Introduction
The immune microenvironment, biology and prognosis
of hepatocellular carcinoma (HCC) have recently been the focus of
extensive research. Local cellular immune status has been shown to
correlate with invasion, recurrence and metastasis in HCC (1). The liver’s unique immunological
properties have also aroused considerable interest (2). Liver fibrosis is the result of chronic
liver injury, during which hepatic stellate cells (HSCs)
proliferate and differentiate into myofibroblasts and express
α-smooth muscle actin (α-SMA) (3,4). They
also express intercellular adhesion molecule 1 (ICAM-1) and
vascular cell adhesion molecule 1 (VCAM-1) and secret chemokines
through CD40 expression, and thus, played a role in the regulation
of lymphocyte recruitment and migration (5,6). Two
recent studies confirmed that T-cell apoptosis can be induced in
liver fibrosis and is spatially associated with activated HSCs,
suggesting a direct interaction between lymphocytes and HSCs in
liver (7,8). In vitro and in vivo
studies have demonstrated that culture-activated HSCs inhibit the
function of T lymphocytes through the promotion of T-cell apoptosis
(9,10).
HCC contains a large number of α-SMA positive HSCs
(tHSCs), which are correlated with the invasion and metastasis of
HCC (11). However, few studies
have examined the immunological function of HSCs and the
interaction between the lymphocytes and the tHSCs. In addition, the
relationship between the T-cell apoptosis and the HCC metastasis
remains unclear. In order to investigate the mechanism whereby
tHSCs regulate T-cell infiltration in HCC, as well as HCC
metastasis, we established a rat HCC metastasis model, and studied
the interactions between the co-cultured T cells and the tHSCs.
Materials and methods
Tumor cell line and animals
The rat HCC cell line, McA-RH7777 (MRH), was
obtained from the American Type Culture Collection (ATCC;
Rockville, MD, USA). Buffalo rats (6–8 weeks old) were obtained
from the Charles River Laboratories (Davis, CA, USA). All animal
studies were conducted in accordance with the animal care policy of
the Fudan University and the Animal Research Committee.
Rat tumor model
Intrahepatic tumor implantation was carried out
according to Yang et al(12)
with minor modifications. MRH cells were injected into the flank of
Buffalo rats. HCC tissue was recovered from the transplanted rats 1
month after tumor cell injection, and cut into cubes ~2×1×1
mm3. The rats were anesthetized with ketamine. A small
subxiphoid midline incision was made to expose the left lateral
lobe of the liver. A small superficial incision was made in the
liver and a cube (size, 2×1×1 mm3) of tumor was
transplanted. The liver incision was closed with 7–0 suture to
avoid any possible early peritoneal seeding of the HCC. Tumour
extensions were determined by ultrasound on days 7, 14, 21 and 28
after tumor establishment.
Lung harvesting
Whole lungs of the recipient rats were harvested on
days 7, 14, 21 and 28 for quantitation of metastasis as follows.
The trachea of the rat was catheterized with a 22-gauge Angiocath
(Becton Dickinson Vascular Access, Sandy, UT, USA) and the lungs
were insufflated with 15% India ink solution. The lungs were
preserved in Fekette’s solution for 24 h, followed by 10% formalin.
The tumor nodules on the surfaces of the lungs and within the lungs
were counted under an operating microscope at ×10 magnification
(13).
Hematoxylin and eosin (H&E)
staining
Cross-sections of liver were checked for the
presence of tumor tissue on days 7, 14, 21 and 28 after
intrahepatic tumor transplantation. Rats were randomly selected for
sacrifice on days 7, 14, 21 and 28 (n=4 each), respectively. Livers
were fixed in formalin, embedded in paraffin, and stained with
H&E for histological examination.
Immunofluorescence
Liver biopsies taken at all check-points were
incubated overnight at room temperature with phosphate-buffered
saline (PBS), 10% sucrose and 4% formaldehyde solution. Specimens
were frozen at −70°C and 7-μm-thick frozen sections were prepared
using a cryostat (Leica CM3000; Meyer Instruments, Houston, TX,
USA). Cellular hyperpermeability was achieved using the 0.3% Triton
(Sigma, Steinheim, Germany). Bovine serum albumin 1% was used to
block non-specific background staining. Following washing with PBS,
the slides were incubated with primary antibody to lymphocyte
markers and anti-α-SMA rabbit primary monoclonal antibody (mAb)
(Roche, Basel, Switzerland) at a dilution of 1:750 for 1 h at room
temperature in the dark, then washed three times with PBS. The
secondary antibody Cy-5 (BD Biosciences, Franklin Lakes, NJ, USA)
was then added, followed by three washes with PBS. In each
double-stained set, a single lymphocyte subset was stained using
anti-α-SMA. Fluoresceine isothiocyanate (FITC)-conjugated mouse
anti-rat CD4 and allophycocyanin (APC)-conjugated mouse anti-rat
CD8 cell mAbs (BD Biosciences) were used, respectively. Sections
were stacked and covered with Fluoromount-G (Birmingham, AL, USA)
to maintain staining and prevent fluorescent bleaching. Sections
were then stored at 4°C until analysis by confocal microscopy
(14,15).
Terminal deoxynucleotidyl
transferase-mediated dUTP nick end labeling (TUNEL) method
For the detection of apoptosis,
periodate-lysine-paraformaldehyde-fixed cryosections were stained
by the TUNEL technique, using an In Situ Apoptosis Detection kit
(Roche), according to the manufacturer’s instructions. Sections
were incubated at 37°C for 1 h with terminal deoxynucleotidyl
transferase in the labeling-safe buffer including FITC, and
observed under a confocal laser scanning microscope.
Double staining
After TUNEL staining, cryosections were incubated
with Cy5-conjugated anti-rat α-SMA mAb (1:500 dilution; Roche) or
phycoerythrin (PE)-conjugated anti-rat CD3 mAb (1:100 dilution;
Serotec) overnight at room temperature. They were observed under a
confocal laser scanning microscope. FITC images and PE or Cy5
images were entered separately into the computer and superimposed
using Adobe Photoshop software (Adobe Systems UK, Uxbridge,
UK).
Isolation and culture of HSCs
Rats were anesthetized by intraperitoneal injection
of ketamine, 10 mg/100 g body weight. HSCs were prepared from the
livers of normal Buffalo rats or HCC tissue, as previously reported
(16). In brief, the abdominal
cavity of the anesthetized animals was opened to canulate the
portal vein. The liver was subsequently perfused at a flow rate of
10 ml/min with Gey’s balanced salt solution (GBSS) without
Ca2+ and Mg2+ for 10 min, followed by 100 ml
of 0.12% pronase E (Roche) dissolved in GBSS with Ca2+
and Mg2+ (GBSS+) for 10 min. Then 100 ml of
0.1% collagenase (Roche) dissolved in GBSS+ was
recirculated for 20 min. The normal liver and HCC tissue were then
excised, dissected and incubated for 30 min at 37°C with continuous
shaking, with 0.04% pronase E, 0.05% collagenase and 0.002% DNase I
(Sigma) in 100 ml GBSS+. After digestion, the cell
suspension was passed through a 0.22-μm mesh and centrifuged at 480
× g for 10 min. Subsequently, cells were purified by 8% Nycodenz
(Sigma) gradient centrifugation. HSCs were grown in DMEM medium
(Gibco-BRL) containing 10% fetal calf serum (FCS) before the
experiments were performed. Cell viability determined by trypan
blue exclusion was >90%. The purity of quiescent (q) HSCs and
tHSCs ranged from 90–95%, as determined by desmin immunostaining.
tHSC activation was determined by α-SMA immunostaining. qHSCs and
tHSCs were obtained by culture on uncoated plastic dishes for 2
days (17).
Culture of dendritic cells (DCs)
Bone marrow cells isolated from F344 rat (Davis, CA,
USA) femurs were lysed of red blood cells using red blood cell
lysis buffer (Sigma), cultured in RPMI-1640 medium containing 10%
v/v heat-inactivated FCS, 20 mmol/l hydroxyethylpiperazine-N-2
ethanesulfonic acid, 2 mmol/l L-glutamine, 0.1 mmol/l non-essential
amino acids, 1 mmol/l sodium pyruvate, 20 μmol/l 2-mercaptoethanol,
and antibiotics (100 U/ml penicillin and 100 mg/ml streptomycin)
(subsequently referred to as complete medium) in the presence of
rat recombinant granulocyte-macrophage colony-stimulating factor (4
ng/ml) and interleukin-4 (1,000 U/ml) (both from BD Bioscences).
Non-adherent cells were released spontaneously from the
proliferating cell clusters, harvested, washed, and resuspended in
complete medium, as previously described (18).
Co-culture of HSCs and splenic T
cells
Proliferation of nylon-wool-eluted Buffalo rat
spleen T cells (2×105/well in 100 μl) was stimulated via
allogeneic DCs in a one-way mixed leukocyte reaction.
Non-stimulated (quiescent, qT) and DC-stimulated (activated, aT)
splenic T cells were added to qHSCs or tHSCs
(1×106/well) and further incubated for 48 h. Three rats
were used for each group. Apoptotic T cells were identified via
double staining with PE-conjugated anti-CD3 mAb and TUNEL.
Following surface CD3 staining, cells were fixed in 4%
paraformaldehyde and permeabilized with 0.1% Triton X-100 and 0.1%
sodium citrate. The TUNEL reaction mixture from the Cell Death
Detection kit (BD Biosciences, San Diego, CA, USA) was then added
according to the manufacturer’s instructions. Cells incubated with
the labeling solution in the absence of terminal transferase were
used as negative controls. The percentage of TUNEL-positive
lymphocytes was then calculated.
Confocal microscopy and image
capture
A confocal laser scanning microscope (14900
Superfusion System; Radnoti, Monrovia, CA, USA) was used to analyze
the stained sections. Fifty images were collected from each liver
using a charge-coupled device camera (Hall 100) and analyzed using
Zeiss LSM Image Browser software. Image processing was performed
using Adobe Photoshop software (Adobe Systems UK) (19).
Quantitative analysis of immunopositive
cells
Livers taken on days 7, 14, 21 and 28 were examined.
Four rats were used per group for each time period and 10 sections
were prepared per animal. The relative areas of α-SMA-positive
cells and numbers of CD4+, CD8+ and
TUNEL+ cells were counted in the HCC area, and the total
area in one microscopic field, at a magnification of ×100. One
microscopic field at ×100 was 11.4 mm2; a total of 57
mm2 of the liver sections were therefore examined per
animal. Cell density was calculated and expressed as cell number
per mm2(7).
Statistical analysis
All statistical analyses were conducted using SPSS
15.0 software. The data were expressed as the means ± standard
deviation (SD). Significant differences were evaluated by unpaired
Student’s t-tests. Correlations among the density of tHSCs,
apoptotic T cells and tumour nodules in the lungs were analyzed
using Spearman’s rank correlation. P<0.05 (two-tailed) was
considered to indicate a statistically significant result.
Results
Tumor model
Orthotopic HCCs were established with no
perioperative complications. All rats (100%) developed tumors
during the observation period (Fig.
1).
Metastatic lung nodules
A previous study demonstrated metastatic lung
nodules at day 30 in an orthotopic MRH hepatoma model (13). This observation was confirmed in our
experiments; 3 out of 6 (50%) animals had significant evidence of
tumor nodules by day 21 (Fig. 2D),
and 6 out of 6 (100%) by day 28 (Fig.
2E). However, tumor nodules were not obvious at days 7 and 14
(Fig. 2B and C). The number of
tumor nodules ranged from 2 to 384 at day 21, (mean, 234.5 nodules
per animal), and from 54 to 579 at day 28, (mean, 318.7 nodules per
animal). There were no tumor nodules in the normal rats, and 0–2
nodules in HCC rats on days 7 and 14 (means, 0.23 and 0.42 nodules
per animal, respectively). No animals had tumor nodules at
extrapulmonary sites. These results demonstrate that the number of
tumor nodules gradually increased with tumor progression (Fig. 2F).
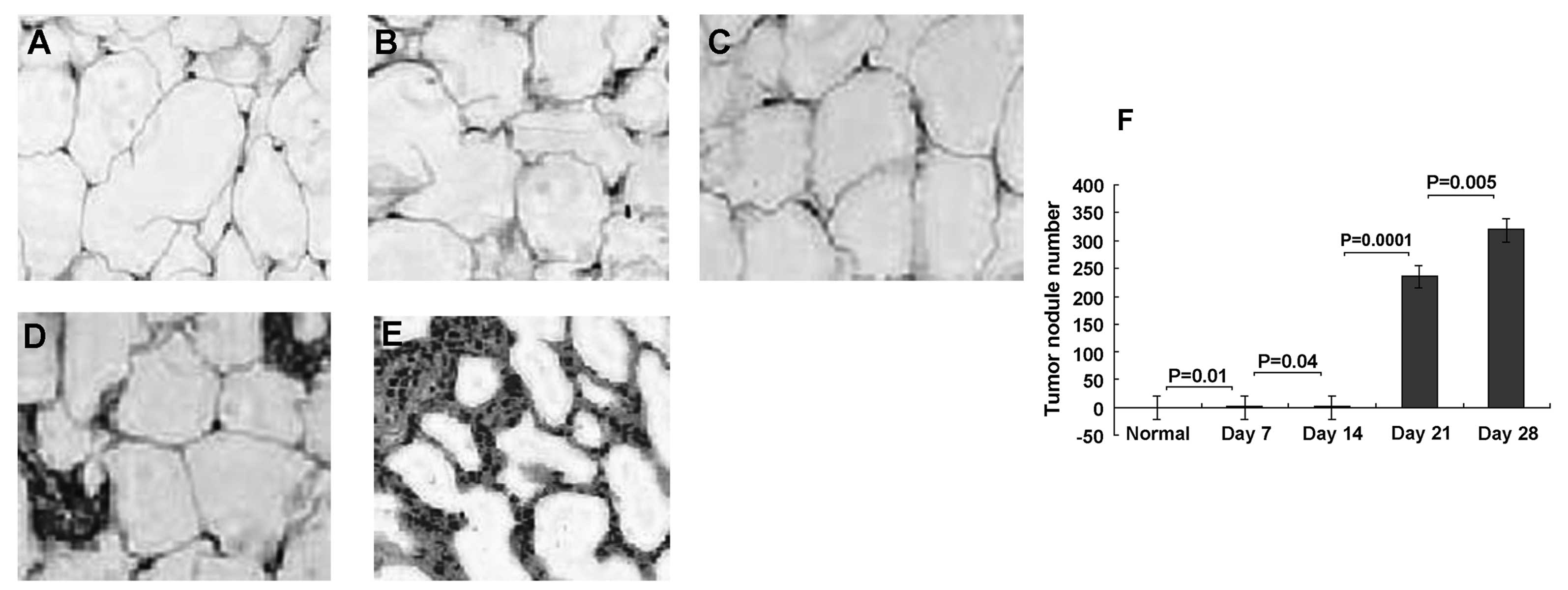 | Figure 2Increased pulmonary tumor nodules in
rat model of orthotopic hepatocellular carcinoma (HCC). Lung
tissues were sectioned and stained with hematoxylin and eosin on
days 0, 7, 14, 21 and 28 days following HCC. Representative
sections from normal and HCC rats are shown. (A) Normal lung
tissue. (B) Lung tissue from HCC rats at days 7, (C) 14, (D) 21,
and (E) 28. (F) Metastatic lung tumor nodules were then quantified
on days 7, 14, 21 and 28. The number of tumor nodules gradually
increased with tumor progression. No tumor nodules were found in
healthy rats. Results are representative of 2 different experiments
with 4 animals in each time subgroup. Results are presented as the
means ± SD, P<0.05. Original magnification, ×200. |
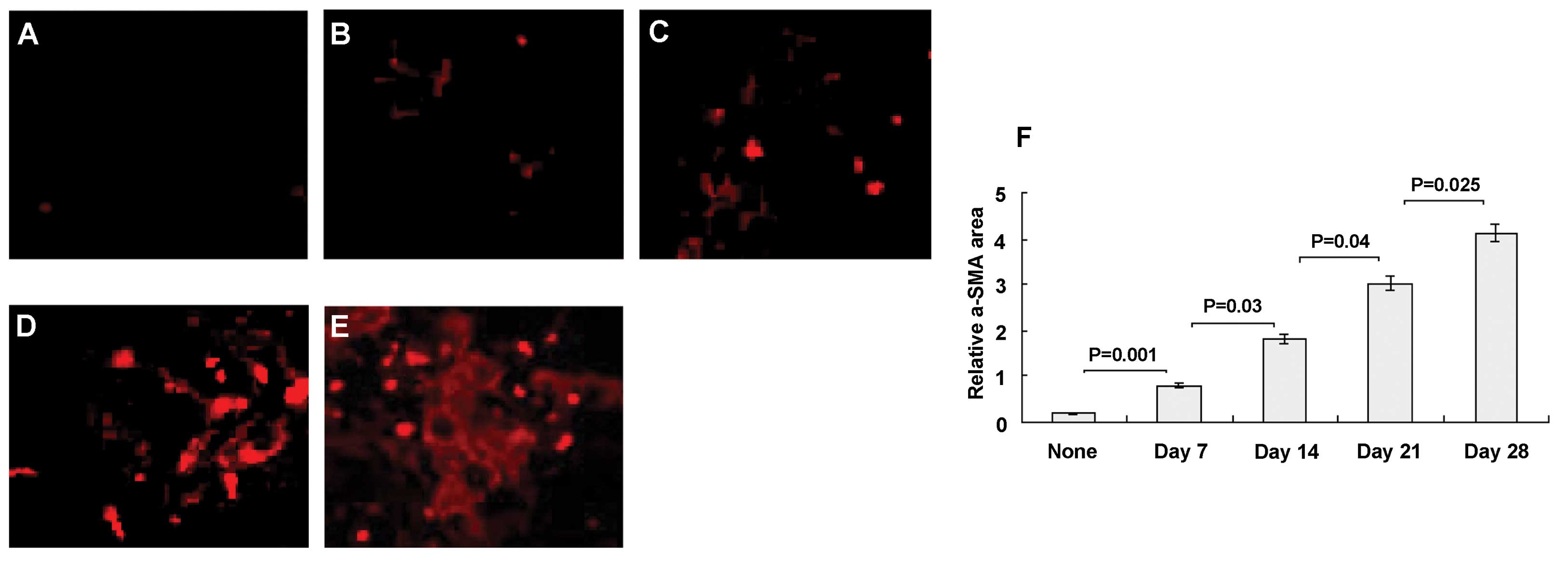
HSC activation in HCC
The expression of α-SMA (a marker of stellate cell
activation) was evaluated in HCC. None of the normal animals showed
abnormal α-SMA staining (Fig. 3A),
except for the normal light staining. In HCC, however, activated
HSCs stained positively with Cy-5 anti-α-SMA and appeared in
situ as red cells (Fig. 3B–E).
Although α-SMA+ cells might include myofibroblasts and
vascular cells as well as activated HSCs, this remains the gold
standard for identification of HSCs (14). The α-SMA signal increased markedly
with time (Fig. 3B–E). The relative
α-SMA area was 0.19±0.12% in normal animals (Fig. 3F), which increased significantly
following HCC progression, to 0.79±0.55 at day 7 (P=0.001),
1.81±1.6 at day 14 (P=0.03), 3.02±1.7 at day 21 (P=0.04) and
4.14±1.9 at day 28 (P=0.025).
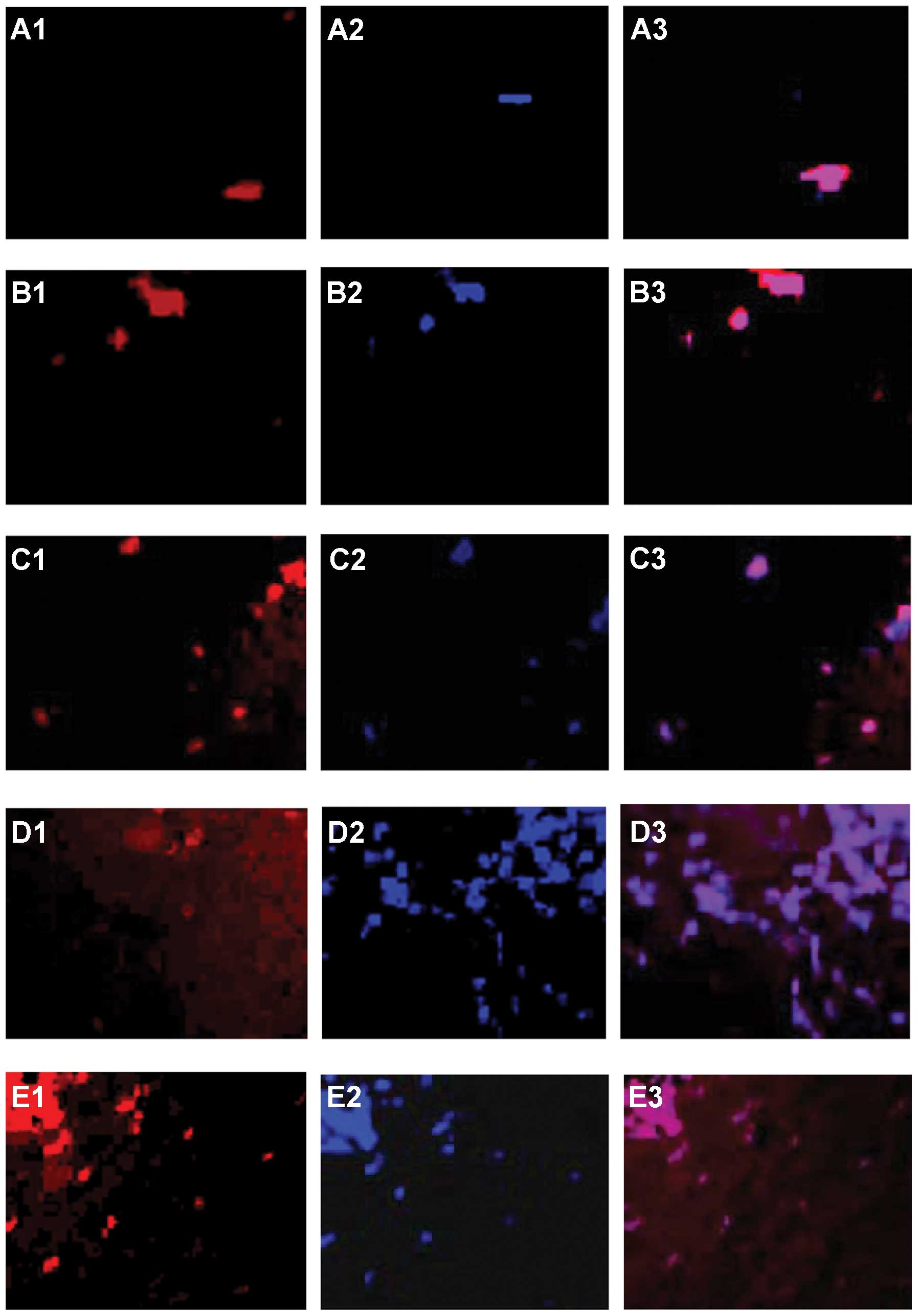
T-cell infiltration in HCC rat liver
Figs. 4–6 show representative stained samples from
normal liver and from HCC livers at 7, 14, 21 and 28 days. Slight
T-cell infiltration occurred at day 7 (Fig. 4B) after tumor implantation, and this
infiltration increased at days 14 (Fig.
4C), 21 (Fig. 4D) and 28
(Fig. 4E), respectively. There were
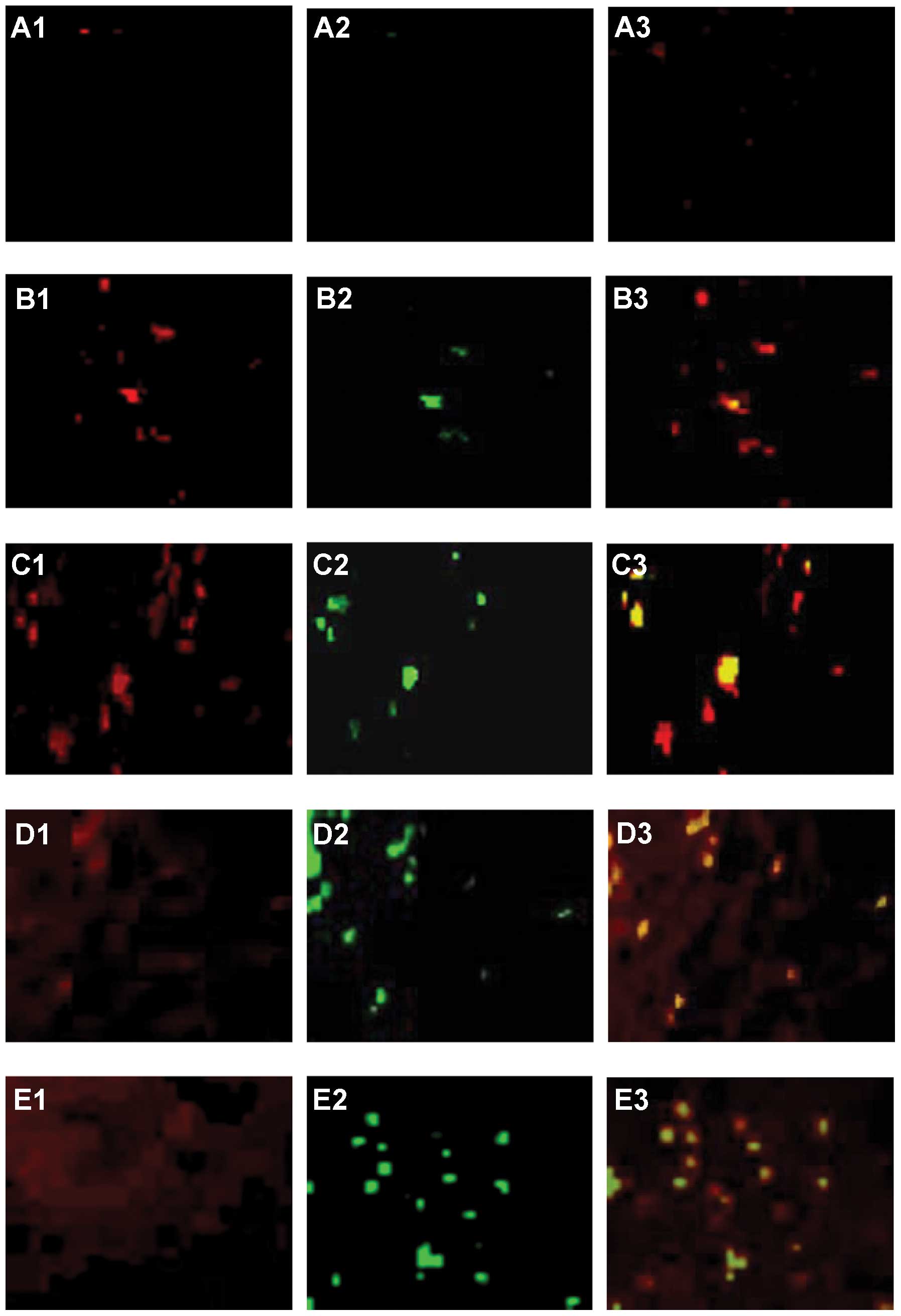
fewer T lymphocytes in normal rat livers (Fig. 4A). Immunofluorescence staining
demonstrated that the number of CD8+ cells per unit
square (Fig. 5A2, B2, C2, D2 and
E2) gradually increased at each sequential check point.
Following tumor progression, CD8+ cells were found very
close to tHSCs. Cy-5-conjugated anti-α-SMA is shown red (Fig. 5A1, B1, C1, D1 and E1) and
APC-conjugated anti-CD8 marker is shown blue in single and merged
stains. Areas stained for both CD8 and α-SMA markers are purple
(Fig. 5A3, B3, C3, D3 and E3).
These results suggest the existence of direct cell to cell
attachment. CD8+ cells were always seen adjacent to
tHSCs. There were few CD8+ cells in normal animals
(Fig. 5F), and the numbers
increased significantly, reaching a plateau within 14 days. The
number of CD8+ cells per unit square (Fig. 5F) peaked at 21 days, and decreased
at 28 days (Fig. 5F). The number of
CD4+ cells per unit square, like CD8+ cells,
also gradually increased at each sequential check point.
CD4+ cells first appeared in situ at day 7
(Fig. 5F). Cy-5-conjugated
anti-α-SMA (Fig. 6A1, B1, C1, D1 and
E1) and FITC-conjugated anti-CD4 markers (Fig. 6A2, B2, C2, D2 and E2) are
illustrated with single (red and green) and merged stains (Fig. 6A3, B3, C3, D3 and E3, yellow-green
cells). CD4+ cells were much less abundant than
CD8+ cells. Like CD8+ cells, however, the CD4
subsets were also mainly found attached to α-SMA-positive tHSCs
(Fig. 6A3, B3, C3, D3 and E3).
There were fewer CD4+ cells in normal rats, and they
increased significantly at 7 days of HCC (Fig. 6F). No marked increase in CD4
staining was seen at day 14, but CD4+ cell numbers were
significantly increased at days 21 and 28 (Fig. 6F).
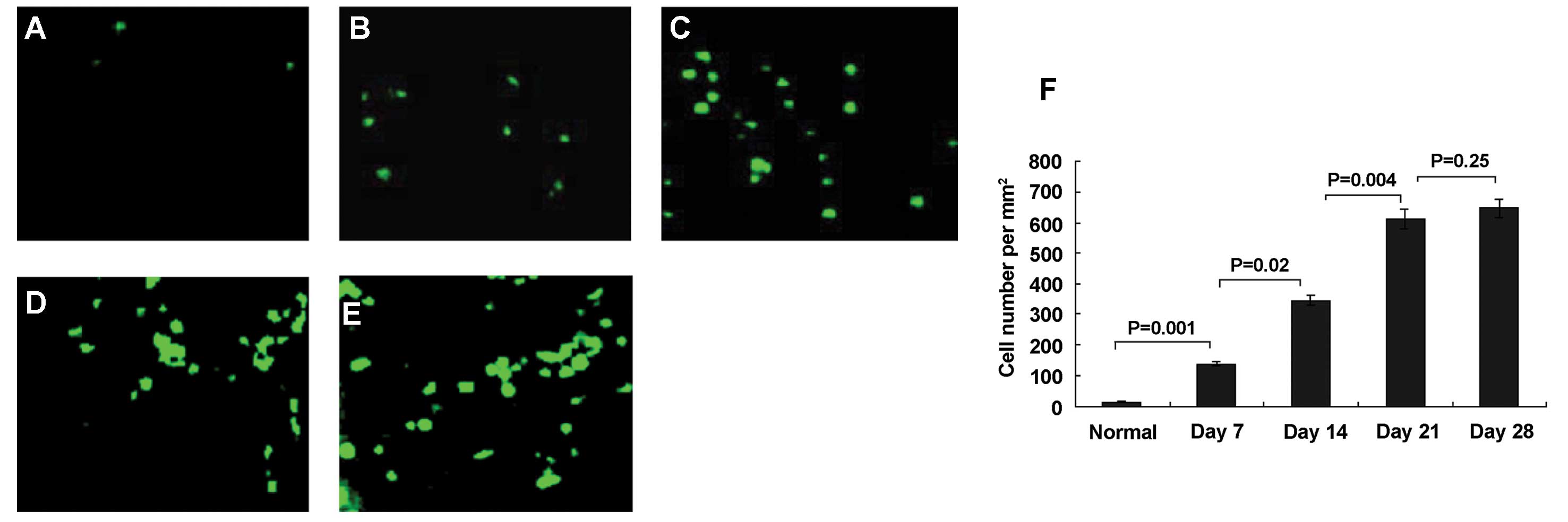 | Figure 4Immunofluorescent staining of normal
and hepatocellular carcinoma (HCC) rat livers by FITC-conjugated
anti-CD3. Fluorescence microscopy of normal and HCC rat livers at
days 7, 14, 21 and 28. (A) Normal liver. (B) HCC livers at days 7,
(C) 14, (D) 21, (E) and 28. There were very few T lymphocytes in
normal livers, and infiltration in HCC livers increased at days 7,
14, 21 and 28, respectively. (F) Measured relative total T
lymphocyte areas in HCC tissue at each time point. Data represent
the means ± SD, P<0.05. Results are representative of 2
different experiments with 4 animals in each time subgroup.
Original magnification, ×200. |
 | Figure 6Double-staining for α-SMA and CD4.
Co-localization of CD4+ cells and intratumoral activated
hepatic stellate cells (tHSC) in situ suggests cell
adhesion. Cells were visualized with confocal laser scanning
microscopy. Cy-5-conjugated α-SMA and FITC-conjugated anti-CD4
markers are illustrated with single (A1, B1, C1, D1 and E1, red;
A2, B2, C2, D2 and E2, green, respectively) and merged stains (A3,
B3, C3, D3 and E3, yellow-green cells). Following tumor
progression, CD4+ cells (green) were only found attached
to tHSCs. There were fewer CD4+ cells in normal rats,
and numbers increased significantly at day 7 in HCC rats. No marked
increase in CD4 staining was seen at day 14, but CD4+
cell numbers were significantly increased at days 21 and 28. (A)
Normal liver. (B) HCC liver at days 7, (C) 14, (D) 21 and (E) 28.
(F) Measured relative CD4+ cells at each time point of
HCC. Data represent the means ± SD, P<0.05. Results are
representative of 2 different experiments with 4 animals in each
time subgroup. Original magnification, ×200. |
 | Figure 5Double staining for α-SMA and CD8.
Co-localization of CD8+ cells and intratumoral activated
hepatic stellate cells (tHSCs) in situ suggests cell
adhesion. Cells were visualized with confocal laser scanning
microscopy. Cy-5-conjugated α-SMA and APC-conjugated anti-CD8
markers are illustrated with single (A1, B1, C1, D1 and E1, red
laser; A2, B2, C2, D2 and E2, blue laser, respectively) and merged
stains (A3, B3, C3, D3 and E3, purple cells). Following tumor
progression, CD8+ cells (blue), were only found attached
to tHSCs. There were fewer CD8+ cells in normal rats
than in HCC rats, and numbers increased significantly, reaching a
plateau within 14 days. CD8+ cells per unit square
peaked at day 21, and decreased at day 28. (A) Normal liver. (B)
HCC liver at days 7, (C) 14, (D) 21 and (E) 28. (F) Measured
relative CD8+ cells at each time point of HCC. Results
are representative of 2 different experiments with 4 animals in
each time subgroup. Data represent the means ± SD, P<0.05.
Original magnification, ×200. |
T-cell apoptosis in HCC
Double staining for CD3 and TUNEL identified some
HCC-infiltrating CD3+ T cells that were also positive
for TUNEL staining (Fig. 7).
Following tumor progression, PE-conjugated anti-CD3- (red; Fig. 7A1, B1, C1, D1 and E1) and
TUNEL-stained cells (green; Fig. 7A2,
B2, C2, D2 and E2) are illustrated as single and merged stains.
Cells stained for both CD3 and TUNEL are yellow-green (Fig. 7B3, C3, D3 and E3). These results
suggest the existence of CD3+ cell apoptosis.
TUNEL+ cells were rare in normal livers (Fig. 7F), but were significantly increased
at days 7 and 14 in HCC livers. The number per unit square of
TUNEL+ cells increased and peaked at day 28 (Fig. 7F). Double staining for α-SMA (red)
and TUNEL (yellow-green) revealed no obvious
α-SMA+/TUNEL+ cells in normal rat livers
(Fig. 8A), but double-stained cells
were clearly found in HCC livers at each time-point (Fig. 8B–E). Direct α-SMA+ HSCs
and TUNEL+ cell adhesion gradually increased on days 7,
14, 21 and 28. TUNEL+ cells were closely associated with
α-SMA+ HSCs at each time point (Fig. 8A3, B3, C3, D3 and E3). Quantitative
analysis demonstrated that the percentage of co-localized
α-SMA+ HSCs and TUNEL+ cells per unit square
gradually increased in the HCC area at each time point, while there
was no appreciable increase in normal rat livers (Fig. 8F).
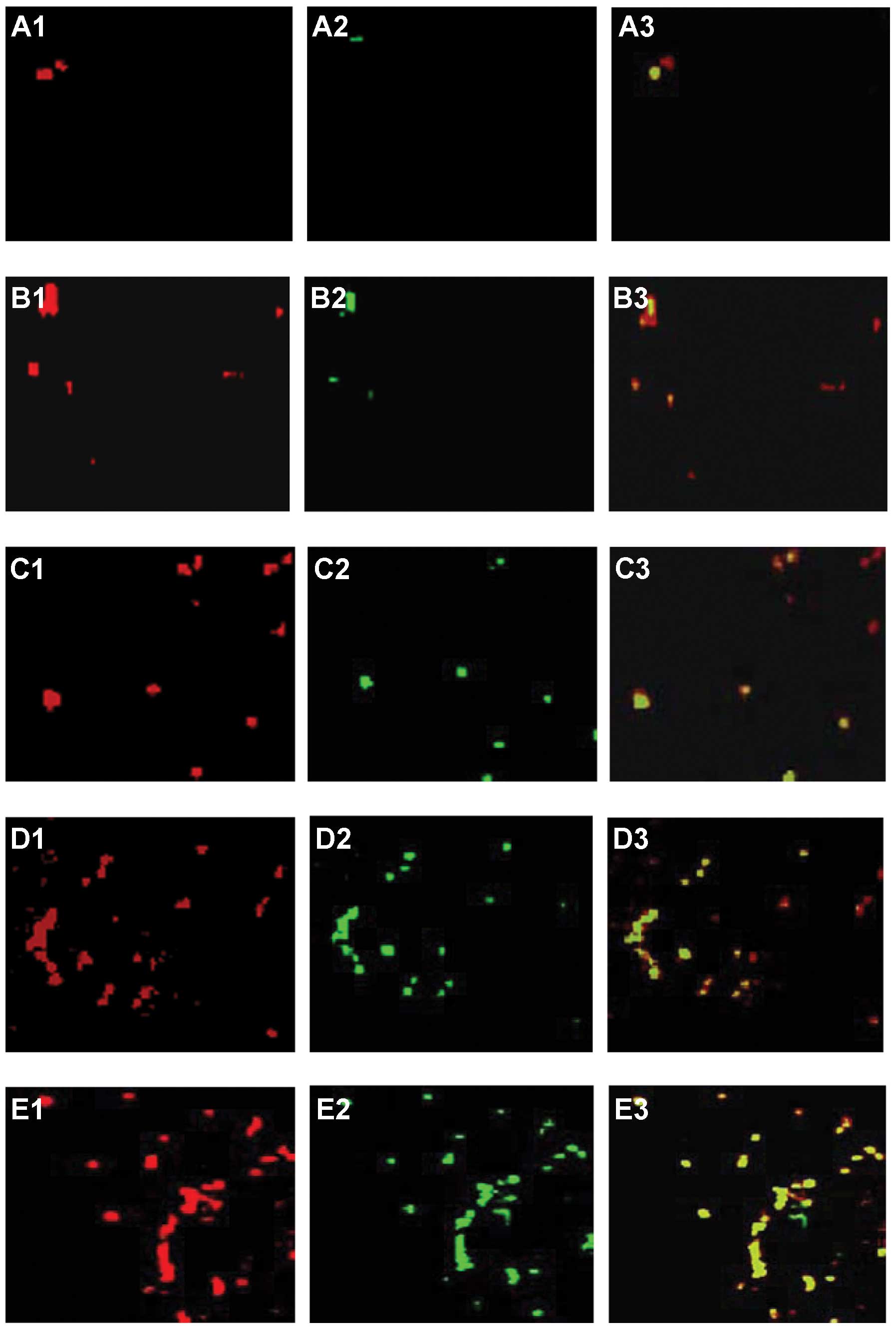 | Figure 7Double staining for T-cell apoptosis
in hepatocellular carcinoma (HCC). Some HCC-infiltrating
CD+ T cells also stained positively for TUNEL. Following
tumor progression, PE-conjugated anti-CD3- (red, A1, B1, C1, D1 and
E1) and TUNEL-stained cells (green, A2, B2, C2, D2 and E2) are
shown as single and merged stains. Overlay staining for both CD3
and TUNEL appears yellow-green (A3, B3, C3, D3 and E3), suggesting
the existence of CD3+ cell apoptosis. (A) Normal liver.
(B) HCC liver at days 7, (C) 14, (D) 21 and (E) 28.
TUNEL+ cells were rare in normal livers, but increased
significantly at days 7 and 14 in HCC livers. The number per unit
square of TUNEL+ cells increased and peaked at day 28.
(F) Measured relative TUNEL+ cells at each time point of
HCC. Data represent the means ± SD, P<0.05. Results are
representative of 2 different experiments with 4 animals in each
time subgroup. Original magnification, ×200. |
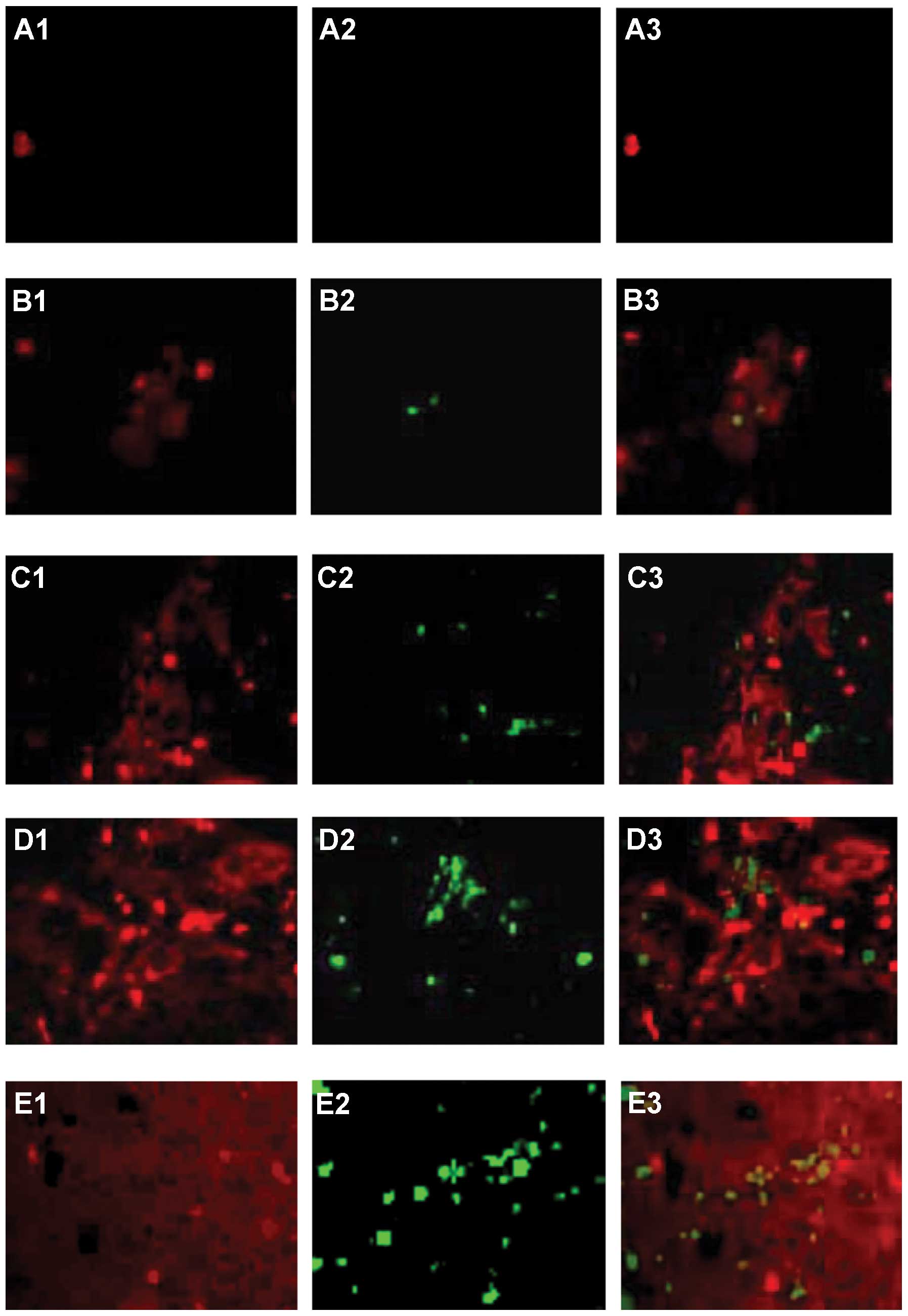 | Figure 8Double staining for TUNEL
(yellow-green) and α-SMA (red). Following tumor progression,
Cy5-conjugated anti-α-SMA+ (red, A1, B1, C1, D1 and E1)
and TUNEL+ cells (green, A2, B2, C2, D2 and E2) are
shown as single and merged stains (A3, B3, C3, D3 and E3). Cells
stained for both α-SMA and TUNEL are yellow-green (A3, B3, C3, D3
and E3), suggesting direct α-SMA+ HSC and
TUNEL+ cell adhesion. α-SMA+ cells were rare
in normal rat livers, but increased in HCC livers.
TUNEL+ cells were sparse in normal rat livers, but
significantly increased in HCC livers at days 7, 14, 21 and 28.
TUNEL+ cells were closely associated with
α-SMA+ HSCs in HCC at each time point, and the
α-SMA+ HSCs and TUNEL+ cell adhesion
gradually increased. (A) Normal liver. (B) HCC liver at days 7, (C)
14, (D) 21 and (E) 28. (F) Co-localized α-SMA+ HSCs and
TUNEL+ cells per unit square in normal and HCC livers at
each time-point. Data represent the means ± SD, P<0.05. Results
are representative of 2 different experiments with 4 animals in
each time subgroup. Original magnification, ×200. |
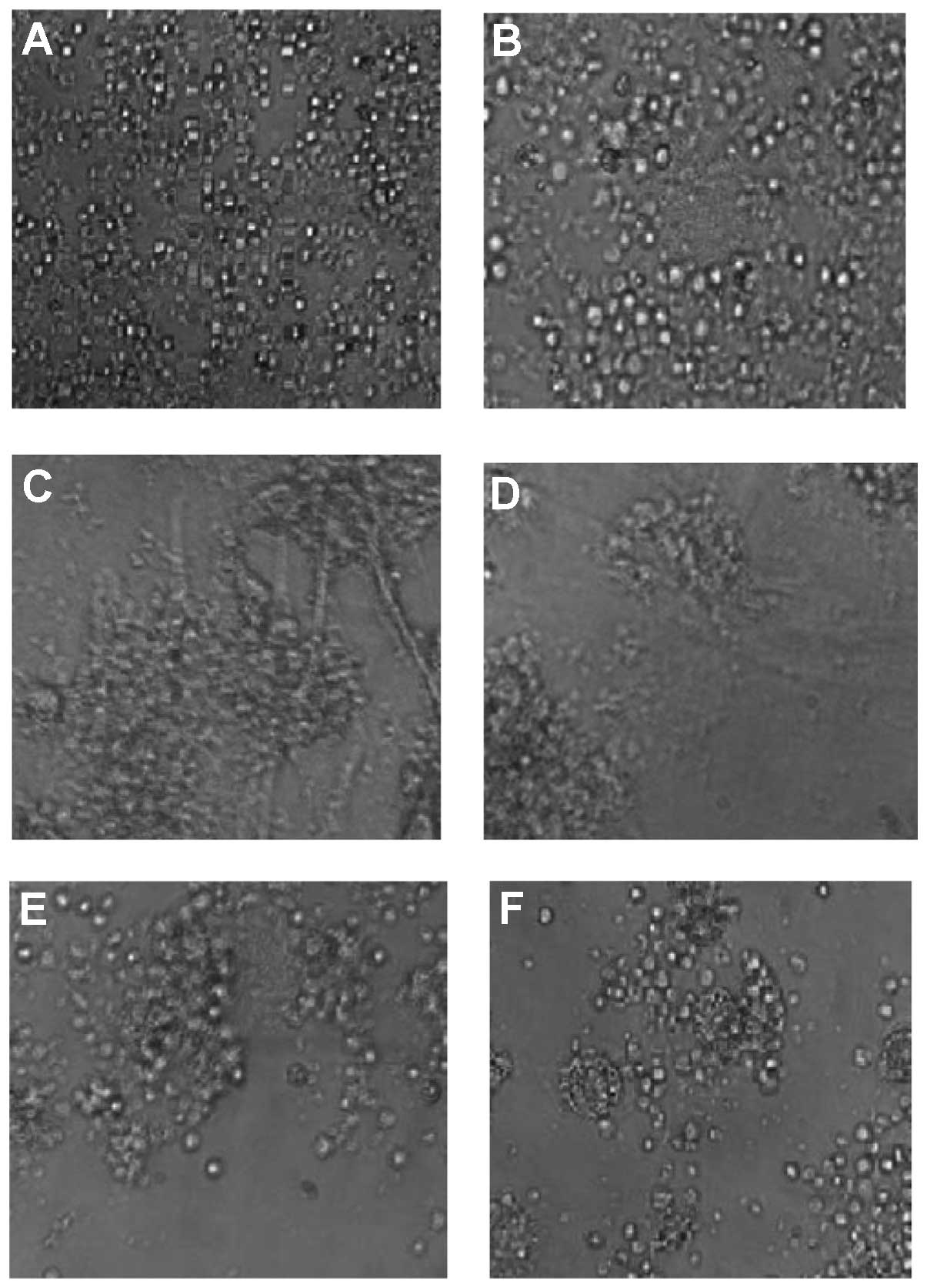
Induction of T-cell apoptosis in
co-culture with HSCs
To confirm the results found in HCC sections, HSCs
were isolated from normal and HCC livers and cultured. HSCs from
the normal livers cultured for 2 days demonstrated quiescent
features (qHSCs), with round or star shapes, abundant lipid
droplets, and a lack of α-SMA expression. HCC HSCs cultured for 2
days (tHSCs) demonstrated activated features (data not shown). We
investigated the adhesion of HSCs and T cells. Light microscopy
clearly demonstrated adhesion of DC-stimulated T cells (aT) cells
to tHSCs in aT/tHSCs co-culture (Fig.
9C). However, T cells were only slightly associated with HSCs
in cultures of qT or aT cells alone (Fig. 9A and B), and in co-cultures of qT
cells/tHSCs (Fig. 9D), aT
cells/qHSCs (Fig. 9E) and qT/qHSCs
(Fig. 9F). We then investigated the
inducing role of tHSCs in T-cell apoptosis using co-cultured
splenic T cells and tHSCs, and double staining for TUNEL and CD3.
Splenic T cells, either non-stimulated (qT) or DC-stimulated (aT),
were co-cultured with qHSCs or tHSCs for 24 h. Culture of qT cells
alone (Fig. 9A) or aT cells alone
(Fig. 9B) for 24 h showed a very
low frequency of apoptosis (Fig. 10A
and B). However, when aT cells were co-cultured with tHSCs
(Fig. 9C), the former often became
round when viewed under a phase-contrast microscope, and stained
positive for TUNEL (Fig. 10C).
Both cell rounding and TUNEL+ staining (Fig. 10D–F) were less frequent in
co-cultures of qT cells/tHSCs (Fig.
9D), aT cells/qHSCs (Fig. 9E)
and qT cells/qHSCs (Fig. 9F). While
the percentage of TUNEL+ cells in total lymphocytes
adhering to HSCs was low in co-cultures of aT cells/qHSCs and qT
cells/tHSCs, it was significantly increased in co-cultures of aT
cells/tHSCs (Fig. 10G).
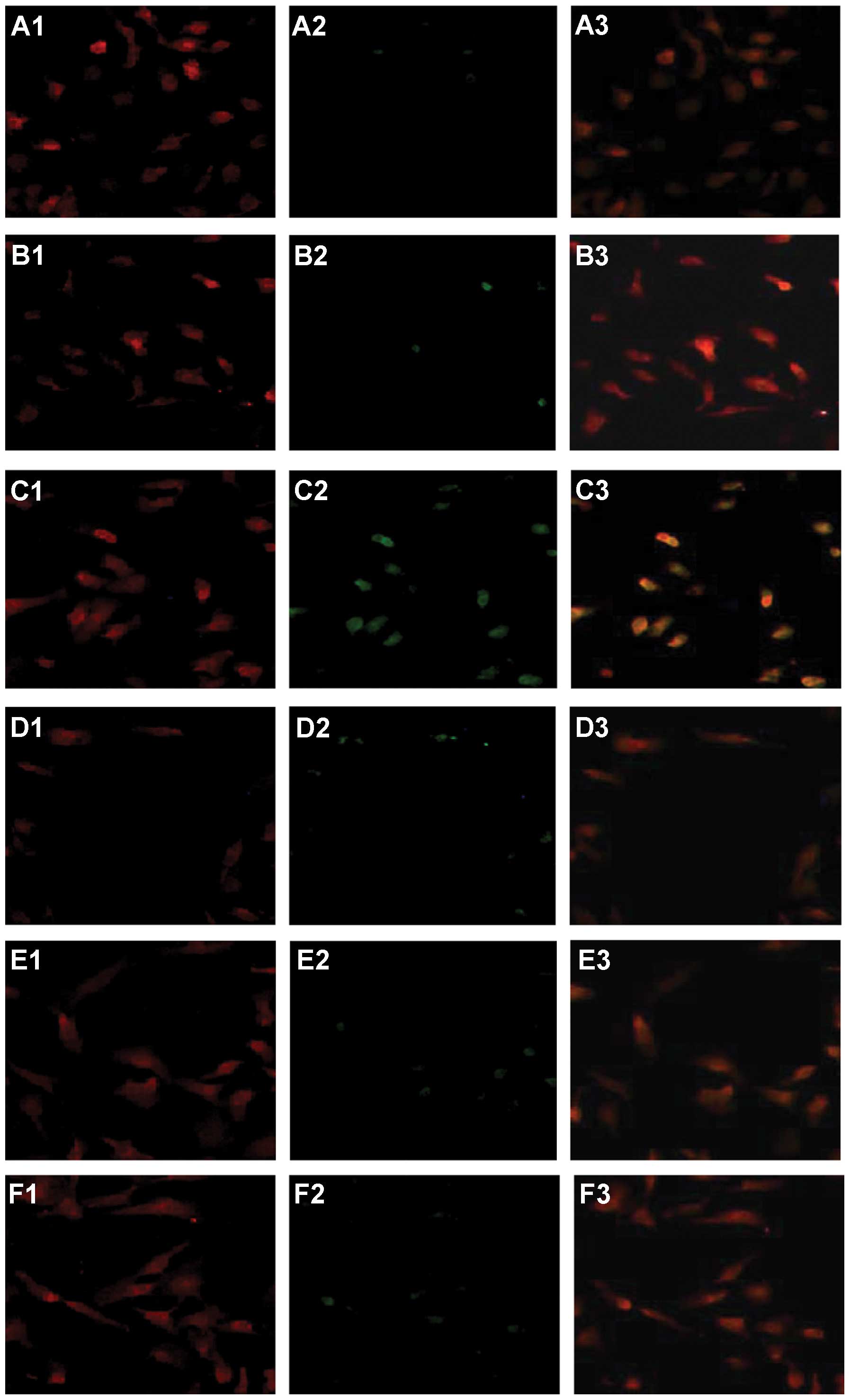 | Figure 10Double staining for TUNEL (green) and
CD3 (red) in quiescent or dendritic cell-stimulated activated T
(aT) lymphocytes co-cultured with quiescent (qHSCs) or intratumoral
activated HSCs (tHSCs). (A) qT cells alone; (B) aT cells alone; (C)
tHSCs/aT cells; (D) tHSCs/qT cells; (E) qHSCs/aT cells; (F)
qHSCs/qT cells. PE-conjugated anti-CD3 (red, A1, B1, C1, D1, E1 and
F1) and TUNEL (green, A2, B2, C2, D2, E2 and F2) are shown as
single and merged stains (A3, B3, C3, D3, E3 and F3). Overlay
staining for both CD3 and TUNEL appears yellow-green (A3, B3, C3,
D3, E3 and F3), suggesting the existence of TUNEL+ CD3
cells. qT cells (A2) or aT cells (B2) cultured alone for 24 h
showed a very low frequency of apoptosis. However, aT cells
co-cultured with tHSCs often stained positive for TUNEL (C2),
whereas TUNEL+ staining was less frequent in co-cultures
of qT cells/tHSCs (D2), aT cells/qHSCs (E2) and qT cells/qHSCs
(F2). (G) Percentage of TUNEL+ T lymphocytes. Results
are representative of three different experiments. Data represent
the means ± SD, the frequency of apoptosis in aT/tHSC co-cultures
compared to all groups, P<0.05. Original magnification,
×200. |
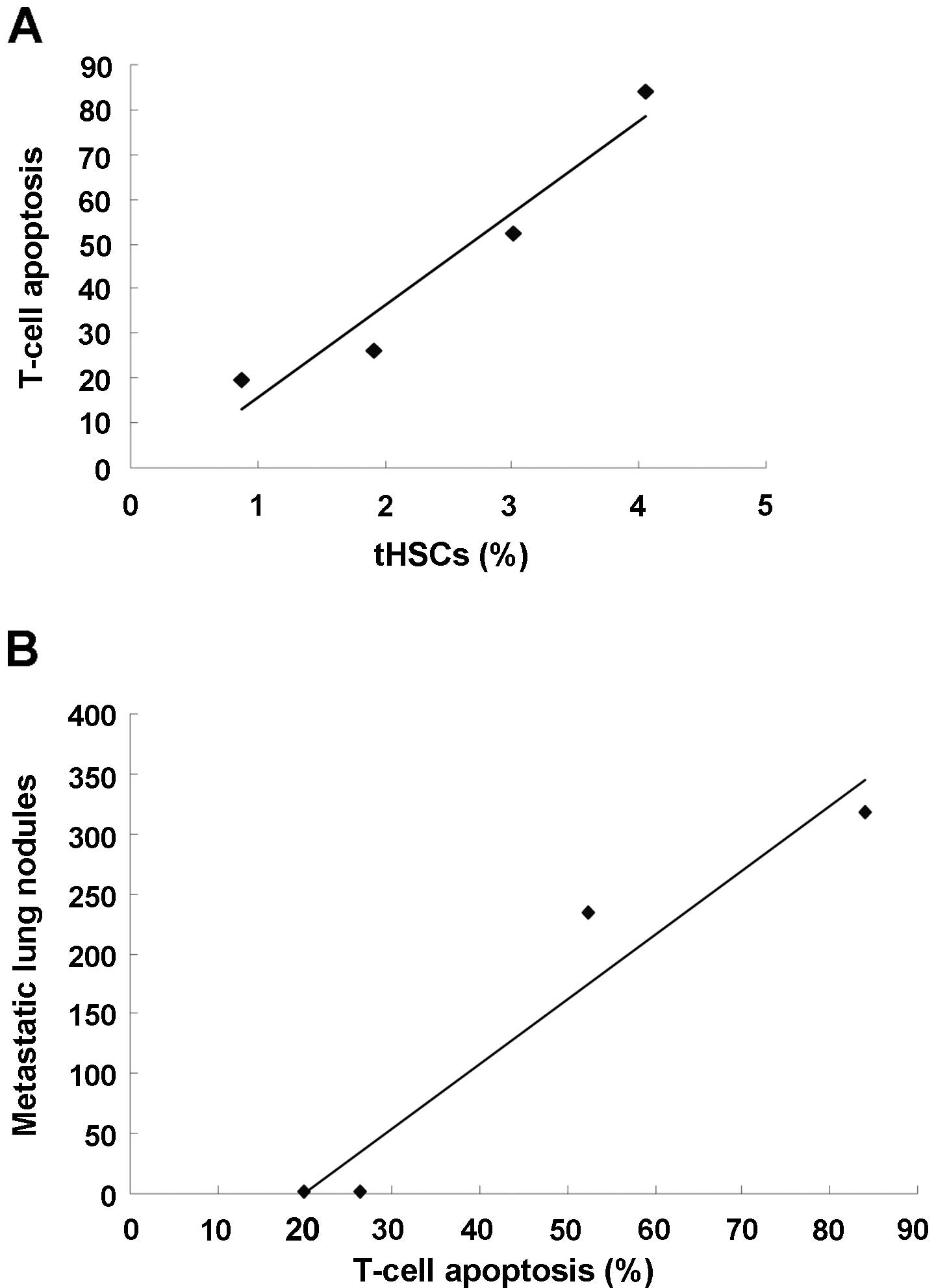
T-cell apoptosis is associated with HCC
metastasis
tHSCs became increasingly prominent in HCC livers
compared to normal livers, following HCC progression at each time
point, accompanied by marked T-cell apoptosis and lung metastasis
(Table I). Correlation analysis
showed that the number of tHSCs was positively correlated with the
percentage of T-cell apoptosis (r=0.861, P<0.05) (Fig. 11A), and the percentage of T-cell
apoptosis was positively correlated with metastasis in the lungs
(r=0.911, P<0.01) (Fig.
11B).
 | Table ItHSCs, T-cell apoptosis and
metastatic lung nodules. |
Table I
tHSCs, T-cell apoptosis and
metastatic lung nodules.
| tHSCs (%) | T-cell apoptosis
(%) | Metastatic lung
nodules |
|---|
| Day 7 (n=4) | 0.87±0.14 | 19.81±5.33 | 0.23±0.92 |
| Day 14 (n=4) | 1.92±0.53 | 26.32±11.38 | 0.42±0.18 |
| Day 21 (n=4) | 3.01±1.28 | 52.31±18.21 | 234.50±92.6 |
| Day 28 (n=4) | 4.05±1.97 | 84.12±29.65 | 318.70±118.5 |
Discussion
Considerable advances have been made in terms of our
understanding of cancer origins. Genetic and cell-biology studies
have indicated that the cooperative activity of parenchymal and
mesenchymal cells is central to carcinogenesis and cancer
progression (20,21). In the liver, hepatocytes represent
the major parenchymal cell type, while the mesenchymal compartment
is composed of various cell types, including Kupffer cells and
HSCs. HSCs are widely accepted as playing a pivotal role in hepatic
tumorigenesis following liver injury (22,23).
HSCs have been previously proposed to exhibit the features of
antigen-presenting cells and to stimulate lymphocyte proliferation
(24). HSCs transdifferentiate into
α-SMA+ myofibroblast-like cells and induce T-cell
apoptosis in liver injury (7).
T-cell apoptosis induced during inflammation contributes to immune
homeostasis in the liver (25,26).
HCC is considered to represent a specific type of liver damage, and
activated HSCs can promote the proliferation and metastasis of HCC
(27). However, the delicate
relationship between the immunological characteristics of tHSCs and
HCC metastasis is far from clear.
In the present study, we investigated T-cell
apoptosis in HCC in an orthotopic rat HCC model, using cubes of
tumor transplanted into rat livers (12). We demonstrated that the role of T
cells in HCC appeared to be modulated through direct attachment to
tHSCs, emphasized by their cell to cell proximity and surface
contact. HCC was confirmed by conventional H&E liver staining,
and direct contacts between T lymphocyte subsets and HSCs were
manifested in HCC livers. We suggest that lymphocytes migrate into
HCC tissue as a result of HCC progression (28,29),
and interact with tHSCs. Confocal imaging revealed significant
increases in intrahepatic CD8+ and CD4+
subsets following tumor progression at each time point. These
results suggest that all lymphocyte subsets infiltrating into HCC
probably interact directly with activated HSCs by adhesion.
T-cell apoptosis was demonstrated by double staining
for TUNEL and CD3. Apoptosis of T cells was more common in HCC
livers compared to normal livers. T-cell apoptosis was also noted
to be spatially associated with tHSCs in HCC, suggesting an
inducing role for the latter cells. However, the involvement of
other cell types, such as macrophages (30), cytotoxic T cells (31) and tumor cells (32), which are reported to possess
apoptosis-inducing functions, could not be excluded. To confirm the
inducing role of tHSCs, we therefore, conducted in vitro
experiments using co-cultures of aT cells and tHSCs. Apoptosis was
increased in these co-cultures, but not in co-cultures of aT cells
with qHSCs, or qT cells with tHSCs, indicating that tHSCs are
responsible for inducing apoptosis of aT cells. Further studies are
planned to investigate the mechanism of T-cell apoptosis with
regard to the involvement of cytokine production or cell-receptor
expression using this culture system. Because the experiment aimed
to demonstrate the requirement for direct contact between
lymphocytes and myofibroblast-like cells, we therefore, examined
the apoptosis-inducing activity of the culture medium obtained from
tHSCs. Supplementation with culture medium from tHSCs did not
significantly increase the percentage of T-cell apoptosis,
confirming the need for direct contact or close proximity with
tHSCs for T-cell apoptosis. These results are in good agreement
with the liver-section results. Activation of T cells has been
reported to enhance their interaction with fibroblasts via a
CD2-dependent adhesion pathway (33), and this interaction serves to
increase the survival of aT cells by inhibiting apoptosis (34,35).
The results of the current investigation support this hypothesis;
while aT cells cultured without HSCs frequently underwent
spontaneous apoptosis, they showed a low frequency of apoptosis in
co-culture with qHSCs. Furthermore, apoptosis of T cells was not
induced in normal livers containing qHSCs, but was induced in HCC
livers containing tHSCs, and in co-culture with tHSCs. The
mechanism of T-cell-apoptosis induction by tHSCs is not known.
Galectin-1 might be involved in this event, because it is produced
by tHSCs but not qHSCs (36), and
induces selective elimination of T cells (37). Some data have demonstrated that aT
cells use CD44 to undergo apoptosis, and dysregulation of this
pathway could lead to increased pathogenesis in a number of
diseases, including hepatitis (38). The upregulated expression of B7-H1
(PD-L1, CD274) in activated HSCs can induce T-cell apoptosis
(9). It has also been reported that
activated HSCs induce transmigration of leukocytes via expression
of adhesion molecules such as ICAM-1 and VCAM-1 (5,39).
Taken together, the present model demonstrated prominent T-cell
infiltration in HCC and indicated that tHSCs play roles in inducing
lymphocyte apoptosis in HCC.
The present study indicated that the number of tHSCs
in HCC gradually increased at days 7, 14, 21 and 28, with a
corresponding progressive increase in the number of T cells.
However, the number of apoptotic T cells also gradually increased,
and lung metastasis ultimately developed. Statistical analysis
showed that tHSCs in HCC were positively correlated with T-cell
apoptosis, and that the percentage of T-cell apoptosis was
positively correlated with the number of lung metastasis nodules.
The present research, thus, demonstrated tHSC-related T-cell
apoptosis, suggesting that tHSC immunosuppression in HCC may
indirectly promote the growth and metastasis of HCC.
In conclusion, the findings of the present study
suggest that T lymphocytes extravasate and accumulate in HCC
tissues during tumor progression. They attach directly to tHSCs and
are induced to undergo apoptosis through interactions with tHSCs,
contributing to lung metastasis of HCC. The present study provides
the first evidence for an immunological function for tHSCs in HCC.
However, further studies are needed to confirm these results and to
identify new targets for inhibiting metastasis of HCC.
Acknowledgements
The authors wish to thank Zen Haiyin from the
Department of Pathology, Zhongshan Hospital, Fudan University, for
her technical help in preparing histological sections. This
research was funded by the National Key Basic Research Development
Program (973) project of China (2004CB518708) and the National
Natural Science Foundation of China (81000909).
References
|
1
|
Unitt E, Rushbrook SM, Marshall A, et al:
Compromised lymphocytes infiltrate hepatocellular carcinoma: the
role of T-regulatory cells. Hepatology. 41:722–730. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mehal WZ, Azzaroli F and Crispe IN:
Immunology of the healthy liver: old questions and new insights.
Gastroenterology. 120:250–260. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Friedman SL: Molecular regulation of
hepatic fibrosis, an integrated cellular response to tissue injury.
J Biol Chem. 275:2247–2250. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Friedman SL: Liver fibrosis - from bench
to bedside. J Hepatol. 38:S38–S53. 2003. View Article : Google Scholar
|
|
5
|
Knittel T, Dinter C, Kobold D, et al:
Expression and regulation of cell adhesion molecules by hepatic
stellate cells (HSC) of rat liver: involvement of HSC in
recruitment of inflammatory cells during hepatic tissue repair. Am
J Pathol. 154:153–167. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Schwabe RF, Schnabl B, Kweon YO and
Brenner DA: CD40 activates NF-κB and c-Jun N-terminal kinase and
enhances chemokine secretion on activated human hepatic stellate
cells. J Immunol. 166:6812–6819. 2001.
|
|
7
|
Kobayashi S, Seki S, Kawada N, et al:
Apoptosis of T cells in the hepatic fibrotic tissue of the rat: a
possible inducing role of hepatic myofibroblast-like cells. Cell
Tissue Res. 311:353–364. 2003.PubMed/NCBI
|
|
8
|
Muhanna N, Horani A, Doron S and Safadi R:
Lymphocyte-hepatic stellate cell proximity suggests a direct
interaction. Clin Exp Immunol. 148:338–347. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yu MC, Chen CH, Liang X, et al: Inhibition
of T-cell responses by hepatic stellate cells via B7-H1-mediated
T-cell apoptosis in mice. Hepatology. 40:1312–1321. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen CH, Kuo LM, Chang Y, et al: In vivo
immune modulatory activity of hepatic stellate cells in mice.
Hepatology. 44:1171–1181. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kurogi M, Nakashima O, Miyaaki H, Fujimoto
M and Kojiro M: Clinicopathological study of scirrhous
hepatocellular carcinoma. J Gastroenterol Hepatol. 21:1470–1477.
2006.PubMed/NCBI
|
|
12
|
Yang R, Rescorla FJ, Reilly CR, et al: A
reproducible rat liver cancer model for experimental therapy:
introducing a technique of intrahepatic tumor implantation. J Surg
Res. 52:193–198. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Freise CE, Liu T, Ascher NL and Roberts
JP: Hepatotoxins and liver transplantation decrease pulmonary
metastases in rats with hepatoma. J Surg Res. 64:198–202. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Forbes SJ, Russo FP, Rey V, et al: A
significant proportion of myofibroblasts are of bone marrow origin
in human liver fibrosis. Gastroenterology. 126:955–963. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sham RL, Packman CH, Abboud CN and
Lichtman MA: Signal transduction and the regulation of actin
conformation during myeloid maturation: studies in HL60 cells.
Blood. 77:363–370. 1991.PubMed/NCBI
|
|
16
|
Ogawa T, Tateno C, Asahina K, et al:
Identification of vitamin A-free cells in a stellate cell-enriched
fraction of normal rat liver as myofibroblasts. Histochem Cell
Biol. 127:161–174. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ikeda K, Wakahara T, Wang YQ, Kadoya H,
Kawada N and Kaneda K: In vitro migratory potential of rat
quiescent hepatic stellate cells and its augmentation by cell
activation. Hepatology. 29:1760–1767. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu L, McCaslin D, Starzl TE and Thomson
AW: Bone marrow-derived dendritic cell progenitors (NLDC
145+, MHC class II+, B7-1dim,
B7-2−) induce alloantigen-specific hyporesponsiveness in
murine T lymphocytes. Transplantation. 60:1539–1545.
1995.PubMed/NCBI
|
|
19
|
Lee JI, Paik YH, Lee KS, et al: A
peroxisome-proliferator activated receptor-γ ligand could regulate
the expression of leptin receptor on human hepatic stellate cells.
Histochem Cell Biol. 127:495–502. 2007.
|
|
20
|
Witz IP and Levy-Nissenbaum O: The tumor
microenvironment in the post-PAGET era. Cancer Lett. 242:1–10.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar
|
|
22
|
Pinzani M, Rombouts K and Colagrande S:
Fibrosis in chronic liver diseases: diagnosis and management. J
Hepatol. 42:S22–S36. 2005. View Article : Google Scholar
|
|
23
|
D’Ovidio KL, Trucksess MW, Devries JW and
Bean G: Effects of irradiation on fungi and fumonisin B(1) in corn,
and of microwave-popping on fumonisins in popcorn. Food Addit
Contam. 24:735–743. 2007.PubMed/NCBI
|
|
24
|
Winau F, Hegasy G, Weiskirchen R, et al:
Ito cells are liver-resident antigen-presenting cells for
activating T cell responses. Immunity. 26:117–129. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Milik AM, Buechner-Maxwell VA, Sonstein J,
et al: Lung lymphocyte elimination by apoptosis in the murine
response to intratracheal particulate antigen. J Clin Invest.
99:1082–1091. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Van Parijs L and Abbas AK: Homeostasis and
self-tolerance in the immune system: turning lymphocytes off.
Science. 280:243–248. 1998.PubMed/NCBI
|
|
27
|
Amann T, Bataille F, Spruss T, et al:
Activated hepatic stellate cells promote tumorigenicity of
hepatocellular carcinoma. Cancer Sci. 100:646–653. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Parmiani G and Anichini A: T cell
infiltration and prognosis in HCC patients. J Hepatol. 45:178–181.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hirano S, Iwashita Y, Sasaki A, Kai S,
Ohta M and Kitano S: Increased mRNA expression of chemokines in
hepatocellular carcinoma with tumor-infiltrating lymphocytes. J
Gastroenterol Hepatol. 22:690–696. 2007.PubMed/NCBI
|
|
30
|
Saio M, Radoja S, Marino M and Frey AB:
Tumor-infiltrating macrophages induce apoptosis in activated CD8(+)
T cells by a mechanism requiring cell contact and mediated by both
the cell-associated form of TNF and nitric oxide. Immunol.
167:5583–5593. 2001.PubMed/NCBI
|
|
31
|
Smith DJ, McGuire MJ, Tocci MJ and Thiele
DL: IL-1 beta convertase (ICE) does not play a requisite role in
apoptosis induced in T lymphoblasts by Fas-dependent or
Fas-independent CTL effector mechanisms. J Immunol. 158:163–170.
1997.PubMed/NCBI
|
|
32
|
Faisal W, Symonds P, Panjwani S, Heng Y
and Murray JC: Cell-surface associated
p43/endothelial-monocyte-activating-polypeptide-II in
hepatocellular carcinoma cells induces apoptosis in T-lymphocytes.
Asian J Surg. 30:13–22. 2007. View Article : Google Scholar
|
|
33
|
Abraham D, Bou-Gharios G, Tulip G, Sumner
H and Olsen I: Regulation of CD2-mediated heterotypic interactions
of murine T lymphocytes. Cell Immunol. 156:342–356. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Crowston JG, Salmon M, Khaw PT and Akbar
AN: T-lymphocyte-fibroblast interactions. Biochem Soc Trans.
25:529–531. 1997.PubMed/NCBI
|
|
35
|
Yarovinsky TO and Hunninghake GW: Lung
fibroblasts inhibit activation-induced death of T cells through
PGE(2)-dependent mechanisms. Am J Physiol Lung Cell Mol Physiol.
281:L1248–L1256. 2001.PubMed/NCBI
|
|
36
|
Kristensen DB, Kawada N, Imamura K, et al:
Proteome analysis of rat hepatic stellate cells. Hepatology.
32:268–277. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Santucci L, Fiorucci S, Cammilleri F,
Servillo G, Federici B and Morelli A: Galectin-1 exerts
immunomodulatory and protective effects on concanavalin A-induced
hepatitis in mice. Hepatology. 31:399–406. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chen D, McKallip RJ, Zeytun A, et al:
CD44-deficient mice exhibit enhanced hepatitis after concanavalin A
injection: evidence for involvement of CD44 in activation-induced
cell death. J Immunol. 166:5889–5897. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hellerbrand, Wang SC, Tsukamoto H, Brenner
DA and Rippe RA: Expression of intracellular adhesion molecule 1 by
activated hepatic stellate cells. Hepatology. 24:670–676. 1996.
View Article : Google Scholar : PubMed/NCBI
|