Introduction
Osteosarcoma is the primary malignant bone tumor
that occurs the most prevalently in young people. The survival rate
has been improved over the past 20 years. It shows an aggressive
growth from the primary site. In addition, it often shows
micrometastases to the lung before it is being diagnosed (1). A complete removal of the tumor using a
wide excision, accompanied by aggressive adjuvant chemotherapy, is
the standard treatment modality for osteosarcoma (2). Despite the intensity and modification
of chemotherapy, its pulmonary metastasis often leads to fatal
outcomes (3,4). Little is known about the exact
molecular pathophysiology of osteosarcoma. It is, therefore,
imperative that it be clarified to improve the long-term survival
of patients with osteosarcoma, and to contribute to the development
of targeted anticancer therapies.
Transforming growth factor-β (TGF-β) is a
multifunctional cytokine that controls a variety of fundamental
biological activities such as the proliferation, differentiation,
migration, adhesion and apoptosis of the cells (5). That is, the TGF-β signaling pathway
has an inhibitory effect on the proliferation of tumor cells.
Therefore, blockage of the TGF-β signaling pathway is involved in
the progression of many carcinomas (6). It has actually been reported that the
human tumorigenesis is characterized by inactivating mutations of
type II receptor (TβRII), Smad2 and Smad4. In addition, there are
also some reports that the TGF-β receptor is often downregulated in
tumor cells, or otherwise unavailable at the cell surface, thus
allowing tumor cells to escape the growth inhibitory activities
(7).
The pseudoreceptor BAMBI (bone morphogenetic protein
and activin membrane-bound inhibitor), formerly known as NMA, is an
inhibitor of the TGF-β signaling pathway. BAMBI exhibits structural
homology to TGF-βRI but lacks an intracellular kinase domain
(8). In most of the high-grade
carcinomas, the degree of BAMBI expression is abnormally increased,
which leads to the proliferation and metastasis of tumor cells
(9,10). In HepG2 hepatoma cells,
transcription of BAMBI is regulated via a feedback loop which
confers binding of Smad3 and Smad4 to the Smad-responsive elements
of the BAMBI promoter. A recent study has also suggested that BAMBI
is involved in Toll-like receptor 4 and lipopolysaccharide mediated
hepatic fibrosis (11). Moreover,
β-catenin can promote the BAMBI expression indirectly (9). Therefore, it is worth examining
whether BAMBI regulates the Wnt signaling.
Wnt acts through a β-catenin-dependent or canonical
Wnt pathway (12), where β-catenin
is considered as the key component of the above signaling pathway.
It consists of highly conserved secreted ligands that bind to
cell-surface receptors called frizzled and lipoprotein receptor
related protein (LRP)(13). In the
presence of Wnt ligands, β-catenin is accumulated in the cytosol
and thereby blocks its degradation. Stabilized β-catenin
translocates into the nucleus, and forms a complex with lymphocyte
enhancer factor (LEF)/T-cell factor (TCF) family of transcription
factors to activate target genes, many of which are involved in the
development and progression of tumors (14). It has, therefore, been proposed that
the Wnt pathway is involved in the pathogenesis and progression of
many human malignancies.
To date, however, no studies have clarified the
mechanisms by which the Wnt pathway is involved in the progression
of osteosarcoma. Given the above background, we examined the role
of BAMBI in the pathophysiology of osteosarcoma and found that its
overexpression affected the intracellular expression of β-catenin,
a mediator of the Wnt signaling pathway, and thereby had an effect
on the invasion and proliferation of osteosarcoma cells.
Our results indicate that the tumor cell motility
and invasion can be blocked by the inhibition of BAMBI using RNA
interference. This confirms that BAMBI plays a key role in the
pathogenesis of osteosarcoma. Taken together, our results provide
evidence linking the Wnt signaling to the progression of human
osteosarcoma.
Materials and methods
Cell culture and reagents
The human osteosarcoma cell line U2-OS, was
purchased from the American Type Culture Collection (ATCC,
Rockville, MD, USA), and HEK293A cells were from stock maintained
in our laboratory. These cells were grown at 37ºC under a
humidified 5% CO2 atmosphere in DMEM medium supplemented
with 10% fetal bovine serum and 100 U/ml penicillin, 100 μg/ml
streptomycin and 2.5 μg/ml amphotericin B. All reagents were
purchased from Sigma (St. Louis, MO, USA) unless otherwise
noted.
Tissue preparation
Eleven paraffin-embedded samples of human
osteosarcoma and normal tissues were obtained from the Chonbuk
National University Hospital. All the tissue samples were fixed in
a 4% formalin solution. Each block was cut into 5-μM sections and
then stained with specific antibodies. All samples were acquired
during surgeries, with written patient consent, and stored at −80ºC
until use.
Transfection of cells
Transient transfections were performed by the
reverse transfection technique using Lipofectamine™ RNAiMAX kit
(Invitrogen, Carlsbad, CA, USA). Three sequences and a negative
control siRNA were purchased from Bioneer Corp. (Daejon, Korea) and
were initially assessed as potential targets for transfection.
Preparation and amplification of adenovirus were performed as
previously described (15).
Adenoviruses encoding BAMBI were created using the ViraPower
Adenovirus Expression system according to the manufacturer's
instruction (Invitrogen). Site-specific recombination between entry
vectors (BAMBI-pENTR) and the adenoviral destination vector
(pAd/PL-DEST) were established with LR Clonase II (Invitrogen). The
control adenovirus (Ad-LacZ) was obtained from R.H. Unger
(University of Texas Southwestern Medical Center, Dallas, TX, USA).
Virus (100 MOI) was added to the cells and then incubated for 6 h
at 37ºC. After conventional culturing of the cells for 48 h, the
cells were collected to perform the experiment.
Histological and immunohistochemical
studies
Tissue sections (5 μm) were stained with hematoxylin
and eosin (H&E) dye for light microscopy. Immunohistochemical
staining was performed under the Dako Envision system (Dako,
Carpinteria, CA, USA) and was immunostained with antibody against
BAMBI (R&D Systems, Minneapolis, MN, USA).
MTT assay for cell proliferation
The viability of cultured cells was determined by
assaying the reduction of
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
to formazan. After transfection with BAMBI adenovirus (100 MOI) or
siRNA, the cells in 96-well plates were washed twice with PBS and
MTT (100 μg/100 μl of PBS) was added to each well. Cells were then
incubated at 37ºC for 1 h, and DMSO (100 μl) was added to dissolve
the formazan crystals. Absorbance was measured at 570 nm with a
model SpectraMax Plus (Molecular Devices, Sunnyvale, CA, USA).
Wound healing assay
Cells were seeded at a density of 106 in
6-well plates and then cultured for 24 h. A wound was made using
the tip of a pipette as previously described (14). After 8 h, the cells were fixed with
3% formaldehyde in PBS for 15 min and images of the wound were
generated using different fields.
Matrigel invasion assay
The invasion assays were carried out using a
chemotaxis chamber (BD Biosciences, San Jose, CA, USA) and coated
with 10 μg/ml Matrigel (BD Biosciences). Then, 100 μl of
105 cells/ml suspension was introduced into each well.
Thus, the cells were allowed to invade for 24 h and then fixed with
methanol. This was followed by crystal violet staining.
Gelatin zymography
The activities of two enzymes, matrix
metalloproteinase (MMP)-2 and -9 were assayed by gelatin zymography
(16). Samples of serum-free
conditioned medium were electrophoresed on a 10% SDS-polyacryamide
gel. After electrophoresis, the gel was washed in 2.5% Triton X-100
for 1 h and then incubated at 37ºC for 24 h in activation buffer
(50 mM Tris-HCl, pH 7.5, 150 mM NaCl, 10 mM CaCl2 and
0.02% NaN3). After staining with Coomassie brilliant
blue R-250, enzymatic activities were detected as clear bands
against the blue background.
Cell cycle analysis
Cells (1–5×105) in 6-well plates were
transfected with Ad-BAMBI, Ad-LacZ (100 MOI) or BAMBI siRNA and
control siRNA for 48 h. After trypsinization and collection, cells
were washed three times with PBS and fixed in 70% ethanol overnight
at 4ºC. Cells were then washed three times in PBS with 0.1% BSA.
Cells were incubated with 5 mg/ml of RNase A (DNase free) and 50
mg/ml of PI for 90 min at 4ºC. The percentages of cells in
different phases of the cell cycle were measured with a FACStar
flow cytometer (Becton-Dickinson, San Jose, CA, USA).
Western blot analysis
The homogenized osteosarcoma tissues or cells were
kept in lysis buffer (M-PER; Thermo Scientific). The lysates were
separated by 10% SDS-PAGE and transferred to PVDF membranes
(Millipore, Billerica, MA, USA) by electroblotting. After blocking
with 5% skim-milk, the blot was probed with primary antibodies for
BAMBI (R&D Systems), β-catenin, cyclin D1, CDK2, CDK6, p21 and
β-actin (all from Santa Cruz Biochemicals, Santa Cruz, CA,
USA).
RNA isolation and the real-time RT-PCR
analysis
Total RNA was extracted from the cells using the
TRIzol reagent (Invitrogen). First-strand cDNA was generated using
the random hexamer primer provided in the first-strand cDNA
synthesis kit (Applied Biosystems, Foster City, CA, USA). Specific
primers for each gene (Table I)
were designed using Primer Express software (Applied Biosystems).
The PCR reaction was carried out by the ABI Prism 7900HT Sequence
Detection system (Applied Biosystems). All the experiments were
performed in triplicate.
 | Table ISequences and accession numbers for
forward and reverse primers for the real-time RT-PCR. |
Table I
Sequences and accession numbers for
forward and reverse primers for the real-time RT-PCR.
| Gene | Sequences for
primers | Accession no. |
|---|
| β-catenin | Forward:
ATTGTCCACGCTGGATTTTC
Reverse: TCGAGGACGGTCGGACT | NM_001904 |
| c-Myc | Forward:
CACCGAGTCGTAGTCGAGGT
Reverse: TTTCGGGTAGTGGAAAACCA | NM_002467 |
| p21 | Forward:
CATGGGTTCTGACGGACAT
Reverse: AGTCAGTTCCTTGTGGAGCC | NM_000389 |
| CDK2 | Forward:
GAATCTCCAGGGAATAGGGC
Reverse: TGGTACCGAGCTCCTGAAAT | NM_001798 |
| CDK6 | Forward:
TGTCTGTTCGTGACACTGTGC
Reverse: ATGCCGCTCTCCACCAT | NM_001259 |
| MMP-2 | Forward:
GGAAAGCCAGGATCCATTTT
Reverse: ATGCCGCCTTTAACTGGAG | NM_004530 |
| MMP-9 | Forward:
TTGGTCCACCTGGTTCAACT
Reverse: ACGACGTCTTCCAGTACCGA | NM_004994 |
| BAMBI | Forward:
GGCAGCATCACAGTAGCATC
Reverse: GATCGCCACTCCAGCTACAT | NM_012342 |
| Cyclin | D1 Forward:
GGCGGATTGGAAATGAACTT
Reverse: TCCTCTCCAAAARGCCAGAG | NM_053056 |
Statistical analysis
Statistical analysis of the data was performed using
ANOVA and Duncan's test. Differences with a P-value <0.05 were
considered to indicate a statistically significant result.
Results
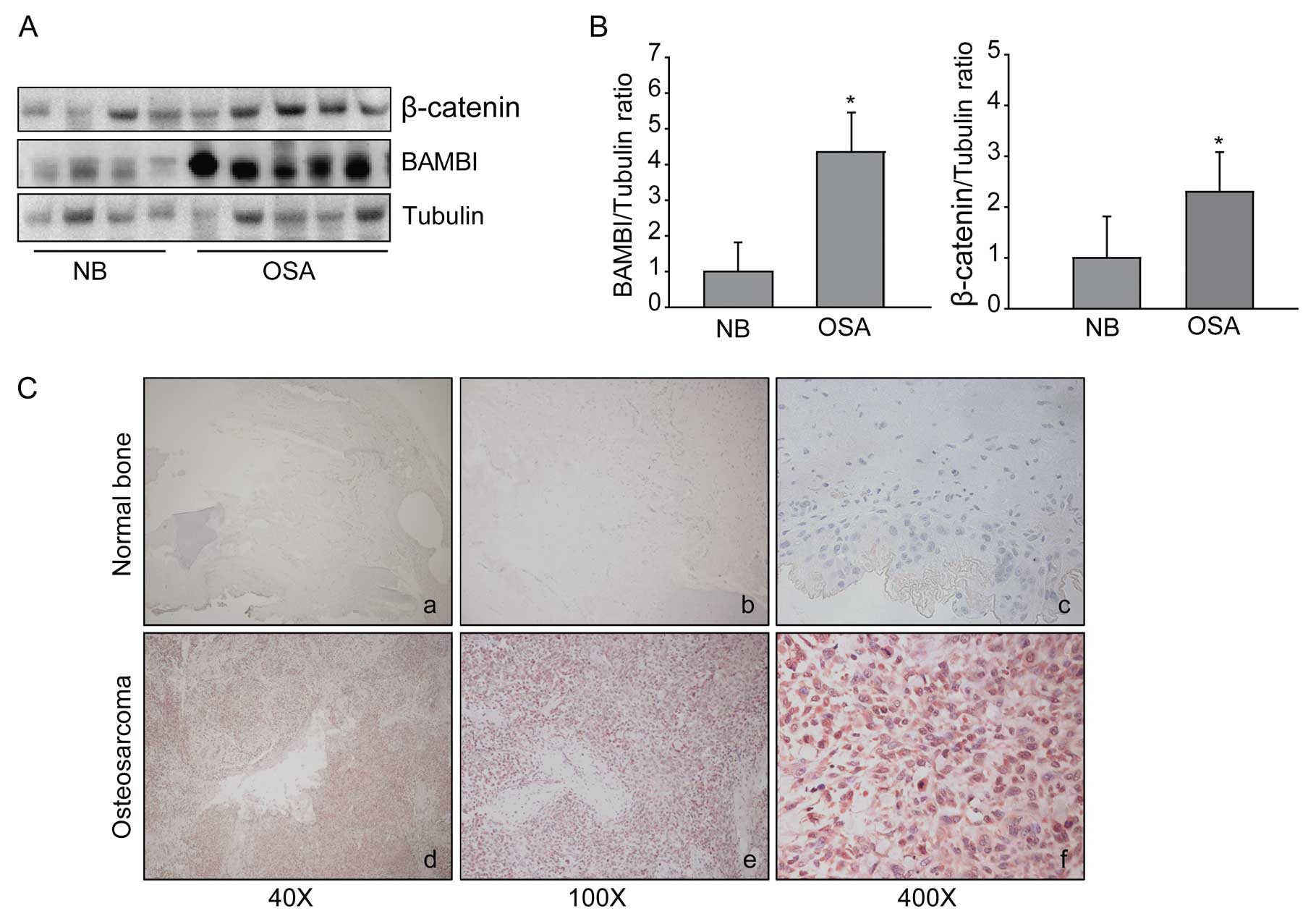
The overexpression of BAMBI in human
osteosarcoma tissues
To assess the effects of BAMBI on the pathogenesis
of osteosarcoma, we analyzed the degree of BAMBI expression using
immunohistochemical studies and western blot analysis in human
osteosarcoma and normal tissues. The western blot analysis showed
that the degree of BAMBI expression was 4-fold higher in
osteosarcoma tissues as compared with normal tissues (Fig. 1A and B). In addition, the degree of
β-catenin expression was also significantly higher in osteosarcoma
tissues (Fig. 1A and B).
Immunohistochemical studies showed that the BAMBI expression was
observed in the cytoplasm of human osteosarcoma cells (Fig. 1C).
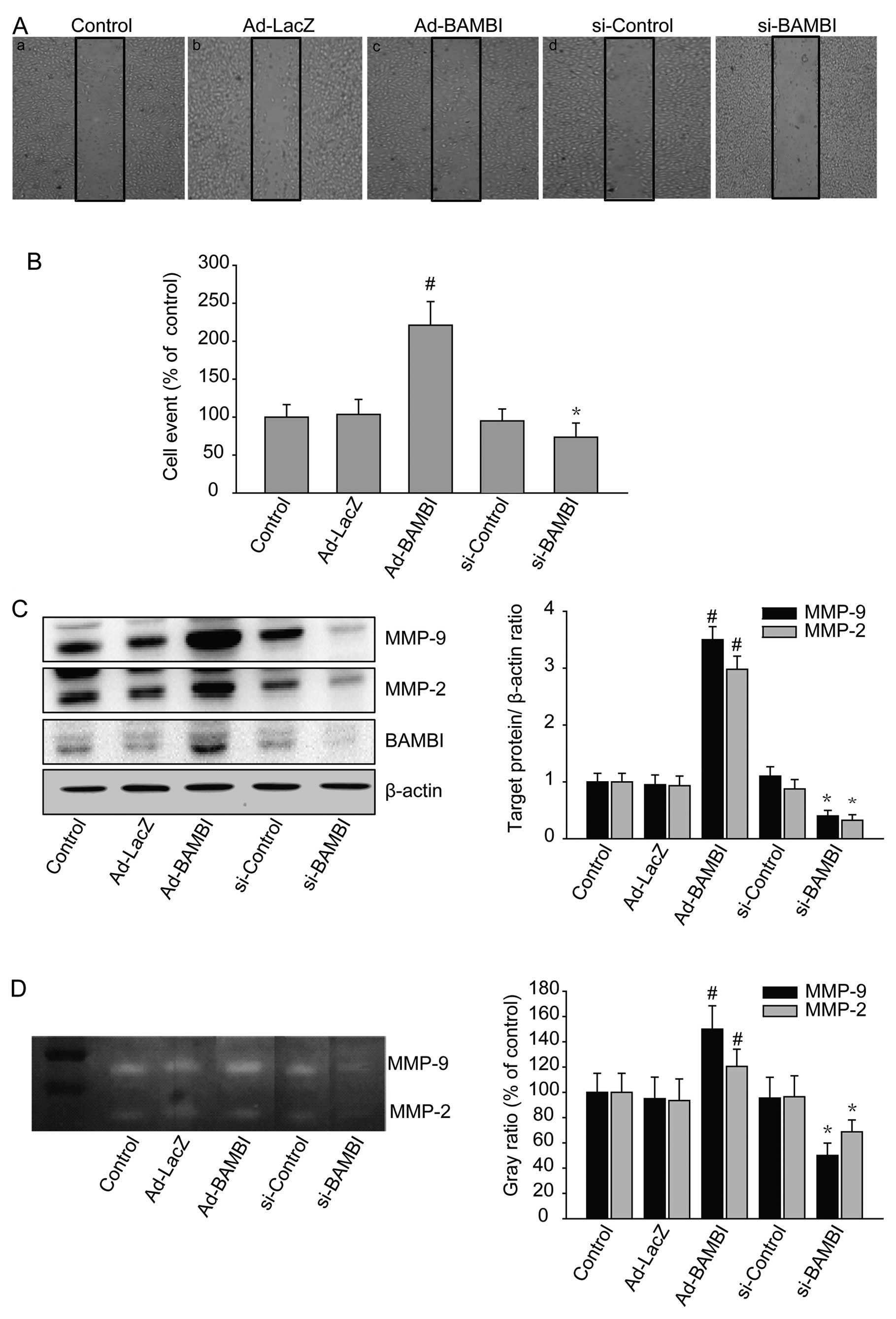
Effects of BAMBI on the migration and
invasion of human osteosarcoma cells
After transfection with BAMBI adenovirus or BAMBI
siRNA for 48 h, we analyzed the migration (Fig. 2A) and invasion (Fig. 2B) of human osteosarcoma cell line
(U2-OS). Moreover, following the transfection, there was a
significant change in the migration of U2-OS in the wound healing
experiment. In the cells with an overexpression of BAMBI, there was
an increase in the invasion of osteosarcoma cells. With the
suppression of the expression of BAMBI (knockdown of endogenous
BAMBI), the migration and invasion of osteosarcoma cells was
effectively blocked. The expression of BAMBI in these cells was
confirmed on a western blot analysis (Fig. 2C). The degree of BAMBI expression
was subject to the transfection. Moreover, the degree of MMP-2
expression was also associated with that of BAMBI expression. It is
generally known that MMP-2 and MMP-9 are released into the culture
medium. We, therefore, analyzed the effects of BAMBI based on their
activities by zymography (Fig. 2D).
This showed that the BAMBI had an effect in improving the
activities of MMP-2 and MMP-9.
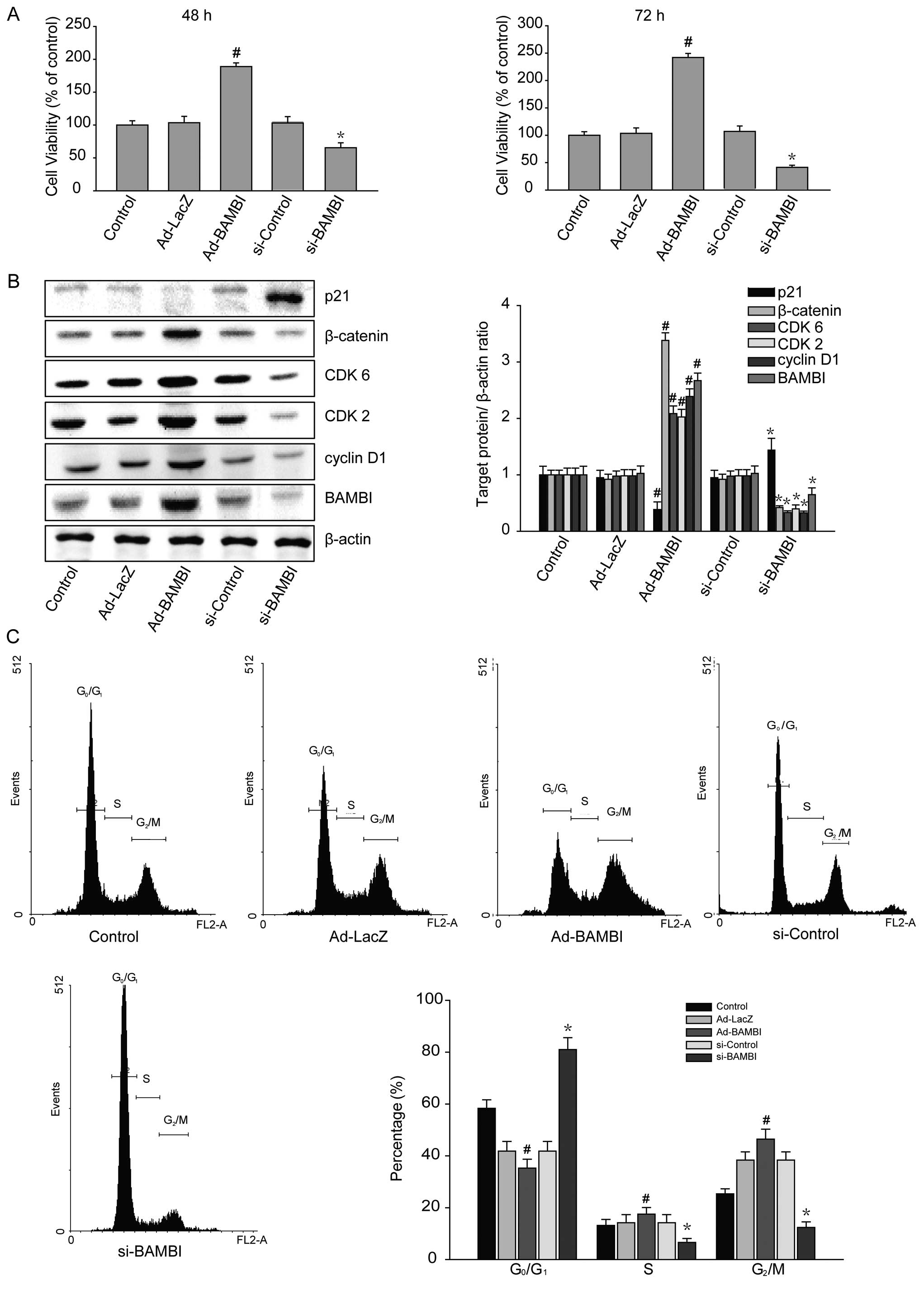
The effects of BAMBI in inducing the cell
proliferation and the transition of G0/G1 to
G2/M in human osteosarcoma cells
After transfection with BAMBI adenovirus or siRNA
for 48 h, cellular proliferation was analyzed in human osteosarcoma
cell line U2-OS. According to the MTT analysis, the overexpression
of BAMBI was associated with the increased proliferation of
osteosarcoma cells by ~189±5.5% at 48 h and 242±7.5% at 72 h
(Fig. 3A). We analyzed the effects
of BAMBI on the cell cycle regulatory molecules that play a key
role in the cell cycle transition. The levels of cyclin D1 and
β-catenin and their associated cyclin-dependent kinases (CDKs),
CDK2 and CDK6, were increased in the cells with an overexpression
of BAMBI (Fig. 3B). In these cells,
however, the degree of the expression of cell cycle inhibitor, p21,
was decreased. Following an analysis of the variability in the
degree of BAMBI expression, the cell cycle was determined by
fluorescence activated cell sorting (FACS) analysis (Fig. 3C). With the overexpression of BAMBI,
the proportion of cells with G0/G1 DNA copies
was decreased to 35.26±3.41%. In the control group, however, it was
58.36±3.27%. The proportion of cells with S and G2/M DNA
copies was 17.52±2.52 and 46.44±3.83%, respectively, with the
overexpression of BAMBI, whereas it was 13.17±2.31 and 25.33±1.96%
in the corresponding order in the control group. In the control
group, comprising virus-transfected cells, it did not change; the
proportion of cells with G0/G1, S and
G2/M DNA copies was 41.81±3.75, 14.22±3.09 and
38.35±3.18%, respectively. Consistent with the findings that the
cell growth was increased, the BAMBI promoted the transition from
G0/G1 to G2/M in the oteosarcoma
cells.
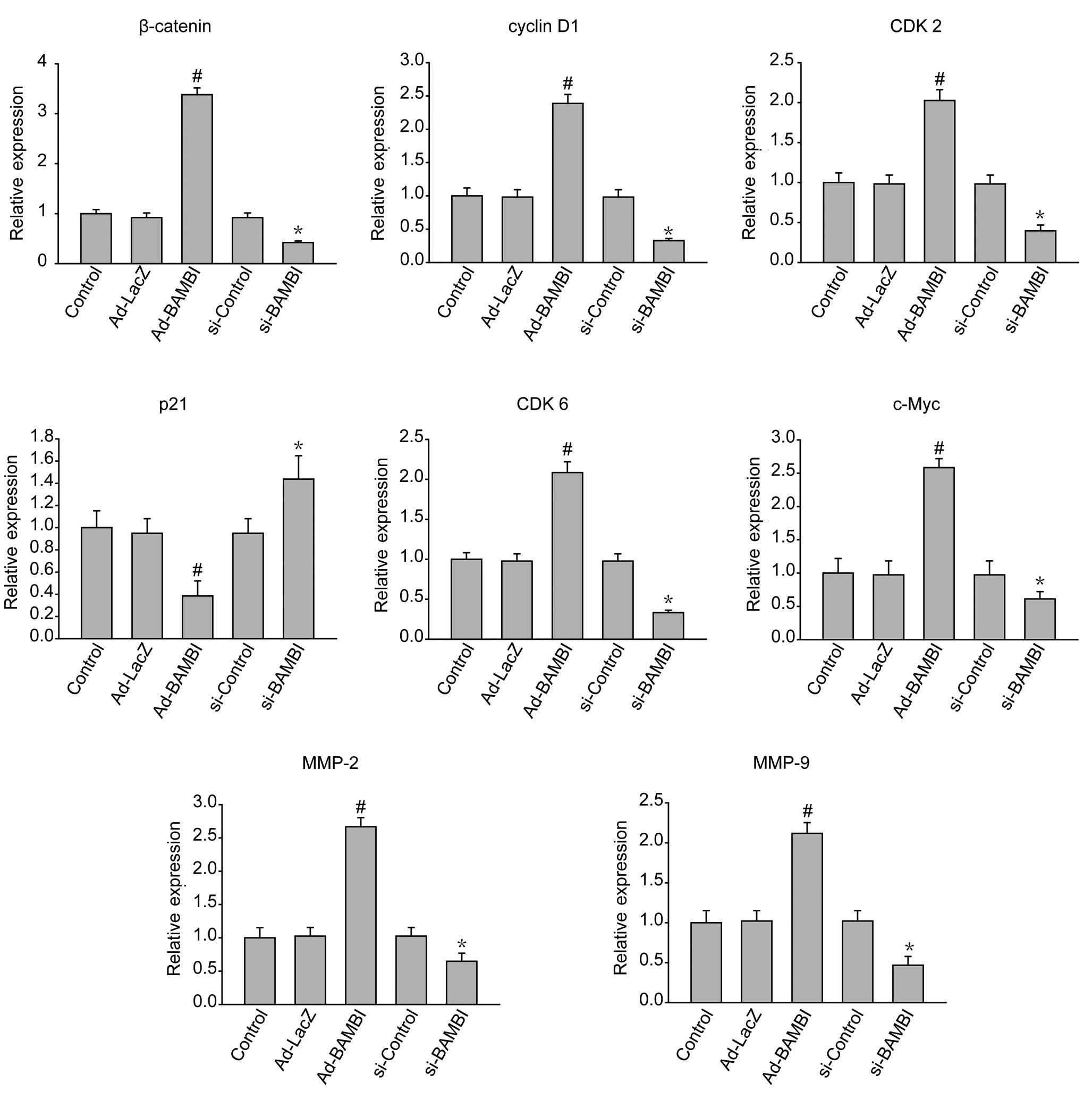
Real-time RT-PCR analysis of
oncogenes
The RNA was isolated from cells transfected with
BAMBI adenovirus or siRNA. Then, the degree of the expression of
several genes related to oncogenesis were analyzed (Fig. 4). The upregulation of BAMBI by
adenovirus or downregulation of BAMBI by RNA interference were
confirmed by measuring protein levels. The β-catenin levels were
consistent with the modification of BAMBI. There were alterations
in the expression of cyclin D1, CDK2, CDK6 and c-Myc, known as
Tcf/Lef target genes (17), and the
relevant cell cycle genes with the treatment with BAMBI. The degree
of the expression of MMP-2 and MMP-9, both of which are involved in
the invasion and metastasis of the tumor, was also increased with
the overexpression of BAMBI.
Discussion
To date, it has been demonstrated that the BAMBI
signaling plays a role in the pathogenesis of many human
malignancies (18). Still, however,
little is known about the biological significance of its role in
the pathogenesis of osteosarcoma. Previous studies have shown that
human osteosarcomas are positive for the expression of both BAMBI
and β-catenin, thus, suggesting that their pathogenesis may be
involved in multiple Wnt signaling pathways. The presence of
metformin-mediated BAMBI suggests that the pathogenesis of
osteosarcoma is associated with the canonical Wnt-β-catenin pathway
(19). Our results provided
additional evidence demonstrating that the BAMBI is involved in the
pathogenesis of osteosarcoma by altering the degree of β-catenin
expression via the Wnt signaling pathway.
In the present study, we performed
immunohistochemistry to compare the degree of BAMBI expression
between the normal tissue and the osteosarcoma specimen sampled
from humans. On immunohistochemical staining of the 11 human tissue
samples, the degree of BAMBI expression was significantly lower in
the normal tissue and that of BAMBI expression was significantly
higher in osteosarcoma specimens. In agreement with these findings,
the western blot analysis showed that the degree of BAMBI
expression was four times higher in osteosarcoma specimen as
compared with the normal tissue. But the degree of β-catenin
expression was significantly higher in the oteosarcoma specimen and
it was only 2.5 times higher as compared with the normal
tissue.
It has been proposed that the BAMBI plays a role as
an antagonist in blocking the signal transduction pathway where the
TGF-β family proteins are involved. Recent studies have shown that
the BAMBI has an effect in regulating the Wnt activity by
increasing the response to the transcription of Wnt, as confirmed
by the Wnt-responsive LEF-luciferase and TOF-luciferase (20). Furthermore, the BAMBI interacts with
receptor Frizzled and LRP-5/6, thus, increasing its effects on the
Wnt signaling pathway. In this context, our results demonstrated
its effects in stimulating the Wnt/β-catenin signaling pathway on
the real-time RT PCR analysis. In addition, our results also showed
that BAMBI stimulated nuclear accumulation of β-catenin. The
overexpression of BAMBI in human osteosarcoma cells strongly
induces β-catenin and the Wnt-induced transcription of target
genes, cyclin D1 and CDKs. Moreover, the Wnt signaling pathway is
blocked by knockdown of endogenous BAMBI.
The Wnt signaling pathway is activated in most human
cancers. β-catenin phosphorylation occurs sequentially by CKIα and
GSK3β, which is followed by degradation via the
ubiquitin-proteasome pathway (21).
Loss-of-function mutations of adenomatous polyposis coli and
stabilization mutations of β-catenin lead to the accumulation of
β-catenin (22). A wide spectrum of
β-catenin targets, such as the proteins involved in cell cycle,
cell-matrix interactions, migration and invasion, have been
identified (23).
The next step of metastasis is the invasion of tumor
cells into the basement membrane, where distinct events are
involved and these include the cell motility and MMP expression.
Invasive tumors exhibit active migration and high levels of MMPs
(24). Our results showed that the
BAMBI might be involved in the upregulation of β-catenin nuclear
translocation in human osteosarcoma cells, thus, suggesting that it
has an effect in modulating the activities of the Wnt signaling
pathway. It is apparent that the effects of BAMBI in promoting the
Wnt signaling pathway are associated with a higher potential of
tumor invasion and a higher degree of MMP-2 expression as compared
with the cells where the BAMBI expression was blocked. Given that
β-catenin preferentially translocates into the nucleus during the
cell migration, it is probable that the BAMBI can increase the
motility of osteosarcoma cells by upregulating the nuclear
translocation of β-catenin. Taken together, our results suggest
that the degree of metastatic potential would be increased in the
cells with an overexpression of BAMBI.
With the nuclear translocation of β-catenin, target
genes, such as cyclin D1 and CDKs, are activated. This leads to the
promotion of the growth of tumor cells (25). Cyclin D1, along with CDK2, are the
driving forces behind the G1-S transition of the cell
cycle, whereas p21 induces G1 and G2/M phase
cell cycle arrest (26). This has
been reported in our previous studies (27). Significant increases in β-catenin,
cyclin D1 and CDK6 were observed in the BAMBI-transfected
osteosarcoma cells. The increase in these checkpoint proteins leads
to uncontrolled cell growth and aberrant cell function and the
cells quickly traverse the G0 to G1 and/or
G1 to S transitions. Finally, we found that BAMBI also
cooperates with the Wnt not only to stimulate the expression of
cyclin D1 and c-Myc but also to promote cell cycle progression.
Osteosarcoma is the most prevalent cancer mainly
affecting adolescents and young adults. Due to a delayed diagnosis
and a lack of the effective treatments, patients with osteosarcoma
are at increased risks of recurrence and distant metastasis
(23). Currently, the surgical
treatment is the best treatment option. In a clinical setting,
however, it is not easy to detect the osteosarcoma at the earliest
opportunity possible for surgical treatment. It has become a very
important issue to develop the optimal diagnostic and treatment
methods for osteosarcoma, which is essential for reducing its
incidence and burden.
To summarize, our results showed that BAMBI
modulated the Wnt-β-catenin pathway in vitro in osteosarcoma
specimen, thus, suggesting that it can upregulate the invasion and
migration of osteosarcoma. In addition, our results showed that
BAMBI enhanced the cell viability by regulating the cell cycle in
the presence of cyclin D1. Collectively, our results indicate that
the BAMBI might have a potential to prevent or treat
osteosarcoma.
Acknowledgements
This study was supported by the National Research
Foundation of Korea (NRF) grant funded by the Korea government
(MEST) (No. 2011-0028928).
References
|
1
|
Bacci G, Longhi A, Versari M, Mercuri M,
Briccoli A and Picci P: Prognostic factors for osteosarcoma of the
extremity treated with neoadjuvant chemotherapy: 15-year experience
in 789 patients treated at a single institution. Cancer.
106:1154–1161. 2006.PubMed/NCBI
|
|
2
|
Ferguson WS and Goorin AM: Current
treatment of osteosarcoma. Cancer Invest. 19:292–315. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bacci G, Briccoli A, Ferrari S, et al:
Neoadjuvant chemotherapy for osteosarcoma of the extremities with
synchronous lung metastases: treatment with cisplatin, adriamycin
and high dose of methotrexate and ifosfamide. Oncol Rep. 7:339–346.
2000.PubMed/NCBI
|
|
4
|
Saeter G, Hoie J, Stenwig AE, Johansson
AK, Hannisdal E and Solheim OP: Systemic relapse of patients with
osteogenic sarcoma. Prognostic factors for long term survival.
Cancer. 75:1084–1093. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Massague J: TGFbeta in cancer. Cell.
134:215–230. 2008. View Article : Google Scholar
|
|
6
|
Osuna D and de Alava E: Molecular
pathology of sarcomas. Rev Recent Clin Trials. 4:12–26. 2009.
View Article : Google Scholar
|
|
7
|
Massague J, Blain SW and Lo RS: TGFβ
signaling in growth control, cancer, and heritable disorders. Cell.
103:295–309. 2000.
|
|
8
|
Onichtchouk D, Chen YG, Dosch R, et al:
Silencing of TGF-β signalling by the pseudoreceptor BAMBI. Nature.
401:480–485. 1999.
|
|
9
|
Fritzmann J, Morkel M, Besser D, et al: A
colorectal cancer expression profile that includes transforming
growth factor β inhibitor BAMBI predicts metastatic potential.
Gastroenterology. 137:165–175. 2009.PubMed/NCBI
|
|
10
|
Yan X, Lin Z, Chen F, et al: Human BAMBI
cooperates with Smad7 to inhibit transforming growth factor-β
signaling. J Biol Chem. 284:30097–30104. 2009.PubMed/NCBI
|
|
11
|
Seki E, De Minicis S, Osterreicher CH, et
al: TLR4 enhances TGF-β signaling and hepatic fibrosis. Nat Med.
13:1324–1332. 2007.
|
|
12
|
van Amerongen R, Mikels A and Nusse R:
Alternative Wnt signaling is initiated by distinct receptors. Sci
Signal. 1:re92008.PubMed/NCBI
|
|
13
|
Nusse R and Varmus HE: Wnt genes. Cell.
69:1073–1087. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Polakis P: Wnt signaling in cancer. Cold
Spring Harb Perspect Biol. 4:2012. View Article : Google Scholar
|
|
15
|
Lee JH, Song MY, Song EK, et al:
Overexpression of SIRT1 protects pancreatic β-cells against
cytokine toxicity by suppressing the nuclear factor-κB signaling
pathway. Diabetes. 58:344–351. 2009.
|
|
16
|
Aizawa J, Sakayama K, Kamei S, et al:
Effect of troglitazone on tumor growth and pulmonary metastasis
development of the mouse osteosarcoma cell line LM8. BMC Cancer.
10:512010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tetsu O and McCormick F: β-catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999.
|
|
18
|
Khin SS, Kitazawa R, Win N, et al: BAMBI
gene is epigenetically silenced in subset of high-grade bladder
cancer. Int J Cancer. 125:328–338. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Subramaniam N, Sherman MH, Rao R, et al:
Metformin-mediated Bambi expression in hepatic stellate cells
induces prosurvival Wnt/β-catenin signaling. Cancer Prev Res.
5:553–561. 2012.PubMed/NCBI
|
|
20
|
Lin Z, Gao C, Ning Y, He X, Wu W and Chen
YG: The pseudoreceptor BMP and activin membrane-bound inhibitor
positively modulates Wnt/β-catenin signaling. J Biol Chem.
283:33053–33058. 2008.PubMed/NCBI
|
|
21
|
Liu C, Li Y, Semenov M, et al: Control of
β-catenin phosphorylation/degradation by a dual-kinase mechanism.
Cell. 108:837–847. 2002.
|
|
22
|
Morin PJ, Sparks AB, Korinek V, et al:
Activation of beta-catenin-Tcf signaling in colon cancer by
mutations in β-catenin or APC. Science. 275:1787–1790. 1997.
|
|
23
|
Willert K and Jones KA: Wnt signaling: is
the party in the nucleus? Genes Dev. 20:1394–1404. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Stetler-Stevenson WG, Aznavoorian S and
Liotta LA: Tumor cell interactions with the extracellular matrix
during invasion and metastasis. Annu Rev Cell Biol. 9:541–573.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Shapiro GI: Cyclin-dependent kinase
pathways as targets for cancer treatment. J Clin Oncol.
24:1770–1783. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiong Y, Hannon GJ, Zhang H, Casso D,
Kobayashi R and Beach D: p21 is a universal inhibitor of cyclin
kinases. Nature. 366:701–704. 1993. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhou L, Park BH, Park JH, et al:
Overexpression of the prolyl isomerase PIN1 promotes cell growth in
osteosarcoma cells. Oncol Rep. 29:193–198. 2013.PubMed/NCBI
|