Introduction
Calcium mobilization from the endoplasmic reticulum
(ER) into the cytosol is a key component of several signaling
networks, including secretion, contraction, metabolism, neuronal
plasticity, gene transcription, cell growth, differentiation,
apoptosis or protein folding. The spatial and temporal control of
intracellular Ca2+ homeostasis is crucial for regulating
diverse Ca2+-dependent physiological events and is
regulated by the sarco(endo)plasmic reticulum
Ca2+-ATPases (SERCAs) via the hydrolysis of ATP. SERCA
proteins are encoded by a family of structurally related and
alternatively spliced transcripts from three distinct genes
(SERCA1, SERCA2 and SERCA3) expressed in a
tissue-dependent and/or developmentally regulated manner (1). Among them, SERCA3 is localized
to 17p13.3 with the encoding of 4698-bp mRNA, and is alternatively
spliced to generate six different transcripts encoding the isoforms
SERCA3a, SERCA3b, SERCA3c, SERCA3d,
SERCA3e and SERCA3f under the control of the
hypertension and protein kinase C, calcineurin, or retinoic acid
receptor signaling pathways. Decreased expression of SERCA3 in
response to biotin supplementation by influencing the nuclear
abundance of Sp1 and Sp3, suppressors of SERCA3, is likely
to affect the oxidative folding of secretory proteins in the ER
(2).
Although SERCA3 transcripts are most abundantly
expressed in lymphoid tissues, intestine, pancreas and prostate,
the highest SERCA3 expression is observed in cells of hematopoietic
lineage (platelets, mast cells and lymphocytes), embryologically
related endothelial cells, secretory epithelial cells (large and
small intestine, thymus and trachea), and Purkinje neurons in the
cerebellum. More interestingly, SERCA3 is always coexpressed with
the ubiquitous isoform SERCA2b (3,4).
Studies using SERCA3-deficient mice have revealed that lack of
SERCA3 alters the endothelium, epithelium-dependent relaxation in
vascular smooth muscle and epithelium-dependent relaxation in
tracheal smooth muscle (5,6). Downregulation of SERCA3 expression is
suggested to be responsible for the impaired glucose responses in
the islets of Langerhans in diabetic mouse and rat models of
non-insulin-dependent diabetes mellitus (7). In humans, missense mutations in the
SERCA3 gene are thought to render patients more susceptible to type
2 diabetes mellitus (8). In
contrast, knockout of the SERCA3 gene leads to defective
endothelium-dependent relaxation of vascular smooth muscle and
endothelial cell Ca2+ signaling, disruption of mixed
(Ca2+) oscillations, increased β-cell depolarization,
and higher insulin response of islets to glucose (6,9). It
was reported that SERCA3 expression is significantly reduced or
lost in colon carcinomas when compared with normal colonic
epithelial cells (10). Treatment
of tumor cells with butyrate or other differentiation inducing
agents was found to result in a marked and specific induction of
the expression of SERCA3, suggesting it constitutes a significant
new differentiation marker (10).
In our previous study, it was found that SERCA3 mRNA was
highly expressed in gastric carcinoma when compared with that in
matched non-neoplastic mucosa (NNM). SERCA3 protein expression was
decreased from gastric NNM, primary to metastatic carcinoma. SERCA3
expression was also found to be negatively related to depth of
invasion, distant metastasis and TNM staging of gastric carcinoma.
These findings suggest that downregulated SERCA3 expression is
closely linked to pathogenesis, invasion, metastasis and prognosis
of gastric carcinomas (11).
Colorectal cancer is one of the most common cancers
in the world, accounting for nearly 10% of newly diagnosed cases of
all cancer. Japan has experienced a marked increase in the
incidence of colorectal cancer, and has recently been listed among
countries with the world’s highest incidence rates (12,13).
Although pathological and genetic observations demonstrate that
colorectal adenoma precedes the majority of adenocarcinoma cases,
the molecular mechanisms underlying colorectal carcinogenesis are
poorly understood. To investigate the roles of SERCA3 expression in
colorectal carcinogenesis and subsequent progression, we examined
the expression of SERCA3 mRNA and protein in colorectal NNM,
adenoma, adenocarcinoma, and compared them with clinicopathological
parameters of carcinomas, including serum carcinoembryonic antigen
(CEA) concentration.
Materials and methods
Cell culture
The colorectal carcinoma cell lines were kindly
provided by Professor Sugiyama, Department of Gastroenterology,
Graduate School of Medical and Pharmaceutical Sciences, University
of Toyama, Japan and Professor Miyagi, Clinical Research Institute,
Kanagawa Cancer Center, Japan. They were maintained in RPMI-1640
(Colo201, Colo205, DLD-1, HCT-15, HCT-116, HT-29, KM-12, SW480 and
SW620) and DMEM (WiDr) supplemented with 10% fetal bovine serum
(FBS), 100 units/ml penicillin, and 100 μg/ml streptomycin in a
humidified atmosphere of 5% CO2 at 37°C. All cells were
harvested by centrifugation, rinsed with phosphate-buffered saline
(PBS) and subjected to total protein extraction by sonication in
RIPA lysis buffer [50 mmol/l Tris-HCl (pH 7.5), 150 mmol/l NaCl, 5
mmol/l EDTA, 0.5% Nonidet P-40, 5 mmol/l dithiothreitol, 10 mmol/l
NaF and protease inhibitor cocktail (Sigma)].
Subjects
Colorectal carcinomas (CRCs, n=481), adjacent
adenoma (n=22), adjacent non-neoplastic mucosa (NNM, n=475),
metastatic foci in lymph node metastasis (n=154) and in liver
(n=24) were collected from surgical resections at the Affiliated
Hospital, Kanagawa Cancer Center, Japan between 1995 and 1999. The
patients with CRC included 278 men and 203 women (age range, 26–85
years; mean age, 64.1 years). Among the patients, 209 cases were
accompanied by lymph node metastasis and 28 cases with liver
metastasis. Thirty cases of CRCs and paired NNM were obtained from
the First Affiliated Hospital of China Medical University, China,
and frozen at −80°C until protein and RNA extraction by
homogenization. None of the patients underwent chemotherapy,
radiotherapy or adjuvant therapy prior to surgery. The patients or
their relatives provided consent for the use of tumor tissue for
clinical research, and our University Ethics Committee approved the
research protocol. We followed up the patients by reviewing their
case documents or by telephone.
Pathology and tissue microarray
(TMA)
All tissues were fixed in 10% neutral formalin,
embedded in paraffin and cut into 4 μm sections. These sections
were stained with hematoxylin and eosin (H&E) to confirm their
histological characteristics. The staging for each colorectal
carcinoma was evaluated according to the Union Internationale
Contre le Cancer (UICC) system (14). Histological architecture of CRCs was
expressed in terms of WHO classification (15). Furthermore, tumor size, depth of
invasion, lymphatic and venous invasion were determined.
Representative areas of the solid tumors were
identified in the H&E-stained sections of the selected tumor
cases, and a 2-mm diameter tissue core per donor block was punched
out and transferred to a recipient block with a maximum of 48 cores
using a tissue microarrayer (Azumaya KIN-1; Azumaya, Tokyo,
Japan).
RT-PCR and DNA sequencing
Total RNA was extracted from the colorectal
carcinoma cell lines or tissues using Qiagen RNeasy Mini kit
(Qiagen, Hilden, Germany) according to the manufacturer’s protocol.
Two micrograms of total RNA was subjected to cDNA synthesis using
the AMV transcriptase and random primers (Takara, Otsu, Japan).
Oligonucleotide primers for PCR were 5′-AGTGCTCCGAAGACAACCC-3′ and
5′-GTTCTCCGAGACGCTGTTG-3′ for SERCA3 (134 bp, 2275–2908,
NM_174953.1); and sense, 5′-CAATGACCCCTTCATTGACC-3′ and antisense,
5′-TGGAAGATGGTGATGGGATT-3′ for GAPDH (135 bp, 201–335,
NM_002046.3). PCR amplification of cDNA was performed in 25-μl
mixtures containing 0.125 μl Pfu (Stratagene, West Cedar Creek,
USA) with 2.0 mmol MgCl2, 2.5 μl 10X PCR buffer, 2 μl
dNTP mixture, 1 μmol/l of each primer set and 100 ng of template
cDNA. PCR conditions were denaturation at 95°C for 10 min, followed
by 35 cycles of denaturation at 95°C for 30 sec, annealing for 30
sec and extension at 72°C for 50 sec. As a termination step, the
extension time of the last cycle was increased to 7 min. The
amplicons were electrophoresed in 2% agarose gel for 30 min.
Real-time PCR was performed according to the protocol of the SYBR
Premix Ex Taq™ II kit (Takara, Tokyo, Japan).
Amplicons were subjected to electrophoresis on 2%
agarose gel and purified with the QIAquick gel extraction kit
(Qiagen). After extraction, the DNA was quantified by a NanoDrop
ND-1000 spectrophotometer (Laboratory & Medical Supplies,
Tokyo, Japan) and then sequenced using a BigDye Terminator v3.1
cycle sequencing kit (Applied Biosystems, Foster City, CA, USA) and
SERCA3 primers according to the recommended protocol. The
sequence data were compared with the human SERCA3 cDNA sequence
(NM_174953.1) using BLAST.
Western blot analysis
Denatured protein was separated on an
SDS-polyacrylamide gel (10% acrylamide) and transferred to an
Amersham Hybond membrane (Amersham, Germany), which was then
blocked overnight in 5% skim milk in TBST (10 mmol/l Tris-HCl, 150
mmol/l NaCl, 0.1% Tween-20). For immunoblotting, the membrane was
incubated for 15 min with the mouse antibody against SERCA3 (1:200)
(XL-6, sc-101265; Santa Cruz Biotechnology, Santa Cruz, CA, USA).
Then, it was rinsed with TBST and incubated with anti-mouse IgG
conjugated to horseradish peroxidase (1:1,000) (Dako, Carpinteria,
CA, USA) for 15 min. All of the incubations were performed in a
microwave oven to allow intermittent irradiation as recommended by
Li et al(16). Bands were
visualized on X-ray film (Fujifilm, Tokyo, Japan) using ECL-Plus
detection reagents (Santa Cruz Biotechnology). Finally, the
membrane was washed with Western Blot Stripping Solution (pH
2.0–3.0; Nacalai, Tokyo, Japan) for 1 h and treated as previously
described except for the anti-GAPDH antibody (1:10,000) (Sigma) as
the internal control.
Immunofluorescence
Cells were grown on glass coverslips, washed twice
with PBS, fixed with 4% formaldehyde for 10 min at room temperature
and permeabilized with 0.2% Triton X-100 in PBS for 10 min at room
temperature. After washing with PBS, cells were incubated overnight
at 4°C with the mouse antibody against SERCA3 (1:50) (Santa Cruz
Biotechnology). The cells were then washed with PBS and incubated
with anti-mouse Alexa Fluor 594 IgG (1:2,000) (Invitrogen). Nuclei
were stained using 1 μg/ml DAPI (Sigma) for 30 min at 37°C.
Finally, coverslips were mounted with SlowFade® Gold
Antifade reagent (Invitrogen) and were observed under a laser
confocal scanning microscope (Leica, Wetzlar, Germany).
Immunohistochemistry
Consecutive sections were deparaffinized with
xylene, rehydrated with alcohol, and subjected to antigen retrieval
by irradiation in target retrieval solution (TRS; Dako) for 15 min
with a microwave oven (Oriental Rotor Co., Ltd., Tokyo, Japan). The
sections were quenched with 3% hydrogen peroxide in absolute
methanol for 2 min to block endogenous peroxidase activity. Bovine
serum albumin (5%) was then applied for 5 min to prevent
non-specific binding. The sections were incubated with the
anti-SERCA3 antibody (1:50) (XL-6, sc-101265; Santa Cruz
Biotechnology) for 15 min, and were then treated with the
anti-mouse conjugated to horseradish peroxidase (Dako) antibody for
15 min. All of the incubations were performed in a microwave oven
to allow intermittent irradiation as previously described (17). After each treatment, the slides were
washed with TBST three times for 1 min. Binding sites were
visualized with 3,3′-diaminobenzidine. After counterstaining with
Mayer’s haematoxylin, the sections were dehydrated, cleared and
mounted. Omission of the primary antibody was used as a negative
control.
As indicated in Fig.
1, SERCA3 protein was positively localized in the cytoplasm.
One hundred cells were randomly selected and counted from 5
representative fields of each section in a blinded manner by two
independent observers (W.-F.G. and H.-C.Z.). Inconsistent data were
assessed by both observers until a final consensus was reached. The
positive percentage of counted cells was graded semi-quantitatively
according to a 4-tier scoring system: negative (−), 0–5%; weakly
positive (+), 6–25%; moderately positive (++), 26–50%; and strongly
positive (+++), >50%.
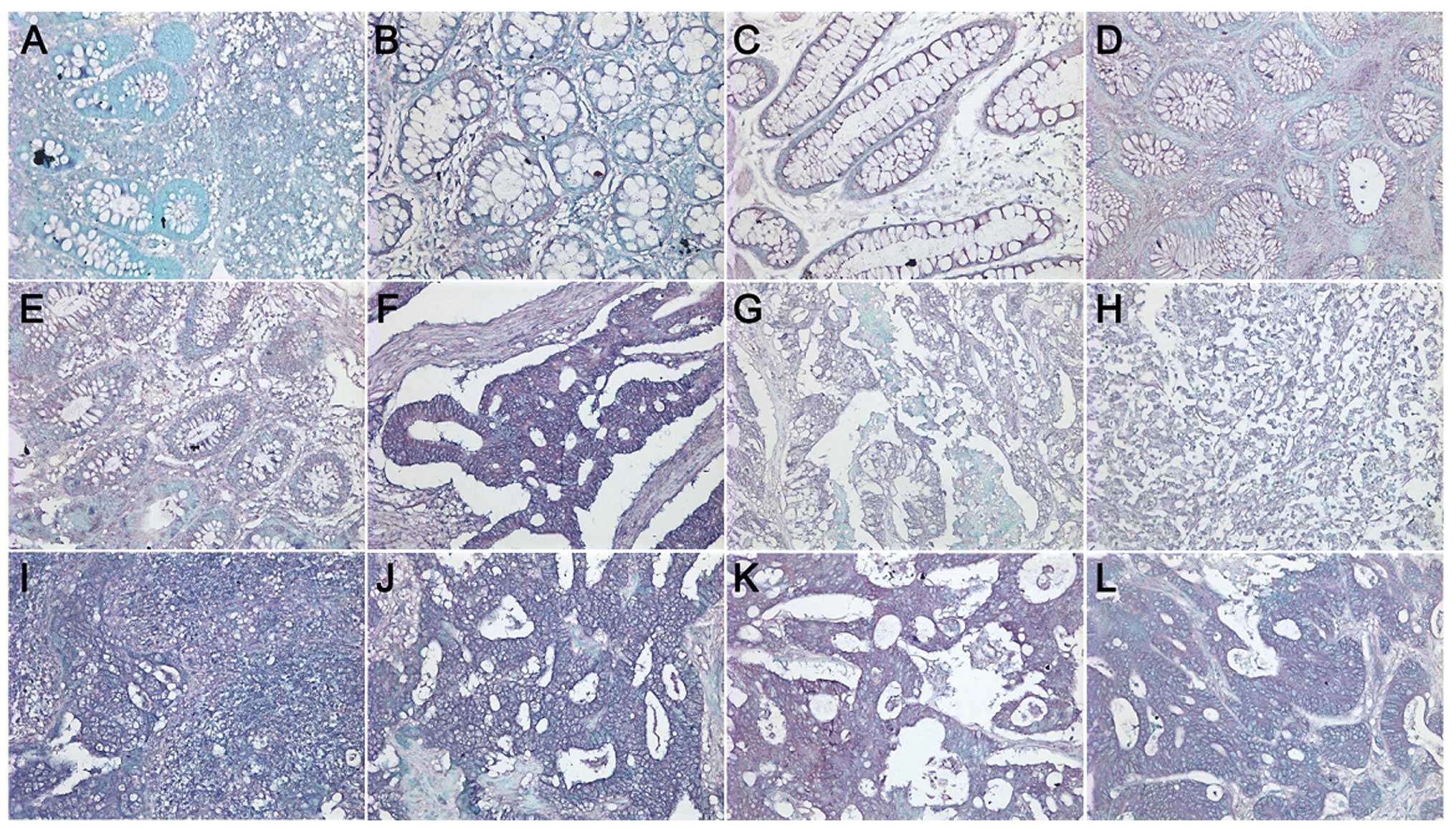
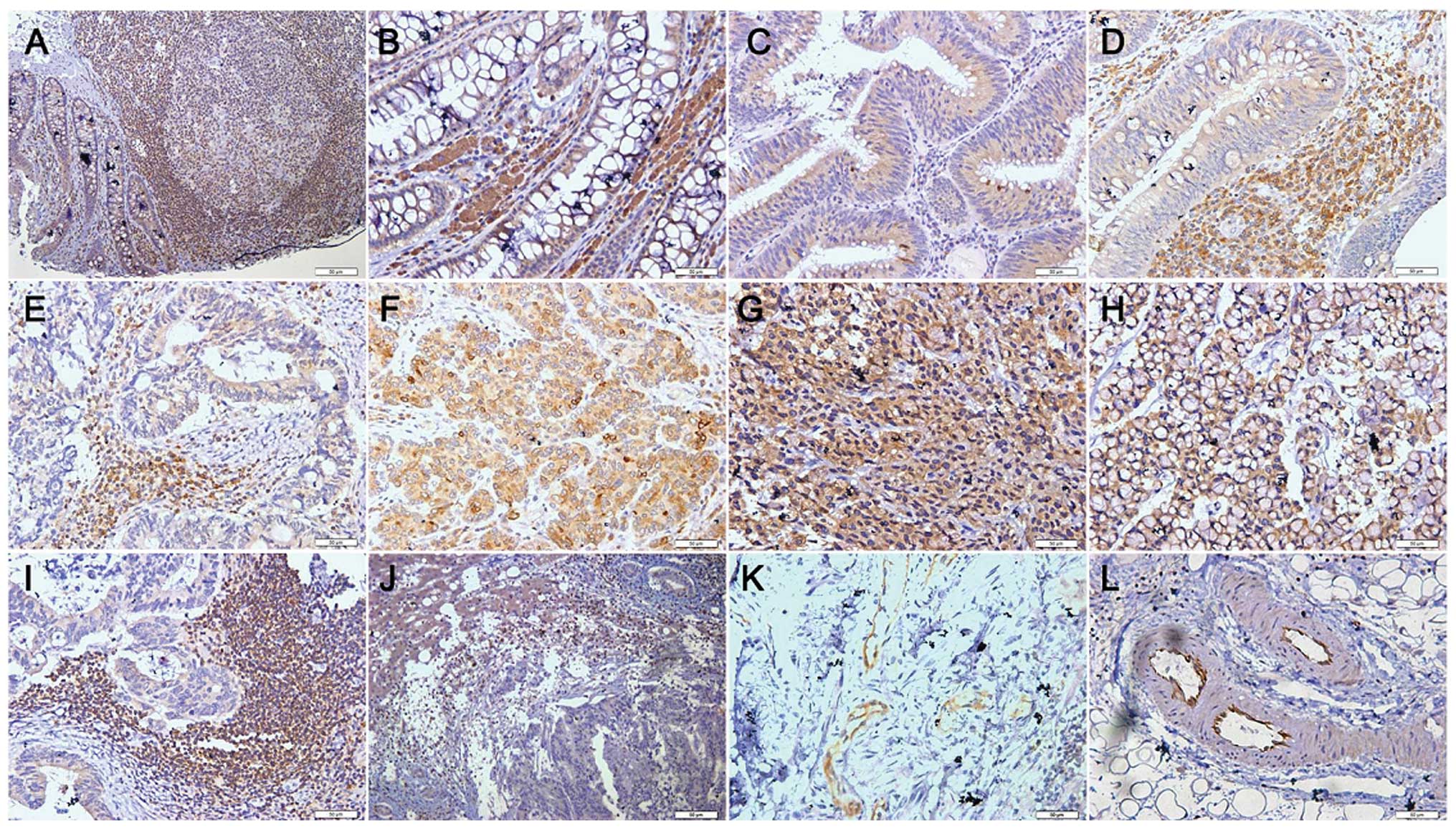 | Figure 1Immunostaining analysis of SERCA3 in
colorectal lesions. SERCA3 was expressed in the (A and B) cytoplasm
of the superficial epithelium, (A) infiltrating inflammatory cells,
(B) macrophages, (A) lymphoid follicle, (C and D) adenoma, and (E)
well-differentiated, (F) moderately-differentiated, and (G)
poorly-differentiated adenocarcinoma, (H) signet ring cell
carcinoma, (I) metastatic carcinoma in lymph node and (J) liver,
(K) endothelial cells in vein and (L) artery. |
In situ hybridization
To perform RNA-DNA in situ hybridization
(ISH) for SERCA3, a digoxygenin-labeled SERCA3 probe was
constructed in 35-cycle PCR using the above-mentioned primers and
50 ng template DNA of 30-cycle products from the DLD-1 cDNA using
Pfu polymerase (Stratagene, La Jolla, CA, USA). Sections (4-μm)
were deparaffinized and digested with 20 μg/ml proteinase K in 50
mmol/l Tris-HCl at 37°C for 10 min. Subsequently, 20 μl of a 1:20
probe dilution in hybridization buffer (22 mmol/l Tris-HCl, pH 7.4,
2.75 mmol/l ethylenediaminetetraacetic acid, 660 mmol/l NaCl, 1X
Denhardt solution, 5.5% dextran sulfate, 0.33% dimethyl sulfoxide,
0.55% Ethoquad® 18/25 and 44% deionized formamide) was
added to each slide. After coverslipping, heating at 95°C for 5
min, and incubation overnight in a humidified chamber at 37°C,
sections were rinsed for 10 min in TBST and incubated with
anti-digoxygenin antibody conjugated with alkaline phosphatase (AP;
Roche Diagnostics, GmbH, Penzberg, Germany) for 20 min at 37°C. The
slides were then washed for 5 min and immersed in solution II (100
mmol/l Tris-HCl, pH 9.5, 100 mmol/l NaCl and 50 mmol/l
MgCl2) for 30 min followed by exposure to NBT (Nitroblue
tetrazolium chloride)/BCIP (5-bromo-4-chloro-3′-indolyl phosphatase
p-toluidine salt) as a chromogen. Finally, counter staining
was performed using methyl green for 2 min.
As indicated in Fig.
2, the mRNA signal of SERCA3 was positively localized in
the cytoplasm. One hundred cells were randomly selected and counted
from 5 representative fields of each section in a blinded manner by
two independent observers (W.-F.G. and H.-C.Z.). Inconsistent data
were assessed by both observers until a final consensus was
reached. The scoring system was the same as that for
immunohistochemistry.
Measurement of carcinoembryonic antigen
(CEA)
Serum CEA was determined using a chemiluminescence
immunoassay (Diagnostic Agnostic Automation Inc.). Briefly, 50 μl
of standard (0–120 ng/ml), specimens and controls was dispensed
into appropriate wells. We then added 100 μl of the enzyme
conjugate reagent into each well, gently mixed and incubated the
plate at room temperature for 60 min. The microtiter wells were
rinsed and flicked with wash buffer. After that, residual water
droplets were removed by striking the well sharply onto absorbent
paper. Finally, 100 μl chemiluminescence substrate solution into
each well was dispensed, mixed gently and subjected to
determination of absorbance.
Statistical analysis
Statistical analysis was performed using the
Spearman correlation test to analyze the rank data, and the
Mann-Whitney U test to differentiate the means of the different
groups. Kaplan-Meier survival plots were generated, and comparisons
were carried out with log-rank statistics. Cox’s proportional
hazards model was employed for multivariate analysis. P<0.05 was
considered to indicate a statistically significant result. SPSS
10.0 software was employed to analyze all data.
Results
SERCA3 expression in colorectal tumors
and carcinoma cell lines
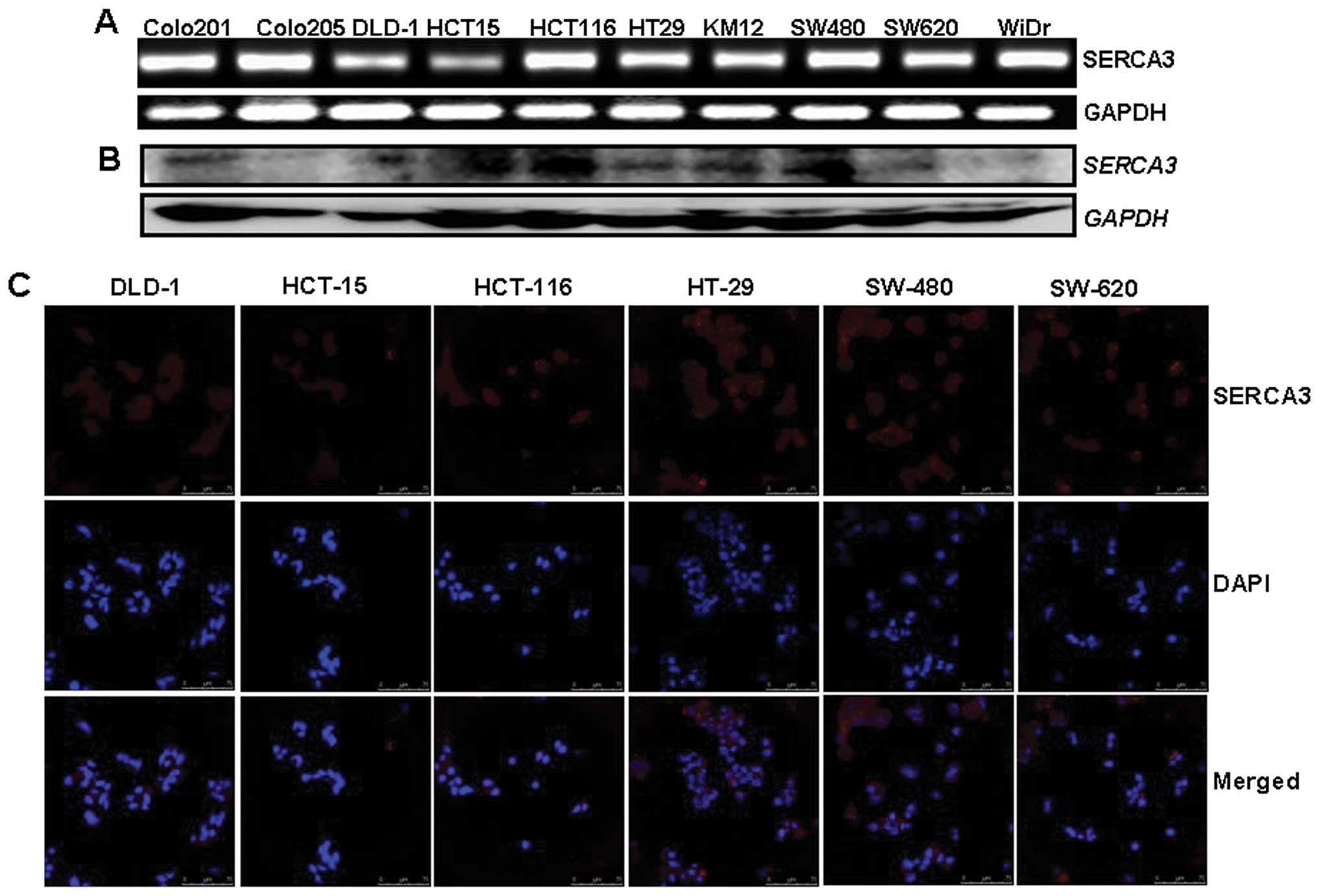
SERCA3 protein was detected in the cytoplasm of all
colorectal carcinoma cell lines by western blot analysis and
immunofluorescence (Fig. 3A and C).
To assess SERCA3 mRNA expression, we employed RT-PCR.
SERCA3 mRNA was expressed at equal levels in these carcinoma
cell lines (Fig. 3B). Additionally,
all of the amplicons were subjected to direct DNA sequencing and
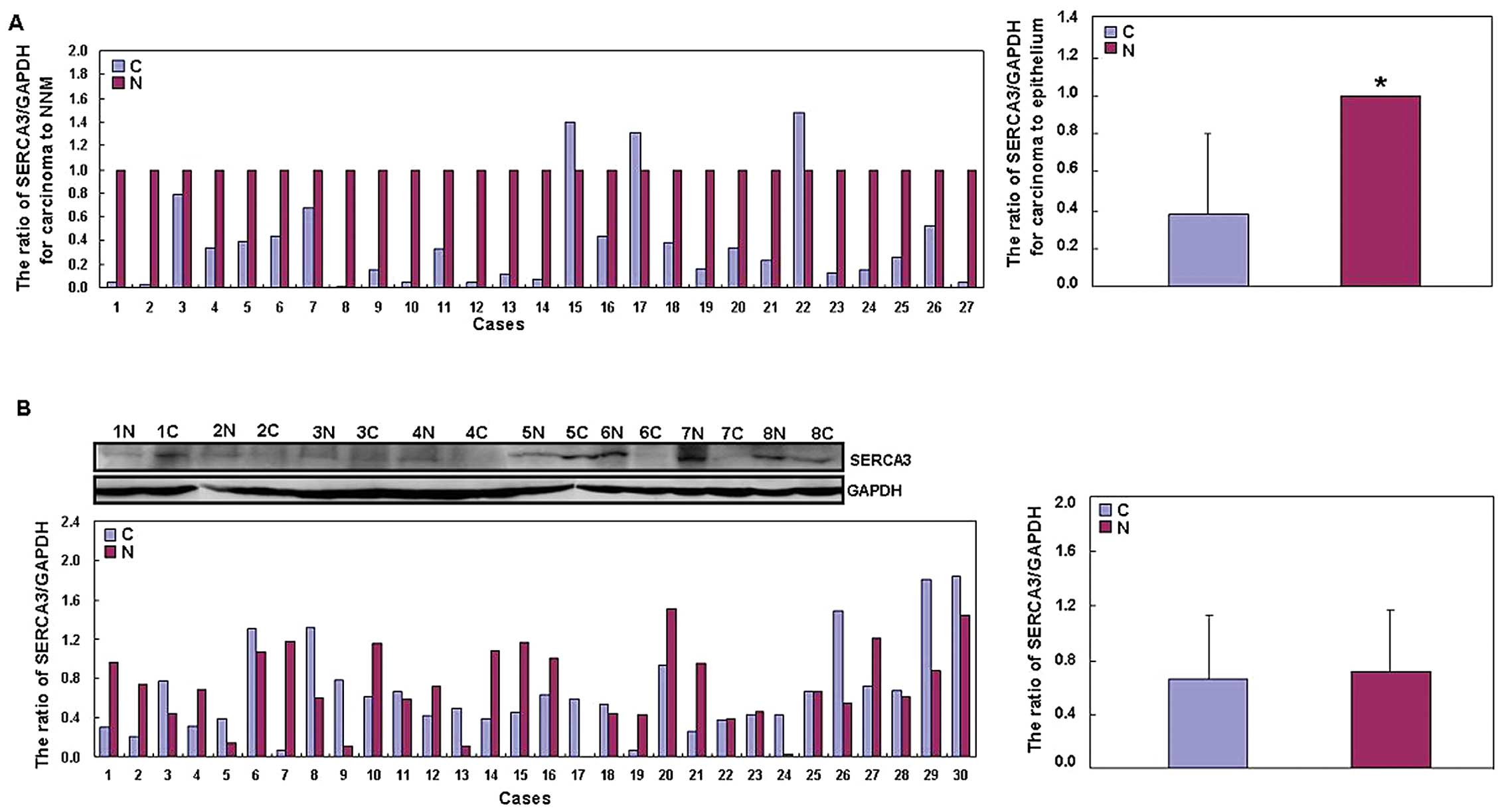
confirmed to be accurate (data not shown). Among 27 frozen samples
of colorectal carcinoma, decreased SERCA3 mRNA expression
was observed in 24 cases (88.9%) when compared with the mRNA
expression in the adjacent mucosa by real-time PCR (Fig. 4A). SERCA3 mRNA expression was
low in the carcinoma samples than that in the matched NNM
(P<0.05). However, there was no difference in SERCA3 expression
between carcinoma and paired NNM by western blot analysis
(P>0.05; Fig. 4B).
As shown in Fig. 1,
SERCA3 was expressed in the cytoplasm of the superficial mucosa,
infiltrating inflammatory cells, macrophages, lymphoid follicle,
adenoma, well-differentiated, moderately differentiated and poorly
differentiated adenocarcinoma, signet ring cell carcinoma,
metastatic carcinoma in lymph node and liver, and endothelial cells
in the vein and artery. SERCA3 expression was detected in 93.3%
(443/475) of the NNM, 95.5% (21/22) of adjacent adenomas, 50.8%
(245/482) of primary carcinomas, 32.5% (50/154) of the metastatic
carcinomas in lymph node and 29.2% (7/24) of metastatic carcinomas
in the liver. When statistics was performed, we considered the
4-graded semi-quantitative scoring and employed Spearman
correlation analysis. Low expression of SERCA3 was observed in the
colorectal carcinoma cases when compared to that in the NNM and
adenomas (P<0.05; Table I). In
contrast, primary carcinoma showed higher SERCA3 expression than
metastatic carcinomas in the lymph node or liver (P<0.05;
Table I).
 | Table ISERCA3 expression in colorectal
non-neoplastic mucosa, adenomas and carcinomas. |
Table I
SERCA3 expression in colorectal
non-neoplastic mucosa, adenomas and carcinomas.
| | SERCA3
expression | |
|---|
| |
| |
|---|
| Groups | n | − | + | ++ | +++ | PR (%) |
|---|
| NNMs | 475 | 32 | 37 | 121 | 285 | 93.3 |
| Adenomas adjacent to
carcinoma | 22 | 1 | 1 | 7 | 13 | 95.5 |
| Primary
carcinoma | 481 | 236 | 85 | 91 | 69 | 50.8a |
| Metastatic carcinoma
in lymph node | 154 | 104 | 24 | 17 | 9 | 32.5b |
| Metastatic carcinoma
in liver | 24 | 17 | 5 | 2 | 0 | 29.2c |
To confirm SERCA3 mRNA expression, in
situ hybridization (ISH) was also employed on TMAs of
colorectal carcinoma, NNM, adenoma, primary and metastatic
carcinoma (Fig. 2). The positive
signal of SERCA3 mRNA was detectable in the cytoplasm of
colorectal epithelial cells, adenoma, carcinoma, infiltrating
inflammatory cells or lymphocytes in lymphatic follicles.
SERCA3 mRNA was detected in colorectal NNM (48.7%, 37/76),
adenoma (57.0%, 53/93), primary carcinoma (64.0%, 71/111),
metastatic carcinoma in lymph node (82.6%, 38/46) and liver (65.4%,
17/26). Statistically, its expression was lower in NNM than that in
the colorectal carcinoma (P<0.05; Table II). SERCA3 mRNA was
expressed at a lower level in the primary carcinoma when compared
to that in the metastatic carcinoma in lymph node (P<0.05;
Table II).
| Table IISERCA3 mRNA expression in
colorectal non-neoplastic mucosa, adenomas and carcinomas. |
Table II
SERCA3 mRNA expression in
colorectal non-neoplastic mucosa, adenomas and carcinomas.
| | SERCA3
expression | |
|---|
| |
| |
|---|
| Groups | n | − | + | ++ | +++ | PR (%) |
|---|
| NNMs | 76 | 39 | 15 | 12 | 10 | 48.7 |
| Adenomas adjacent
to carcinoma | 93 | 40 | 13 | 18 | 22 | 57.0 |
| Primary
carcinoma | 111 | 40 | 27 | 22 | 22 | 64.0a |
| Metastatic
carcinoma in lymph node | 46 | 8 | 2 | 18 | 18 | 82.6 |
| Metastatic
carcinoma in liver | 26 | 9 | 3 | 2 | 12 | 65.4 |
Association of SERCA3 expression with
clinicopathological parameters of the colorectal carcinoma
cases
As shown in Table
III, SERCA3 expression was negatively correlated with lymphatic
invasion (P<0.05), but not with age, gender, depth of invasion,
venous invasion, lymph node metastasis, distant metastasis, TNM
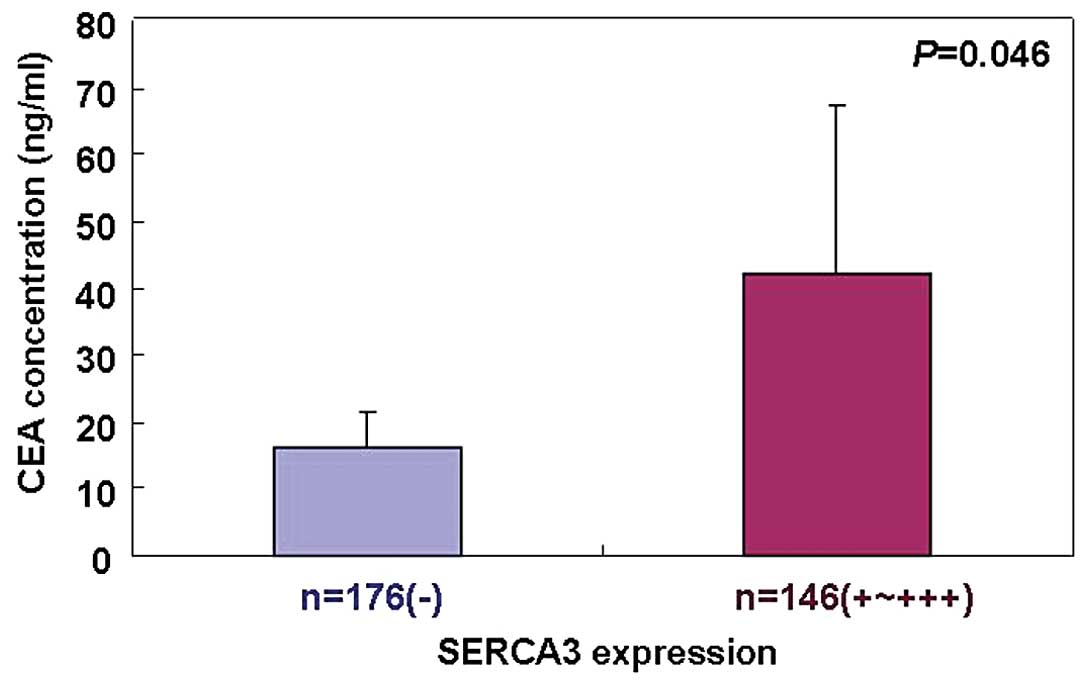
stage or degree of differentiation (P>0.05). The serum CEA
concentration was higher in the carcinoma patients with positive
SERCA3 expression than that in patients without SERCA3 positivity
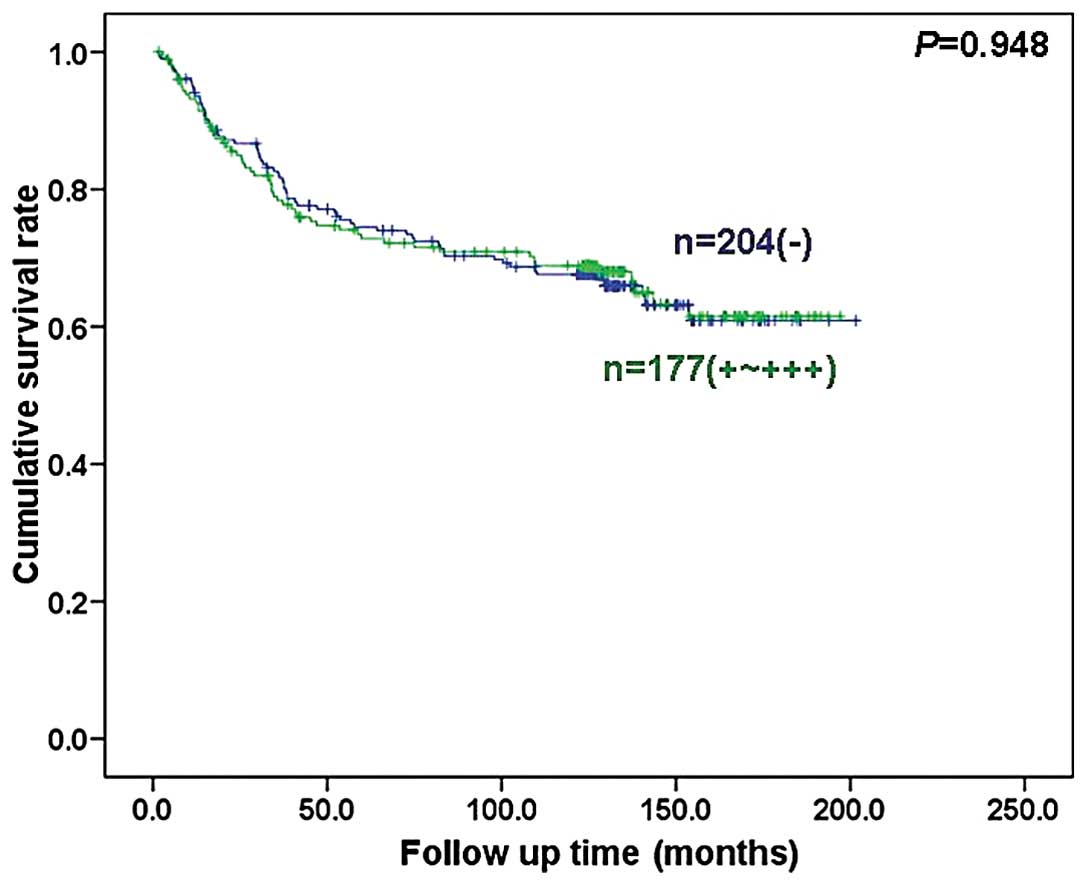
(P<0.05; Fig. 5). Follow-up
information was available for 381 colorectal carcinoma patients for
a period ranging from 1.6 months to 16.8 years (median, 10.5
years). Survival curves for colorectal carcinomas were stratified
according to SERCA3 expression (Fig.
6). Univariate analysis using the Kaplan-Meier method indicated
that the cumulative survival rate of patients was not linked to
SERCA3 expression status (P>0.05), even when the data were
stratified according to the depth of invasion (data not shown).
Multivariate analysis using Cox’s proportional hazard model
indicated that depth of invasion and distant metastasis
(P<0.05), but not age, gender, lymphatic and venous invasion,
lymph node metastasis, liver metastasis, differentiation, TNM stage
or SERCA3 expression (P>0.05; Table
IV) are independent prognostic factors for overall survival of
the colorectal carcinoma parients.
| Table IIIRelationship between SERCA3
expression and clinicopathological features of the CRC cases. |
Table III
Relationship between SERCA3
expression and clinicopathological features of the CRC cases.
| Clinicopathological
features | n | SERCA3
expression | PR (%) | P-value |
|---|
|
|---|
| − | + | ++ | +++ |
|---|
| Age (years) | | | | | | | 0.941 |
| <65 | 260 | 126 | 48 | 50 | 36 | 51.5 | |
| ≥65 | 221 | 110 | 37 | 41 | 33 | 50.2 | |
| Gender | | | | | | | 0.865 |
| Male | 278 | 134 | 56 | 51 | 37 | 51.8 | |
| Female | 203 | 102 | 29 | 40 | 32 | 49.8 | |
| Depth of
invasion | | | | | | | 0.641 |
|
Tis-T2 | 123 | 58 | 23 | 22 | 20 | 52.8 | |
|
T3–T4 | 338 | 168 | 58 | 63 | 49 | 50.3 | |
| Lymphatic
invasion | | | | | | | 0.015 |
| − | 195 | 82 | 38 | 39 | 36 | 57.9 | |
| + | 207 | 112 | 32 | 38 | 25 | 45.9 | |
| Venous
invasion | | | | | | | 0.473 |
| − | 193 | 99 | 33 | 30 | 31 | 48.7 | |
| + | 196 | 92 | 33 | 43 | 28 | 53.1 | |
| Lymph node
metastasis | | | | | | | 0.211 |
| − | 253 | 119 | 43 | 50 | 41 | 53.0 | |
| + | 209 | 108 | 37 | 38 | 26 | 48.3 | |
| Distant
metastasis | | | | | | | 0.895 |
| − | 441 | 216 | 76 | 84 | 65 | 51.0 | |
| + | 36 | 17 | 8 | 7 | 4 | 52.8 | |
| TNM stage | | | | | | | 0.424 |
| I–II | 242 | 116 | 42 | 44 | 40 | 52.1 | |
| III–IV | 217 | 109 | 38 | 43 | 27 | 49.8 | |
|
Differentiation | | | | | | | 0.136 |
|
Well-differentiated | 194 | 88 | 32 | 38 | 36 | 54.6 | |
|
Moderately-differentiated | 231 | 118 | 44 | 45 | 24 | 48.9 | |
|
Poorly-differentiated | 32 | 16 | 5 | 4 | 7 | 50.0 | |
| Table IVMultivariate analysis of the
clinicopathological variables for the overal survival of the CRC
patients. |
Table IV
Multivariate analysis of the
clinicopathological variables for the overal survival of the CRC
patients.
| Clinicopathological
parameters | Relative risk (95%
CI) | P-value |
|---|
| Age (≥65
years) | 0.875
(0.553–1.384) | 0.567 |
| Gender
(female) | 0.850
(0.544–1.327) | 0.474 |
| Depth of invasion
(T2–4) | 8.027
(2.439–26.424) | 0.001 |
| Lymphatic invasion
(+) | 1.242
(0.729–2.116) | 0.426 |
| Venous invasion
(+) | 1.017
(0.598–1.731) | 0.950 |
| Lymph node
metastasis (+) | 1.810
(0.549–5.961) | 0.329 |
| Distant
metastasis | 4.826
(2.292–10.162) | <0.001 |
| UICC staging
(I–II) | 1.657
(0.439–6.257) | 0.456 |
| Differentiation
(moderately and poorly) | 1.129
(0.790–1.614) | 0.505 |
| SERCA3 expression
(+−+++) | 1.076
(0.681–1.701) | 0.754 |
Discussion
Endoplasmic reticulum calcium homeostasis is
involved in a multitude of intracellular signaling, as well as
‘housekeeping’ functions that control cell growth, differentiation,
protein synthesis or apoptosis in eukaryotic cells. In several cell
systems, SERCA3 expression is selectively induced during
differentiation, whereas SERCA3 expression is decreased during
tumorigenesis and blastic transformation (1,2). In
the present study, we, for the first time, examined in situ
SERCA3 expression in a large number of colorectal mucosal and
carcinoma samples. It was found that SERCA3 protein was mainly
localized in the cytoplasm of the superficial epithelium,
infiltrating inflammatory cells, macrophages, lymphoid follicle,
adenoma, adenocarcinoma, signet ring cell carcinoma, and
endothelial cells in vein and artery. It was suggested that the
SERCA3 expression pattern has cellular specificity, which
determines its biological functions. However, the mechanisms of its
cell-specific characteristics warrant further investigation.
To clarify the in situ expression pattern and
the clinicopathological significance of the SERCA3 protein, IHC and
ISH were performed on TMAs of colorectal lesions. Statistically,
SERCA3 expression was reduced in the colorectal carcinoma, when
compared with that in the adjacent non-neoplastic mucosa and
adenoma in line with previous reports (11,18).
Real-time PCR showed that SERCA3 mRNA was weakly expressed
in carcinoma in comparison with the adjacent NNM. The data
indicated that downregulation of SERCA3 expression may contribute
to the malignant transformation of colorectal epithelial cells as a
late event and its transcript expression as an early event. In our
frozen samples, we found no difference in SERCA3 expression between
most of the colorectal carcinoma and the matched mucosa. The
paradoxical phenomenon may be explained by the different detecting
approaches due to the presence of SERCA3 expression in stromal
cells from colorectal samples, which were excluded from
morphological data by virtue of their specific histomorphological
features and topographic location in the tissue sections. The short
half life of SERCA3 might be due to its downregulated expression in
colorectal carcinoma possibly via proteosome-mediated degradation.
Strangely, higher SERCA3 mRNA expression was detected in the
carcinoma by ISH, when compared with that in the adjacent NNM,
which may be due to different sample treatment and detection
approaches. Brouland et al(18) reported that SERCA3 expression was
increased along the crypts as cells differentiated in normal
colonic mucosa and in hyperplastic polyps; was moderately and
heterogeneously expressed in colonic adenomas with expression
levels inversely correlated with the degree of dysplasia; and was
barely detectable in adenocarcinomas. During the multi-step process
of colon carcinogenesis, the decrease in SERCA3 expression appears
to be linked to enhanced APC/β-catenin/TCF4 signaling and deficient
Sp1-like factor-dependent transcription (19,20).
In the present study, SERCA3 expression was lower in
the metastatic carcinomas in the liver and lymph node, and the
cases with lymphatic invasion although it was not linked to
aggressive behaviors of the colorectal carcinomas, such as
invasion, metastasis and TNM stage. In our previous report, SERCA3
expression was inversely correlated with depth of invasion, distant
metastasis and TNM stage of gastric cancer. Combined with these
findings, we believe that SERCA3 expression may be involved in the
development of gastrointestinal carcinomas. Previously, we reported
that SERCA3 expression was positively linked to the favorable
prognosis of gastric carcinoma (11). However, this relationship did not
exist for the patients with colorectal carcinoma, which might be
explained by the absence of a correlation of its expression with
aggressive behaviors. Multivariate analysis demonstrated that the
depth of invasion and distant metastasis are independent prognostic
factors for overall survival of the colorectal carcinoma
patients.
Brouland et al(18) found that treatment of gastric or
colorectal carcinoma cells with butyrate or other established
differentiation-inducing agents resulted in a marked induction in
the expression of SERCA3 and higher resting cytosolic calcium
concentrations, suggesting that SERCA3 constitutes a significant
new differentiation marker that may prove useful for the analysis
of the phenotype of gastrointestinal adenocarcinomas. However, no
relationship between SERCA3 expression and the degree of
differentiation of colorectal carcinoma was found in the present
study although the depolarization of SERCA3 was observed with worse
differentiation. This result may be due to the small number of
poorly-differentiated carcinoma cases. In agreement with the
present finding, there was no difference in SERCA3 expression
between intestinal- and diffuse-type carcinoma and even between
intestinal and diffuse components of mixed-type carcinoma of the
stomach (11).
CEA is a glycosyl phosphatidyl inositol (GPI)-cell
surface anchored glycoprotein and serves as functional colon
carcinoma L-selectin and E-selectin ligands. It was found that
serum from individuals with gastric, pancreatic, lung, breast and
medullary thyroid carcinoma had higher levels of CEA than healthy
individuals (>2.5 ng/ml). In colorectal carcinoma, CEA might be
employed to predict early progression and worse prognosis (21–23).
Reportedly, as little as 1% serum admixed with tumor cells results
in a CEA release up to 200% greater than that of serum-free
controls, which is dependent on calcium (24). Since SERCA3 is involved in calcium
mobility of the endoplasmic reticulum, the positive link between
SERCA3 expression and serum CEA level is rational in patients with
colorectal carcinoma. However, further investigation of how SERCA3
regulates CEA secretion in malignancies is warranted.
In summary, the present study revealed that aberrant
SERCA3 expression impacts the malignant transformation of
colorectal epithelial cells. Nevertheless, the biological functions
of SERCA3 in colorectal carcinomas need further investigation.
Acknowledgements
The present study was supported by the Shenyang
Outstanding Talent Foundation of China; the Shenyang Science and
Technology Grant (F11-264-1-10 and F12-277-1-01); the Natural
Scientific Foundation of China (nos. 81172371 and 81201886); and
the grant-in aid for the Scientific Research from the Ministry of
Education, Culture, Sports and Technology of Japan (23659958).
References
|
1
|
Hovnanian A: SERCA pumps and human
diseases. Subcell Biochem. 45:337–363. 2007. View Article : Google Scholar
|
|
2
|
Wuytack F, Dode L, Baba-Aissa F, et al:
The SERCA3-type of organellar Ca2+ pumps. Biosci Rep.
15:299–306. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Baba-Aïssa F, Raeymaekers L, Wuytack F, et
al: Purkinje neurons express the SERCA3 isoform of the organellar
type Ca(2+)-transport ATPase. Brain Res Mol Brain Res. 41:169–174.
1996.PubMed/NCBI
|
|
4
|
Chandrasekera PC, Kargacin ME, Deans JP,
et al: Determination of apparent calcium affinity for endogenously
expressed human sarco(endo)plasmic reticulum calcium-ATPase isoform
SERCA3. Am J Physiol Cell Physiol. 296:C1105–C1114. 2009.
View Article : Google Scholar
|
|
5
|
Kao J, Fortner CN, Liu LH, et al: Ablation
of the SERCA3 gene alters epithelium-dependent relaxation in mouse
tracheal smooth muscle. Am J Physiol Lung Cell Mol Physiol.
277:L264–L270. 1999.PubMed/NCBI
|
|
6
|
Liu LH, Paul RJ, Sutliff RL, et al:
Defective endothelium-dependent relaxation of vascular smooth
muscle and endothelial cell Ca2+ signalling in mice
lacking sarco(endo)plasmic reticulum Ca2+-ATPase isoform
3. J Biol Chem. 272:30538–30545. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Roe MW, Philipson LH, Frangakis CJ, et al:
Defective glucose-dependent endoplasmic reticulum Ca2+
sequestration in diabetic mouse islets of Langerhans. J Biol Chem.
269:18279–18282. 1994.PubMed/NCBI
|
|
8
|
Varadi A, Lebel L, Hashim Y, et al:
Sequence variants of the sarco(endo)plasmic reticulum
Ca2+-transport ATPase 3 gene (SERCA3) in Caucasian type
II diabetic patients (UK Prospective Diabetes Study 48).
Diabetologia. 42:1240–1243. 1999.PubMed/NCBI
|
|
9
|
Arredouani A, Guiot Y, Jonas JC, et al:
SERCA3 ablation does not impair insulin secretion but suggests
distinct roles of different sarcoendoplasmic reticulum
Ca2+ pumps for Ca2+ homeostasis in pancreatic
β-cells. Diabetes. 51:3245–3253. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gélébart P, Kovács T, Brouland JP, et al:
Expression of endomembrane calcium pumps in colon and gastric
cancer cells. Induction of SERCA3 expression during
differentiation. J Biol Chem. 277:26310–26320. 2002.PubMed/NCBI
|
|
11
|
Xu XY, Yang X, Takahashi H, et al: The
down-regulated SERCA3 expression is closely linked to pathogenesis,
invasion, metastasis and poor prognosis of gastric carcinomas.
Tumour Biol. 33:1845–1854. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Colorectal Cancer Facts and Figures.
American Cancer Society; 2005
|
|
13
|
Yoshida D, Kono S, Moore MA, et al:
Colorectal polypectomy and risk of colorectal cancer by subsite:
the Fukuoka Colorectal Cancer study. Jpn J Clin Oncol. 37:597–602.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hamilton SR and Aaltonen LA: WHO
Classification of Tumors: Pathology and Genetics of Tumors of the
Digestive System. IARC Press; Lyon: 2000
|
|
15
|
Sobin LH and Wittekind CH: TNM
Classification of Malignant Tumours. 6th edition. John Wiley &
Sons; Hoboken, NJ: 2002
|
|
16
|
Li W, Murai Y, Okada E, et al: Modified
and simplified western blotting protocol: use of intermittent
microwave irradiation (IMWI) and 5% skim milk to improve binding
specificity. Pathol Int. 52:234–238. 2002.PubMed/NCBI
|
|
17
|
Kumada T, Tsuneyama K, Hatta H, Ishizawa
S, et al: Improved 1-h rapid immunostaining method using
intermittent microwave irradiation: practicability based on 5 years
application in Toyama Medical and Pharmaceutical University
Hospital. Mod Pathol. 17:1141–1149. 2004.
|
|
18
|
Brouland JP, Gélébart P, Kovàcs T, et al:
The loss of sarco/endoplasmic reticulum calcium transport ATPase 3
expression is an early event during the multistep process of colon
carcinogenesis. Am J Pathol. 167:233–242. 2005. View Article : Google Scholar
|
|
19
|
Arbabian A, Brouland JP, Gélébart P, et
al: Endoplasmic reticulum calcium pumps and cancer. Biofactors.
37:139–149. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Brouland JP, Valleur P and Papp B:
Expression of SERCA pumps during cell differentiation and
tumorigenesis: application to colonic carcinogenesis. Ann Pathol.
26:159–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Petrioli R, Licchetta A, Roviello G, et
al: CEA and CA19.9 as early predictors of progression in
advanced/metastatic colorectal cancer patients receiving
oxaliplatin-based chemotherapy and bevacizumab. Cancer Invest.
3:65–71. 2012. View Article : Google Scholar
|
|
22
|
Lan YT, Lin JK, Lin TC, et al: Prognostic
significance of perioperative change of CEA level in colorectal
patients when pre-operative level is normal.
Hepatogastroenterology. 59:717–720. 2012.PubMed/NCBI
|
|
23
|
Lin JK, Lin CC, Yang SH, et al: Early
postoperative CEA level is a better prognostic indicator than is
preoperative CEA level in predicting prognosis of patients with
curable colorectal cancer. Int J Colorectal Dis. 26:1135–1141.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yeatman TJ, Duan C, Mao W, et al:
Augmentation of carcinoembryonic antigen release from intact,
viable tumor cells by a factor in human serum. Ann Surg Oncol.
2:336–342. 1995. View Article : Google Scholar : PubMed/NCBI
|