Introduction
Colorectal cancer (CRC) is the third most common
cancer in men and the second in women worldwide (1) and has a high propensity for liver
metastasis (2). The primary cause
of mortality in patients with CRC is liver metastasis (3), and the 5-year overall survival (OS) is
only 25–40% (3,4). The molecular mechanisms underlying CRC
metastasis are not completely understood. Early treatment targeting
CRC liver metastatic foci may be important for improving patient
survival. Therefore, there is an urgent need to identify molecules
that facilitate the metastasis of CRC to the liver, which would be
potential therapeutic targets for treating patients with CRC and
liver metastases.
The GATA factors belong to an evolutionarily
conserved family of C2-type zinc finger proteins. There are six
members of the vertebrate GATA family. GATA1, 2 and 3 are mainly
expressed in the hematopoietic lineages and GATA4, 5 and 6 are
expressed in endodermally derived tissues, such as the gut, liver
and lungs (5). In the adult small
intestine, GATA4, 5 and 6 are expressed in a partially overlapping
pattern along the crypt/villus axis (6,7).
GATA4, 5 and 6 regulate various differentiation marker genes
expressed in gastrointestinal tissue by binding to the WGATAR
sequences within the regulatory regions of these genes and
interacting with other ubiquitous and tissue-enriched
transcriptional regulators (8–10). In
addition to differentiation, GATA4, 5 and 6 have been associated
with cell survival, cell proliferation, and neoplastic
transformation of various cell types (11–15).
Among gastrointestinal cancers, GATA4 is amplified in up to 10% of
esophageal adenocarcinomas and Barrett metaplasia (16). The transition from normal esophageal
epithelium to Barrett metaplasia to adenocarcinoma is associated
with upregulation of GATA6 (17).
Furthermore, the GATA6 gene is amplified in pancreaticobiliary
cancer (18). A strong expression
of GATA6 in the proliferative crypt compartment of the intestine
has suggested that GATA6 may be associated with cellular
proliferation. In support of this, GATA6 is strongly expressed in
CRC-derived cell lines and is upregulated in CRC (19). However, the roles of GATA6 in the
prognosis and the metastasis of CRC to the liver have yet to be
explored.
In the present study, we first investigated the
expression of GATA6 in a series of CRC patients and its
correlations with patient prognosis and liver metastasis. Then, the
role of GATA6 in CRC invasion and metastasis was studied in CRC
cells.
Materials and methods
CRC patient samples
A total of 89 CRC patients undergoing surgical
resection between January 1, 2004 and October 1, 2010, in Guangzhou
First People’s Hospital, Guangzhou Medical College, were included.
One hundred and six cancerous samples, 35 adjacent normal, 39 liver
metastatic samples from 89 CRC patients were collected. The median
age of the patients was 54 years at operation, range 28–81 years.
Follow-up and clinicopathological characteristics including tumor
location, stage, and differentiation were recorded. OS was
calculated from the date of surgery until the date of last contact.
This study was approved by the Ethics Committee of the Guangzhou
First People’s Hospital, Guangzhou Medical College.
Immunohistochemical analysis (IHC)
Surgical samples containing primary tumors, adjacent
normal tissues and liver metastatic tumors were collected in 10%
buffered formalin. The details of deparaffinization and IHC were
previously described (20).
Briefly, following deparaffinization, the endogenous peroxidase
activity was blocked with 3% H2O2. The array
slides were incubated with normal goat serum for 20 min, and then
applied with primary antibody for 20 min at room temperature. After
7 min of H2O2 treatment, the array slides
were incubated with horseradish peroxidase-labeled polymer
conjugated with corresponding antibodies for 30 min. Each slide was
counterstained with hematoxylin (Dako, Carpinteria, CA, USA). PBS
was used as a negative control.
The rabbit anti-human GATA6 monoclonal antibody
(Sigma-Aldrich, USA) was used for IHC staining (1:100 dilution). To
reduce the image reader bias, an automated imaging system was
employed to obtain digital images of the stained sections for
subsequent quantitative analyses. In addition, each sample was
evaluated by two independent investigators in a double-blind
manner. Investigators reviewed and assessed the subcellular
localization (cytoplasm vs. nucleus), staining intensity
(integrated optical density), and/or percentage of stained cells
(total area or percentage of cells positive) for each image.
Discrepancies in samples were resolved after joint review by the
readers.
CRC cell culture and transfection
The two CRC cell lines (Colo-205 and SW-480 cells)
were obtained from the American Type Culture Collection and
maintained in RPMI-1640 with 10% calf serum (HyClone) at 37°C with
5% CO2.
Cells were transfected with lentiviral vectors
encoding short hairpin RNA targeting human GATA6 for GATA6
knockdown (shGATA6) or a scrambled shRNA as control (shControl)
(Sigma-Aldrich). Multiplicity of infection was 10. Cells were
cultured for 72 h after transfection. Cells were grown to 80%
confluency in 60-mm dishes. GATA6 (10 lg) plasmid (ExGATA6) or
empty plasmid (ExControl) was transfected using Lipofectamine
Reagent (Invitrogen). Cells were cultured for 48 h after
transfection.
Cell invasion assay
Cells (2×105) were suspended in 400 μl
serum-free RPMI-1640 medium and seeded in the top chamber that had
been coated with a layer of extracellular matrix (BD Biosciences,
USA). The complete medium with serum (500 μl) was added to the
bottom chamber. After 48 h of incubation, the cells which had
invaded through the extracellular matrix layer to the lower surface
of the filters were stained. Images of three randomly selected
fields of the fixed cells were captured, and cells were counted.
Experiments were repeated independently three times.
Cell migration assay
Cells were seeded in a 6-well plate, grown until
confluence and then starved for 24 h. A linear wound was made by
scraping a pipette tip. The cell motility in terms of wound closure
was measured by photographing at three random fields 72 h after
wounding. Experiments were repeated independently three times.
Real-time PCR
Total RNA was extracted from CRC cells using RNA
Extraction kit (Qiagen, China). The cDNA synthesis was performed
according to the manufacturer’s instructions (SYBR-Green PCR kit;
Qiagen). Quantitative PCR was performed by SYBR-Green PCR kit
(Qiagen) using a LightCycler system. PCR reaction conditions for
all assays were 94°C for 30 sec, followed by 40 cycles of
amplification (94°C for 5 sec, 58°C for 30 sec and 72°C for 30
sec). β-actin mRNA was used to normalize RNA. Primer sequences
were: GATA6 forward, CCAACTTCCACCTCTTCTAAC and reverse,
TTGACCCGAATACTTGAGC; β-actin forward, CCATGTACGTTGCTATCCAGG and
reverse, TCTCCTT AATGTCACGCACGA.
Western blot analysis
Nuclear protein or total proteins of CRC cells were
isolated using RIPA (Cell Signaling Technology). For
immunoblotting, equal amounts of proteins were separated on 5–8%
SDS-PAGE and were electrophoretically transferred onto
nitrocellulose membranes (Millipore), which were blocked in TBST
containing 5% milk for 2 h at room temperature and blotted with
antibody overnight at 4°C: anti-GATA6 (1:1,000; Abcam) and
anti-histone H3 (1:500; Cell Signaling Technology). After washing
with TBST and incubating with either anti-rabbit or anti-mouse
horseradish peroxidase-conjugated secondary antibody (Cell
Signaling Technology) for 2 h at room temperature, immunocomplexes
were visualized using the chemiluminescence (GE Healthcare Life
Sciences, USA) following the manufacturer’s protocol.
Statistical analysis
Data were analyzed using SPSS 17.0. Continuous data
were measured by the t-test. For categorical data, Chi-square
analysis or Fisher’s exact test was used. Multivariate logistic
regression was used to adjust for covariate effects on the odds
ratio (OR). Kaplan-Meier was applied for OS. P<0.05 was
considered to indicate a statistically significant difference.
Results
GATA6 expression in primary CRC,
metastatic liver lesions and normal tissues
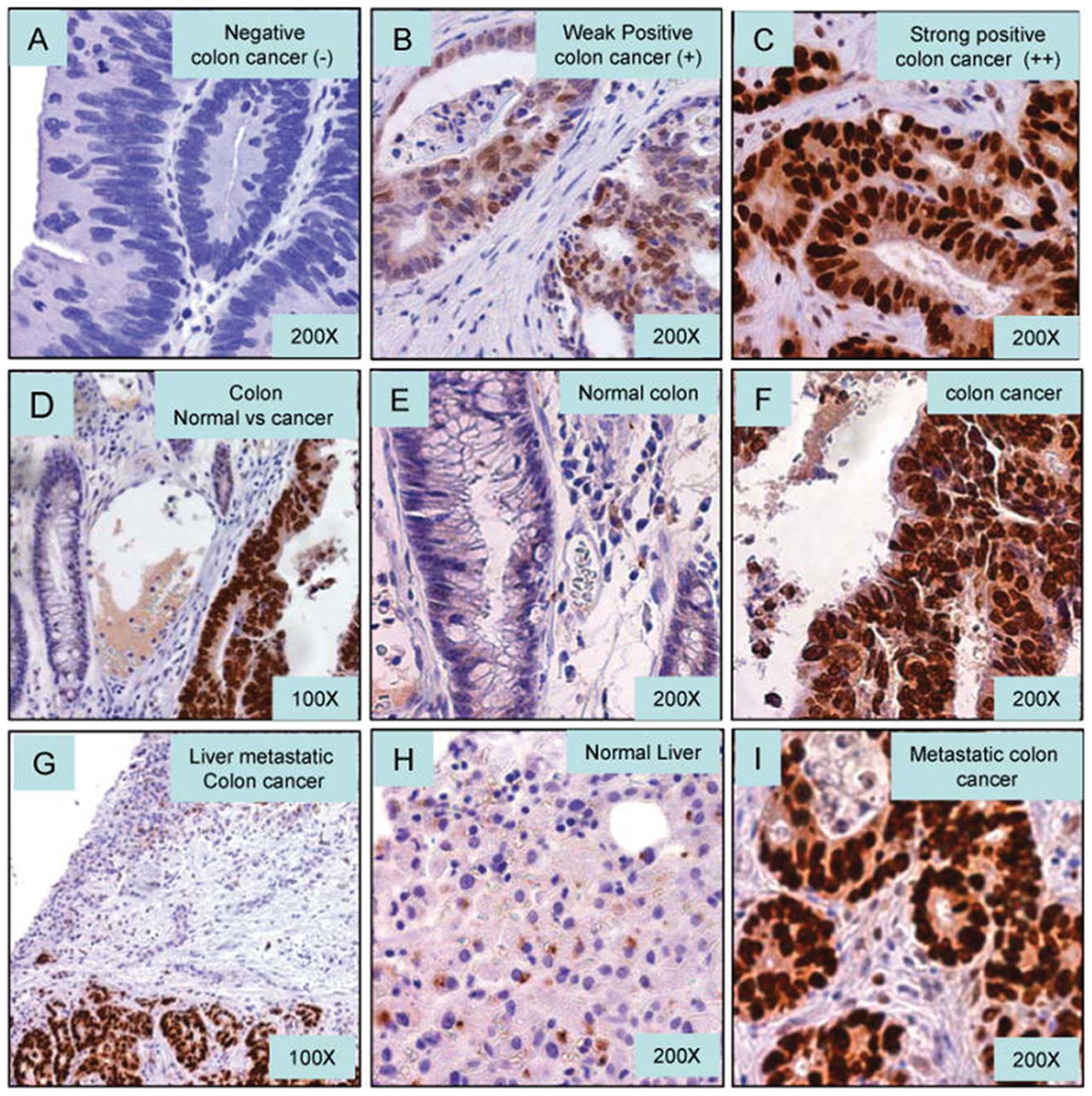
We examined GATA6 protein expression in metastatic
CRC by IHC. In general, GATA6 was predominantly nuclear staining in
IHC. Some perinuclear granulation in cytoplasm was also observed.
GATA6 expression was quantified using a visual grading system based
on the extent of staining. Only immunoreactivity in the nucleus was
evaluated. GATA6 was defined as: negative nuclear staining (−),
<5%; positive (+), ≥5% and <50% or strong positive (++),
>50% positive nuclear staining from CRC cells. The GATA6 IHC
standards of negative (−), weak positive (+) and strong positive
(++) are displayed in Fig. 1A–C,
respectively. GATA6 expression was predominantly observed in the
primary cancer cells, but not in the adjacent normal colorectal
epithelium (Fig. 1D–F). Meanwhile,
strong positive GATA6 was also observed in metastatic CRC to liver
(Fig. 1G–I).
Expression of GATA6 is associated with
hepatic metastasis of CRC
Based on the IHC staining of GATA6, 32 of 89 CRC
samples were defined as GATA6 nuclear positive staining [including
weak positive (+) and strong positive (++)]. The TNM stage of CRC
was based on clinical diagnosis. According to the univariate
analysis results, the GATA6 staining was positively and
significantly associated with hepatic metastasis (P<0.01) and
tumor invasion (P<0.05) of CRC, but not with age, gender, tumor
location and lymph node involvement (Table I).
 | Table IClinicopathological characteristics
and GATA6 expression of CRC. |
Table I
Clinicopathological characteristics
and GATA6 expression of CRC.
| Characteristics | No. of cases | No. of positive GATA6
(%) | P-value |
|---|
| Age |
| <40 | 4 | 0 (0.0) | |
| 40–49 | 5 | 2 (40.0) | |
| 50–59 | 20 | 7 (35.0) | |
| 60–69 | 30 | 13 (43.3) | |
| 70–79 | 23 | 7 (30.4) | |
| >80 | 7 | 3 (42.9) | 0.567 |
| Gender |
| Male | 42 | 18 (42.9) | |
| Female | 47 | 14 (29.8) | 0.2 |
| Location |
| Rectum | 7 | 5 (71.4) | |
| Colon | 82 | 27 (32.9) | |
| Proximal | 53 | 17 (32.1) | |
| Distal | 36 | 15 (41.7) | 0.158 |
| Tumor invasion |
| Within propria | 20 | 8 (40) | |
| Out propria | 69 | 24 (34.8) | 0.027a |
| Lymph node |
| Negative | 38 | 15 (39.5) | |
| Positive | 51 | 17 (33.3) | 0.55 |
| Hepatic
metastasis |
| No | 50 | 13 (26.0) | |
| Yes | 39 | 19 (48.7) | 0.001a |
To validate this finding, non-conditional logistic
analysis was employed for univariate and multivariate analyses. The
GATA6 positive and negative were stratified as unfavorable and
favorable subsets, respectively. Tumor invasion, lymph node
involvement, and hepatic metastasis were considered as the endpoint
in logistic analysis. The expression of GATA6 significantly
impacted the risk of hepatic metastasis but not tumor invasion and
lymph node involvement according to either univariate or
multivariate analysis. After adjusting for age and gender, the OR
of GATA6-positive for hepatic metastasis was 3.53 (95% CI,
1.37–9.70) (Table II). Therefore,
our analyses revealed that GATA6 significantly affected hepatic
metastasis of CRC, suggesting that GATA6 may be used to
prognosticate CRC.
| Table IIUnivariate and multivariate logistic
analysis for GATA6 and TNM stage of CRC. |
Table II
Univariate and multivariate logistic
analysis for GATA6 and TNM stage of CRC.
| n=89 |
|---|
|
|
|---|
| OR (95% CI) | Adjusted ORa (95% CI) |
|---|
| Tumor invasion |
| T0–2 | Reference | Reference |
| T3–4 | 0.91
(0.32–2.74) | 0.74
(0.24–2.35) |
| Lymph node
involvementb |
| N0 | Reference | Reference |
| N1 or N2 | 0.77
(0.32–1.84) | 0.48
(0.17–1.31) |
| Hepatic
metastasisc |
| M0 | Reference | Reference |
| M1 | 2.70
(1.12–6.72)d | 3.83
(1.14–14.14)d |
Positive GATA6 expression correlates with
poor prognosis in CRC
The prognostic significance of GATA6 was determined
by GATA6 staining and the corresponding clinical follow-up records.
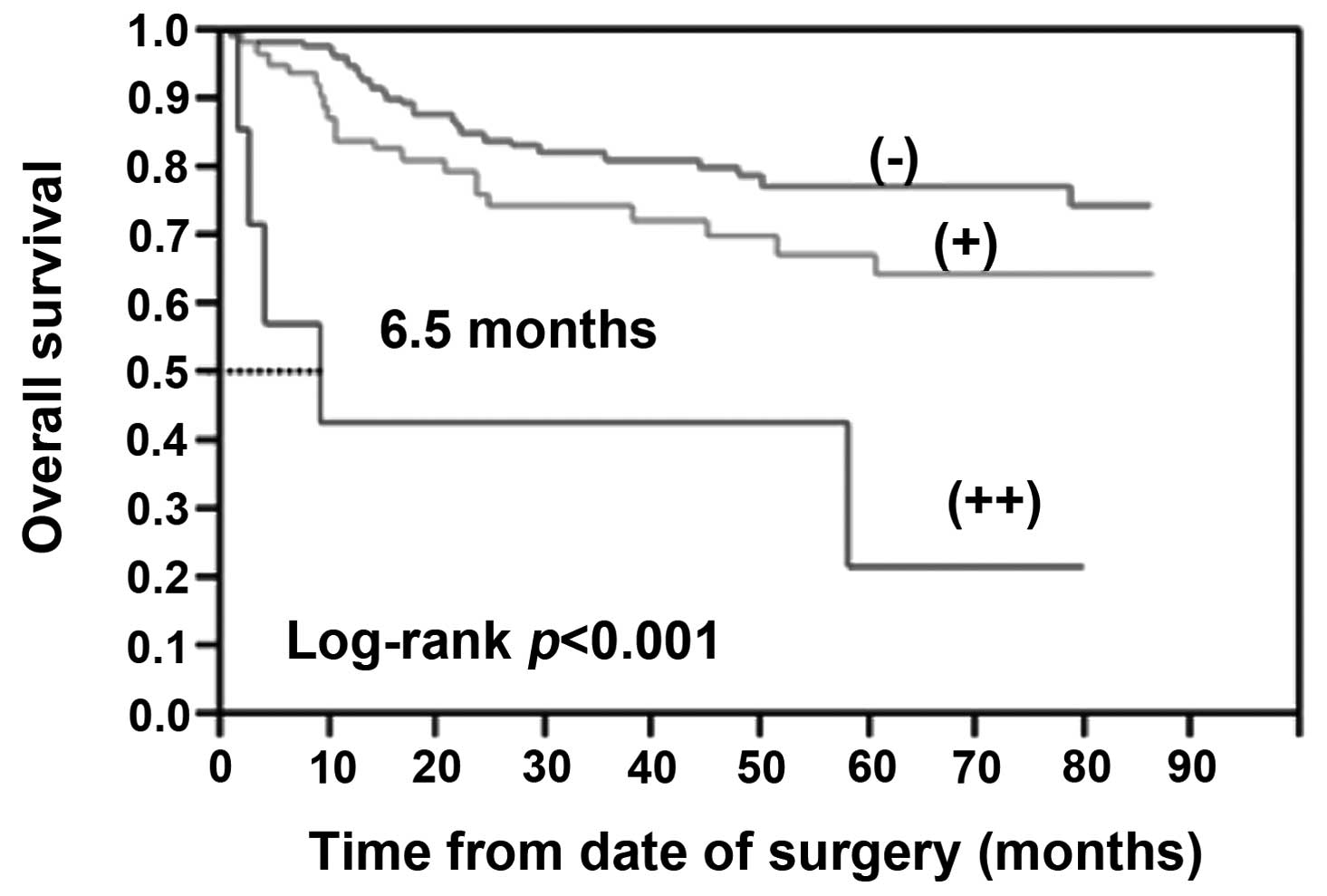
Kaplan-Meier survival analysis revealed a correlation between
higher GATA6 expression levels and shortened OS times (Fig. 2). The Log-rank test indicated that
the positive GATA6 expression was significantly related to OS
(P<0.05). The strong positive GATA6 (++) exhibited a more
significantly reduced survivability (P<0.001). Taken together,
these observations indicate that overexpression of GATA6 is
significantly associated with CRC liver metastasis and poor
prognosis in patients with CRC.
GATA6 promotes CRC cell migration,
invasion and metastasis
As clinical data showed positive correlation between
GATA6 and metastatic behavior in CRC, the roles of GATA6 in
invasion and metastasis were investigated in two human CRC cell
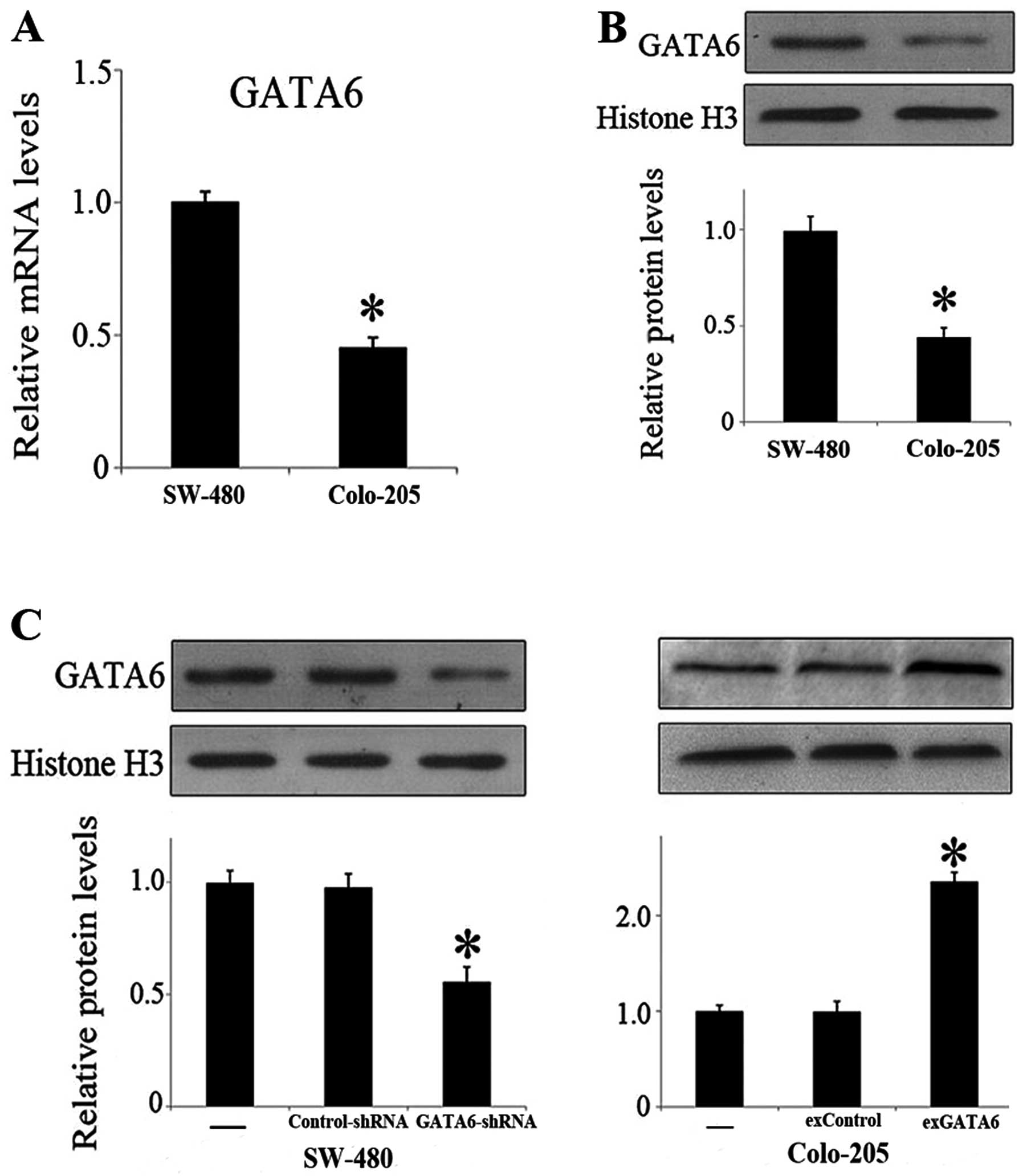
lines, Colo-205 and SW-480 cells. Both mRNA and nuclear protein
levels of GATA6 were significantly higher in SW-480 cells than in
Colo-205 cells (Fig. 3A and B), and
therefore, knockdown of GATA6 was performed in SW-480 cells by
GATA6-shRNA lentivirus transfection and overexpression of GATA6 was
performed in Colo-205 cells by GATA6 plasmid transfection,
respectively. The GATA6 nuclear protein levels after transfection
were also validated (Fig. 3C). By
transwell assay, the number of invading cells was markedly reduced
by knockdown of GATA6 in SW-480 cells and was markedly increased by
overexpression of GATA6 in Colo-205 cells (Fig. 4A). Consistently, cell migration was
significantly reduced by GATA6 knockdown and enhanced by GATA6
overexpression by wound-healing assay (Fig. 4B). Collectively, the data suggested
a role of GATA6 in promoting CRC cell invasion and metastasis.
Discussion
In the present study, we examined the expression
level of GATA6 in CRC to assess its potential role as a prognostic
or predictive marker. Our findings revealed that the positive
expression of GATA6 was significantly associated with the hepatic
metastasis. The adjusted ORs of GATA6 for the risk of hepatic
metastasis were 3.53 (95% CI, 1.37–9.70). We also found that the
positive expression of GATA6 was an indicator of poor prognosis.
Kaplan-Meier survival analysis revealed a correlation between
higher GATA6 expression levels and shorter OS. The log-rank test
indicated that the positive GATA6 expression was significantly
related to OS. In vitro, the cell invasion and migration of
established CRC cell lines were decreased by GATA6 knockdown and
enhanced by GATA6 overexpression. Therefore, our findings suggest
that the positive expression of GATA6 is associated with the
hepatic metastasis of CRC and a reduced patient survival.
GATA6 functions as a promoter or suppressor
according to the tumor origin. GATA6 activates the expression of
tumor suppressor, Dab2, and regulates the activity of LKB tumor
suppressor protein (21,22). Loss of GATA6 protein function due to
epigenetic silencing of GATA6 gene or GATA6 protein exclusion from
the nuclei has been reported in ovarian cancer (12,23).
Although GATA6 was expressed in normal adrenal cortex, GATA6
expression was downregulated in adrenocortical tumors (15). Furthermore, GATA6 was downregulated
in hyperplastic neointimal smooth muscle cells and forced
expression of GATA6 restored normoplasia (24). A recent study revealed tumor
suppressor activity of GATA6 in astrocytomas (25). In contrast to the tumor-suppressing
activity of GATA6 in these tissues, the promoter function of GATA6
is reported mainly in digestive malignancies, such as tumorigenesis
in the esophagus, pancreas and intestines (13,17,18,26)
indicating that GATA6 may be a digestive-lineage tumor promoter,
which, however, requires further elucidation. The studies above are
in agreement with our findings, and the present study in
vitro showed GATA6 induces migratory and invasive behavior,
promoting invasion and metastasis, and confers survival
advantages.
However, the mechanism by which overexpressed GATA6
participates in the initiation and/or progression of CRC is not
known. GATA6 binding sites are present in the regulatory regions of
members of the Wnt family of secreted glycoproteins such as Wnt 2,
4, 6, 7b and 8b, and GATA6 regulates the expression of Wnt 2, 7b
and 8 and the Wnt receptor Fzd2 (27–30).
Dysregulated expression of GATA6 in preneoplastic colorectal
lesions may lead to overexpression of target Wnts, which may
trigger the Wnt signaling pathway implicated in the pathogenesis of
colorectal cancer (31). Continued
overexpression of GATA6 in benign and malignant lesions may aid in
maintaining the activated Wnt-β-catenin signaling during
progression of CRC. In addition to Wnts, GATA6 also regulates the
expression of the members of the transforming growth factor-β
(TGF-β) family of proteins such as BMP4. The BMP4 promoter contains
consensus GATA binding sites, and these sites are essential for
GATA6-induced activation of the BMP4 promoter (32). BMP4 is overexpressed in malignant
and metastatic CRC compared with benign lesions and normal mucosa
(33). A recent genome-wide study
showed the association between BMP4 and CRC (34). BMP4 activates the Smad signaling
pathway and induces epithelial-mesenchymal transition and uPA
production in CRC cells and promotes cell migration and invasion.
In support of this, a previous study showed that forced expression
of BMP4 in HCT116 CRC cells induced uPA gene expression and
enhanced cell migration and invasion (33). GATA6 and the related protein, GATA4,
physically and functionally interact with Smads and play an
essential role in TGF-β signal transduction (8). Thus, it is conceivable that the
overexpression of GATA6 will serve dual functions: it will lead to
excessive production of BMP4 and it will participate in the
activation of BMP4-induced Smad pathway signaling, leading to cell
migration and invasion.
Although results from previous studies suggested
that GATA6 functions as an oncogene that is overexpressed in
various types of human cancer, it remains unclear how GATA6
promotes cancer invasion and metastasis in these types of cancer.
GATA6 may affect CRC cell migratory and invasive properties
independently. Multiple extracellular matrix degrading proteases
such as matrix metalloproteinases 2 and 9 are regulated by GATA2 in
endothelial cells (35). Since
GATA6 is expressed in CRC, the expression of these matrix
metalloproteinases in colorectal tumors may be regulated by GATA6.
GATA6 may also be involved in other pathways independent of
proteases to influence the cell migratory properties. GATA6 and the
related GATA4 proteins are targeted by the small GTPase RhoA
pathway, an evolutionarily conserved pathway that regulates the
migratory behavior of normal and cancerous cells (36).
In conclusion, our finding that GATA6 overexpression
is an informative biomarker, which is associated with poor
prognosis of patients with CRC, has potential implications for CRC
survival prediction, choice of treatment regimens and future
development of treatment strategies.
Acknowledgements
The study was supported by the Guangdong Provincial
Science & Technology Projects (2009B060700019 and
2013B031800103).
References
|
1
|
Ferlay J, Shin HR, Bray F, et al:
Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int
J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ochiai H, Nakanishi Y, Fukasawa Y, et al:
A new formula for predicting liver metastasis in patients with
colorectal cancer: immunohistochemical analysis of a large series
of 439 surgically resected cases. Oncology. 75:32–41. 2008.
View Article : Google Scholar
|
|
3
|
Bakalakos EA, Kim JA, Young DC and Martin
EW Jr: Determinants of survival following hepatic resection for
metastatic colorectal cancer. World J Surg. 22:399–404. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Choti MA, Sitzmann JV, Tiburi MF, et al:
Trends in long-term survival following liver resection for hepatic
colorectal metastases. Ann Surg. 235:759–766. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Molkentin JD: The zinc finger-containing
transcription factors GATA-4, -5, and -6. Ubiquitously expressed
regulators of tissue-specific gene expression. J Biol Chem.
275:38949–38952. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Divine JK, Staloch LJ, Haveri H, et al:
GATA-4, GATA-5, and GATA-6 activate the rat liver fatty acid
binding protein gene in concert with HNF-1α. Am J Physiol
Gastrointest Liver Physiol. 287:G1086–G1099. 2004.PubMed/NCBI
|
|
7
|
Fang R, Olds LC and Sibley E:
Spatio-temporal patterns of intestine-specific transcription factor
expression during postnatal mouse gut development. Gene Expr
Patterns. 6:426–432. 2006. View Article : Google Scholar
|
|
8
|
Belaguli NS, Zhang M, Rigi M, Aftab M and
Berger DH: Cooperation between GATA4 and TGF-beta signaling
regulates intestinal epithelial gene expression. Am J Physiol
Gastrointest Liver Physiol. 292:G1520–G1533. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fluck CE and Miller WL: GATA-4 and GATA-6
modulate tissue-specific transcription of the human gene for
P450c17 by direct interaction with Sp1. Mol Endocrinol.
18:1144–1157. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhou B, Francis TA, Yang H, et al: GATA-6
mediates transcriptional activation of aquaporin-5 through
interactions with Sp1. Am J Physiol Cell Physiol. 295:C1141–C1150.
2008. View Article : Google Scholar
|
|
11
|
Agnihotri S, Wolf A, Picard D, Hawkins C
and Guha A: GATA4 is a regulator of astrocyte cell proliferation
and apoptosis in the human and murine central nervous system.
Oncogene. 28:3033–3046. 2009.PubMed/NCBI
|
|
12
|
Capo-chichi CD, Roland IH, Vanderveer L,
et al: Anomalous expression of epithelial
differentiation-determining GATA factors in ovarian tumorigenesis.
Cancer Res. 63:4967–4977. 2003.PubMed/NCBI
|
|
13
|
Kwei KA, Bashyam MD, Kao J, et al: Genomic
profiling identifies GATA6 as a candidate oncogene amplified in
pancreatobiliary cancer. PLoS Genet. 4:e10000812008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Perlman H, Suzuki E, Simonson M, Smith RC
and Walsh K: GATA-6 induces p21(Cip1) expression and G1 cell cycle
arrest. J Biol Chem. 273:13713–13718. 1998.PubMed/NCBI
|
|
15
|
Vuorenoja S, Rivero-Muller A, Kiiveri S,
et al: Adrenocortical tumorigenesis, luteinizing hormone receptor
and transcription factors GATA-4 and GATA-6. Mol Cell Endocrinol.
269:38–45. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miller CT, Moy JR, Lin L, et al: Gene
amplification in esophageal adenocarcinomas and Barrett’s with
high-grade dysplasia. Clin Cancer Res. 9:4819–4825. 2003.
|
|
17
|
Kimchi ET, Posner MC, Park JO, et al:
Progression of Barrett’s metaplasia to adenocarcinoma is associated
with the suppression of the transcriptional programs of epidermal
differentiation. Cancer Res. 65:3146–3154. 2005.
|
|
18
|
Fu B, Luo M, Lakkur S, Lucito R and
Iacobuzio-Donahue CA: Frequent genomic copy number gain and
overexpression of GATA-6 in pancreatic carcinoma. Cancer Biol Ther.
7:1593–1601. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shureiqi I, Zuo X, Broaddus R, et al: The
transcription factor GATA-6 is overexpressed in vivo and
contributes to silencing 15-LOX-1 in vitro in human colon cancer.
FASEB J. 21:743–753. 2007. View Article : Google Scholar
|
|
20
|
Liu X, Zhou B, Xue L, et al:
Metastasis-suppressing potential of ribonucleotide reductase small
subunit p53R2 in human cancer cells. Clin Cancer Res. 12:6337–6344.
2006.PubMed/NCBI
|
|
21
|
Morrisey EE, Musco S, Chen MY, Lu MM,
Leiden JM and Parmacek MS: The gene encoding the mitogen-responsive
phosphoprotein Dab2 is differentially regulated by GATA-6 and
GATA-4 in the visceral endoderm. J Biol Chem. 275:19949–19954.
2000. View Article : Google Scholar
|
|
22
|
Setogawa T, Shinozaki-Yabana S, Masuda T,
Matsuura K and Akiyama T: The tumor suppressor LKB1 induces p21
expression in collaboration with LMO4, GATA-6, and Ldb1. Biochem
Biophys Res Commun. 343:1186–1190. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Caslini C, Capo-chichi CD, Roland IH,
Nicolas E, Yeung AT and Xu XX: Histone modifications silence the
GATA transcription factor genes in ovarian cancer. Oncogene.
25:5446–5461. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mano T, Luo Z, Malendowicz SL, Evans T and
Walsh K: Reversal of GATA-6 downregulation promotes smooth muscle
differentiation and inhibits intimal hyperplasia in balloon-injured
rat carotid artery. Circ Res. 84:647–654. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kamnasaran D, Qian B, Hawkins C, Stanford
WL and Guha A: GATA6 is an astrocytoma tumor suppressor gene
identified by gene trapping of mouse glioma model. Proc Natl Acad
Sci USA. 104:8053–8058. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Haveri H, Westerholm-Ormio M, Lindfors K,
et al: Transcription factors GATA-4 and GATA-6 in normal and
neoplastic human gastrointestinal mucosa. BMC Gastroenterol.
8:92008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Alexandrovich A, Arno M, Patient RK, Shah
AM, Pizzey JA and Brewer AC: Wnt2 is a direct downstream target of
GATA6 during early cardiogenesis. Mech Dev. 123:297–311. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Katoh M and Katoh M: Conserved POU/OCT-
and GATA-binding sites in 5′-flanking promoter region of mammalian
WNT8B orthologs. Int J Oncol. 30:1273–1277. 2007.PubMed/NCBI
|
|
29
|
Weidenfeld J, Shu W, Zhang L, Millar SE
and Morrisey EE: The WNT7b promoter is regulated by TTF-1, GATA6,
and Foxa2 in lung epithelium. J Biol Chem. 277:21061–21070. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang Y, Goss AM, Cohen ED, et al: A
Gata6-Wnt pathway required for epithelial stem cell development and
airway regeneration. Nat Genet. 40:862–870. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
de Lau W, Barker N and Clevers H: WNT
signaling in the normal intestine and colorectal cancer. Front
Biosci. 12:471–491. 2007.PubMed/NCBI
|
|
32
|
Nemer G and Nemer M: Transcriptional
activation of BMP-4 and regulation of mammalian organogenesis by
GATA-4 and -6. Dev Biol. 254:131–148. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Deng H, Makizumi R, Ravikumar TS, Dong H,
Yang W and Yang WL: Bone morphogenetic protein-4 is overexpressed
in colonic adenocarcinomas and promotes migration and invasion of
HCT116 cells. Exp Cell Res. 313:1033–1044. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Houlston RS, Webb E, Broderick P, et al:
Meta-analysis of genome-wide association data identifies four new
susceptibility loci for colorectal cancer. Nat Genet. 40:1426–1435.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han X, Boyd PJ, Colgan S, Madri JA and
Haas TL: Transcriptional upregulation of endothelial cell matrix
metalloproteinase-2 in response to extracellular cues involves
GATA-2. J Biol Chem. 278:47785–47791. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Charron F, Tsimiklis G, Arcand M, et al:
Tissue-specific GATA factors are transcriptional effectors of the
small GTPase RhoA. Genes Dev. 15:2702–2719. 2001. View Article : Google Scholar : PubMed/NCBI
|