Introduction
B7-H1, also known as programmed death ligand-1
(PD-L1) or CD274, is a co-inhibitory member of the B7 family
(1). The B7-H1 protein is a cell
surface glycoprotein which is only expressed on macrophage lineage
of cells in normal tissues (1–2). The
immunoglobulin (Ig)V-like domain of B7-H1 plays an essential role
in the interaction with its receptor (3). The receptor for B7-H1 is programmed
death-1 (PD-1, CD279), which is a co-inhibitory receptor (4). In addition to PD-1, other receptors
for B7-H1 may exist (4). The
ligation of B7-H1 on antigen-presenting cells (APC) with PD-1 on T
cells induces anergy in naive T cells and apoptosis in effector T
cells (1). Thus, it is now well
accepted that B7-H1 is a negative regulator of T cell immunity.
B7-H1 protein is overexpressed in various
malignancies including lung (5),
brain (6), melanoma (3), breast (7–8), and
renal (9) cancer. Furthermore, a
correlation between the expression of B7-H1 on cancer cells and
patient clinic pathological status has been shown in some of these
malignancies (7–9). Ligation of B7-H1 on cancer cells with
PD-1 on tumor-specific T cells has been demonstrated to suppress
T-cell activation and proliferation, and to induce T-cell apoptosis
(10). Tumor cells exploit this
regulatory interaction as a mechanism of immune evasion.
Furthermore, cell surface B7-H1 could serve as a receptor to
protect cancer cells from apoptosis induced by Fas binding or the
protein kinase inhibitor staurosporine (11).
Aside from tumor cells, some immune cells in the
tumor microenvironment also expressed B7-H1, including macrophages
and dendritic cells (DCs) (12).
Cell surface B7-H1 on DCs is involved in the induction and
maintenance of T-cell anergy (12).
Signaling through B7-H1 on DCs is required for the induction of
adaptive CD4+ Foxp3+ regulatory T cells
(CD4+Foxp3+Treg) and the modulation
CD4+Foxp3+Treg cell-mediated immune
inhibition (13). It is significant
that the induced Tregs were able to suppress antitumor immunity
(14,15). Thus, B7-H1 may play an important
role in the immune evasion from the host immune system in cancer
patients.
B7-H1 may be regarded as a tumor associated antigen
as well as an immune-suppressor playing an important role in the
immune escape of tumor cells. Blockade of the B7-H1 signal pathway
may represent novel strategies to inhibit tumor growth and enhance
T-cell tumor immunity in cancer. Previous study showed that
administration of mouse B7-H1 monoclonal antibody inhibited the
growth of B7-H1/P815 in vivo(3). Moreover, blockade of the B7-H1/PD-1
pathway with soluble PD-1-IgV protein was able to enhance CTL
activity and antitumor effects on tumor-bearing mice (16).
Active vaccination against B7-H1 is an approach that
may present advantages over passive mAb therapy by inducing a
polyclonal Ab response, potentially leading to better activation of
Ab-dependent effector functions. However, the efficacy of active
vaccination may be limited by the fact that B7-H1 is a self Ag with
poor immunogenicity. In this study, insertion of foreign T-helper
cell epitope (17) was designed to
bypass B7-H1-specific CD4+ Th tolerance. The approach of
such a Th chimeric vaccine was effective toward self Ags in mice,
such as the proinflammatory cytokine TNF-α (18), B-lymphocyte stimulator (BLyS)
(19), and HER2 (20).
In this study, we coupled the P2 tetanus toxoid
T-helper epitope (QYIKANSKFIGITE) (21,22) to
the N terminal end of B7-H1 IgV-like domain (known as recombinant
human B7-H1 mutant, rhB7-H1M). The result demonstrated that the
rhB7-H1M protein vaccination was highly effective in both
preventive and therapeutic animal models.
Materials and methods
Construction, expression and purification
of rhB7-H1M proteins
The rhB7-H1M gene fragment was commercially
synthesized according to the sequence of human B7-H1 in the NCBI
databank (gi:20070268) (Bioasia Company, China). The DNA fragment
was then cloned into plasmid pQE-30 (Qiagen, Germany). rhB7-H1M
protein was expressed in E. coli by
isopropyl-1-thio-β-galactopyranoside (IPTG) induction. The isolated
inclusion bodies were resuspended in solubilizing buffer B with 6
mol/l urea. The supernatant was applied to the Ni2+
chelate affinity column. Bound proteins were eluted with 250 mmol/l
imidazole in buffer B and refolded by dialyzing against buffers
with decreasing denaturant in 20 mmol/l Tris-HCl (pH 9.0): first 4
mol/l urea, then 2 mol/l urea and 1 mmol/l EDTA, and finally PBS
(pH 7.4). The protein was characterized by SDS-PAGE, western
blotting, size exclusion chromatography high performance liquid
chromatography (SEC-HPLC).
SEC-HPLC analysis
SEC-HPLC analysis was performed on a Beckman’s HPLC
system (Beckman Coulter, USA). The protein sample in PBS was
injected onto a 7.5×300 mm G2000SW column (Tosoh, Japan). Peaks
were detected by monitoring at a wavelength of 280 nm. The purity
of the rhB7-H1M protein was calculated as a percentage of the total
peak area detected.
Mice, cell lines, and tumor
challenges
Healthy female BALB/c mice (6–8 weeks old, purchased
from the National Rodent Laboratory Animal Resource, Shanghai,
China) were cared for under institutional animal care protocols in
the Experimental Animal Center of the Fourth Military Medical
University.
SP2/0 (mouse myeloma cell line) and HT-29 (human
colorectal carcinoma cell line) were maintained in RPMI-1640 medium
(Sigma-Aldrich, USA). HT-29/B7-H1-expressing cell line was obtained
by stimulating with 20 ng/ml human interferon-γ (IFN-γ) (R&D
Systems, USA) for 48 h.
For injection, tumor cells were harvested, washed
and suspended at the desired concentration in serum-free medium.
Mice were injected s.c. in the lower back with 100 μl cell
suspension. Tumor growth was measured with calipers and is
presented as the average of the products of two bisecting
diameters.
Immunizations
Proteins for immunization were emulsified 1:1 (v/v)
in IFA (Sigma-Aldrich). Mice were injected s.c. in the neck region
with 40 μg of protein in a total volume of 100 μl. Unless otherwise
stated, vaccinations were performed every other week for a total of
three times.
Flow cytometry
To determine the antigen-binding capacity of
antiserum, HT-29/B7-H1(+) and SP2/0 cells were incubated with
antiserum samples collected from mice vaccinated by rhB7-H1M,
anti-B7-H1 mAb (positive control), or normal serum (vaccinated by
adjuvant, negative control) and analyzed by the flow cytometer
(Becton-Dickinson, USA). For blocking the binding capacity of mouse
serum, tumor cells were incubated with the antiserum samples which
were pretreated with B7-H1/Fc for 30 min.
To identify the changes of
CD4+Foxp+ T cells, lymph cells from mouse
peripheral blood were incubated with anti-CD4 phycoerythrin (PE),
anti-Foxp3-fluorescein isothiocyanate (FITC), or appropriate
isotype controls (eBioscience, USA), and then analyzed by flow
cytometry.
Complement-dependent cytotoxicity (CDC)
assay
The SP2/0 cells were washed and 50 μl of
1×106 cells/ml were plated per well in 96-well plates
(Costar, USA). Cells were incubated with antibody or antiserum
diluted 1:10 with normal guinea pig serum (used as complement). MTT
was added. The plate was incubated for 4 h and DMSO was added. The
OD was measured at 570 nm. Anti-B7-H1mAb (0.1 μg/well) was used as
a positive control. The experiments were repeated at least three
times.
ELISPOT assay
Splenocytes from vaccinated mice were isolated and
restimulated with B7-H1-expressing SP2/0 cells (splenocytes:tumor
cells, 30:1) and cultured at 37°C with 5% CO2 for 24 h.
SP2/0 cells were killed by 60 Gy irradiation and immediately used
as antigen presenting cells specifically for B7-H1. The
restimulated splenocytes were then added to anti-mouse IFN-γ mAb
precoated 96-well plates (2.5×105/well) and further
incubated at 37°C with 7% CO2 for 4 h. The experiment
was performed according to the manufacturer’s instructions (IFN-γ
ELISPOT kit from Invitrogen, USA). The spots were counted using a
dissecting microscope. The spot numbers were the mean of
triplicates in each vaccinated group.
Passive transfer
Sera from BALB/c mice vaccinated with rhB7-H1M were
pooled, and the B7-H1-specific Ab concentration was determined by
ELISA using a standard curve. A total of 100–200 μl of the pooled
sera was injected into the peritoneum of naive BALB/c mice 1 day
before s.c. challenge with 5×106 SP2/0 tumor cells.
Control animals were injected with 500 μg nonspecific mouse
IgG.
Statistical analysis
All data were analyzed by Student’s t-test. Results
are expressed as the means ± standard deviations (SD). A P-value of
<0.05 was considered to indicate a statistically significant
difference.
Results
rhB7-H1M protein is expressed and
purified from E. coli
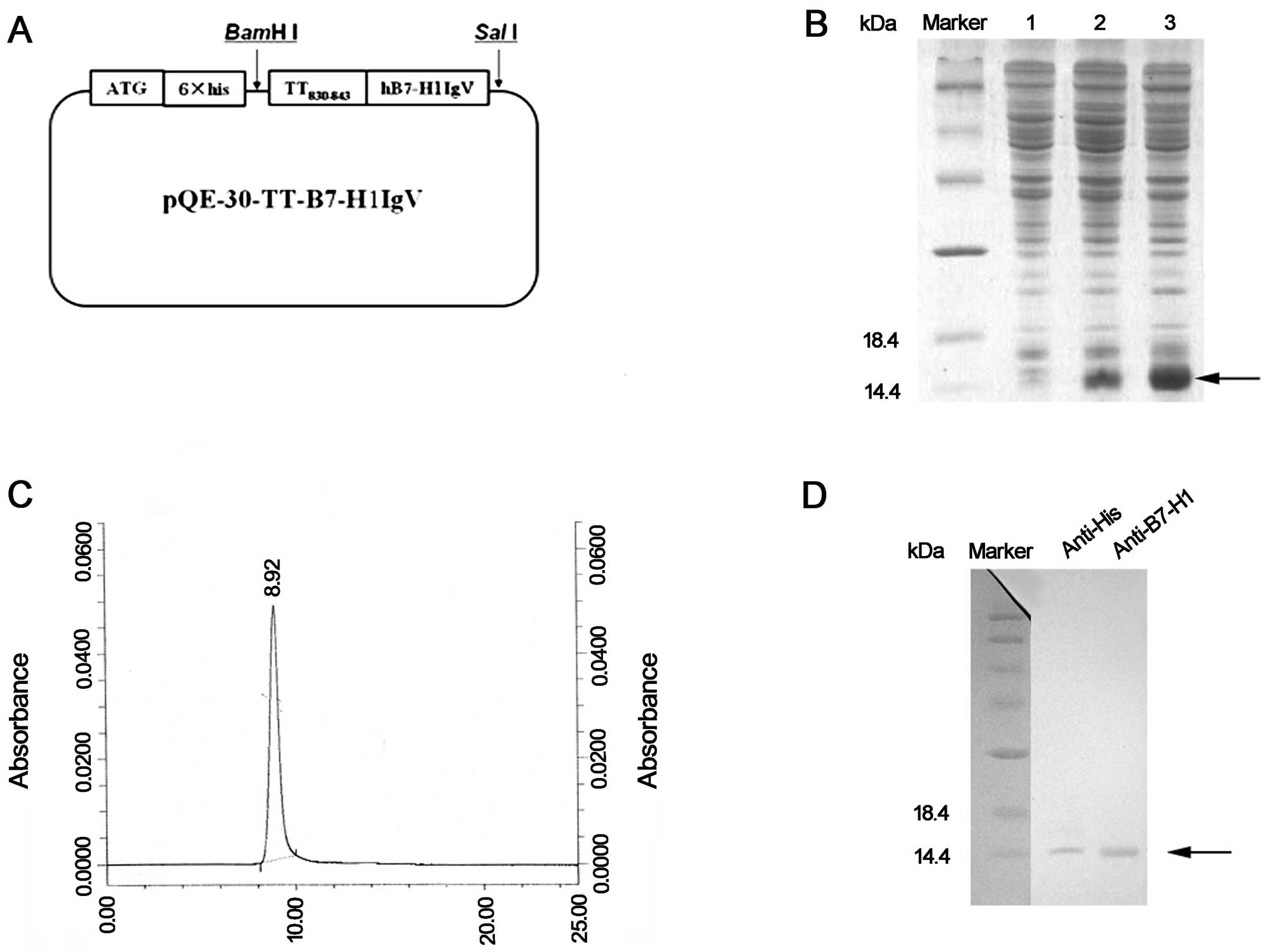
The DNA fragments encoding rhB7-H1M were cloned into
the vector, pQE30 (Fig. 1A). The
recombinant plasmid was transformed into E. coli to express
the fusion protein with an N-terminal six-histidine tag. The
IPTG-induced target proteins were detected by SDS-PAGE. The
observed molecular weight of rhB7-H1M was ~15 kDa, consistent with
the expected size. These fusion proteins, mainly existing in
inclusion bodies (Fig. 1B, lane 3),
were solubilized and purified by Ni2+ chelate affinity
chromatography under denaturing conditions. The protein was then
refolded by dialysis. The purity of the target protein was at least
95% as identified by HPLC (Fig.
1C). The recombinant protein was further analyzed by western
blotting with anti-His and anti-hB7-H1 antibodies (Fig. 1D).
rhB7-H1M protein vaccine mediates tumor
protection in mice
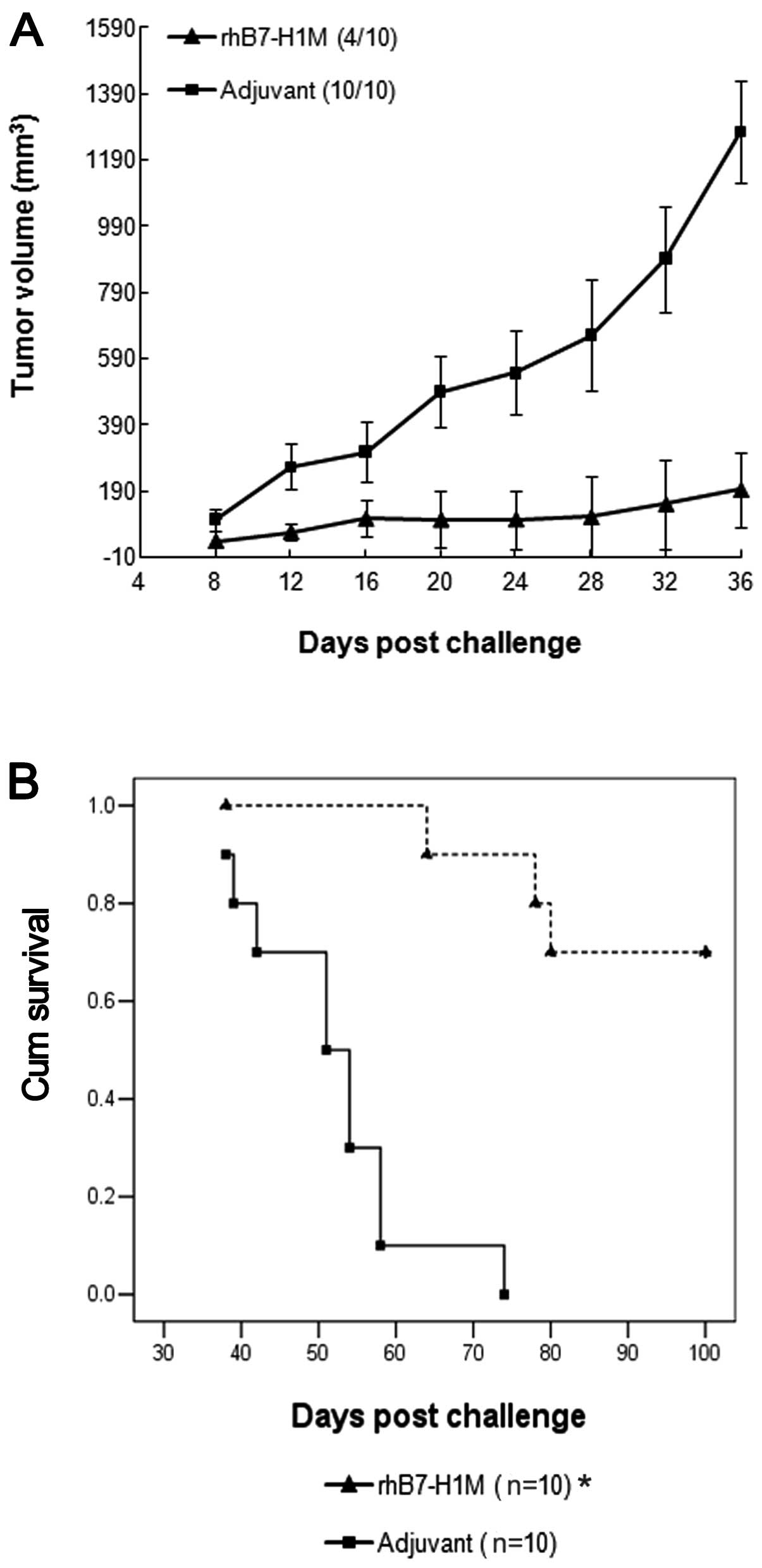
The rhB7-H1M protein vaccine was evaluated for the
ability to elicit a protective immune response against challenge
with a syngeneic B7-H1-expressing SP2/0 tumor cell line. Two groups
of 10 BALB/c mice were vaccinated with either rhB7-H1M protein or
with adjuvant, negative control, respectively. Two weeks later, the
mice were challenged with 5×106 SP2/0 cells and tumor
growth was monitored. The mice vaccinated with adjuvant, negative
control developed large solid tumors within 12–20 days of
subcutaneous administration. Vaccination with the rhB7-H1M protein
significantly inhibited the growth of the tumor cells (Fig. 2A) and only 40% (4/10) of the mice
developed small, slow growing tumors. Thirty-six days after the
challenge, the mice were sacrificed and if a solid tumor was
present, its mass was determined. In the group vaccinated with the
rhB7-H1M protein, the mean mass of tumors from only four mice was
significantly less than the mean mass of the tumors in the
adjuvant, negative control group (1.84 vs. 6.90 g; P<0.05). In
addition, the life span of another two groups of BALB/c mice with
the same treatment as above was observed. The increase in survival
rate in mice vaccinated with the rhB7-H1M protein was also
statistically significant (P<0.05) (Fig. 2B).
rhB7-H1M protein vaccine treats
established tumors
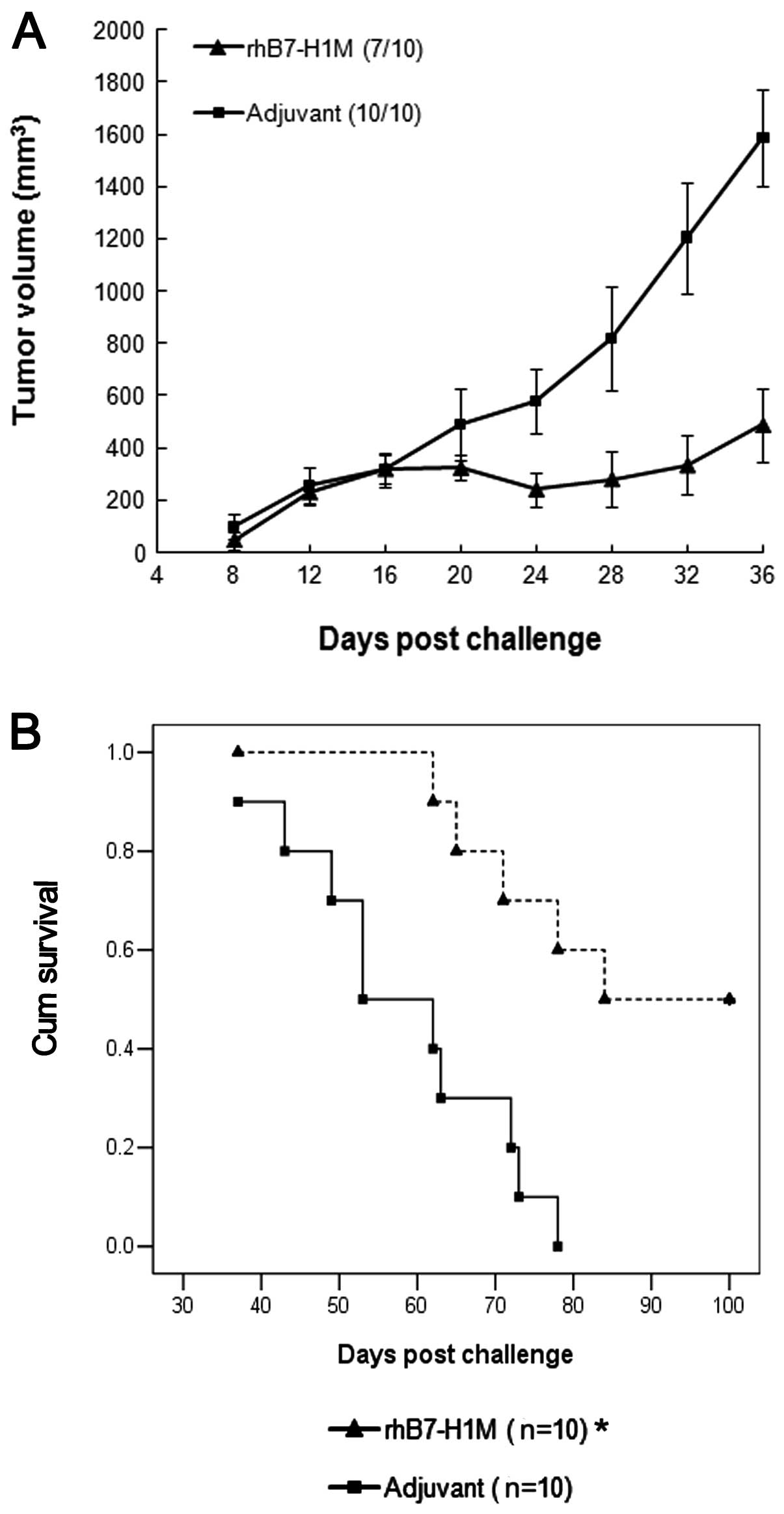
To test the therapeutic efficacy of the vaccine in a
situation that more closely resembles the clinical setting, we
allowed tumors to establish before vaccinating with the rhB7-H1M
protein. At Day 0, BALB/c mice were inoculated with
5×106 SP2/0 cells. At Day 7, the mice were immunized
with rhB7-H1M protein or adjuvant, negative control. Treatment with
the rhB7-H1M protein had a therapeutic effect on tumor growth
(Fig. 3A). The average tumor growth
rate was decreased in some instances. Similarly, tumor incidence
was reduced in the rhB7-H1M group, as 30% (3/10) of rhB7-H1M
vaccinated mice remained tumor-free, vs. 0% (0 of 10) of adjuvant,
negative control mice. Thirty-six days after the challenge, the
mice were sacrificed. The tumor weight of the rhB7-H1M vaccination
group was significantly lower (3.30±0.76 g) than that of the
adjuvant, negative control group (6.98±1.01 g) (P<0.05).
Furthermore, we observed statistically significant differences of
survival time in another two groups of BALB/c mice with the same
treatment as above (Fig. 3B) and
the rhB7-H1M vaccination prolonged the life span of tumor bearing
mice (P<0.05).
Cellular and humoral immunity
To investigate the immunological mechanism
underlying the therapeutic effect of the rhB7-H1M vaccine, we
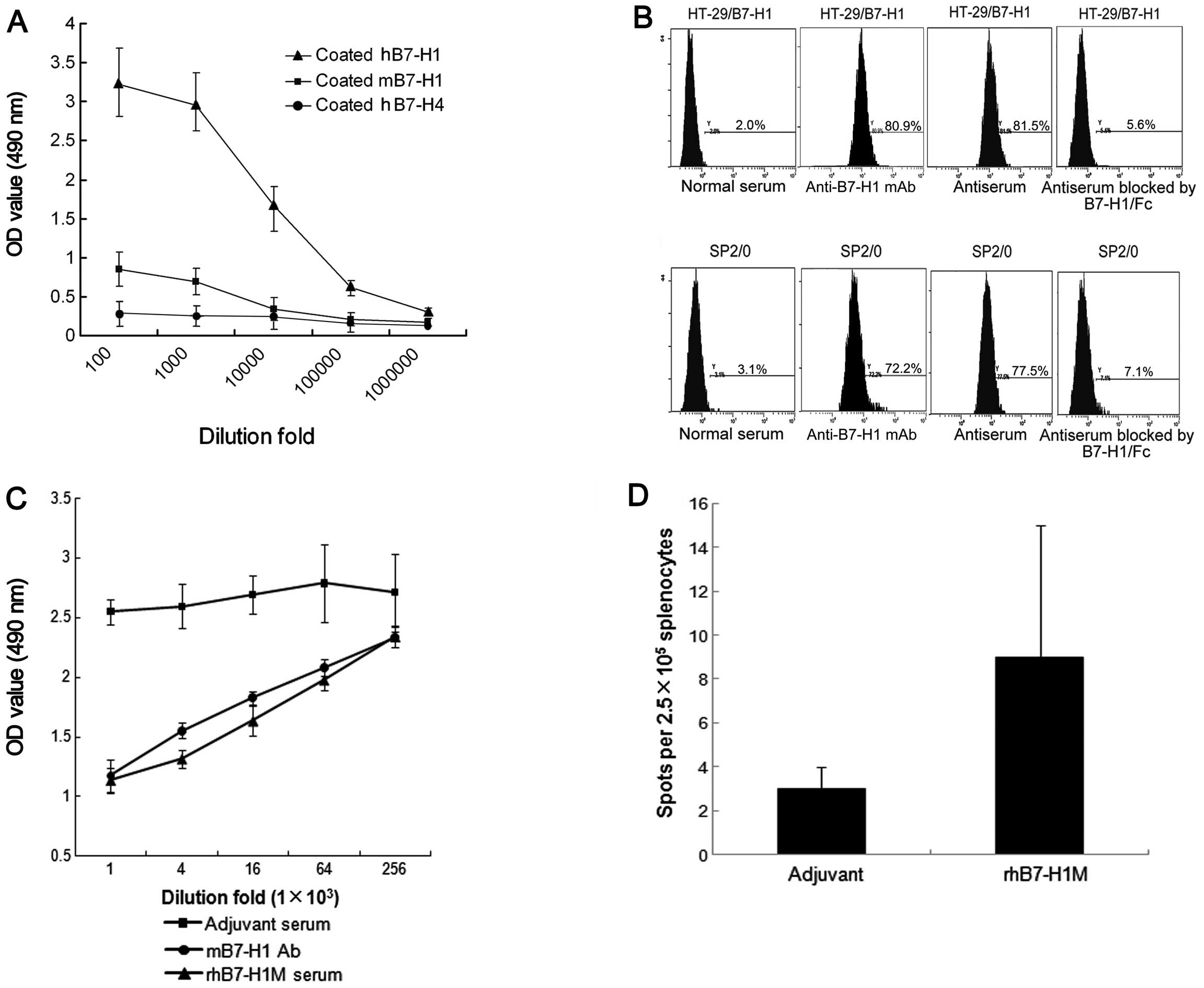
measured the anti-B7-H1 antibody in mouse serum. The vaccine
elicited strong anti-hB7-H1 and anti-mB7-H1 antibody responses
(Fig. 4A). However, there was
almost no cross-binding activity of antiserum with B7-H4, which
belongs to the same family as B7-H1. The results suggested that
antibodies induced by the rhB7-H1M vaccine can interact with human
as well as mouse B7-H1 antigens. The ability of the serum
antibodies to recognize the native cell surface B7-H1 protein was
tested by flow cytometry. The serum antibodies were able to bind to
B7-H1-expressing HT-29 and SP2/0 cells (HT-29/B7-H1, 81.5%; SP2/0,
77.5%) (Fig. 4B). To determine
binding specificity, rhB7-H1/Fc protein and rmB7-H1/Fc protein were
used to compete with the native membrane B7-H1 respectively. The
competitive inhibition rates were 93.1 and 90.8%, respectively
(Fig. 4B). These data indicated
that the rhB7-H1M vaccine resulted in the production of antibodies
to a mixture of native structures.
In addition, antisera from BALB/c mice immunized
with the rhB7-H1M protein were capable of depleting
B7-H1-expressing SP2/0 cells in vitro through CDC (Fig. 4C). The killing rate of SP2/0 cells
reached 53.4±9.4% (1:1,000-fold diluted anti-B7-H1 mAb) and
48.7±8.2% (1:1,000-fold diluted antiserum), respectively. The
killing rate decreased as dilution of anti-B7-H1 mAb and
antiserum.
To measure B7-H1-specific cellular immune response,
the number of IFN-γ-producing B7-H1-specific T-cell precursors was
determined by ELISPOT assay. There was a weak generation of IFN-γ+
spots stimulated in splenocytes from mice immunized with the
rhB7-H1M protein vaccine (Fig. 4D).
Therefore, it was suggested that cellular immunity may not play a
major role in mediating therapeutic effects of the rhB7-H1M protein
vaccine.
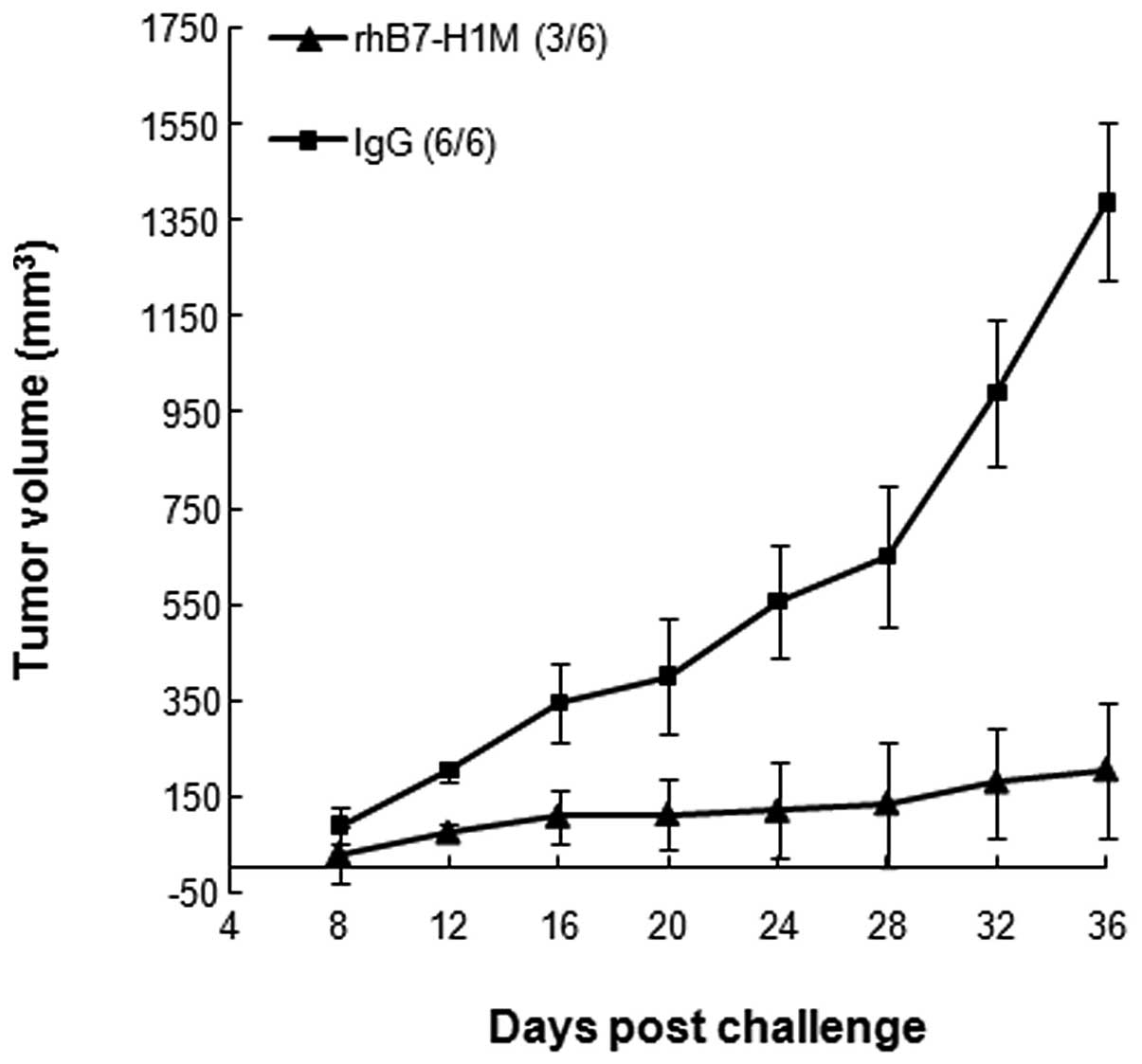
Passive transfer of B7-H1-specific Abs
protects naive mice from tumor challenge
The high Ab response and weak cellular immune
response suggested that Abs may be responsible for the inhibition
of tumor growth. To confirm this, antisera from BALB/c mice
immunized with rhB7-H1M protein were injected into the peritoneum
of BALB/c mice that were subsequently challenged with SP2/0 tumor
cells. Tumor grew at normal rates in mice injected with control
IgG, while tumor growth rate was significantly reduced in mice that
received B7-H1-specific antisera (Fig.
5). Collectively, these results suggest that the rhB7-H1M
vaccine may mediate tumor rejection mainly through Abs induced by
itself in this animal model.
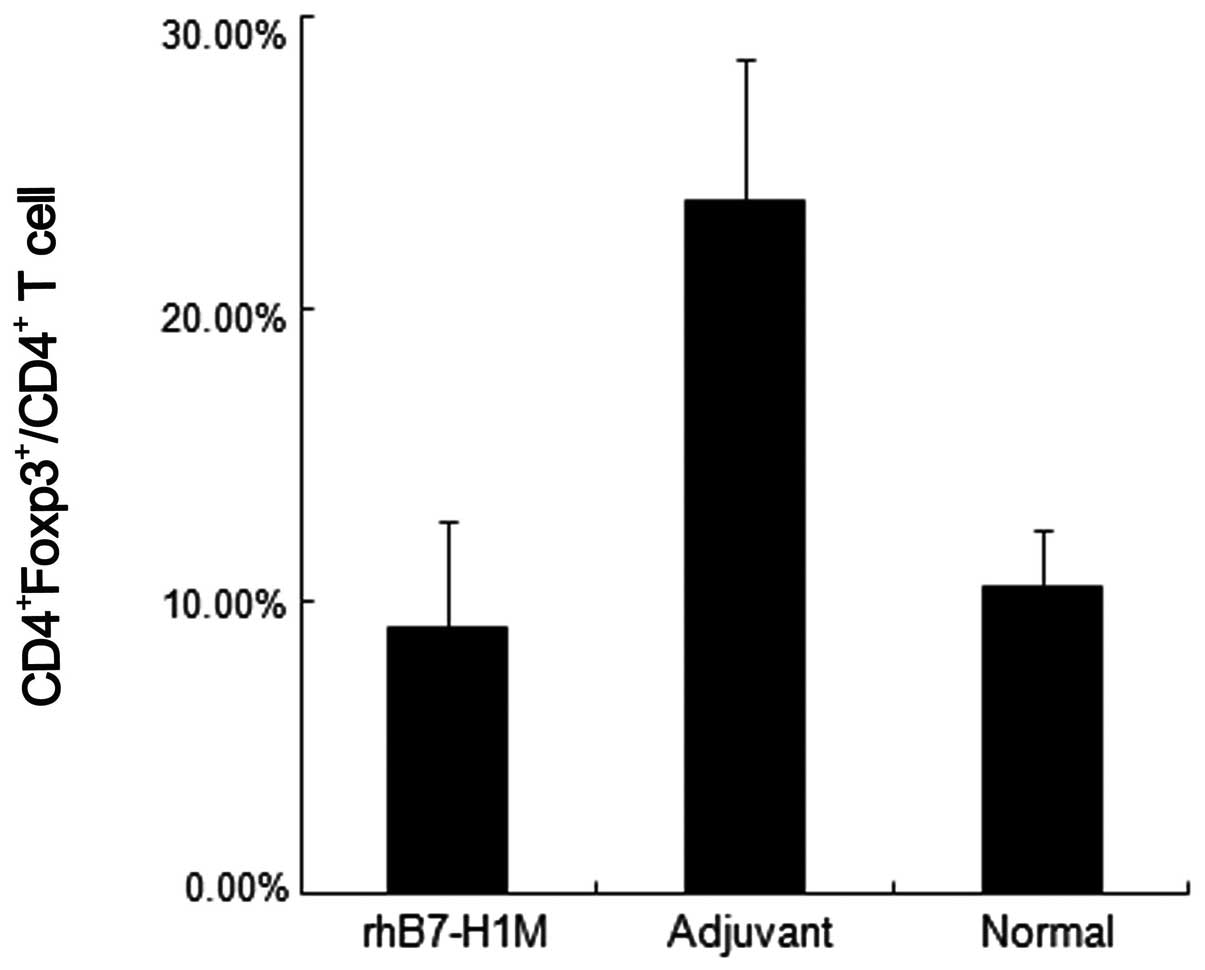
rhB7-H1M vaccine reduces the level of
CD4+Foxp3+Treg cells in tumor-bearing
mice
As discussed above, B7-H1 in the tumor
microenvironment is involved in the induction of
CD4+Foxp3+Treg through the B7-H1/PD-1 signal
pathway. To identify whether the rhB7-H1M protein vaccine could
block the induction of CD4+Foxp3+Treg, we
detected the ratio of CD4+Foxp3+Treg cells to
CD4+ T cells in peripheral blood in the therapeutic mice
models. Similarly, the mice in the vaccination group showed a
significant decrease in the percentage of
CD4+Foxp3+Treg cells compared to the
adjuvant, negative control group and the level of
CD4+Foxp3+Treg cells in the vaccination group
were almost equal to the levels in normal mice (Fig. 6).
Discussion
The molecule of the B7 family belongs to the
immunoglobulin (Ig) superfamily. The IgV-like domain of this family
is responsible for the interaction of ligands and receptors
(2,23). The sequence identity of the amino
acids between human and mouse B7-H1 IgV domain is up to 91%. In
this study, the antiserum by rhB7-H1M vaccine can bind to native
mouse and human B7-H1. It is indicated that the rhB7-H1M vaccine
could break species restriction and induce cross-reactive Abs
against B7-H1 IgV domain of mice and humans. Thus, functional
studies by xenogeneic hB7-H1 vaccination are feasible in mice.
In this study, rhB7-H1M protein vaccines elicited
high B7-H1-specific Ab titers and efficiently prevented
transplantable tumor growth, and eradicated pre-established tumors
in mice models. Together with passive transfer of Abs, it is
suggested that Ab-mediated immune responses are critical in
antitumor immunity induced by the rhB7-H1M protein vaccine. The
effective therapeutic role for B7-H1-specific Abs was previously
suggested by Iwai et al(10). In the present study, we observed
that polyclonal anti-B7-H1 Abs could directly deplete
B7-H1-expressing tumor cells through the CDC effect in
vitro. The mechanisms of antibody involvement in the
destruction of tumor cells in vivo require further
investigation.
By contrast, lower cellular immune response appeared
negligible and did not correlate with tumor protection. However,
additional evidence is required to better understand the level of
T-cell-mediated immunity induced by the rhB7-H1M vaccine. For
example, whether depletion of CD8+ T cells or
CD4+ T cells decreased the efficacy of antitumor
immunity was not tested in the current study, but warrants further
investigation.
In addition, inflammatory mediators such as IFN-γ
are implicated in the upregulation of B7-H1 expression in several
tumor cell lines (24). T cells or
natural killer cells in tumor tissue or tumor cells themselves
secrete several cytokines, including IFN-γ. Therefore, one possible
scenario is that TILs and tumor cells secrete IFN-γ in the
beginning, followed by upregulation of B7-H1 in tumor cells;
thereafter, the upregulated B7-H1 induces CD8+ T-cell
apoptosis through interaction with PD-1 (25). Finally, these factors make up a
positive feedback circle in the tumor microenvironment to help
tumor immune escape. In this study, Abs induced by rhB7-H1M
vaccination may block B7-H1/PD-1 interaction in vivo. The
blockade may destroy this circle and rescue T-cell function which
ultimately translates into antitumor immunity (26).
As discussed above, B7-H1 is involved in inducing
CD4+Foxp3+Treg through the B7-H1/PD-1 pathway
(11). CD4+
Foxp3+Treg could induce anergy of tumor specific CTL.
Thus, these negative immunoregulatory checkpoints form another
network of tumor immune escape. Our preliminary results showed that
rhB7-H1M vaccination could decrease the percentage of
CD4+Foxp3+Treg cells in tumor-bearing mice,
although the mechanisms remain unknown. Thus, the rhB7-H1M vaccine
may partly reverse the condition of immune suppression in a tumor
microenvironment and expose tumor cells to the host antitumor
immunity.
In summary, the present study demonstrated the
utility of the rhB7-H1M protein vaccine is potentially an effective
means of cancer therapy. Current studies are underway to evaluate
the rhB7-H1M vaccine in primate models in an attempt to develop
vaccines useful for treating human B7-H1-expressing carcinomas.
Acknowledgements
The authors thank the members of the Biotechnology
Center of the Fourth Military Medical University for their
excellent technical support and we especially thank Dr Qiuyang
Daniel Zhang for the suggestion on the experiment design. This
study was funded by the National Natural Foundation of China,
NSFC30900537.
References
|
1
|
Dong H, Zhu G, Tamada K and Chen L: B7-H1,
a third member of the B7 family, co-stimulates T-cell proliferation
and interleukin-10 secretion. Nat Med. 5:1365–1369. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Dong H, Strome SE, Salomao DR, et al:
Tumor-associated B7-H1 promotes T-cell apoptosis: a potential
mechanism of immune evasion. Nat Med. 8:793–800. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hansen JD, Du Pasquier L, Lefranc MP,
Lopez V, Benmansour A and Boudinot P: The B7 family of
immunoregulatory receptors: a comparative and evolutionary
perspective. Mol Immunol. 46:457–472. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Freeman GJ, Long AJ, Iwai Y, et al:
Engagement of the PD-1 immunoinhibitory receptor by a novel B7
family member leads to negative regulation of lymphocyte
activation. J Exp Med. 192:1027–1034. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Konishi J, Yamazaki K, Azuma M, Kinoshita
I, Dosaka-Akita H and Nishimura M: B7-H1 expression on non-small
cell lung cancer cells and its relationship with tumor-infiltrating
lymphocytes and their PD-1 expression. Clin Cancer Res.
10:5094–5100. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wintterle S, Schreiner B, Mitsdoerffer M,
et al: Expression of the B7-related molecule B7-H1 by glioma cells:
a potential mechanism of immune paralysis. Cancer Res.
63:7462–7467. 2003.PubMed/NCBI
|
|
7
|
Ghebeh H, Mohammed S, Al-Omair A, et al:
The B7-H1 (PD-L1) T lymphocyte-inhibitory molecule is expressed in
breast cancer patients with infiltrating ductal carcinoma:
correlation with important high-risk prognostic factors. Neoplasia.
8:190–198. 2006. View Article : Google Scholar
|
|
8
|
Hasan A, Ghebeh H, Lehe C, Ahmad R and
Dermime S: Therapeutic targeting of B7-H1 in breast cancer. Expert
Opin Ther Targets. 15:1211–1225. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thompson RH, Kuntz SM, Leibovich BC, et
al: Tumor B7-H1 is associated with poor prognosis in renal cell
carcinoma patients with long-term follow-up. Cancer Res.
66:3381–3385. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Iwai Y, Ishida M, Tanaka Y, Okazaki T,
Honjo T and Minato N: Involvement of PD-L1 on tumor cells in the
escape from host immune system and tumor immunotherapy by PD-L1
blockade. Proc Natl Acad Sci USA. 99:12293–12297. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wilcox RA, Feldman AL, Wada DA, et al:
B7-H1 (PD-L1, CD274) suppresses host immunity in T-cell
lymphoproliferative disorders. Blood. 114:2149–2158. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Selenko-Gebauer N, Majdic O, Szekeres A,
et al: B7-H1 (programmed death-1 ligand) on dendritic cells is
involved in the induction and maintenance of T cell anergy. J
Immunol. 170:3637–3644. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fukaya T, Takagi H, Sato Y, et al: Crucial
roles of B7-H1 and B7-DC expressed on mesenteric lymph node
dendritic cells in the generation of antigen-specific
CD4+Foxp3+ regulatory T cells in the
establishment of oral tolerance. Blood. 116:2266–2276. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo X, Tarbell KV, Yang H, et al:
Dendritic cells with TGF-beta1 differentiate naive
CD4+CD25− T cells into islet-protective
Foxp3+ regulatory T cells. Proc Natl Acad Sci USA.
104:2821–2826. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yamazaki S, Bonito AJ, Spisek R, Dhodapkar
M, Inaba K and Steinman RM: Dendritic cells are specialized
accessory cells along with TGF− for the differentiation
of Foxp3+ CD4+ regulatory T cells from
peripheral Foxp3 precursors. Blood. 110:4293–4302. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang C, Wu S, Xue X, et al: Antitumor
immunotherapy by blockade of the PD-1/PD-L1 pathway with
recombinant human PD-1-IgV. Cytotherapy. 10:711–719. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dalum I, Jensen MR, Hindersson P, Elsner
HI and Mouritsen S: Breaking of B cell tolerance toward a highly
conserved self protein. J Immunol. 157:4796–4804. 1996.PubMed/NCBI
|
|
18
|
Dalum I, Butler DM, Jensen MR, et al:
Therapeutic antibodies elicited by immunization against TNF-alpha.
Nat Biotechnol. 17:666–669. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xue X, Feng G, Li M, et al: Amelioration
of experimental autoimmune encephalomyelitis by BLyS autovaccine.
Vaccine. 26:2873–2881. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Renard V, Sonderbye L, Ebbehoj K, et al:
HER-2 DNA and protein vaccines containing potent Th cell epitopes
induce distinct protective and therapeutic antitumor responses in
HER-2 transgenic mice. J Immunol. 171:1588–1595. 2003. View Article : Google Scholar
|
|
21
|
Valmori D, Pessi A, Bianchi E and Corradin
G: Use of human universally antigenic tetanus toxin T cell epitopes
as carriers for human vaccination. J Immunol. 149:717–721.
1992.PubMed/NCBI
|
|
22
|
Yu Z, Healy F, Valmori D, Escobar P,
Corradin G and Mach JP: Peptide-antibody conjugates for tumour
therapy: a MHC-class-II-restricted tetanus toxin peptide coupled to
an anti-Ig light chain antibody can induce cytotoxic lysis of a
human B-cell lymphoma by specific CD4 T cells. Int J Cancer.
56:244–248. 1994. View Article : Google Scholar
|
|
23
|
Suh WK, Wang S, Duncan GS, et al:
Generation and characterization of B7-H4/B7S1/B7x-deficient mice.
Mol Cell Biol. 26:6403–6411. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gallina G, Dolcetti L, Serafini P, et al:
Tumors induce a subset of inflammatory monocytes with
immunosuppressive activity on CD8+ T cells. J Clin
Invest. 116:2777–2790. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zang X and Allison JP: The B7 family and
cancer therapy: costimulation and coinhibition. Clin Cancer Res.
13:5271–5279. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blank C, Kuball J, Voelkl S, et al:
Blockade of PD-L1 (B7-H1) augments human tumor-specific T cell
responses in vitro. Int J Cancer. 119:317–327. 2006. View Article : Google Scholar : PubMed/NCBI
|