AKT immunoreactivity. Significant positive
correlations were established between p-mTOR and its upstream
cytoplasmic p-AKT or its downstream p-p70S6K, consistent with
p-mTOR mediating the downstream effects of p-AKT on protein
synthesis and proliferation in NSCLC (41). The correlation between p-AKT and
p-mTOR in NSCLC has also been reported in other studies (16,21)
but has been questioned by Yoshizawa et al(18). Dobashiet al(22) observed little overlap in the
localization of the 2 molecules, except for adenocarcinomas
harbouring EGFR gene mutation, resulting in markedly
increased levels of p-AKT and p-mTOR. Analogous correlations
between p-mTOR and pS6 or eIF4E, the partner of 4E-BP1, have been
documented by Yoshizawa et al(18). Of note, we failed to elicit a
correlation between p-mTOR and p-4E-BP1, reinforcing the assumption
that an unknown pathway may be responsible for eIF4E/4e-BP1
phosphorylation, independent of p-mTOR (18). We also documented a positive
association between p85αPI3K or p110γPI3K with cytoplasmic p-AKT
which also applies to AKT downstream effectors. It should be noted
that the above correlations remained significant in the subgroups
of adenocarcinomas and squamous cell carcinomas, apart from the
ones between cytoplasmic p-AKT and p-mTOR in adenocarcinomas. In
their study on colorectal cancer, Johnson et al(42) revealed a strong correlation between
p85αPI3K and p-AKT (particularly p-AKT2) in agreement with our
findings. The positive correlation between p85αPI3K and p110γPI3K
nuclear H-scores attests to the functional liaison between the 2
subunits of PI3K. Zito et al(27) also observed the co-expression of the
2 subunits on their NSCLC specimens.
Surprisingly, we did not observe the negative
correlation between PTEN and p-AKT, neither by immunohistochemistry
nor by molecular analysis, as reported by Tang et
al(20) in NSCLC. Our data are
corroborated by Lim et al(30) in NSCLC, as well as by studies on
breast (43), renal cell (44) and urothelial bladder (45) carcinomas. In this context, it has
been hypothesised that in cancer, PTEN loss activates the Rho
family proteins, leading to increased cell migration and invasion
through pathways independent of AKT (43,46).
In the present study, PTEN expression also failed to correlate with
any other molecule of the PI3K/AKT/mTOR cascade consistent with the
fact that mTOR and its downstream proteins, p-4E-BP1 and p70S6K,
can also be activated by other pathways, not regulated by PTEN,
such as the Raf/MAPK signaling pathway (47). However, all the PTEN mutant cases
displayed cytoplasmic p-AKT immunoexpression, consistent with the
abrogation of PTEN inhibitory function on PI3K ability to
phosphorylate AKT (14).
As regards the expression of the phosphorylated
molecules and p85αPI3K in normal lung tissue, the alveolar
epithelium displayed no immunoreactivity, whereas the seromucous
glands and bronchial epithelium stained weakly for p-mTOR,
p85αPI3K, p-p70S6K and p-AKT (nuclear). These findings postulate
the essential role of the above proteins in the neoplastic
transformation of lung epithelial cells (21). PTEN was present in the normal
bronchi and alveolar epithelium, as well as in seromucous glands.
Notably, p-4E-BP1 was not observed in normal lung tissue.
An interesting finding is that p-mTOR expression was
considerably higher in adenocarcinomas, lending support to the
results of Liu et al(21)
and Dobashi et al(22) and
disputing those of Yoshizawa et al(18), reporting higher p-mTOR levels in
squamous cell carcinoma. It should be noted though, that the
preferential enhancement of mTOR activation was not observed in
adenocarcinoma cultured cells, suggesting the potential implication
of mTOR in the morphogenesis of adenocarcinoma (22). The same applied to p110γ nuclear
expression, in accordance with the recently reported prevalence of
PI3K subunit expression in adenocarcinoma (27). From a clinical perspective, these
findings raise the issue of the selective response of patients with
adenocarcinoma to mTOR or PI3K inhibitors. On the contrary,
p-4E-BP1 expression levels prevailed in squamous cell carcinoma.
Previous reports, however, assign eIF4E expression rather to lung
adenocarcinomas, where it is thought to facilitate invasiveness
(48,49).
Among the molecules investigated, p-4E-BP1
immunoexpression levels increased in parallel with tumour status.
The same applied to nuclear p-AKT expression as regards stage,
whereas p-p70S6K (marginally) prevailed in low-grade/low T-category
cases. A similar inverse correlation of marginal significance was
observed between p-mTOR and histological grade. These findings
concur with those of Liu et al(21) and advocate that the activation of
mTOR and p70S6K appears more critical during the early stages of
lung carcinogenesis as opposed to 4E-BP1 and AKT, which is a rather
late event, contributing to the acquisition of a more aggressive
phenotype. In harmony with our results, eIF4E overexpression was
reportedly more prevalent in advanced stages of lung
adenocarcinomas (25), whereas
published data regarding the association of p-AKT and stage are
controversial (20,26). PTEN loss also marginally prevailed
in high-grade cases, as in the study of Tang et al(20) and more importantly, the presence of
the PTEN mutant genotype was associated with nodal
metastases. A similar association, albeit of marginal significance,
was obtained between the p110γ expression level and nodal status.
The overall impression gained from these observations is the
diverse role of PI3K/mTOR signalling during the evolution of the
neoplastic process in NSCLC.
One of the most significant findings emerging from
the present study is the adverse prognostic significance of
p-4E-BP1 expression in adenocarcinomas established in univariate
analysis and remaining in multivariate analysis as a function of
its interaction with p-AKT expression and histological type. p-AKT
cytoplasmic expression on its own attained a marginal significance
in univariate analysis in this regard. The survival time was also
substantially shorter for patients with a co-expression of p-AKT
and p-4E-BP1 than for those with either the single-positive or
double-negative phenotype, implying that multi-targeted
interventions may be more effective than the use of single
inhibitors (21). For example,
rapamycin has been shown to induce the phosphorylation of AKT and
eIF4E, which was suppressed by a PI3K inhibitor (50). To the best of our knowledge,
although there are a few studies investigating the prognostic
significance of the PI3K/p-AKT/mTOR pathway in NSCLC (18–23,26–32),
the prognostic utility of p-4E-BP1 expression in this regard has
not been documented thus far.
The oncogenic effects of 4E-BP1 phosphorylation
cannot be disentangled from those of eIF4E which coordinates
translation initiation with the consequent preferential enhancement
of the synthesis of growth-promoting or oncogenic proteins
facilitating angiogenesis, invasion and metastasis. The strongest
evidence supporting the role of the 4E-BP1/p-eIF4E interaction in
NSCLC patient survival comes from 3 independent studies assigning a
poor survival probability to p-eIF4E overexpression (18,24,25) in
lung adenocarcinomas. Current belief holds that the activation of
4E-BP1 represents the convergence point of several oncogenic
pathways apart form mTOR, hence providing a better reflection of
tumour aggressiveness than the upstream genetic alterations
(51).
The prognostic role of the remaining components of
the PI3K/AKT/mTOR pathway remains a controversial issue, with the
majority of studies proposing p-AKT and/or (p-) mTOR as adverse
(18,20,21,28–32)
and others as favourable (19,26)
prognosticators. The overexpression of p-p70S6K (or pS6) (21,23),
p85 or p110 subunits of PI3K (27),
as well as PTEN loss (20)
reportedly confer shorter survival. These inconsistencies may
reflect the type of antibodies used (against phosphorylated or
total molecules), quantification methods, variability in
stage/histology and the use of various cut-off points for
categorization. It also becomes clear that not all components of
the pathway have a similar impact on prognosis. Another issue that
remains to be addressed is our inability to substantiate the
prognostic value of stage in our series, which may be related to
the fact that stages IIIB and IV were not represented.
In conclusion, in the present study, we provide
evidence that alterations of the PI3K/AKT/mTOR pathway components
are differentially implicated in the pathogenesis and
aggressiveness of NSCLC. Our data stands in favour of nuclear
p-4E-BP1 immunoexpression as a molecular marker of prognostic value
in adenocarcinomas, particularly when combined with p-AKT. Careful
evaluation of these parameters may help predict which tumours are
most sensitive to PI3K/AKT signalling inhibitors (51).
References
|
1
|
Travis WD; IASLC Staging Committee.
Reporting lung cancer pathology specimens. Impact of the
anticipated 7th edition TNM classification based on recommendations
of the IASLC Staging Committee. Histopathology. 54:3–11. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Siegel R, Ward E, et al: Cancer
statistics: 2007. CA Cancer J Clin. 57:43–66. 2007. View Article : Google Scholar
|
|
3
|
Vivanco I and Sawyers CL: The
phosphatidyloinositol 3-kinase AKT pathway in human cancer. Nat Rev
Cancer. 2:489–501. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sarbassov DD, Ali SM and Sabatini DM:
Growing roles for the mTOR pathway. Curr Opin Cell Biol.
17:596–603. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sarbassov DD, Ali SM, Kim DH, et al:
Rictor, a novel binding pattern of mTOR, defines a
rapamycin-insensitive and raptor-independent pathway that regulates
the cytoskeleton. Curr Biol. 14:1296–1302. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hay N and Sonenberg N: Upstream and
downstream of mTOR. Genes Dev. 18:1926–1945. 2004. View Article : Google Scholar
|
|
7
|
Gingras AC, Gypi SP, Raught B, et al:
Regulation of 4E-BP1 phosphorylation: a novel two-step mechanism.
Genes Dev. 13:1422–1437. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bjornsti MA and Houghton PJ: The TOR
pathway: a target for cancer therapy. Nat Rev Cancer. 4:335–348.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu J, Zhang Y, McIlroy J, et al:
Regulation of the p85/p110 phosphstidyloinositol 3′-kinase:
stabilization and inhibition of the p110alpha catalytic subunit by
the p85 regulatory subunit. Mol Cell Biol. 18:1379–1387. 1998.
|
|
10
|
Miled N, Yan Y, Hon WC, et al: Mechanism
of two classes of cancer mutations in the phosphoinositide 3-kinase
catalytic subunit. Science. 317:239–242. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yamamoto H, Hisayuki S, Masaharu N, et al:
PI3KCA mutations and copy number gains in human lung cancer. Cancer
Res. 68:693–621. 2008. View Article : Google Scholar
|
|
12
|
Scrima M, De Marco C, Fabiani F, et al:
Signaling networks associated with AKT activation in non-small cell
lung cancer (NSCLC): new insights on the role of
phosphatidyl-inositol-3 kinase. PLoS One. 7:e304272012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Marti A and Felip E: PI3K pathway in
NSCLC. Front Oncol. 1:552012.
|
|
14
|
Stambolic V, Suzuki A, de la Pompa JL, et
al: Negative regulation of PKB/Akt-dependent cell survival by the
tumor suppressor PTEN. Cell. 95:29–39. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wangpaichitr M, Wu C, You M, et al:
Inhibition of mTOR restores cisplatin sensitivity through
down-regulation of growth and anti-apoptotic proteins. Eur J
Pharmacol. 591:124–127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Balsara BR, Pei J, Mitsuuchi Y, et al:
Frequent activation of AKT in non-small cell lung carcinomas and
preneoplastic bronchial lesions. Carcinogenesis. 25:2053–2059.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Okudela K, Suzuki M, Kageyama S, et al:
PIK3CA mutation and amplification in human lung cancer. Pathol Int.
57:664–671. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yoshizawa A, Fukuoka J, Shimizu S, et al:
Overexpression of phospho-eIF4E is associated with survival through
AKT pathway in non-small cell lung cancer. Clin Cancer Res.
16:240–248. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Anagnostou VK, Bepler G, Syrigos KN, et
al: High expression of mammalian target of rapamycin is associated
with better outcome for patients with early stage lung
adenocarcinoma. Clin Cancer Res. 15:4157–4164. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang JM, He QY, Guo RX, et al:
Phosphorylated AKT overexpression and loss of PTEN expression in
non-small cell lung cancer confers poor prognosis. Lung Cancer.
51:181–191. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu D, Huang Y, Chen B, et al: Activation
of mammalian target of rapamycin pathway confers adverse outcome in
nonsmall cell lung carcinoma. Cancer. 117:3763–3673. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dobashi Y, Suzuki S, Matsubara H, et al:
Critical and diverse involvement of Akt/mammalian target of
rapamycin signaling in human lung carcinomas. Cancer. 115:107–118.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
McDonald JM, Pelloski CE, Ledoux A, et al:
Elevated phospho-S6 expression is associated with metastasis in
adenocarcinoma of the lung. Clin Cancer Res. 14:7832–7837. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Khoury T, Alrawi S, Ramnath N, et al:
Eukaryotic initiation factor-4E and cyclin D1 expression associated
with patient survival in lung cancer. Clin Lung Cancer. 10:58–66.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang R, Genng J, Wang JH, et al:
Overexpression of eukaryotic initiation factor 4E (eIF4E) ad its
clinical significance in lung adenocarcinoma. Lung Cancer.
66:237–244. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oh MH, Lee HJ, Yoo SB, et al:
Clinicopathological correlations of mTOR and pAkt expression in
non-small cell lung cancer. Virchows Arch. 460:601–609. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zito CR, Jilaveanu LB, Anagnostou V, et
al: Multi-level targeting of the phosphatidyloinositol-3-kinase
pathway in non-small cell lung cancer cells. PLoS One.
7:e313312012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Gately K, Al-Alao B, Dhillon T, et al:
Overexpression of the mammalian target of rapamycin (mTOR) and
angioinvasion are poor prognostic factors in early stage NSCLC: a
verification study. Lung Cancer. 75:217–222. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dhillon T, Mauri FA, Belezza G, et al:
Overexpression of the mammalian target of rapamycin: a novel
biomarker for poor survival in resected early stage non-small cell
lung cancer. J Thorac Oncol. 5:314–319. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lim WT, Zhang WH, Miller CR, et al: PTEN
and phosphorylated AKT expression and prognosis in early- and
late-stage non-small cell lung cancer. Oncol Rep. 17:853–857.
2007.PubMed/NCBI
|
|
31
|
Shi Y, Chen L, Li J, et al: Prognostic and
predictive values of pERK1/2 and pAkt-1 expression in non-small
cell lung cancer patients treated with adjuvant chemotherapy.
Tumour Biol. 32:381–389. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
David O, Jett J, LeBeau H, et al:
Phospho-Akt overexpression in non-small cell lung cancer confers
significant stage-independent survival disadvantage. Clin Cancer
Res. 10:6865–6871. 2004. View Article : Google Scholar
|
|
33
|
Korkolopoulou P, Levidou G, Trigka EA, et
al: A comprehensive immunohistochemical and molecular approach to
the PI3K/AKT/mTOR (phosphoinositide 3-kinase/v-akt murine thymoma
viral oncogene/mammalian target of rapamycin) pathway in bladder
urothelial carcinoma. BJU Int. E1237–E1248. 2012. View Article : Google Scholar
|
|
34
|
Levidou G, Saetta AA, Gigelou F, et al:
ERK/pERK expression and B-raf mutations in colon adenocarcinomas:
correlation with clinicopathological characteristics. World J Surg
Oncol. 10:472012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rojo F, Najera L, Lirola J, et al:
4E-binding protein 1, a cell signaling hallmark in breast cancer
that correlates with pathologic grade and prognosis. Clin Cancer
Res. 13:81–89. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fujisaka Y, Yamada Y, Yamamoto N, et al: A
phase 1 clinical study of temsirolimus (CCI-779) in Japanese
patients with advanced solid tumors. Jpn J Clin Oncol. 40:732–738.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pal SK, Figlin RA and Reckamp KL: The role
of targeting mammalian target of rapamycin in lung cancer. Clin
Lung Cancer. 9:340–345. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
An SJ, Chen ZH, Su J, et al:
Identification of enriched driver gene alterations in subgroups of
non-small cell lung cancer patients based on histology and smoking
status. PLoS One. 7:e401092012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thomas G: The S6 kinase signaling pathway
in the control of development and growth. Biol Res. 35:305–313.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Riemenschneider MJ, Betensky RA, Pasedag
SM, et al: AKT activation in human glioblastomas enhances
proliferation via TSC1 and S6 kinase signaling. Cancer Res.
66:5618–5623. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Han S, Khuri FR and Roman J: Fibronectin
stimulates non-small cell lung cancinoma cell growth through
activation of Akt/mammalian target of rapamycin/S6 kinase and
inactivation of LKB1/AMP-activated protein kinase signal pathways.
Cancer Res. 66:315–323. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Johnson SM, Gulhati P, Rampy BA, et al:
Novel expression patterns of PI3K/Akt/mTOR signaling pathway
components in colorectal cancer. J Am Coll Surg. 210:767–778. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Bose S, Chandran S, Mirocha JM and Bose N:
The Akt pathway in human breast cancer: a tissue-array-based
analysis. Mod Pathol. 19:238–245. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Pantuck AJ, Seligson DB, Klatte T, et al:
Prognostic relevance of the mTOR pathway in renal cell carcinoma:
implications for molecular patient selection for targeted therapy.
Cancer. 109:2257–2267. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sun CH, Chang YH and Pan CC: Activation of
the PI3K/Akt/mTOR pathway correlates with tumour progression and
reduced survival in patients with urothelial carcinoma of the
urinary bladder. Histopathology. 58:1054–1063. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liliental J, Moon SY, Lesche R, et al:
Genetic deletion of the Pten tumor suppressor gene promotes cell
motility by activation of Rac1 and Cdc42 GTPases. Curr Biol.
10:401–404. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dai B, Kong YY, Ye DW, et al: Activation
of the mammalian target of rapamycin signaling pathway in prostate
cancer and its association with patient clinicopathological
characteristics. BJU Int. 104:1009–1016. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Seki N, Takasu T, Mandai K, et al:
Expression of eukaryotic initiation factor 4E in atypical
adenomatous hyperplasia and adenocarcinoma of the human peripheral
lung. Clic Cancer Res. 8:3046–3053. 2002.PubMed/NCBI
|
|
49
|
Rosenwald IB, Hutzler MJ, Wang S, et al:
Expression of eukaryotic translation initiation factors 4E and
2alpha is increased frequently in bronchioalveolar but not in
squamous cell carcinomas of the lung. Cancer. 92:2164–2171. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Sun SY, Rosenberg LM, Wang X, et al:
Activation of Akt and eIF4E survival pathways by rapamycin-mediated
mammalian target of rapamycin inhibition. Cancer Res. 65:7052–7058.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Armengol G, Rojo F, Castellvi J, et al:
4E-binding protein 1: a key molecular ‘funnel factor’ in human
cancer with clinical implications. Cancer Res. 67:7551–7555.
2007.
|
|
52
|
Dumstorf CA, Konicek BW, McNulty AM, et
al: Modulation of 4E-BP1 function as a critical determinant of
enzastaurin-induced apoptosis. Mol Cancer Ther. 9:3158–3163. 2010.
View Article : Google Scholar : PubMed/NCBI
|