Introduction
Cholangiocarcinoma is a malignant tumor originating
from the bile duct epithelium (1,2).
Complete surgical resection is currently the only curative
treatment for patients with cholangiocarcinoma. However, due to
difficulties in the early diagnosis and its high metastatic
potential, most patients with cholangiocarcinoma have regional or
distant metastasis at the time of clinical diagnosis (3–5). Few
specific diagnostic or therapeutic tools are available because of
the limited information on the molecular pathogenesis of
cholangiocarcinoma. Therefore, new therapeutic targets are urgently
needed.
Epigenetic gene silencing is an important mechanism
that results in loss of gene expression and that mediates, along
with genetic mutations, the initiation and progression of human
cancer. Polycomb group proteins regulate and mediate epigenetic
transcriptional silencing (6,7). They
are involved in the maintenance of embryonic and adult stem cells
and in repression of key tumor-suppressor pathways, which may
contribute to their oncogenic function (8). The enhancer of zeste homolog 2 (EZH2)
is the catalytic subunit of the polycomb repressive complex 2
(PRC2), which also includes the suppressor of zeste 12 (SUZ12)
protein and embryonic ectoderm development (EED) protein. EZH2 acts
as a histone lysine methyltransferase that mediates trimethylation
of lysine 27 of histone H3 (3mH3K27) to silence expression of PRC2
target genes involved in lineage differentiation (9,10).
EZH2 is overexpressed in a variety of malignancies including
prostate cancer, breast cancer, melanoma, uterine cancer, gastric
cancer and renal cell cancer. EZH2 expression levels are correlated
with aggressiveness, metastasis and a poor patient prognosis in
most of these cancers (11–15). EZH2 is barely expressed in normal
tissues of various types (16).
More recently, we found that cholangiocarcinoma also
overexpresses EZH2 and that high expression of EZH2 is correlated
with poor patient prognosis (17).
Furthermore, an activating mutation in EZH2 has been identified in
a subset of B-cell lymphomas (18).
Overexpression of EZH2 enhances aggressiveness in prostatic cancer
cells (19) and produces a
neoplastic phenotype characterized by independent growth and cell
invasion in immortalized mammary epithelial cells and bronchial
epithelial cells (13,20). Conversely, depletion of EZH2 results
in reduced proliferation, increased apoptosis, and inhibition of
tumorigenicity in cancer cells (12,19,21,22)
including cholangiocarcinoma cells (17). These findings indicate that EZH2 may
be an appropriate therapeutic target for various types of
cancers.
A cyclopentenyl analog of 3-deazaadenosine,
3-deazaneplanocin A (DZNep), inhibits the activity of
S-adenosyl-L-homocysteine (AdoHcy) hydrolase, the enzyme
responsible for the reversible hydrolysis of AdoHcy to adenosine
and homocysteine (23). This
inhibition results in the intracellular accumulation of AdoHcy,
which leads to inhibition of S-adenosyl-L-methionine-dependent
lysine methyltransferase activity. Presently, DZNep was shown to
reduce levels of the PRC2 complex, including EZH2, in breast cancer
cells and cause concomitant loss of 3mH3K27 and repression of
epigenetically silenced target genes (24). Moreover, DZNep was found to inhibit
proliferation and promote apoptosis in several types of cancer
cells (24–29), and the synergistic interaction of
DZNep and gemcitabine enhanced the inhibition of cell proliferation
and apoptosis in pancreatic cancer cells (30).
However, data on the activity of DZNep in
cholangiocarcinoma cells are scarce. In the present study, we
demonstrated the effects of DZNep alone and in combination with
gemcitabine on cholangiocarcinoma cells.
Materials and methods
Cell lines and chemicals
The cholangiocarcinoma cell lines RBE and TFK-1 were
purchased from RIKEN Bioresource Center (Ibaraki, Japan). RBE and
TFK-1 cell lines were cultured in RPMI-1640 (Invitrogen, Tokyo,
Japan) containing 10% fetal bovine serum (FBS). All cultures were
maintained in a 5% CO2 air-humidified atmosphere at
37°C. DZNep and gemcitabine were provided by Cayman Chemical Japan
and Eli Lilly Japan, respectively.
Cell growth assay
Cells were seeded in a 96-well plate at a density of
5000 cells per well and treated with or without DZNep (0.5 μM) or
gemcitabine (30 nM) or the combination. Viable cholangiocarcinoma
cell numbers were measured with the Cell Counting Kit-8 (CCK-8)
containing 2-(2-methoxy-4-
nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium
(WST-8) (Dojin Laboratories, Kumamoto, Japan) for 24, 48 and 72 h,
according to the manufacturer’s instructions. OD 450 was measured
using an automatic microplate reader (Molecular Devices, Osaka,
Japan). Each experiment was performed in triplicate.
Flow cytometric analysis of cell cycle
distribution and detection of apoptosis
RBE and TFK-1 cells were seeded in a 12-well plate.
After 72 h of treatment with or without DZNep (0.5 nM) or
gemcitabine (30 nM) or the combination, cells were trypsinized,
adjusted to 5×105/ml, and fixed in 70% ethanol overnight
at −20°C. Subsequently, cells were pelleted and resuspended in
phosphate-buffered saline containing 1 μg/ml RNase A (Sigma, Tokyo,
Japan) and 100 μg/ml propidium iodide (PI) (Sigma). Cell cycle
analyses were performed using a FACS Aria II (BD Biosciences, Tokyo
Japan). The distribution of cells in different cell cycle phases
was calculated using FlowJo software (Tomy Digital Biology Co.,
Ltd., Tokyo, Japan). Phosphatidylserine externalization was
detected by Annexin V staining (Millipore, Tokyo, Japan) following
the manufacturer’s instructions. Annexin V-positive and PI-negative
cells were detected as early apoptotic cells.
Statistical analysis
Statistical analysis was performed using the JMP
program (SAS Institute, Cary, NC, USA). Quantitative data are
expressed as means ± SD, unless otherwise stated. The χ2
test was used to analyze the relationship between categorical
variables. For all statistical analyses, the level of significance
was set at 0.05.
Results
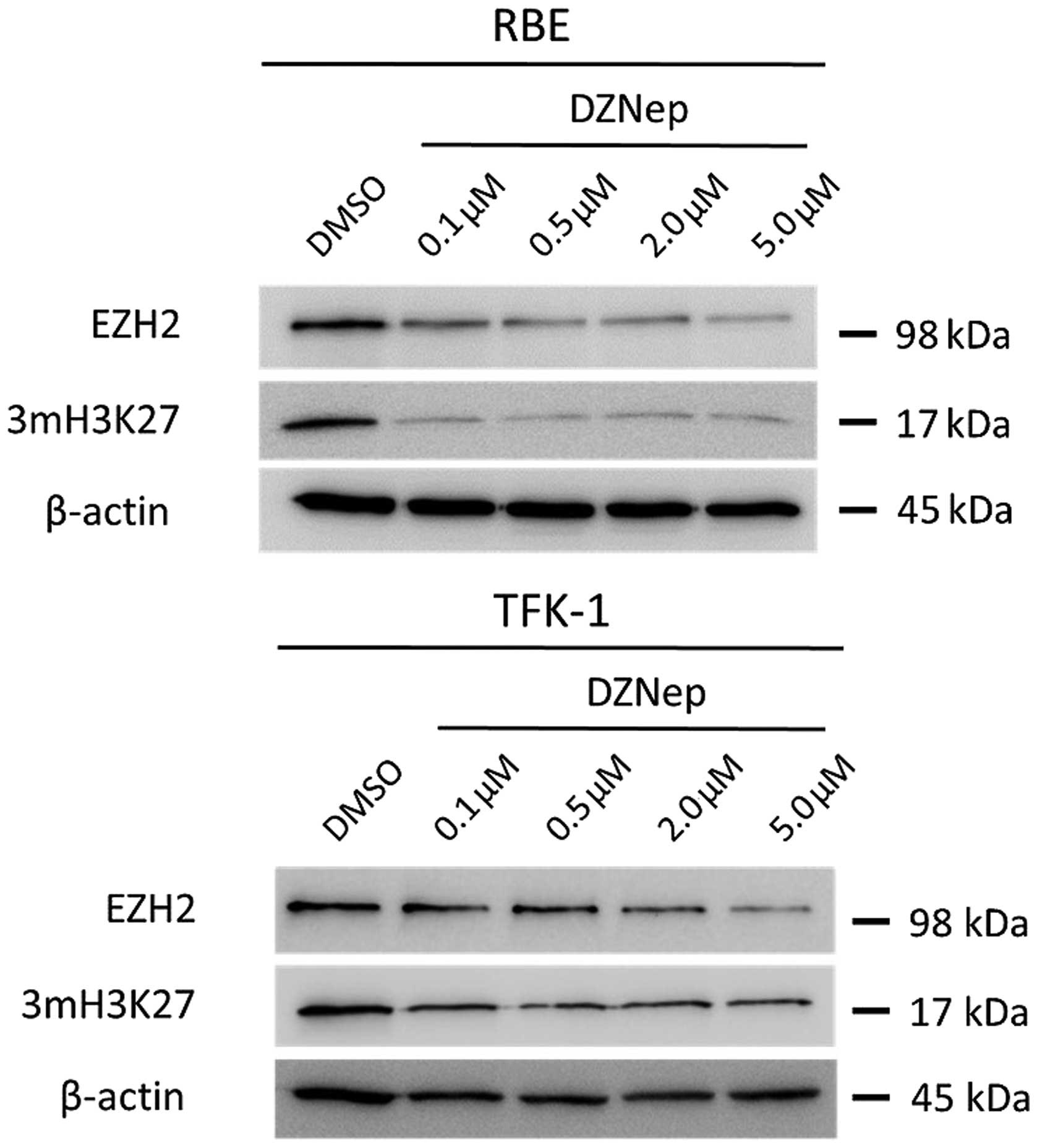
DZNep reduces EZH2 expression and
demethylated H3K27 in cholangiocarcinoma cells
The expression of EZH2 was studied at the protein
level in untreated RBE and TFK-1 cells and in cells treated with
0.1–5 μM DZNep. As shown in Fig. 1,
DZNep reduced the expression of EZH2 in a dose-dependent manner. In
addition, we investigated the expression of the 3mH3K27 protein,
which was also reduced by DZNep in a dose-dependent manner in both
cell lines.
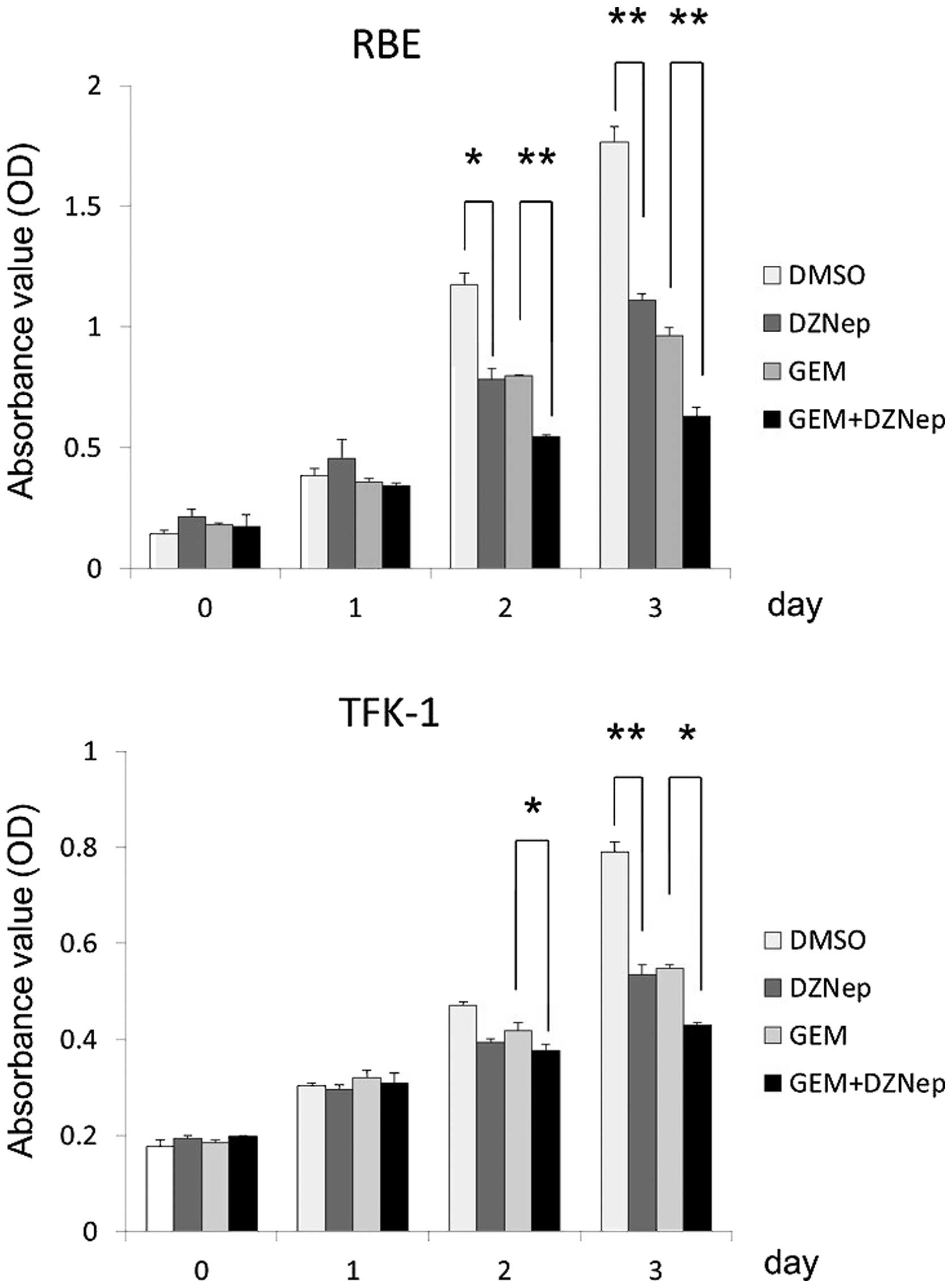
DZNep inhibits the growth of
cholangiocarcinoma cells and interacts synergistically with
gemcitabine
We assessed the effect of DZNep-mediated EZH2
repression on the growth of RBE and TFK-1 cholangiocarcinoma cells
(Fig. 2). The cell growth assay
demonstrated that 0.5 μM DZNep caused significantly inhibited
proliferation of cholangiocarcinoma cells. As compared with control
cells on day 3, DZNep reduced the growth of RBE and TFK-1 cells to
67.0 and 67.6%, respectively. Similarly, 30 nM gemcitabine also
significantly inhibited cell proliferation to 58.4 and 69.5% (in
RBE and TFK-1 cells, respectively) as compared with the control
cells on day 3. The combination of 0.5 μM DZNep and 30 nM
gemcitabine enhanced the antiproliferative activity of gemcitabine.
The combination of DZNep and gemcitabine reduced the proliferation
of RBE and TFK-1 cells to 61.2 and 78.6% compared with gemcitabine
alone, and to 35.7 and 53.1% compared with the control cells
(Fig. 2).
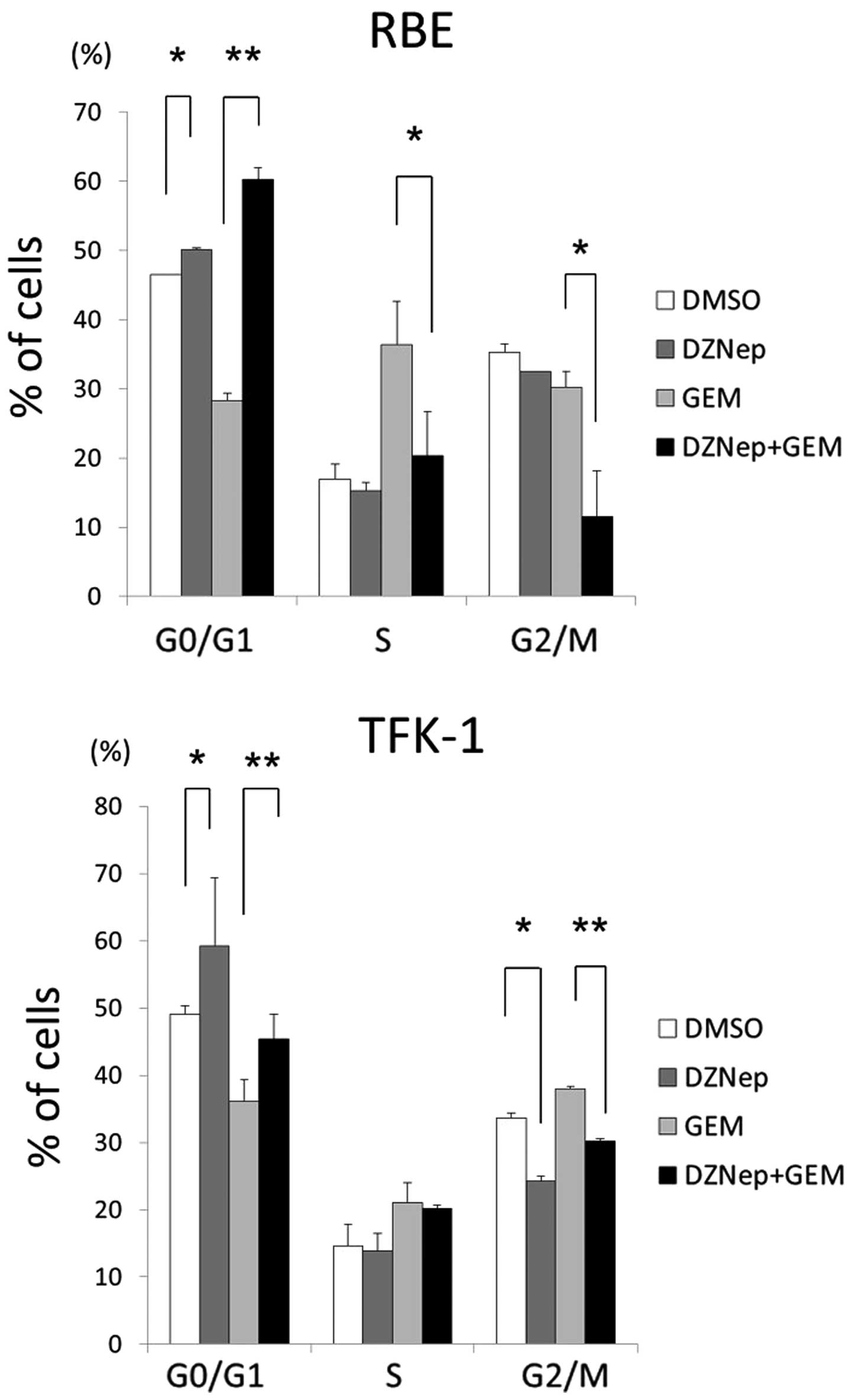
DZNep induces cell cycle arrest and the
combination of DZNep and gemcitabine enhances cell cycle
arrest
We used flow cytometry to determine whether the
reduction in proliferation was due to cell cycle arrest or
apoptosis in the two cholangiocarcinoma cell lines. DZNep alone and
in combination with gemcitabine affected the cell cycle of
cholangiocarcinoma cells (Fig. 3).
In particular, DZNep increased the percentage of RBE and TFK-1
cells in the G0/G1 phase from 46.5 to 50.2% and from 49.1 to 59.2%,
respectively (p<0.05). Gemcitabine alone reduced the number of
cells in the G0/G1 phase to 28.3 and 36.1%, respectively
(p<0.05). The combination of DZNep and gemcitabine significantly
increased the percentage of RBE and TFK-1 cells in the G0/G1 phase
compared with gemcitabine alone (from 28.3 to 60.2% and from 36.1
to 45.4%, respectively). Additionally, DZNep reduced the percentage
of cells in the S and G2/M phases, and the combination of DZNep and
gemcitabine significantly reduced the percentage of cells in the S
and G2/M phases compared with gemcitabine alone.
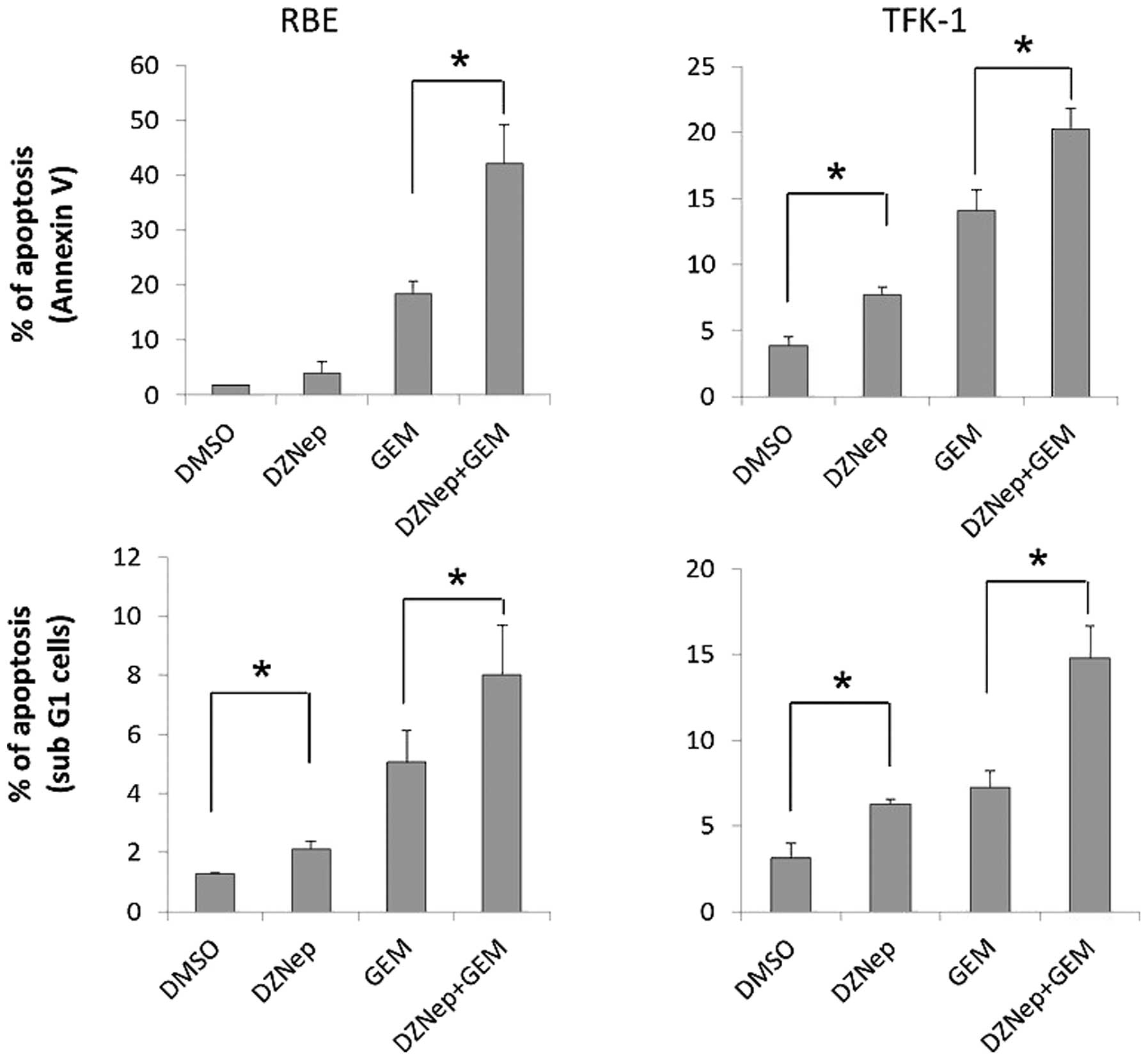
DZNep induces cell apoptosis and the
combination of DZNep and gemcitabine enhances apoptosis
Flow cytometry using Annexin V and PI staining, and
analysis of the sub-G1 region demonstrated that DZNep or
gemcitabine alone significantly enhanced apoptosis when compared
with the control cells (Fig. 4).
Treatment with 0.5 μM DZNep increased apoptosis 2-fold, and
treatment with 30 nM gemcitabine resulted in ~5-fold increase in
apoptosis. Furthermore, the combination of DZNep and gemcitabine
significantly increased the number of apoptotic cells when compared
with either agent alone (~5-fold and 2-fold increases in the RBE
and TFK-1 cells, respectively). We also investigated expression of
cleaved caspase-3 and cleaved polyADP ribose polymerase (PARP) as
markers of apoptosis to confirm the apoptotic effect of the
combination of DZNep and gemcitabine. A slight increase in these
proteins was noted followed treatment with DZNep alone, but the
combination of DZNep and gemcitabine resulted in a significant
increase in apoptotic proteins in the RBE and TFK-1 cells (Fig. 5). These results confirmed the flow
cytometric analysis.
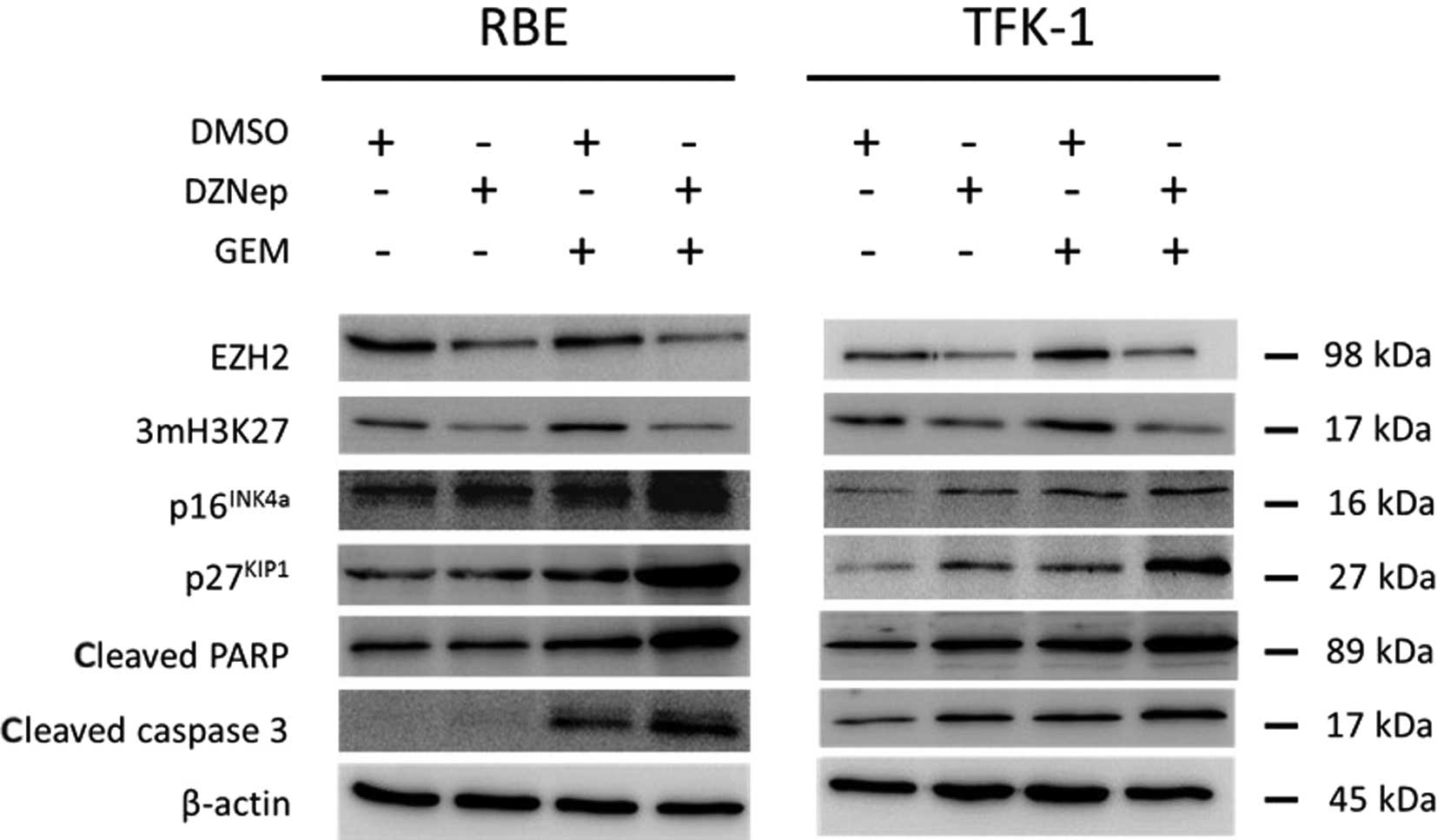
DZNep depletes EZH2 and 3mH3K27 and
increases p16INK4a and p27KIP1 in
cholangiocarcinoma cells and the combination of DZNep and
gemcitabine enhances these effects
DZNep reduced the expression of EZH2 protein and
3mH3K27 in cholangiocarcinoma cell lines (Fig. 5). The combination of DZNep and
gemcitabine also reduced the expression of EZH2 and 3mH3K27 when
compared with gemcitabine alone. We determined the effect of DZNep
on EZH2 target genes such as p16INK4a and
p27KIP1. Expression of p16INK4a and
p27KIP1 was increased by DZNep. The combination of DZNep
and gemcitabine enhanced the increase in expression of
p16INK4a and p27KIP1.
Discussion
The present study demonstrated that the combination
of the EZH2 inhibitor DZNep and the cytotoxic compound gemcitabine
exhibited strong synergistic effects on the cholangiocarcinoma cell
lines. EZH2 has a master regulatory role in the fate of native
embryogenic cells (31), as well as
in cancer development via methylation-mediated repression of the
transcription of several genes (32,33).
Overexpression of EZH2 is a marker of advanced and metastatic
disease in many solid tumors, including cholangiocarcinoma
(12,13,34).
EZH2 is crucial for regulating the cell cycle by repressing several
tumor-suppressor genes such as p16INK4a,
p27KIP1 and RUNX3, and by mediating the pRb pathway. In
addition, EZH2 regulates E2F1-dependent apoptosis directly in
cancer cells, through modulation of Bim expression.
Our previous study on RBE and TFK-1 cells showed
that suppression of EZH2 expression by RNA interference with siRNA
targeting EZH2 markedly inhibited cellular proliferation in
vitro, and drastically induced both cell cycle arrest and
apoptosis (17,35). Knockdown of EZH2 by siRNA increased
expression of p16INK4a and
p27KIP1, which are considered to be the
targets of EZH2 (17). According to
other research, transfection of shEZH2 construct cells sensitized
MIA-PaCa-2 and Pac04.02 cells to doxorubicin and gemcitabine in
pancreatic cancer (21), suggesting
that the combination of EZH2 inhibitors and gemcitabine might
overcome the intrinsic chemoresistance of cholangiocarcinoma. To
the best of our knowledge, this is the first study evaluating the
pharmacological interaction of the small-molecule EZH2 inhibitor
DZNep with gemcitabine in cholangiocarcinoma cells.
In the cholangiocarcinoma cell lines, we observed a
significant reduction of both EZH2 and 3mH3K27 expression in a
dose-dependent manner. Analyses of the cell cycle distribution and
apoptosis indicated that the growth suppression by DZNep was
associated with G1 cell cycle arrest and increased apoptosis in
cholangiocarcinoma cells. This conclusion is consistent with the
findings from acute myeloid leukemia (AML) cells treated with DZNep
(27). Knockdown of EZH2 by siRNA
has also been shown to induce G1 cell cycle arrest and to increase
apoptosis in cholangiocarcinoma cells and other types of carcinoma
cells (36). DZNep induced
accumulation of p16INK4a and p27KIP1 in RBE
and TFK-1 cells in the present study, and this may have resulted in
G1 arrest and increased apoptosis. An association between either
p16INK4a or p27KIP1 accumulation and G1 cell
cycle arrest and apoptosis has been shown in various types of cells
(37,38). Fiskus et al reported that
DZNep treatment induced p16INK4a, p21CIP1,
p27KIP1 and FBXO32 while reducing cyclin E and HOXA9
levels in human AML cells (27).
Taken together, these results indicate that growth suppression by
DZNep is associated with G1 phase arrest and apoptosis in
cholangiocarcinoma cells, partly via p16INK4a and
p27KIP1 accumulation.
The Advanced Biliary Cancer (ABC)-02 study showed
that systemic combination chemotherapy of gemcitabine and cisplatin
prolonged the survival of patients with inoperable
cholangiocarcinoma, making it a treatment standard, and gemcitabine
is still the key drug used in chemotherapy of advanced
cholangiocarcinoma (39,40). In the present study, we demonstrated
that the combination of DZNep and gemcitabine has a synergistic
effect on two representative cholangiocarcinoma cell lines, RBE and
TFK-1. This synergistic interaction against cell proliferation was
associated with a significant increase in apoptosis induction and
cell cycle arrest. The combination of gemcitabine and DZNep
increased the expression of p16INK4a and
p27KIP1 when compared with gemcitabine alone, so that
the synergistic effect might be partly explained by the significant
upregulation of p16INK4a and p27KIP1.
In conclusion, EZH2 inhibitor DZNep appears to be a
promising anticancer agent, by attacking key mechanisms involved in
the proliferation, cell cycle control and apoptosis of
cholangiocarcinoma cells. Moreover, the favorable modulation of
p16INK4a and p27KIP1 makes DZNep an optimal
candidate for combination therapy with gemcitabine. The synergistic
effects observed in the present study may have critical
implications for the rational development of innovative regimens
consisting of DZNep and gemcitabine to improve the effectiveness of
the treatment of cholangiocarcinoma.
References
|
1
|
Alpini G, McGill JM and Larusso NF: The
pathobiology of biliary epithelia. Hepatology. 35:1256–1268. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roberts SK, Ludwig J and Larusso NF: The
pathobiology of biliary epithelia. Gastroenterology. 112:269–279.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Welzel TM, Graubard BI, El-Serag HB, et
al: Risk factors for intrahepatic and extrahepatic
cholangiocarcinoma in the United States: a population-based
case-control study. Clin Gastroenterol Hepatol. 5:1221–1228. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shaib YH, El-Serag HB, Nooka AK, et al:
Risk factors for intrahepatic and extrahepatic cholangiocarcinoma:
a hospital-based case-control study. Am J Gastroenterol.
102:1016–1021. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tao LY, He XD, Qu Q, et al: Risk factors
for intrahepatic and extrahepatic cholangiocarcinoma: a
case-control study in China. Liver Int. 30:215–221. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ringrose L and Paro R: Epigenetic
regulation of cellular memory by the Polycomb and Trithorax group
proteins. Annu Rev Genet. 38:413–443. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sakamoto Y, Watanabe S, Ichimura T, et al:
Overlapping roles of the methylated DNA-binding protein MBD1 and
polycomb group proteins in transcriptional repression of HOXA genes
and heterochromatin foci formation. J Biol Chem. 282:16391–16400.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sparmann A and van Lohuizen M: Polycomb
silencers control cell fate, development and cancer. Nat Rev
Cancer. 6:846–856. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cao R and Zhang Y: The functions of
E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr
Opin Genet Dev. 14:155–164. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simon JA and Lange CA: Roles of the EZH2
histone methyltransferase in cancer epigenetics. Mutat Res.
647:21–29. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bachmann IM, Halvorsen OJ, Collett K, et
al: EZH2 expression is associated with high proliferation rate and
aggressive tumor subgroups in cutaneous melanoma and cancers of the
endometrium, prostate, and breast. J Clin Oncol. 24:268–273. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Varambally S, Dhanasekaran SM, Zhou M, et
al: The polycomb group protein EZH2 is involved in progression of
prostate cancer. Nature. 419:624–629. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kleer CG, Cao Q, Varambally S, et al: EZH2
is a marker of aggressive breast cancer and promotes neoplastic
transformation of breast epithelial cells. Proc Natl Acad Sci USA.
100:11606–11611. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsukawa Y, Semba S, Kato H, Ito A,
Yanagihara K and Yokozaki H: Expression of the enhancer of zeste
homolog 2 is correlated with poor prognosis in human gastric
cancer. Cancer Sci. 97:484–491. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wagener N, Macher-Goeppinger S, Pritsch M,
et al: Enhancer of zeste homolog 2 (EZH2) expression is an
independent prognostic factor in renal cell carcinoma. BMC Cancer.
10:5242010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takawa M, Masuda K, Kunizaki M, et al:
Validation of the histone methyltransferase EZH2 as a therapeutic
target for various types of human cancer and as a prognostic
marker. Cancer Sci. 102:1298–1305. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakagawa S, Okabe H, Sakamoto Y, et al:
Enhancer of zeste homolog 2 (EZH2) promotes progression of
cholangiocarcinoma cells by regulating cell cycle and apoptosis.
Ann Surg Oncol. Jul 26–2013.(Epub ahead of print).
|
|
18
|
McCabe MT, Graves AP, Ganji G, et al:
Mutation of A677 in histone methyltransferase EZH2 in human B-cell
lymphoma promotes hypertrimethylation of histone H3 on lysine 27
(H3K27). Proc Natl Acad Sci USA. 109:2989–2994. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Karanikolas BD, Figueiredo ML and Wu L:
Comprehensive evaluation of the role of EZH2 in the growth,
invasion, and aggression of a panel of prostate cancer cell lines.
Prostate. 70:675–688. 2010.PubMed/NCBI
|
|
20
|
Watanabe H, Soejima K, Yasuda H, et al:
Deregulation of histone lysine methyltransferases contributes to
oncogenic transformation of human bronchoepithelial cells. Cancer
Cell Int. 8:152008. View Article : Google Scholar
|
|
21
|
Ougolkov AV, Bilim VN and Billadeau DD:
Regulation of pancreatic tumor cell proliferation and
chemoresistance by the histone methyltransferase enhancer of zeste
homologue 2. Clin Cancer Res. 14:6790–6796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wagener N, Holland D, Bulkescher J, et al:
The enhancer of zeste homolog 2 gene contributes to cell
proliferation and apoptosis resistance in renal cell carcinoma
cells. Int J Cancer. 123:1545–1550. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Glazer RI, Hartman KD, Knode MC, et al:
3-Deazaneplanocin: a new and potent inhibitor of
S-adenosylhomocysteine hydrolase and its effects on human
promyelocytic leukemia cell line HL-60. Biochem Biophys Res Commun.
135:688–694. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tan J, Yang X, Zhuang L, et al:
Pharmacologic disruption of Polycomb-repressive complex 2-mediated
gene repression selectively induces apoptosis in cancer cells.
Genes Dev. 21:1050–1063. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hayden A, Johnson PW, Packham G and Crabb
SJ: S-adenosylhomocysteine hydrolase inhibition by
3-deazaneplanocin A analogues induces anti-cancer effects in breast
cancer cell lines and synergy with both histone deacetylase and
HER2 inhibition. Breast Cancer Res Treat. 127:109–119. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Miranda TB, Cortez CC, Yoo CB, et al:
DZNep is a global histone methylation inhibitor that reactivates
developmental genes not silenced by DNA methylation. Mol Cancer
Ther. 8:1579–1588. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fiskus W, Wang Y, Sreekumar A, et al:
Combined epigenetic therapy with the histone methyltransferase EZH2
inhibitor 3-deazaneplanocin A and the histone deacetylase inhibitor
panobinostat against human AML cells. Blood. 114:2733–2743. 2009.
View Article : Google Scholar
|
|
28
|
Jiang X, Tan J, Li J, et al: DACT3 is an
epigenetic regulator of Wnt/beta-catenin signaling in colorectal
cancer and is a therapeutic target of histone modifications. Cancer
Cell. 13:529–541. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suva ML, Riggi N, Janiszewska M, et al:
EZH2 is essential for glioblastoma cancer stem cell maintenance.
Cancer Res. 69:9211–9218. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Avan A, Crea F, Paolicchi E, et al:
Molecular mechanisms involved in the synergistic interaction of the
EZH2 inhibitor 3-deazaneplanocin A with gemcitabine in pancreatic
cancer cells. Molecular Cancer Ther. 11:1735–1746. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xu CR, Cole PA, Meyers DJ, Kormish J, Dent
S and Zaret KS: Chromatin ‘prepattern’ and histone modifiers in a
fate choice for liver and pancreas. Science. 332:963–966. 2011.
|
|
32
|
Chang CJ, Yang JY, Xia W, et al: EZH2
promotes expansion of breast tumor initiating cells through
activation of RAF1-beta-catenin signaling. Cancer Cell. 19:86–100.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chase A and Cross NC: Aberrations of EZH2
in cancer. Clin Cancer Res. 17:2613–2618. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Toll AD, Dasgupta A, Potoczek M, et al:
Implications of enhancer of zeste homologue 2 expression in
pancreatic ductal adenocarcinoma. Human Pathol. 41:1205–1209. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen Y, Xie D, Yin Li W, et al: RNAi
targeting EZH2 inhibits tumor growth and liver metastasis of
pancreatic cancer in vivo. Cancer Lett. 297:109–116. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kikuchi J, Takashina T, Kinoshita I, et
al: Epigenetic therapy with 3-deazaneplanocin A, an inhibitor of
the histone methyltransferase EZH2, inhibits growth of non-small
cell lung cancer cells. Lung Cancer. 78:138–143. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Resnitzky D, Hengst L and Reed SI: Cyclin
A-associated kinase activity is rate limiting for entrance into S
phase and is negatively regulated in G1 by p27Kip1. Mol Cell Biol.
15:4347–4352. 1995.PubMed/NCBI
|
|
38
|
Girard F, Strausfeld U, Fernandez A and
Lamb NJ: Cyclin A is required for the onset of DNA replication in
mammalian fibroblasts. Cell. 2067:1169–1179. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Faris JE and Zhu AX: Targeted therapy for
biliary tract cancers. J Hepatobiliary Pancreat Sci. 19:326–336.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Thomas MB: Biological characteristics of
cancers in the gallbladder and biliary tract and targeted therapy.
Crit Rev Oncol Hematol. 61:44–51. 2007. View Article : Google Scholar : PubMed/NCBI
|