Introduction
Glioma, one of the most aggressive malignancies, is
the most common primary brain tumor in adults, accounting for 80%
of primary malignant brain tumors (1,2).
Astrocytoma presents the most common type. To date, despite major
research efforts and progress in therapy, including neurosurgical
technology, radiation therapy, chemotherapy and molecular therapy,
the prognosis of gliomas remains poor. Patients with malignant
glioma have a median survival rate of only 15 months (3–5).
Excessive proliferation, resistance to apoptotic stimuli,
neovascularization, disseminated tumor growth and suppression of
antitumor immune surveillance are key biological features that
contribute to the malignant phenotype of gliomas (6,7).
Maximal surgical resection of the tumor mass,
combined with chemotherapy and/or radiotherapy, is the standard of
care (8,9). However, chemotherapy has been hampered
due to resistance of these tumor cells to available agents and
challenges in delivering agents targeting the tumor cells. Gliomas
often respond to radiotherapy, but subsequent recurrence is almost
inevitable, which suggests insufficient ability to kill the
tumorigenic cells.
Radioresistance is a widespread challenge in
malignant glioma therapy (10). In
a previous study from our laboratory, we found that
phosphoglycerate kinase 1 (PGK1) was significantly upregulated in
radioresistant astrocytomas (11),
which suggested that PGK1 may be involved in the radioresistant
phenotype and may be a potential biomarker for developing better
therapeutic methods.
PGK1 is an ATP-generating glycolytic enzyme in the
glycolytic pathway catalyzing the conversion of
1,3-diphosphoglycerate to 3-phosphoglycerate and further affects
the DNA replication and repair (12). It was previously shown that the
increased expression of PGK1 was detected in colon cancer tissue
from metastatic patients (13),
lung adenocarcinoma (14), gastric
(15), prostate (16), pancreatic (17), and multidrug resistant ovarian
cancer, but was moderate in adjacent non-tumor tissues (18) and resistant in ovarian cancer cells
(19).
In view of the involvement of PGK1 in malignant
tumors, in the present study, U251 cells, a human glioma cell line,
were selected as our in vitro model to investigate whether
PGK1 is involved in the radioresistance. Our results showed that an
increased PGK1 expression was observed in radioresistant U251
cells, and it could promote radioresistance in U251 human glioma
cells.
Materials and methods
Cell culture
Human glioma U251 cells, obtained from Nanjing
KeyGen Biotech. (Co. Ltd., Nanjing, China) were cultured in
Dulbecco’s modified Eagle’s medium (DMEM; Gibco-Invitrogen,
Carlsbad, CA, USA) supplemented with 10% fetal calf serum
(Gibco-Invitrogen), 100 U/ml penicillin and 100 μg/ml streptomycin.
Cells were incubated at 37°C in a humidified incubator containing
5% CO2.
Establishment of radioresistant U251
cells (RR-U251)
U251 cells were irradiated as monolayer culture by
the γ-rays of a 60Co source at 0.5 Gy/min for 10 min per
time until the accumulation was 60 Gy (20).
Transfection
The shRNA-PGK1 plasmid and the high expression
plasmid pcDNA3.1-PGK1 were synthesized by Shanghai GenePharma Co.,
Ltd. (Shanghai, China). shRNA-PGK1 was designed to target the
coding region of the human PGK1 sequence
(5′-GCAAGGATGTTCTGTTCTTGA-3′; Gene ID:5230). A negative control
(5′-GTTCTCCGAACGTGTC ACGT-3′) was also obtained from Shanghai
GenePharma.
When U251 cells in 6-well plates were grown to
70–80% confluence, 4 μg shRNAs-PGK1 or the high expression plasmids
and 8 μl Lipofectamine™ 2000 (Invitrogen) were diluted in 240 μl
Opti-MEM medium separately, after 5-min standing, the diluted
plasmids and Lipofectamine 2000 were mixed and added into each well
after 20 min standing (final concentration for plasmids was 2
μg/ml). Six hours after transfection, the medium was replaced with
fresh DMEM containing 10% fetal bovine serum (FBS), and the cells
were incubated for an additional 18 h (24 h after transfection) for
radiation treatment.
Total RNA extraction and real-time
polymerase chain reaction (real-time PCR)
Total RNA was extracted in each group (normal U251
cells and established RR-U251 cells) with SV Total RNA Isolation
system (Promega, Madison, WI, USA) according to the manufacturer’s
protocol. The total RNA was then reverse transcribed to cDNA with
reverse transcription System (Promega). mRNA expression was
determined by real-time PCR using GoTaq® qPCR Master Mix
under standard thermocycler conditions (Eppendorf AG22331, Hamburg,
Germany).
Sequences of PCR primer pairs were: human GAPDH,
forward, 5′-CGCTGAGTACGTCGTGGAGTC-3′ and reverse,
5′-GCTGATGATCTTGAGGCTGTTGTC-3′. Human PGK1, forward,
5′-ATGCTGAGGCTGTCACTCGG-3′ and reverse,
5′-CACAGCAAGTGGCAGTGTCTCC-3′.
The PCR amplification began with an initial
denaturation step at 95°C for 2 min followed by 40 cycles at 95°C
for 45 sec, 58°C for 45 sec, and 72°C for 45 sec. Relative gene
expression values were determined using the 2−ΔΔCT
method as previously described (21). The human GAPDH gene was used as an
endogenous control for sample normalization. Results were presented
as fold increases relative to the expression of human GAPDH.
Western blot analysis
U251 cells were washed with cold phosphate-buffered
saline (PBS) three times and then scraped from the well. The cells
were lysed with lysis buffer (Thermo Fisher Scientific, Runcorn,
UK). The cell lysate was centrifuged and the supernatant was
collected. All samples were diluted in loading buffer (Nanjing
Sunshine Biotechnology, Co., Ltd., Nanjing, China) and boiled for 3
min. Samples (30 μg) were loaded on a 10% sodium dodecyl
sulfate-polyacrylamide gel by electrophoresis (SDS-PAGE).
Prestained markers were used to estimate molecular weight. Proteins
were transferred to polyvinylidene difluoride membranes (Nanjing
Sunshine Biotechnology), blocked by 5% skim milk and incubated
overnight at 4°C with antibodies from mouse against human PGK1
diluted in TBST buffer (1:1,000; Abcam, Cambridge, MA, USA),
followed by a goat anti-mouse horseradish peroxidase-conjugated
secondary antibody at 37°C for 1 h (1:10,000). The membranes were
washed three additional times with TBST, and antibody-antigen
complexes were revealed using an enhanced chemiluminescent (ECL)
technique, using a SuperSignal West Pico Trial kit obtained from
Thermo Fisher Scientific according to the manufacturer’s
instructions. The membrane was stripped and reprobed with a primary
antibody against α-tubulin (1:400 dilution; Nanjing Sunshine
Biotechnology) as a control. Quantitation was performed by computer
with commercial software (ImageQuan Molecular Dynamics, Sunnyvale,
CA, USA).
Cell viability assay
The cell viability was determined by the MTT
[3-(4,5-dimethylthiazol)-2,5-diphenyltetrazolium bromide] assay
(Sigma-Aldrich, St. Louis, MO, USA). Briefly, with or without
radiation by 60Co, the normal U251 cells and RR-U251
cells 5×103 cells/well were seeded in 96-well plates and
allowed to attach for 48 h. Each group contained eight wells. MTT
(20 μl) was added to each well and the cells were incubated at 37°C
for 4 h. The reaction was then stopped by adding 150 μl of dimethyl
sulfoxide (DMSO) for 10 min. Optical density (OD) was obtained at a
wavelength of 570 nm using a microplate reader. Viability was
calculated by the following equation: Cell viability (%) = 100 ×
(OD treatment/OD control) %.
Cell migration assay
The wound-healing assay (WHA) method was used to
evaluate cell migration ability. The normal U251 and RR-U251 cells,
treated with shRNA-PGK1, overexpression PGK1 or untreated, were
cultured in 6-well plates until ~80–90% confluence was reached. The
monolayer cells, with or without radiation by 60Co, were
scratched out using the tip of a 10 μl pipette to create the wound
line. To monitor the migration, images were captured with a
microscope at 0 and 24 h at the same site, and the migration
distance was measured. The ratio of the increased area by cell
migration after 24 h to that at 0 h was calculated to quantitate
the extent of migration. Migration ratio = (Width0h -
Width24h)/Width0h × 100%.
Cell invasion assessment
Cell invasion abilities were examined using a
24-well Transwell culture chamber system. The filter membrane with
8-μm pores was coated with Matrigel (1:8 diluted in DMEM medium; BD
Biosciences, Franklin Lakes, NJ, USA). The normal U251 and RR-U251
cells, treated with shRNA-PGK1, overexpression PGK1 or untreated,
were cultured in 6-well plates until confluence was reached. The
monolayer cells, before irradiation or 6 h after radiation by
60Co, were trypsinized and resuspended in serum-free
DMEM at a density of 1×105/ml. The 100 μl suspension was
seeded into the upper chambers. DMEM supplemented with 15% FBS was
added to the lower chamber. Following incubation for 24 h, the
upper chamber was taken out and the cells remaining on the upper
surface of the membrane were removed very lightly using a medical
cotton bud. The invaded cells on the other side were fixed with
dehydrated alcohol and stained with 0.1% crystal violet for several
minutes. The chamber was viewed under a light microscope and images
were captured in 5 randomly selected ×200 fields for each well.
Three separate experiments were performed.
Statistical analysis
The analysis was performed using SPSS 13.0.
Statistical significance was analyzed using the Student’s t-test
and the Mann-Whitney U test. Data are expressed as the means ±
standard deviation (SD). P<0.05 was considered to indicate a
statistically significant difference.
Results
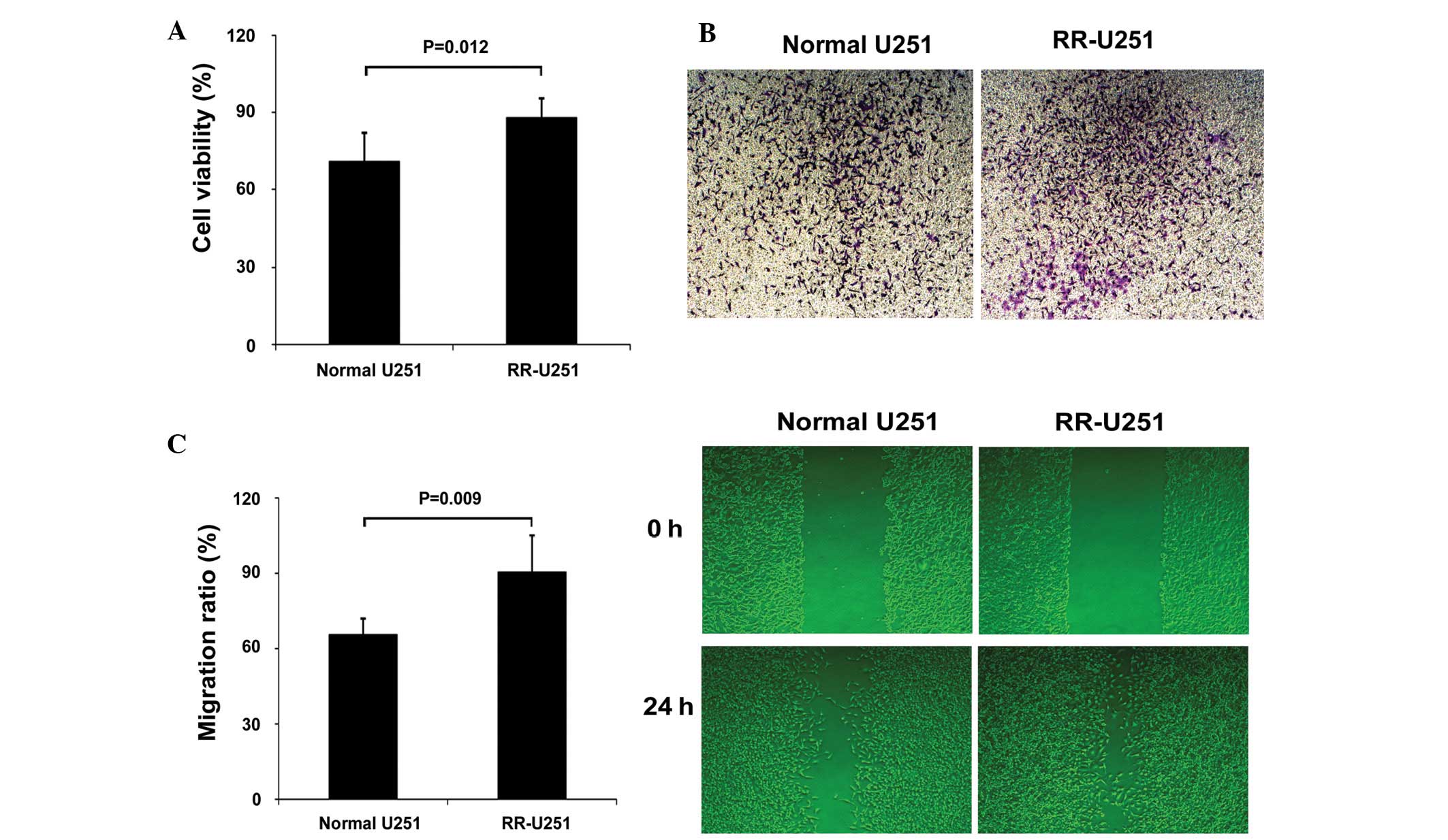
RR-U251 cells and measurement of the
radiosensitivity
RR-U251 cells were established from normal U251
cells, irradiated by a 60Co source at 0.5 Gy/min for 10
min per time until the accumulation was 60Gy, and the
radiosensitivity was characterized by measuring the cell viability,
migration and invasion abilities after radiation treatment. The
results showed that the cell viability was significantly increased
in the radiation-treated RR-U251 cells, compared with
radiation-treated normal U251 cells. Similar results were also
observed in cell migration and invasion assays. All results
suggested that the radiosensitivity was decreased in RR-U251 cells
(Fig. 1).
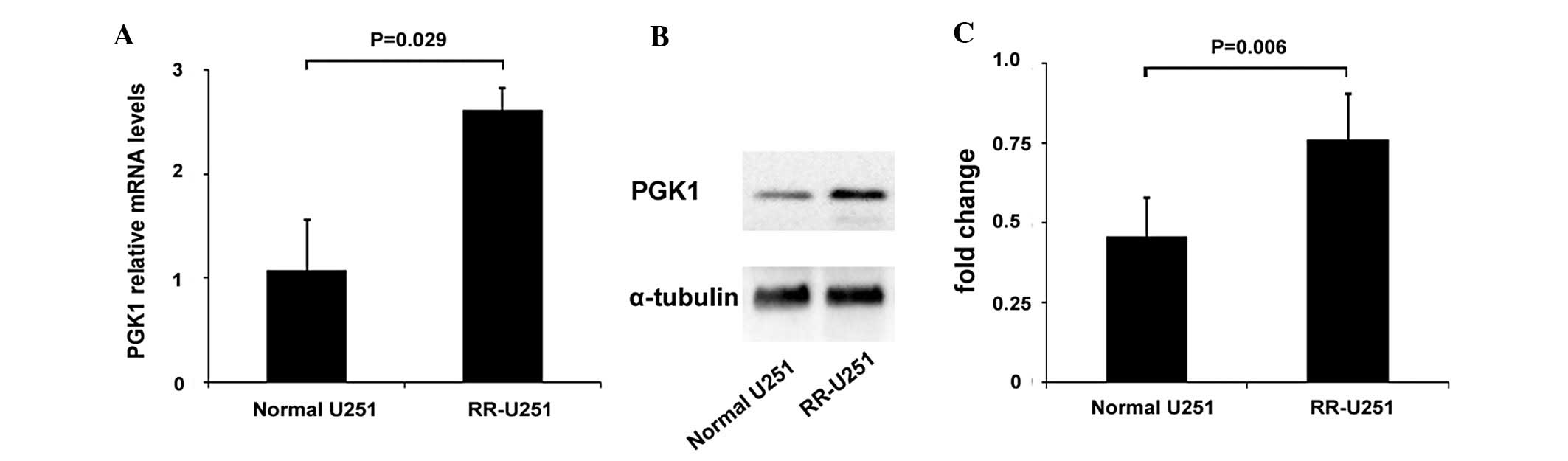
Elevated expression of PGK1 in RR-U251
cells
The PGK1 expression was determined by real-time PCR
and western blot analysis. Compared with normal U251 cells,
elevated expression of both mRNA and protein levels of PGK1 was
observed in RR-U251 cells (Fig.
2).
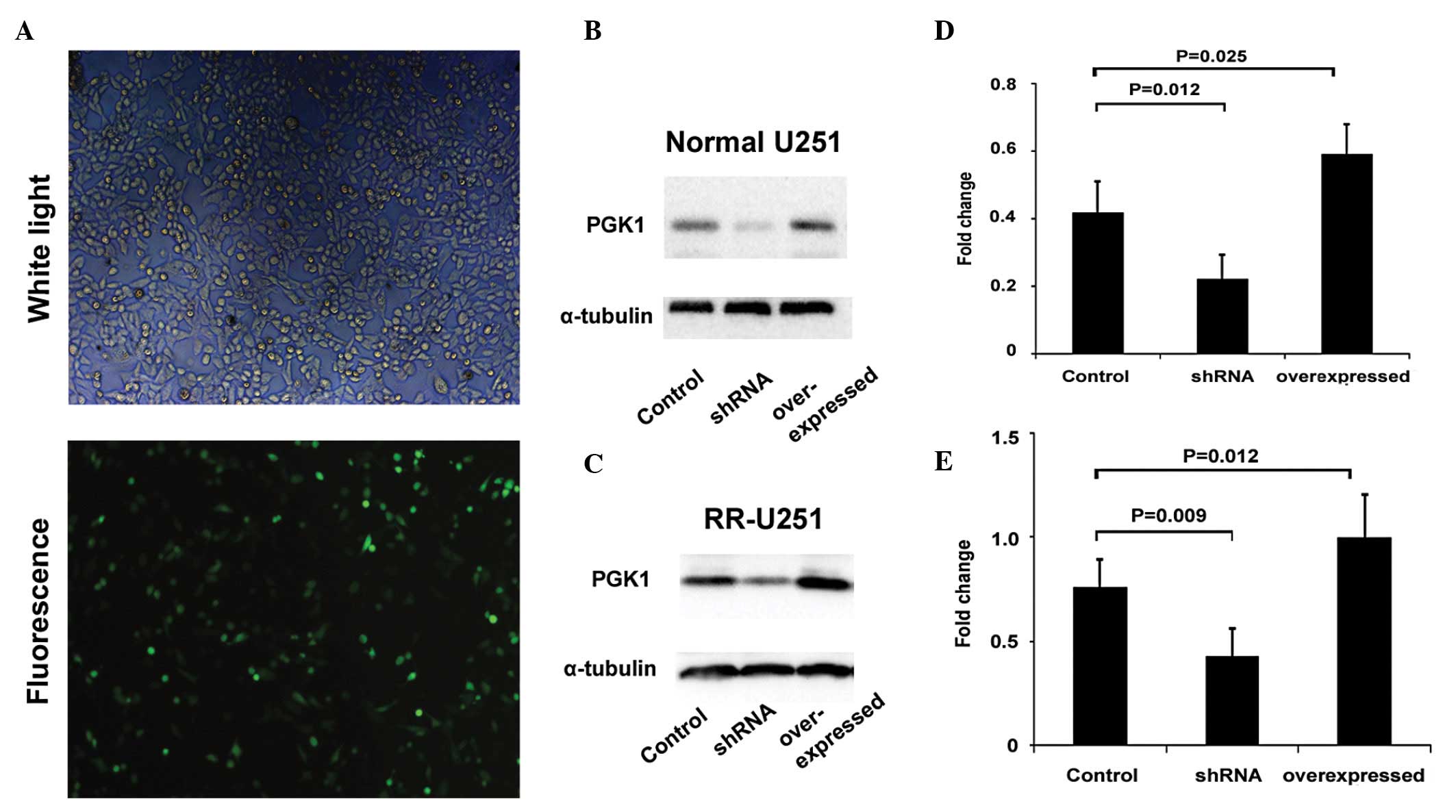
Confirmation of the transfection
efficiency
Normal U251 and RR-U251 cells were transfected with
shRNA-PGK1 to decrease the expression of PGK1. As shown in Fig. 3A, transfection of shRNA-PGK1 led to
a stable exogenous gene expression with ~50–60% efficiency in U251
cells as indicated by the GFP-labeled reporter fluorescence. We
further confirmed that both normal U251 and RR-U251 cells, which
were transfected with shRNA-PGK1, showed a significantly lower
expression of PGK1 as compared with those transfected with an
irrelevant sequence control. The cells, transfected with high
expression plasmid pcDNA3.1-PGK1, also showed higher PGK1
expression as compared with controls confirmed by western blot
analysis (Fig. 3B and C). We also
tested the PGK1 expression 24 and 48 h after transfection, and
found that after 24 h the role was more efficient (data not shown).
The time-point of 24 h after transfection was, therefore, used in
the following experiments with radiation treatment.
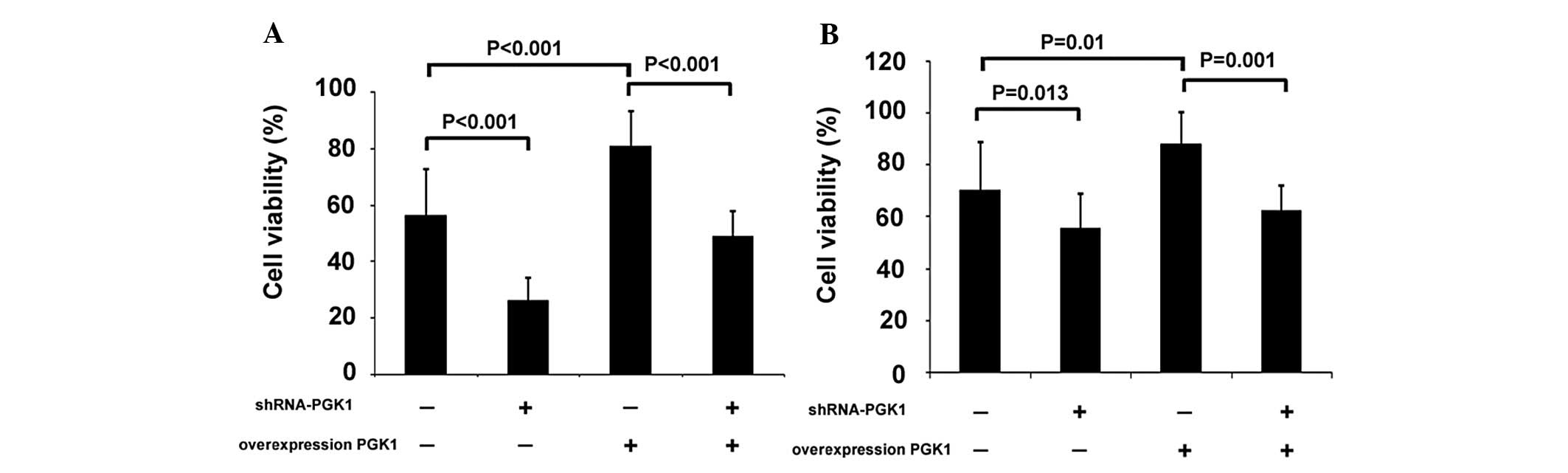
PGK1 modulates U251 cell
proliferation
The proliferation rates of U251 cells with enhanced
or decreased expression of PGK1 were determined by the MTT assay.
Compared to control cells, cells transfected with shRNA-PGK1, both
normal U251 and RR-U251 cells, proliferated at a significantly
lower rate. In contrast, overexpression of PGK1 by infection with
high expression plasmid pcDNA3.1-PGK1 resulted in significantly
enhanced proliferation in both normal U251 and RR-U251 cells. We
further downregulated PGK1 expression in PGK1 overexpressed U251
cells, using shRNA-PGK1 transfection. The shRNA-PGK1-transfected
cells showed significantly lower proliferation rate compared with
the control group, both in normal U251 and RR-U251 cells (Fig. 4).
PGK1 modulates U251 cell migration
ability
The migration ability of U251 cells with enhanced or
decreased expression of PGK1 was determined by the WHA method.
Following radiation therapy, both shRNA-PGK1-transfected normal
U251 and RR-U251 cells showed significantly decreased migration
abilities, compared with control cells. In contrast,
PGK1-overexpressed cells showed significantly enhanced migration
abilities in both normal U251 and RR-U251 cells. We also used
shRNA-PGK1 transfection to further downregulate PGK1 expression in
PGK1-overexpressed U251 cells. The migration ability of
PGK1-overexpressed cells was significantly decreased after
transfection with shRNA-PGK1, in both normal U251 and RR-U251 cells
(Fig. 5).
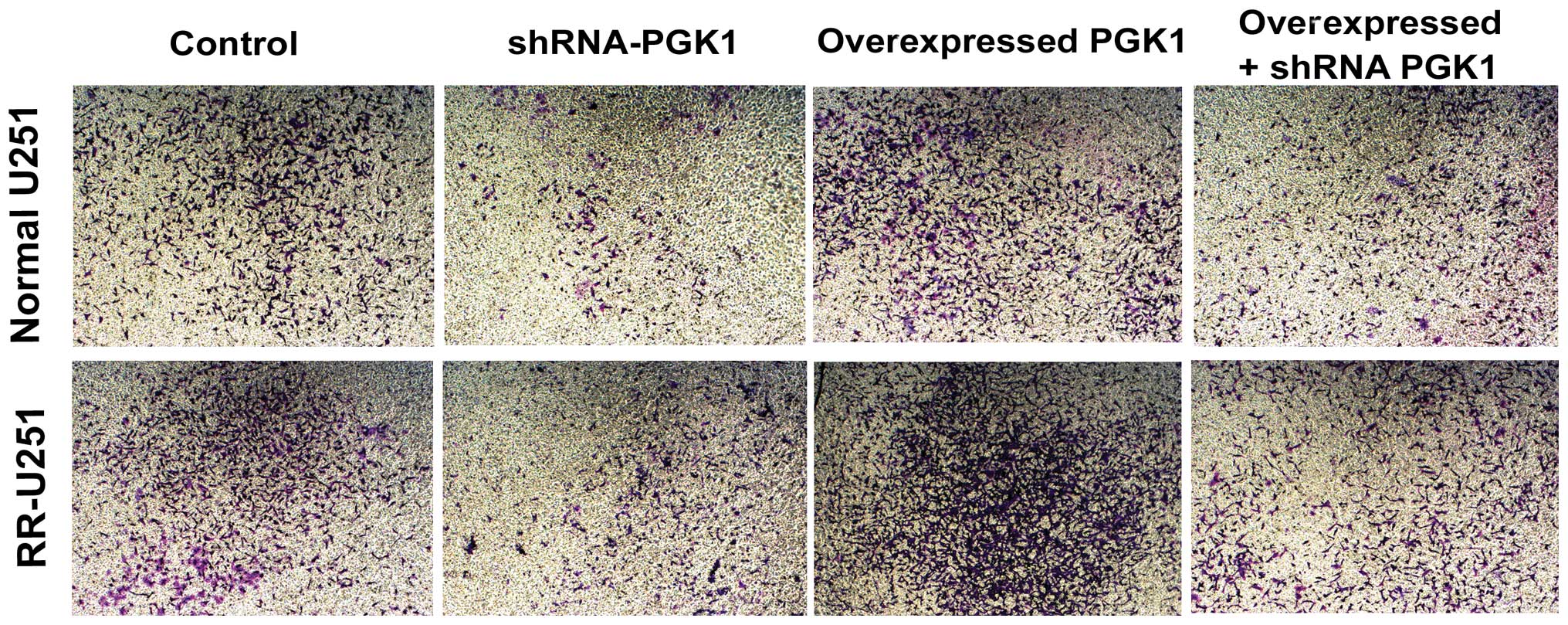
PGK1 modulates U251 cell invasion
ability
We also assessed the role of PGK1 in cell invasion
by Transwell assay. As shown in Fig.
6, compared with control cells, the invasion potential of U251
cells transfected with shRNA-PGK1 was significantly decreased both
in normal U251 and RR-U251 cells, while cells transfected with
pcDNA3.1-PGK1 displayed markedly increased invasive ability.
Following transfection with shRNA-PGK1, the invasion ability of
PGK1-overexpressed U251 cells also decreased compared with the
control group. These results suggest that PGK1 modulates U251 cell
invasion (Fig. 6).
Discussion
In the present study, we first observed that the
expression of PGK1 was upregulated in RR-U251 cells, which are
resistant to radiation. Further study confirmed that cell viability
and cell migration were much higher, and cells invaded more when
PGK1 was overexpressed, while blocking PGK1 in the cell viability,
cell migration and invasion assay revealed significant inhibition
of these parameters, suggesting that PGK1 was involved in the
radioresistance in human glioma by promoting the tumorigenic
properties, including cell viability, cell migration and invasion
ability.
Our previous study showed that patients with
radioresistant astrocytomas, compared to patients with
radiosensitive astrocytomas, displayed much higher protein level of
the glycolytic enzyme PGK1 (11).
We proposed that PGK1 was involved in the radioresistance in
glioma. U251 cells are glioma cancer cells that can induce highly
aggressive tumors of the central nervous system (22) and they are an accepted model to
study the biology of gliomas (23).
PGK1 is a metabolism-related ATP-generating glycolytic enzyme
overexpressed in a number of malignancies and generally correlates
with poor survival (14,24,25).
It catalyzes the reversible conversion of 1,3-diphosphoglycerate to
3-phosphoglycerate and further affects DNA replication and repair
(12). Increasing glycolysis in
radioresistant astrocytomas suggests a status of hypoxia. In a
range of human tumor sites, the presence of hypoxia is a negative
prognostic indicator for outcome following radiotherapy (26–28).
As is well known, hypoxia plays a central role in the pathobiology
of glioma (9). The key mediator,
the transcription factor hypoxia inducible factor 1 (HIF-1), is
stabilized under hypoxic conditions and promotes the production of
vascular endothelial growth factor (VEGF), which is critical for
neovascularization. Hypoxia-driven physiologic changes also include
the glycolytic pathway regulation and blood-vessel formation
(29) and also promote glioma cell
migration and invasion (30). PGK1,
a key enzyme of anaerobic glycolytic metabolism, is also one of the
target genes of HIF-1 (31), and we
found that PGK1 was upregulated in both radioresistant astrocytomas
and RR-U251 cells. Therefore, we speculate that in glioma
conditions, hypoxia augments HIF-1 secretion and consequently
affects PGK1 expression, resulting in resistance of glioma
radiotherapy. This hypothesis is in general agreement with that
reported by Lam et al (32),
who found that the induction of HIF-1 in hypoxic conditions can
induce the expression of PGK in non-small cell lung carcinoma
cells. We hypothesized that a vicious circle may arise between
increased PGK1 and radioresistance in glioma.
On the other hand, in the present study, our new
data concerning PGK1 provided comprehensive evidence of the Warburg
effect, a phenomenon first described more than 80 years ago that
cancer cells maintain a high glycolytic rate even in the presence
of oxygen (33). To explain the
conversion of glucose to lactate by cancer cells in the presence of
oxygen, Warburg speculated that tumor mitochondria are decreased or
functionally impaired. Previous studies renewed the Warburg effect
and clarified its exact role as cause, correlate, or facilitator of
cancer (34,35). Meanwhile it is largely accepted and
shown that solid tumor cells employ glycolytic enzymes, such as
PGK1, to produce ATP when their supply of oxygen is limited
(36). Furthermore, PGK1 has been
shown to be overexpressed in several malignancies, such as
metastatic colon cancer (13), lung
adenocarcinoma (14), gastric
(15), prostate (16), pancreatic (17) and ovarian cancer (18–19).
Meanwhile, our results were generally consistent with a recent
study performed on ovarian cancer, in which siRNA-PGK1 sensitized
chemoresistant human ovarian cancer cell lines to cisplatin
(37). These findings support the
comprehension between glucose metabolism and cancer generation and
may serve as a potential therapeutic target to prevent metastasis
of cancer cells. Our results presented here suggest that PGK1 could
promote radioresistance in human U251 cells, and manipulation of
PGK1 may offer a novel therapeutic approach for the treatment of
glioma.
Interest in cancer stem cells is rapidly increasing.
In glioma, studies have shown that stem cells could promote
radioresistance by preferential activation of the DNA damage
response (38). Whether PGK1 plays
a potential role in regulating radioresistance of glioma stem cells
remains unclear and further studies are required to clarify this
issue.
Our previous study showed that patients with
radioresistant astrocytomas, compared to patients with
radiosensitive astrocytomas, displayed much higher protein level of
PGK1 (11). However, the results
are a long way from using the levels of PGK1 in human gliomas as a
potential marker for predicting the radioresistance in clinical
application. More glioma cell lines should be included to confirm
the role of PGK1 in the glioma. Further studies using PGK1
neutralization or PGK1 knockout mice are needed to examine the
exact role of PGK1 in the radioresistance of glioma animal models.
Also, the relationship between PGK1 levels and the survival rate
after radiotherapy in glioma patients (radioresistant or
non-radioresistant) should also be investigated prospectively. This
knowledge is necessary before a role of PGK1 in the treatment of
glioma can be envisaged.
Acknowledgements
The present study was supported by the grant
NSFC81172390 from the National Science Foundation of China and the
grant ZKX10021 from the Health Bureau of Nanjing.
References
|
1
|
Cancer incidence in five continents. VIII.
IARC Sci Publ; pp. 1–781. 2002
|
|
2
|
Schwartzbaum JA, Fisher JL, Aldape KD and
Wrensch M: Epidemiology and molecular pathology of glioma. Nat Clin
Pract Neurol. 2:494–503. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maher EA, Furnari FB, Bachoo RM, et al:
Malignant glioma: genetics and biology of a grave matter. Genes
Dev. 15:1311–1333. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Stupp R, Mason WP, van den Bent MJ, et al:
Radiotherapy plus concomitant and adjuvant temozolomide for
glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Krakstad C and Chekenya M: Survival
signalling and apoptosis resistance in glioblastomas: opportunities
for targeted therapeutics. Mol Cancer. 9:1352010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Poonnoose SI and Daniel RT: Radiological
evidence of glioma invasion of the central nervous system along
tracts. Surg Neurol. 54:194–196. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tate MC and Aghi MK: Biology of
angiogenesis and invasion in glioma. Neurotherapeutics. 6:447–457.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Grossman SA and Batara JF: Current
management of glioblastoma multiforme. Semin Oncol. 31:635–644.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Preusser M, de Ribaupierre S, Wohrer A, et
al: Current concepts and management of glioblastoma. Ann Neurol.
70:9–21. 2011. View Article : Google Scholar
|
|
10
|
Zinn PO, Colen RR, Kasper EM and Burkhardt
JK: Extent of resection and radiotherapy in GBM: A 1973 to 2007
surveillance, epidemiology and end results analysis of 21,783
patients. Int J Oncol. 42:929–934. 2013.PubMed/NCBI
|
|
11
|
Yan H, Yang K, Xiao H, Zou YJ, Zhang WB
and Liu HY: Over-expression of cofilin-1 and phosphoglycerate
kinase 1 in astrocytomas involved in pathogenesis of
radioresistance. CNS Neurosci Ther. 18:729–736. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
VandeBerg JL: The phosphoglycerate kinase
isozyme system in mammals: biochemical, genetic, developmental, and
evolutionary aspects. Isozymes Curr Top Biol Med Res. 12:133–187.
1985.PubMed/NCBI
|
|
13
|
Ahmad SS, Glatzle J, Bajaeifer K, et al:
Phosphoglycerate kinase 1 as a promoter of metastasis in colon
cancer. Int J Oncol. 43:586–590. 2013.PubMed/NCBI
|
|
14
|
Chen G, Gharib TG, Wang H, et al: Protein
profiles associated with survival in lung adenocarcinoma. Proc Natl
Acad Sci USA. 100:13537–13542. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zieker D, Konigsrainer I, Tritschler I, et
al: Phosphoglycerate kinase 1 a promoting enzyme for peritoneal
dissemination in gastric cancer. Int J Cancer. 126:1513–1520.
2010.PubMed/NCBI
|
|
16
|
Wang J, Ying G, Jung Y, et al:
Characterization of phosphoglycerate kinase-1 expression of stromal
cells derived from tumor microenvironment in prostate cancer
progression. Cancer Res. 70:471–480. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cecconi D, Palmieri M and Donadelli M:
Proteomics in pancreatic cancer research. Proteomics. 11:816–828.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Duan Z, Lamendola DE, Yusuf RZ, Penson RT,
Preffer FI and Seiden MV: Overexpression of human phosphoglycerate
kinase 1 (PGK1) induces a multidrug resistance phenotype.
Anticancer Res. 22:1933–1941. 2002.PubMed/NCBI
|
|
19
|
Lincet H, Guevel B, Pineau C, et al:
Comparative 2D-DIGE proteomic analysis of ovarian carcinoma cells:
toward a reorientation of biosynthesis pathways associated with
acquired platinum resistance. J Proteomics. 75:1157–1169. 2012.
View Article : Google Scholar
|
|
20
|
Lin TY, Chang JT, Wang HM, et al:
Proteomics of the radioresistant phenotype in head-and-neck cancer:
Gp96 as a novel prediction marker and sensitizing target for
radiotherapy. Int J Radiat Oncol Biol Phys. 78:246–256. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
|
|
22
|
Rich JN and Bigner DD: Development of
novel targeted therapies in the treatment of malignant glioma. Nat
Rev Drug Discov. 3:430–446. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakamizo A, Marini F, Amano T, et al:
Human bone marrow-derived mesenchymal stem cells in the treatment
of gliomas. Cancer Res. 65:3307–3318. 2005.PubMed/NCBI
|
|
24
|
Hwang TL, Liang Y, Chien KY and Yu JS:
Overexpression and elevated serum levels of phosphoglycerate kinase
1 in pancreatic ductal adenocarcinoma. Proteomics. 6:2259–2272.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fantin VR, St-Pierre J and Leder P:
Attenuation of LDH-A expression uncovers a link between glycolysis,
mitochondrial physiology, and tumor maintenance. Cancer Cell.
9:425–434. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brizel DM, Dodge RK, Clough RW and
Dewhirst MW: Oxygenation of head and neck cancer: changes during
radiotherapy and impact on treatment outcome. Radiother Oncol.
53:113–117. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fyles AW, Milosevic M, Wong R, et al:
Oxygenation predicts radiation response and survival in patients
with cervix cancer. Radiother Oncol. 48:149–156. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Knocke TH, Weitmann HD, Feldmann HJ,
Selzer E and Potter R: Intratumoral pO2-measurements as
predictive assay in the treatment of carcinoma of the uterine
cervix. Radiother Oncol. 53:99–104. 1999.
|
|
29
|
Semenza GL: Defining the role of
hypoxia-inducible factor 1 in cancer biology and therapeutics.
Oncogene. 29:625–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fujiwara S, Nakagawa K, Harada H, et al:
Silencing hypoxia-inducible factor-1α inhibits cell migration and
invasion under hypoxic environment in malignant gliomas. Int J
Oncol. 30:793–802. 2007.
|
|
31
|
Semenza GL, Roth PH, Fang HM and Wang GL:
Transcriptional regulation of genes encoding glycolytic enzymes by
hypoxia-inducible factor 1. J Biol Chem. 269:23757–23763.
1994.PubMed/NCBI
|
|
32
|
Lam W, Bussom S and Cheng YC: Effect of
hypoxia on the expression of phosphoglycerate kinase and antitumor
activity of troxacitabine and gemcitabine in non-small cell lung
carcinoma. Mol Cancer Ther. 8:415–423. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim JW and Dang CV: Cancer’s molecular
sweet tooth and the Warburg effect. Cancer Res. 66:8927–8930.
2006.
|
|
35
|
Vander Heiden MG, Cantley LC and Thompson
CB: Understanding the Warburg effect: the metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009.PubMed/NCBI
|
|
36
|
Daly EB, Wind T, Jiang XM, Sun L and Hogg
PJ: Secretion of phosphoglycerate kinase from tumour cells is
controlled by oxygen-sensing hydroxylases. Biochim Biophys Acta.
1691:17–22. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lepleux C, Abeilard-Lemoisson E, Duval M,
Icard P and Lincet H: siPGK1 sensitizes chemoresistant human
ovarian cancer cell lines to cisplatin. Anticancer Res.
32:4277–4286. 2012.PubMed/NCBI
|
|
38
|
Bao S, Wu Q, McLendon RE, et al: Glioma
stem cells promote radioresistance by preferential activation of
the DNA damage response. Nature. 444:756–760. 2006. View Article : Google Scholar : PubMed/NCBI
|