Introduction
CD44 is a cell membrane glycoprotein, which mediates
the response of cells to their cellular microenvironment and
regulates growth, survival, differentiation and motility (1–3). The
function of CD44 depends on its ligands. Hyaluronic acid mediates
the tumor-suppressor function of CD44, while growth factors
regulate the growth promotion function of CD44 (4). CD44 is encoded by a single gene
consisting of 20 exons. Exons 1–5 and exons 16–20 are
constitutively spliced, and are included in all of the CD44 mRNA
isoforms. Exons 6–16 (V exons) are differently included or skipped
to generate a large variety of splicing variants (5). The amino terminal domain of the
standard isoform is separated from the plasma membrane by an
extracellular, membrane-proximal stem structure of CD44 protein.
The stem structure can be different due to the alternative splicing
of stem-encoding variant exons (1,6).
Among the various CD44 isoforms, the V6
exon-containing isoforms (CD44V6) have been implicated in
tumorigenesis (6), tumor cell
invasion and metastasis (7,8). It was shown that CD44V4-V7 conferred
metastatic potential to cells of a non-metastatic rat tumor cell
line (7). Immunohistochemistry
analysis demonstrated a much higher expression of CD44V6 in various
types of tumors when compared with that in normal tissues (9–11). Due
to its significantly high expression, CD44V6 antibody-based cancer
therapy was developed (12,13). The CD44V6-containing isoform forms a
complex with the extracellular hepatocyte growth factor (HGF) and
its tyrosine kinase receptor Met (14,15).
Formation of this complex (CD44V6-HGF-Met) activates Met-dependent
Ras signaling (14,16) through the association of ERM
(ezrin-radixin-moesin) (17–19) to
the cytoplasmic tail of CD44. However, the splicing mechanism of
CD44V6 is not yet clear.
Pre-mRNA splicing is essential for gene expression
in higher eukaryotes (20).
Alternative splicing produces diverse proteins from a gene.
Regulation of alternative splicing plays key roles in signal
transduction and development. Deregulation of alternative splicing
causes various types of diseases including cancer (21–24).
Pre-mRNA splicing requires critical sequences on pre-mRNA called
splicing signals, which include the 5′ splice site, the 3′ splice
site, the polypyrimidine tract (PPT) and branch point (25,26).
Pre-mRNA splicing is regulated by cis-acting elements and
trans-acting elements (27–29).
Cis-acting elements are also called splicing enhancers or
inhibitors which are specific RNA sequences located at exons or
introns. Trans-acting elements are proteins which promote
exon inclusion or skipping.
SC35 is an SR (serine-arginine rich) protein that
includes RRMs (RNA recognition motifs) and RS (arginine-serine
rich) domain (30). SR proteins
participate in multiple steps of splicing including U1 snRNP
binding to the 5′ splice site and U2 snRNP binding to the branch
point. The RS domain of SR proteins functions as an activator,
whereas RRMs provide the binding sites for RNA (31–33).
SR proteins also play additional roles in transcription, RNA
stability, mRNA transport and mRNA translation (34). SC35 plays key roles in constitutive
and alternative splicing in higher eukaryotes.
In the present study, we created a stable cell line
which reports V6 exon skipping and inclusion of CD44 pre-mRNA with
green fluorescence protein (GFP) or red fluorescence protein (RFP),
independently. With this cell line, we identified that the V6 exon
and flanking introns contain SC35 responsive elements. Furthermore,
we found that overexpression of SC35 promoted C5-V6-C6 isoform
production of CD44; knockdown of SC35 reduced CD44V6
expression.
Materials and methods
Construction of pFlare-v6 mini-gene
A reporter construct was generated using standard
cloning techniques. The CD44 genomic DNA, which includes the v6
exon and flanking introns (500 bp each), was PCR amplified from the
Human Genomic DNA Library (Promega, Madison, WI, USA). The PCR
product was cloned in MluI and BamHI enzyme sites in
pFlare9A-Dup34 which was provided by Dr D.L. Black from the Howard
Hughes Medical Institute (Worcester, MA, USA) (35).
Cell culture and generation of the stable
cell line
Breast cancer cell line, MCF-7, was obtained from
ATCC and maintained in Dulbecco’s modified Eagle’s medium (DMEM)
supplemented with 10% fetal bovine serum (FBS) at 37°C in a
humidified 5% CO2 atmosphere. Plasmid transfection into
MCF-7 cells was carried out with polyethyleneimide (PEI) according
to the manufacturer’s instruction. PEI (2 μg) was mixed with 1 μg
of the pFlare-v6 plasmid in 100 μl of DMEM and incubated for 10
min. The mixture was applied to cells in 900 μl of DMEM
supplemented with FBS. Four hours later, the medium was replaced.
The transfected cells were selected under 1 mg/ml G418 (Sigma) for
3 weeks. Stably transfected clones were validated by RT-PCR and
fluorescence microscopy.
RT-PCR
Total RNA was extracted from transfected MCF-7 cells
using RiboEx reagent (GeneAll) following the manufacturer’s
protocol. Total RNA (1 μg) was reverse transcribed using oligo(dT)
primer using ImProm-II™ reverse transcriptase (Promega) following
the manufacturer’s protocol. The cDNA (1 μl) was amplified by PCR
using G-Taq polymerase (Cosmo Genetech Co., Ltd., Seoul, Korea).
The primers used are as following: pFlarev6 forward (5′-GGA AGA GTT
GGT GGT GAG G-3′) and pFlarev6 reverse (5′-GGT GCA GAT GAA CTT CAG
G-3′); v6 forward (5′-TCC AGG CAA CTC CTA GTA GT-3′) and v6 reverse
(5′-CAG CTG TCC CTG TTG TCG AA-3′); GAPDH forward (5′-ACC ACA GTC
CAT GCC ATC A-3′) and GAPDH reverse (5′-TCC ACC ACC CTG TTG CTG
TA-3′). For the endogenous CD44, RT-PCR was conducted as previously
described (36). A specific primer,
CD44RT (5′-ATG CAA ACT GCA AGA ATC-3′) was used for reverse
transcription. PCR was carried out with C5v6 forward (5′-ATC CCT
GCT ACC ATC CAG GCA AC-3′) and CD44E7 reverse (5′-TTT GCT CCA CCT
TCT TGA CTC C-3′).
Fluorescence microscopy
Fluorescence analyses were performed using an
Olympus IX71 inverted microscope and a ×20 LCPlanF1 Ph1 objective.
Images were composed using MetaMorph software and saved as JPEG
files.
shRNA treatment
The shRNA lentivirus was generated by
co-transfection of the pLKO.1 plasmid encoding the SC35 mRNA
matching sequence or the non-silencing sequence (Open Biosystems)
and PSPAX2 and PMD2G helper plasmids into 293T cells using
polyethyleneimide (PEI). The medium was replaced after 24 h and
incubated for another 24 h. The supernatants containing the
lentiviruses were harvested with a 0.45-μm filter. MCF-7 cells were
seeded in a 6-well plate one day prior to infection. The
lentiviruses containing the supernatants were added to the cells
supplemented with 8 μg/ml Polybrene. After a 72-h infection, RNAs
were extracted for RT-PCR.
Results
A stable cell line reports V6 exon
splicing of CD44 pre-mRNA
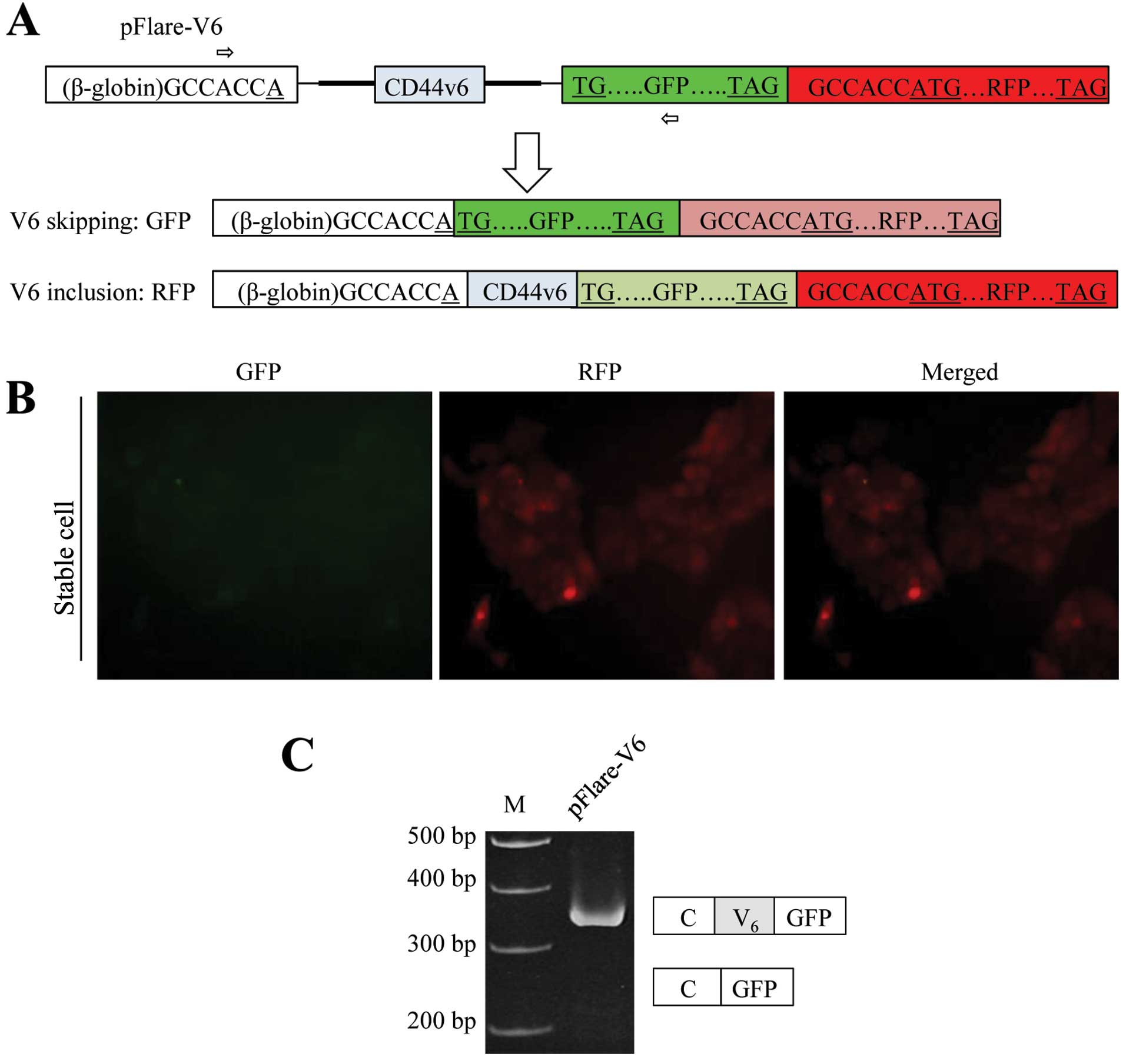
In order to identify the regulatory factors for V6
exon splicing of CD44 pre-mRNA, we constructed a mini-gene with the
pFlare-RFP/GFP reporter system. It was previously shown that the
start codon of GFP is split into a constant β-globin exon and GFP
exon (35). V6 exon and flanking
introns of CD44 were inserted between the constitute exon and GFP
exon (Fig. 1A). GFP is expressed
when the flanking test exon (V6 exon of CD44) is skipped, whereas
another start codon on RFP is out of frame in the pFlare-V6
plasmid. If V6 exon of CD44 is included, the start codon for GFP
expression is abolished. Then another start codon which is located
at RFP will be used for translation of RFP (Fig. 1A). The pFlare-V6 plasmid was
transfected into MCF-7 cells, and the stable cell line
(pFlare-V6-MCF-7) was established by G418 selection for three
weeks. The results in Fig. 1B show
that the red fluorescence protein (RFP) was highly expressed
whereas green fluorescence protein (GFP) was almost not detected in
the stable cell line. Therefore, we hypothesized that the CD44V6
included form should be the dominant isoform, and that the skipped
form is the minimum isoform for the pFlare-V6 stable cell line. In
order to test this possibility, we performed RT
(reverse-transcriptase)-PCR analysis. RNA was extracted from the
stable cell line. Primer sets that base pair with constant exon and
RFP separately were used for RT-PCR analysis (Fig. 1A). The results in Fig. 1C show that the CD44V6 included
isoform was dominantly expressed from the stable cell line; whereas
the CD44V6 skipped isoform was not detectable at a significant
level. Thus, the expression of RFP and GFP indicates V6 exon
splicing. Therefore, the stable cell line expressing pFlare-V6 was
able to be applied for the identification of factors which regulate
V6 exon splicing of CD44 through targeting V6 exon and flanking
introns.
V6 exon and flanking introns of CD44
contain SC35 responsive elements
In order to test the possibility that SC35 regulates
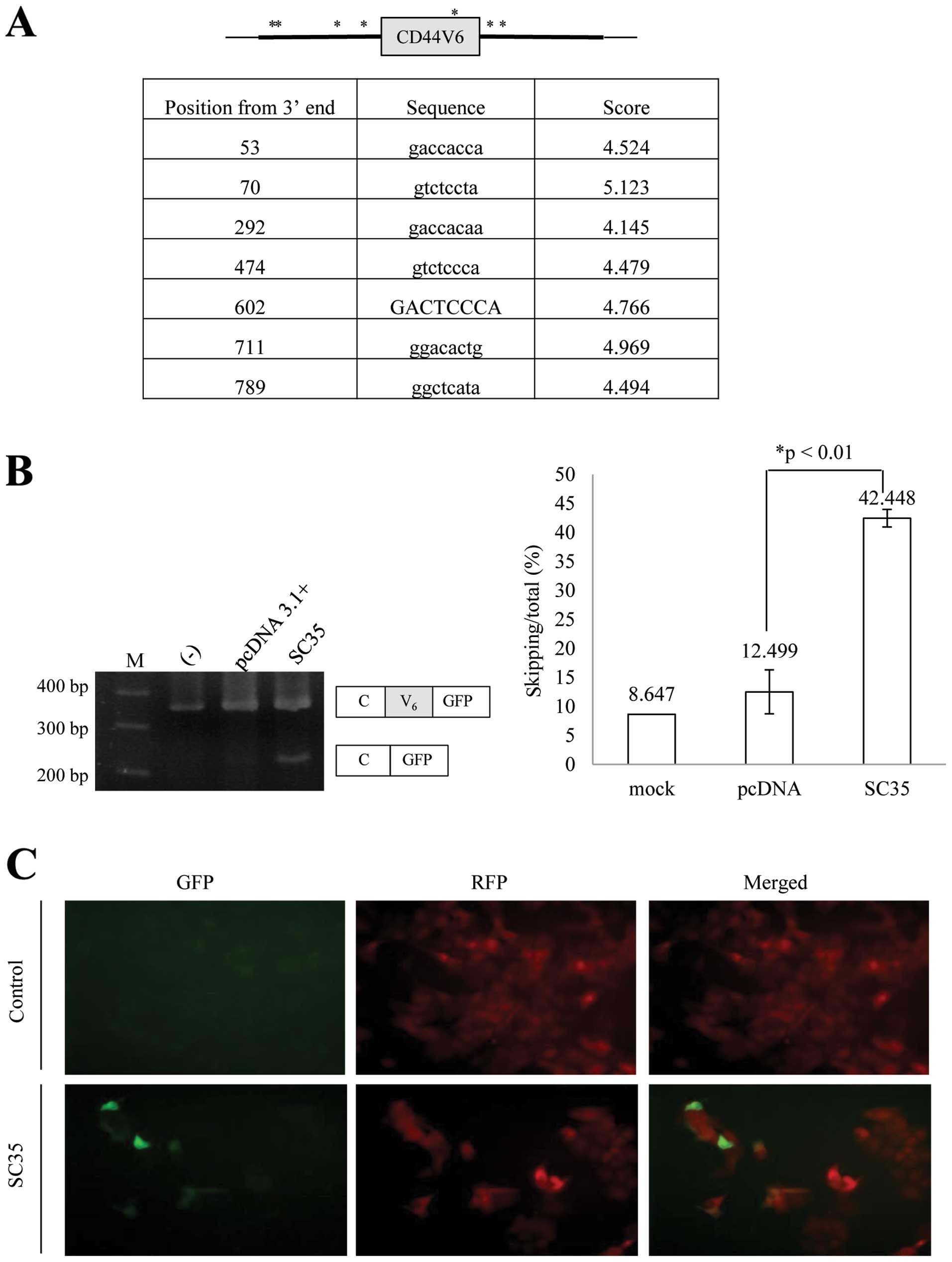
CD44V6 splicing, we applied a bioinformatics approach. Using a
bioinformatics tool (ESE finder) (37), we found that there is one potential
SC35 binding site on the V6 exon, four potential binding sites on
the upstream introns and two potential binding sites on the
downstream introns, which are located at 53, 70, 292, 474, 602, 711
and 789 bp downstream from the 3′ end of the inserted V6 exon and
flanking introns (Fig. 2A). We
hypothesized that SC35 regulates CD44V6 splicing. To test this
possibility, we expressed SC35 in the pFlare-V6-expressing cells.
RNA was extracted; RT-PCR analysis was performed for the splicing
of the V6 exon. As expected, the results in Fig. 2B show that SC35 significantly
promoted-skipping of the V6 exon (by ~34%). By contrast, expression
of the control plasmid (pcDNA3.1+) did not cause a significant
change in V6 exon splicing. This result was consistent with the
fluorescent expression of stable cells as shown in Fig. 2C. Thus, we concluded that CD44V6 and
flanking introns contain SC35 response elements.
SC35 promotes the production of the
endogenous C5-V6-C6 isoform of CD44
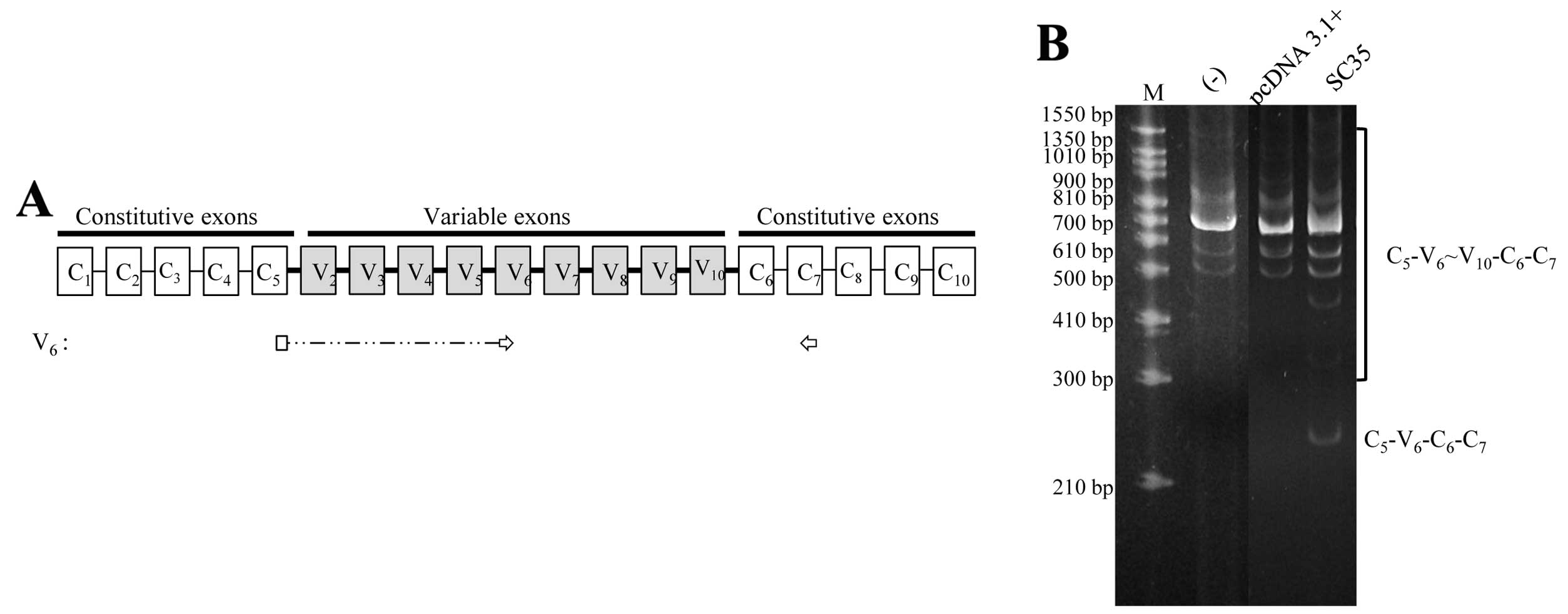
We next aimed to ascertain whether SC35 regulates
the splicing of the endogenous V6 exon. To detect V6-containing
isoforms, we used primer sets that are base paired with junction
C5V6 and C7 separately (Fig. 3A).
The isoforms detected also included V7, V8, V9 or V10 exons. The
MCF-7 cell line is a non-metastatic human breast cancer cell line.
The results in Fig. 3B showed that
the MCF-7 cell line does not express the C5-V6-C6 isoform of CD44,
in which only V6 is included but other various exons are excluded.
Expression of SC35 promotes the production of the C5-V6-C6 isoform.
The production of other V6 exon-containing isoforms, which includes
other various isoforms in addition to V6 (C5-V6~V10-C6), was not
altered upon SC35 expression. Therefore, we concluded that SC35
promotes the production of the C5-V6-C6 isoform.
Knockdown of SC35 reduces endogenous
CD44V6 expression
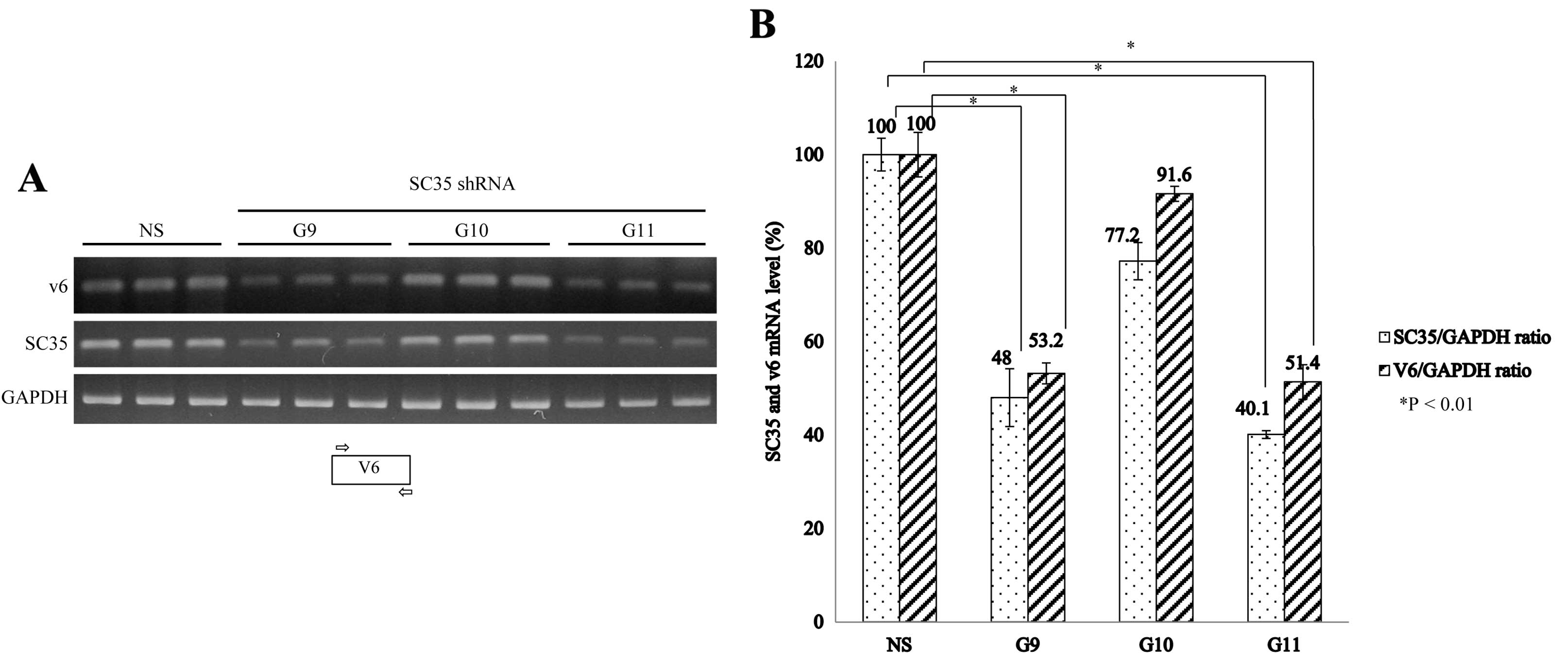
We next aimed to ascertain whether SC35 knockdown
affects V6 exon splicing of CD44. We applied three types of shRNAs
(G9, G10 and G11) to knockdown SC35 mRNA. Fig. 3A shows that G9 and G11 shRNAs
significantly reduced the expression of SC35 as shown with RT-PCR
with SC35-specific primer sets. We found that G10 shRNA which was
designed to target SC35 and non-silencing shRNA did not reduce SC35
expression. To test the effects carefully, we performed triplicate
experiments for every shRNA. RNA was extracted from the cells
treated with shRNA virus and untreated cells; RT-PCR was performed
with primer sets that were base paired with the V6 exon (Fig. 4A). Thus, the PCR products
represented all of the V6-containing isoforms (CD44V6). The RT-PCR
results were normalized to the ratio of SC35/GAPDH and V6/GAPDH
(Fig. 4B). The results in Fig. 4A show that V6 expression was
significantly reduced after infection with the G9 and G11 shRNA
virus. In contrast, V6 expression was not significantly altered
upon G10 and non-silencing shRNA virus infection. Therefore, we
concluded that knockdown of SC35 reduces expression of the CD44V6
isoforms. The quantitation of results is shown in Fig. 4B. G9 and G11 shRNAs reduced the V6
expression by ~48 and ~49% as shown by the average of the V6/GAPDH
ratio. However, G10 shRNA did not induce a significant decrease in
V6 exon expression. V6 expression was reduced after G9 and G11
shRNA infection as shown by the V6/GAPDH ratio, whereas G10 shRNA
infection did not induce the alteration of V6 expression
significantly. Therefore, we concluded that knockdown of SC35
induces a decrease in CD44V6 expression.
Discussion
SR proteins play important roles in constitutive and
alternative splicing (38–40). SC35 has been reported to play an
active role in transcriptional elongation (41) and mRNA stability (42). In the present study, we performed a
systematic RT-PCR analysis to determine the role of SC35 in CD44
pre-mRNA splicing. In the present study, we created a stable cell
line that expressed the V6 exon of CD44 and its flanking introns.
Expression of RFP (red florescence protein) indicated inclusion of
the V6 exon, whereas (green florescence protein) GFP expression
indicated V6 exon skipping. With this stable cell line, we found
that SC35 promotes V6 exon skipping in the mini-gene. These results
indicate that V6 exon and flanking introns of CD44 pre-mRNA contain
SC35 response elements. RT-PCR analysis with endogenous CD44
pre-mRNA demonstrated that SC35 promotes the C5-V6-C6 expression of
the CD44 pre-mRNA. By contrast, knockdown of SC35 with shRNA
reduced expression of the V6-containing isoform. Collectively, our
results indicate that SC35 promotes CD44V6 inclusion.
V6 exon and flanking introns of CD44
pre-mRNA contain response elements for SC35
Our strategy for constructing a pFlare-V6 mini-gene
is that GFP is expressed when the V6 exon of CD44 is skipped. By
contrast, RFP is expressed while CD44V6 is included. Our microscopy
results demonstrated that the stable cell line expressed only RFP.
RT-PCR results showed that the V6 exon of CD44 was dominantly
included in the pFlare-V6-transfected MCF cells. Consistent with
the results, microscopic analysis demonstrated that the stable cell
line expressed only RFP. pFlare plasmid contained the V6 exon and
flanking introns (500 nt each) of CD44 pre-mRNA. The first and last
exon of the mini-gene (pFlare-V6) was not from the CD44 gene; thus,
the mini-gene can be applied for the identification of
trans-acting response elements on V6 exon and flanking
introns. Based on a bioinformatics approach we hypothesized that V6
and flanking introns contain potential SC35-binding sites. As
expected, SC35 regulates V6 exon splicing of CD44 in the pFlare-V6
pre-mRNA. Our results demonstrated that CD44V6 and flanking introns
contain SC35 response elements.
SC35 promotes the production of the
C5-V6-C6 isoform of CD44
The existence of SC35 response elements on CD44V6
and flanking introns raised the possibility that SC35 regulates
endogenous V6 exon splicing of CD44 pre-mRNA. We found that SC35
induced the expression of the C5-V6-C6 isoform of CD44, which was
not significantly expressed in the non-metastatic breast cancer
cell line MCF-7.
Knockdown of SC35 reduces the expression
of the CD44V6 isoform
We found that SC35 knockdown induced a decrease in
CD44V6 expression. Therefore, we conclude that SC35 has a critical
function in the regulation of CD44V6 splicing.
Acknowledgements
The present study was supported by the Mid-Career
Researcher Program through a National Research Foundation (NRF)
grant (2013029711) funded by the Ministry of Education, Science,
and Technology (MEST), Korea; and a Systems Biology Infrastructure
Establishment grant provided by the Gwangju Institute of Science
and Technology (GIST) in 2013.
References
|
1
|
Bourguignon LY: CD44-mediated oncogenic
signaling and cytoskeleton activation during mammary tumor
progression. J Mammary Gland Biol Neoplasia. 6:287–297. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Zoller M: CD44: can a cancer-initiating
cell profit from an abundantly expressed molecule? Nat Rev Cancer.
11:254–267. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Marhaba R and Zoller M: CD44 in cancer
progression: adhesion, migration and growth regulation. J Mol
Histol. 35:211–231. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Herrlich P, Morrison H, Sleeman J, et al:
CD44 acts both as a growth- and invasiveness-promoting molecule and
as a tumor-suppressing cofactor. Ann NY Acad Sci. 910:106–120.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Screaton GR, Bell MV, Bell JI and Jackson
DG: The identification of a new alternative exon with highly
restricted tissue expression in transcripts encoding the mouse
Pgp-1 (CD44) homing receptor. Comparison of all 10 variable exons
between mouse, human, and rat. J Biol Chem. 268:12235–12238.
1993.
|
|
6
|
Herrera-Gayol A and Jothy S: Adhesion
proteins in the biology of breast cancer: contribution of CD44. Exp
Mol Pathol. 66:149–156. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gunthert U, Hofmann M, Rudy W, et al: A
new variant of glycoprotein CD44 confers metastatic potential to
rat carcinoma cells. Cell. 65:13–24. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rudy W, Hofmann M, Schwartz-Albiez R, et
al: The two major CD44 proteins expressed on a metastatic rat tumor
cell line are derived from different splice variants: each one
individually suffices to confer metastatic behavior. Cancer Res.
53:1262–1268. 1993.PubMed/NCBI
|
|
9
|
Kaufmann M, Heider KH, Sinn HP, von
Minckwitz G, Ponta H and Herrlich P: CD44 variant exon epitopes in
primary breast cancer and length of survival. Lancet. 345:615–619.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tempfer C, Losch A, Heinzl H, et al:
Prognostic value of immunohistochemically detected CD44 isoforms
CD44v5, CD44v6 and CD44v7–8 in human breast cancer. Eur J Cancer.
32A:2023–2025. 1996.
|
|
11
|
Sinn HP, Heider KH, Skroch-Angel P, et al:
Human mammary carcinomas express homologues of rat
metastasis-associated variants of CD44. Breast Cancer Res Treat.
36:307–313. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Reber S, Matzku S, Gunthert U, Ponta H,
Herrlich P and Zoller M: Retardation of metastatic tumor growth
after immunization with metastasis-specific monoclonal antibodies.
Int J Cancer. 46:919–927. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Seiter S, Arch R, Reber S, et al:
Prevention of tumor metastasis formation by anti-variant CD44. J
Exp Med. 177:443–455. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Orian-Rousseau V, Chen L, Sleeman JP,
Herrlich P and Ponta H: CD44 is required for two consecutive steps
in HGF/c-Met signaling. Genes Dev. 16:3074–3086. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Recio JA and Merlino G: Hepatocyte growth
factor/scatter factor induces feedback up-regulation of CD44v6 in
melanoma cells through Egr-1. Cancer Res. 63:1576–1582.
2003.PubMed/NCBI
|
|
16
|
Cheng C, Yaffe MB and Sharp PA: A positive
feedback loop couples Ras activation and CD44 alternative splicing.
Genes Dev. 20:1715–1720. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tsukita S, Oishi K, Sato N, Sagara J and
Kawai A: ERM family members as molecular linkers between the cell
surface glycoprotein CD44 and actin-based cytoskeletons. J Cell
Biol. 126:391–401. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Legg JW and Isacke CM: Identification and
functional analysis of the ezrin-binding site in the hyaluronan
receptor, CD44. Curr Biol. 8:705–708. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yonemura S, Hirao M, Doi Y, Takahashi N,
Kondo T and Tsukita S: Ezrin/radixin/moesin (ERM) proteins bind to
a positively charged amino acid cluster in the juxta-membrane
cytoplasmic domain of CD44, CD43, and ICAM-2. J Cell Biol.
140:885–895. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wahl MC, Will CL and Luhrmann R: The
spliceosome: design principles of a dynamic RNP machine. Cell.
136:701–718. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cooper TA, Wan L and Dreyfuss G: RNA and
disease. Cell. 136:777–793. 2009. View Article : Google Scholar
|
|
22
|
Xu Q and Lee C: Discovery of novel splice
forms and functional analysis of cancer-specific alternative
splicing in human expressed sequences. Nucleic Acids Res.
31:5635–5643. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kim E, Goren A and Ast G: Insights into
the connection between cancer and alternative splicing. Trends
Genet. 24:7–10. 2008. View Article : Google Scholar
|
|
24
|
Venables JP, Klinck R, Bramard A, et al:
Identification of alternative splicing markers for breast cancer.
Cancer Res. 68:9525–9531. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Black DL: Mechanisms of alternative
pre-messenger RNA splicing. Annu Rev Biochem. 72:291–336. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen H, Zheng X, Luecke S and Green MR:
The U2AF35-related protein Urp contacts the 3′ splice site to
promote U12-type intron splicing and the second step of U2-type
intron splicing. Genes Dev. 24:2389–2394. 2010.PubMed/NCBI
|
|
27
|
Konig H, Moll J, Ponta H and Herrlich P:
Trans-acting factors regulate the expression of CD44 splice
variants. EMBO J. 15:4030–4039. 1996.
|
|
28
|
Cho S, Moon H, Yang X, et al: Validation
of trans-acting elements that promote exon 7 skipping of
SMN2 in SMN2-GFP stable cell line. Biochem Biophys Res Commun.
423:531–535. 2012.
|
|
29
|
Lee J, Zhou J, Zheng X, et al:
Identification of a novel cis-element that regulates alternative
splicing of Bcl-x pre-mRNA. Biochem Biophys Res Commun.
420:467–472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Busch A and Hertel KJ: Evolution of SR
protein and hnRNP splicing regulatory factors. Wiley Interdiscip
Rev RNA. 3:1–12. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hertel KJ: Combinatorial control of exon
recognition. J Biol Chem. 283:1211–1215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Senapathy P, Shapiro MB and Harris NL:
Splice junctions, branch point sites, and exons: sequence
statistics, identification, and applications to genome project.
Methods Enzymol. 183:252–278. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen H and Green MR: RS domain-splicing
signal interactions in splicing of U12-type and U2-type introns.
Nat Struct Mol Biol. 14:597–603. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhong XY, Wang P, Han J, Rosenfeld MG and
Fu XD: SR proteins in vertical integration of gene expression from
transcription to RNA processing to translation. Mol Cell. 35:1–10.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stoilov P, Lin CH, Damoiseaux R, Nikolic J
and Black DL: A high-throughput screening strategy identifies
cardiotonic steroids as alternative splicing modulators. Proc Natl
Acad Sci USA. 105:11218–11223. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
van Weering DH, Baas PD and Bos JL: A
PCR-based method for the analysis of human CD44 splice products.
PCR Methods Appl. 3:100–106. 1993.PubMed/NCBI
|
|
37
|
Cartegni L, Wang J, Zhu Z, Zhang MQ and
Krainer AR: ESEfinder: a web resource to identify exonic splicing
enhancers. Nucleic Acids Res. 31:3568–3571. 2003.PubMed/NCBI
|
|
38
|
Hernandez F, Perez M, Lucas JJ, Mata AM,
Bhat R and Avila J: Glycogen synthase kinase-3 plays a crucial role
in tau exon 10 splicing and intranuclear distribution of SC35.
Implications for Alzheimer’s disease. J Biol Chem. 279:3801–3806.
2004.PubMed/NCBI
|
|
39
|
D’Souza I and Schellenberg GD:
Determinants of 4-repeat tau expression. Coordination between
enhancing and inhibitory splicing sequences for exon 10 inclusion.
J Biol Chem. 275:17700–17709. 2000.PubMed/NCBI
|
|
40
|
Graveley BR: Sorting out the complexity of
SR protein functions. RNA. 6:1197–1211. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lin S, Coutinho-Mansfield G, Wang D,
Pandit S and Fu XD: The splicing factor SC35 has an active role in
transcriptional elongation. Nat Struct Mol Biol. 15:819–826. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Qian W, Iqbal K, Grundke-Iqbal I, Gong CX
and Liu F: Splicing factor SC35 promotes tau expression through
stabilization of its mRNA. FEBS Lett. 585:875–880. 2011. View Article : Google Scholar : PubMed/NCBI
|