Introduction
OSF is a chronic insidious disease which is
predominantly found in individuals of Asian descent (1,2). OSF
is predisposing to cancer, and the rate of carcinoma incidence in
OSF cases reaches to 7.6% (3). The
hallmark of this disease is submucosal progressive fibrosis that
affects the upper digestive tract involving the oral cavity,
oropharynx and the upper third of the esophagus. Histologically,
most OSF cases are characterized by epithelial atrophy and
accumulation of collagen fibers in the lamina propria while a
minority is characterized by epithelial atypical hyperplasia and
progressive loss of vascularity. The majority of patients are
intolerant to spicy foods, and experience a burning sensation in
the mouth, xerostomia, and limitation of mouth opening, swallowing
and tongue movement (4–8). Epidemiologic studies have indicated
that betel quid (BQ) chewing is one of the major risk factors of
OSF and oral squamous cell carcinoma (OSCC) (9–12).
Arecoline, the most abundant areca alkaloid, has been suggested as
a possible carcinogen which was found to be cytotoxic and genotoxic
in several types of cells (13–17).
However, the mechanisms which are responsible for arecoline-induced
OSF and carcinogenesis are not fully known.
Apoptosis is one of the major forms of programmed
cell death which is dependent on caspase activity (18). This death program can be elicited by
different extracellular or intracellular stimuli that activate
common downstream cell-death machinery. The extrinsic pathway, also
known as the direct, or type I pathway, is triggered by the
ligand-induced activation of cell membrane-anchored death
receptors, and finally induces caspase-8 cleavage and activation
(19). The intrinsic pathway is
activated by cellular damage in which the mitochondrion plays a
central role and eventually activates caspase-9 (20–22).
Both cleaved-caspase-8 and caspase-9 further regulate other caspase
members, including caspase-3 and caspase-7, to initiate a caspase
cascade leading to apoptosis (23).
Bid was first cloned in 1996 as a novel death agonist that
heterodimerizes with either agonists (Bax) or antagonists (Bcl-2)
(24). Bid is localized in the
cytosolic fraction of cells as an inactive precursor. Caspase-8
cleaves Bid at aspartic acid residue 60 (Asp60), leading to the
release of a truncated form containing the carboxy-terminal part of
the protein (25). Cleaved Bid
(tBid) translocates to mitochondria and initiates mitochondrial
outer membrane permeabilization (MOMP), and eventually induces
cytochrome c release and mitochondrial damage (26,27).
Thus, Bid relays an apoptotic signal from the cell surface to the
mitochondria.
In the present study, we investigated the effects of
arecoline on the HaCaT epithelial and Hel fibroblast cell lines.
First, human keratinocyte cells of the HaCaT cell line and human
embryo lung fibroblasts of the Hel cell line were exposed to high
doses of arecoline for a short time to observe the survival rate
and apoptosis. Secondly, the molecular mechanism of
arecoline-induced HaCaT cell apoptosis was studied.
Materials and methods
Reagents and antibodies
Arecoline was purchased from Sigma (St. Louis, MO,
USA). Antibodies to detect cleaved-PARP (9541), caspase-3 (9665)
and Bid (2002) were purchased from Cell Signaling Technology, Inc.
(Beverly, MA, USA). The antibody against β-actin was purchased from
Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture
Keratinocytes of the naturally immortalized normal
cell line, HaCaT, and the human embryonic lung fibroblast cell line
(human normal fibroblast cell line), Hel, were kindly provided by
the College of Life Science, Hunan Normal University, China. HaCaT
and Hel cells were grown in Dulbecco’s modified Eagle’s medium
supplemented with 10% FBS (Invitrogen, Carlsbad, CA, USA), 100 U/ml
penicillin, and 100 mg/ml streptomycin and cultured at 37°C in a
humidified incubator with 5% CO2.
Cell proliferation assay
Cells were seeded (5×103/well) in 96-well
plates and incubated for 24 h and then exposed to different doses
of arecoline. After incubation for 12, 24, 48 or 72 h, 20 μl of
CellTiter 96 Aqueous One Solution (Promega, Madison, WI, USA) was
added, and then cells were incubated for 1 h at 37°C in a 5%
CO2 incubator. Absorbance was measured at 492 nm.
Apoptosis and flow cytometric
analysis
Cells (2×105) were seeded into 6-well
plates and treated without or with different concentrations of
arecoline for 24 h. Cells were trypsinized and washed twice with
cold PBS and then resuspended in 1X Binding Buffer containing
Annexin V-fluorescein isothiocyanate (FITC) and propidium iodide
according to the supplier’s instructions (BD Biosciences, San Jose,
CA, USA). The cells were incubated for 15 min at room temperature
(20–25°C) in the dark and assessed by flow cytometry using a BD
FACS Calibur cytometer as soon as possible (within 1 h). Each
sample was run twice.
Western blot analysis
Cells were harvested by trypsinization and pelleted
by centrifugation. Cell pellets were lysed in NP-40 lysis buffer
supplemented with protease inhibitors. Protein concentrations were
determined using the Bradford assay (Bio-Rad Laboratories,
Philadelphia, PA, USA). Proteins were separated by SDS-PAGE and
electrically transferred to polyvinylidene difluoride membranes
(Millipore, Billerica, MA, USA). After blocking in 5% non-fat dry
milk in TBS, the membranes were hybridized to specific primary
antibodies overnight at 4°C, washed three times with TBS Tween-20,
and then incubated with secondary antibodies conjugated with
horseradish peroxidase for 1 h at room temperature. Next, the
membranes were washed three times in TBS Tween-20 at room
temperature. The protein bands were visualized using ECL
chemiluminescence reagents (Pierce Chemical Co., Rockford, IL, USA)
according to the manufacturer’s protocol.
Statistical analysis
All the statistical analyses were performed using
SPSS software (version 13.0). The experiments were performed in
triplicate. The quantitative data are expressed as mean values ±
standard deviation. The significant differences between two groups
were assessed by a two-tailed Student’s t-test. P<0.05 was
considered to represent a statistically significant difference.
Results
Effect of arecoline on HaCaT and Hel cell
morphology
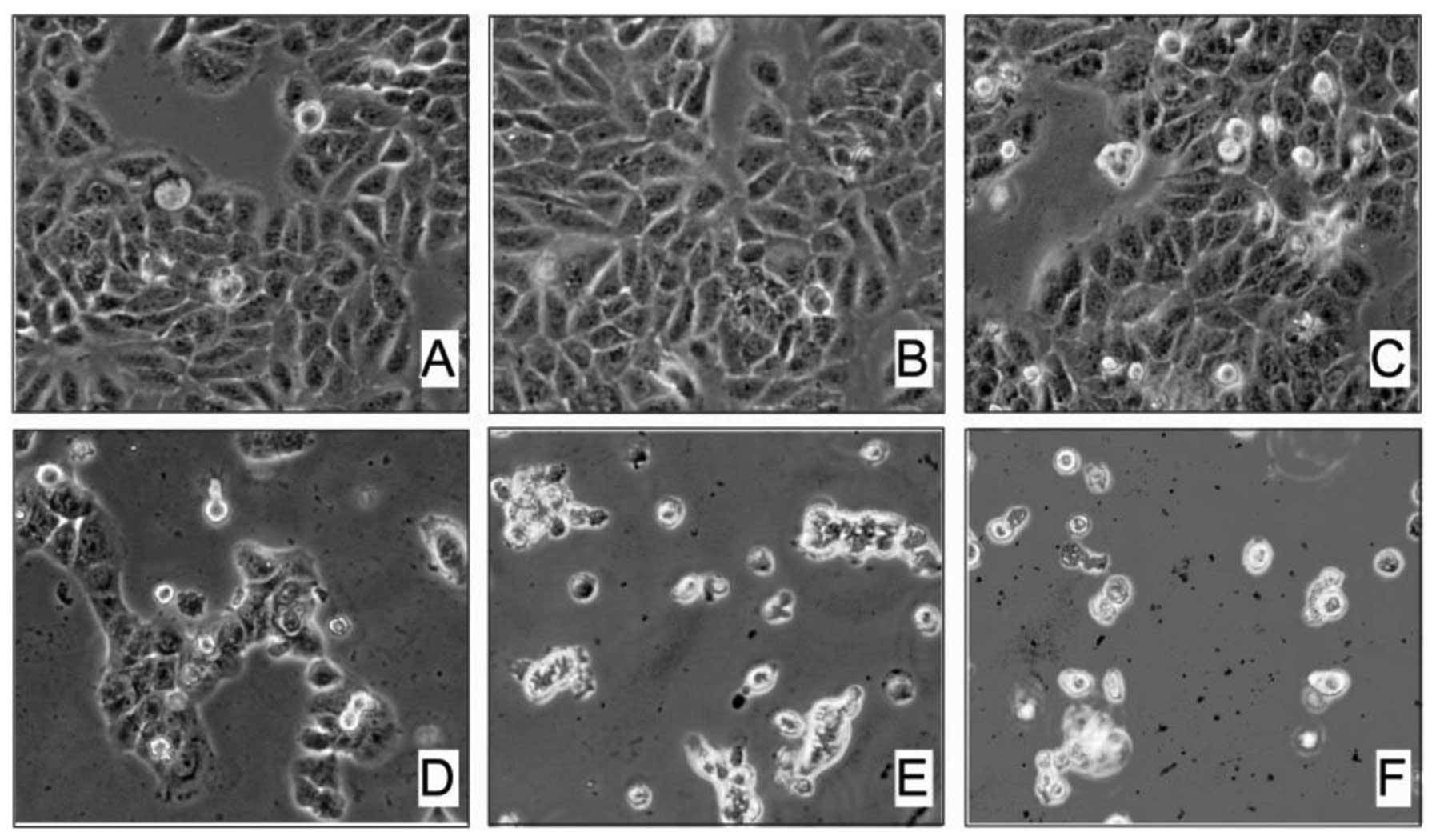
In the present study, we first examined the effect
of arecoline on HaCaT and Hel cell morphology. Exposure of HaCaT
cells to different concentrations of arecoline for 48 h markedly
induced changes in cell morphology. Our results showed that 0.1 mM
arecoline had no obvious cytotoxic effect on HaCaT cells (Fig. 1A and B). However, after we exposed
HaCaT cells to higher concentrations of arecoline from 0.2 to 0.5
mM for 48 h, we found that a significant proportion of cells became
detachment and round in shape. The number of rounded cells
increased proportionally to the arecoline concentration, suggesting
a dose-dependent effect of arecoline on the HaCaT cells.
Additionally, high concentrations of arecoline directly induced
HaCaT cell death. There was massive cell death and a lack of
margins between the cells when the concentration of arecoline was
increased to 0.3, 0.4 and 0.5 mM (Fig.
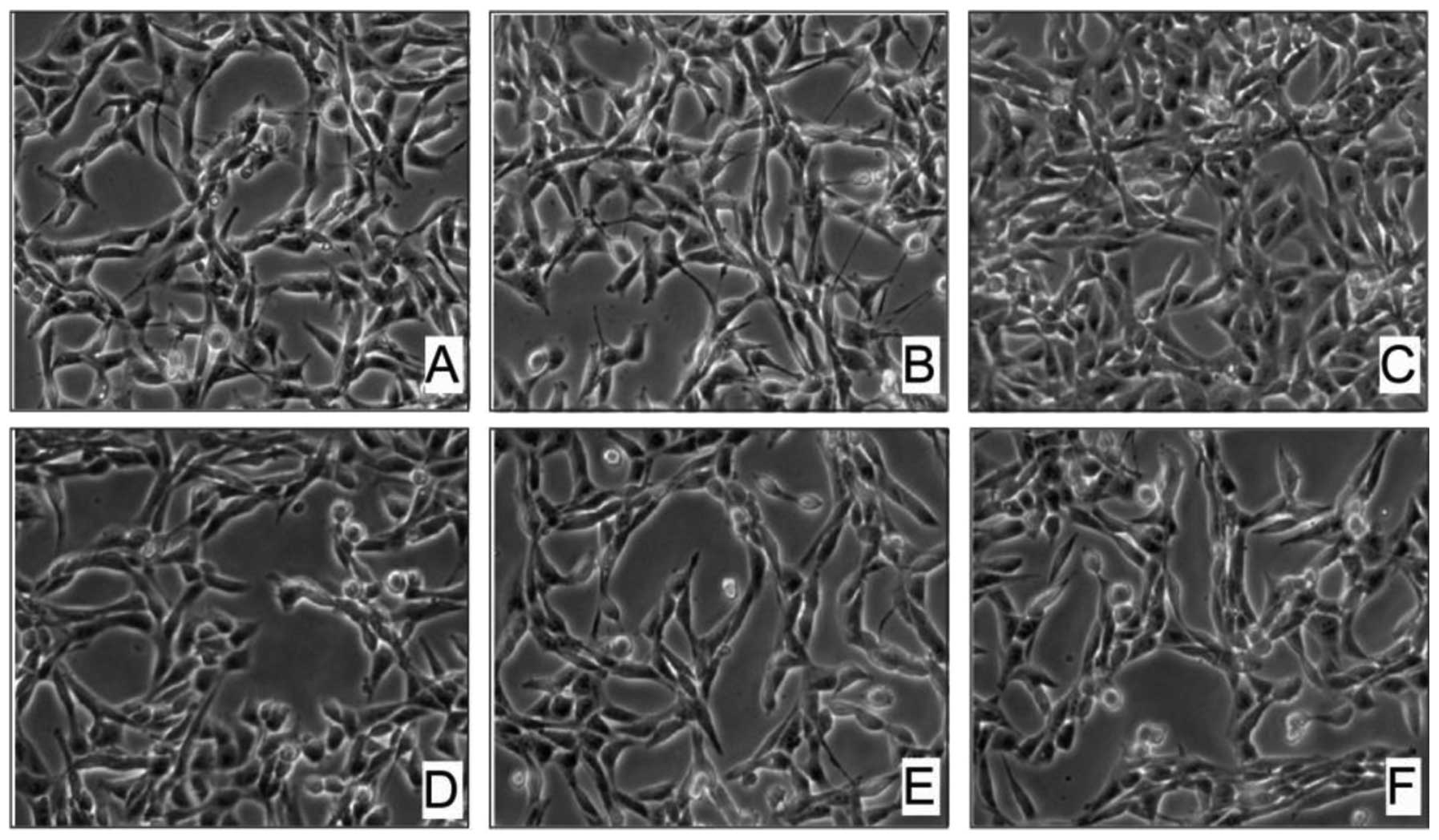
1C–F). In contrast, the Hel cells were less affected by
arecoline exposure. Even when the concentration of arecoline was
increased to 0.4 and 0.5 mM, the Hel cells were still alive and the
cell margins were bright and compact; the morphology began to
change slightly (Fig. 2). Taken
together, these data indicate that arecoline had a dose-dependent
effect on HaCaT cell morphology but not on Hel cell morphology.
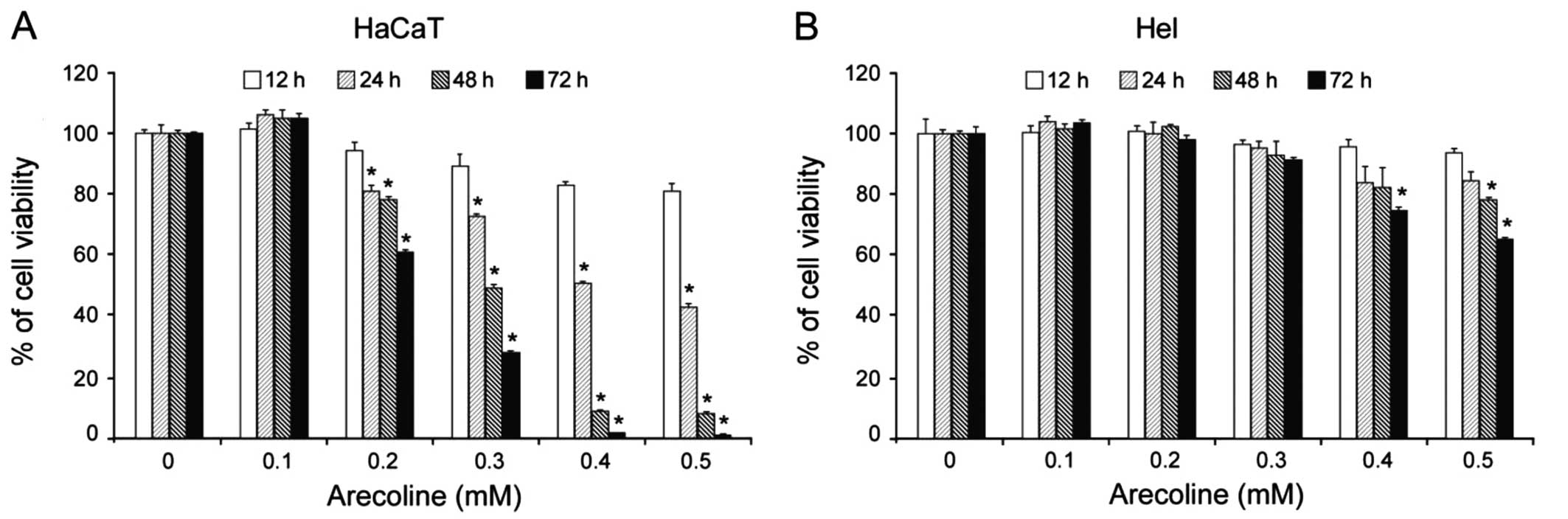
Arecoline inhibits the proliferation of
HaCaT cells
Based on our results, we next determined the effect
of arecoline on HaCaT and Hel cell growth. As expected, incubation
with arecoline significantly reduced the viability of HaCaT cells
in a dose- and time-dependent manner. Exposure to 0.2 mM arecoline
for 24 h began to inhibit HaCaT cell proliferation. Moreover,
exposure to relatively high concentrations (0.4 and 0.5 mM) of
arecoline for 72 h inhibited the viability of almost all of the
HaCaT cells (Fig. 3A). However,
arecoline had no such extensive effect on Hel cells. Only high
concentrations (0.4 and 0.5 mM) of arecoline reduced the viability
of Hel cells (Fig. 3B).
Collectively, these results suggest that arecoline inhibits the
proliferation of HaCaT epithelial cells, but not Hel
fibroblasts.
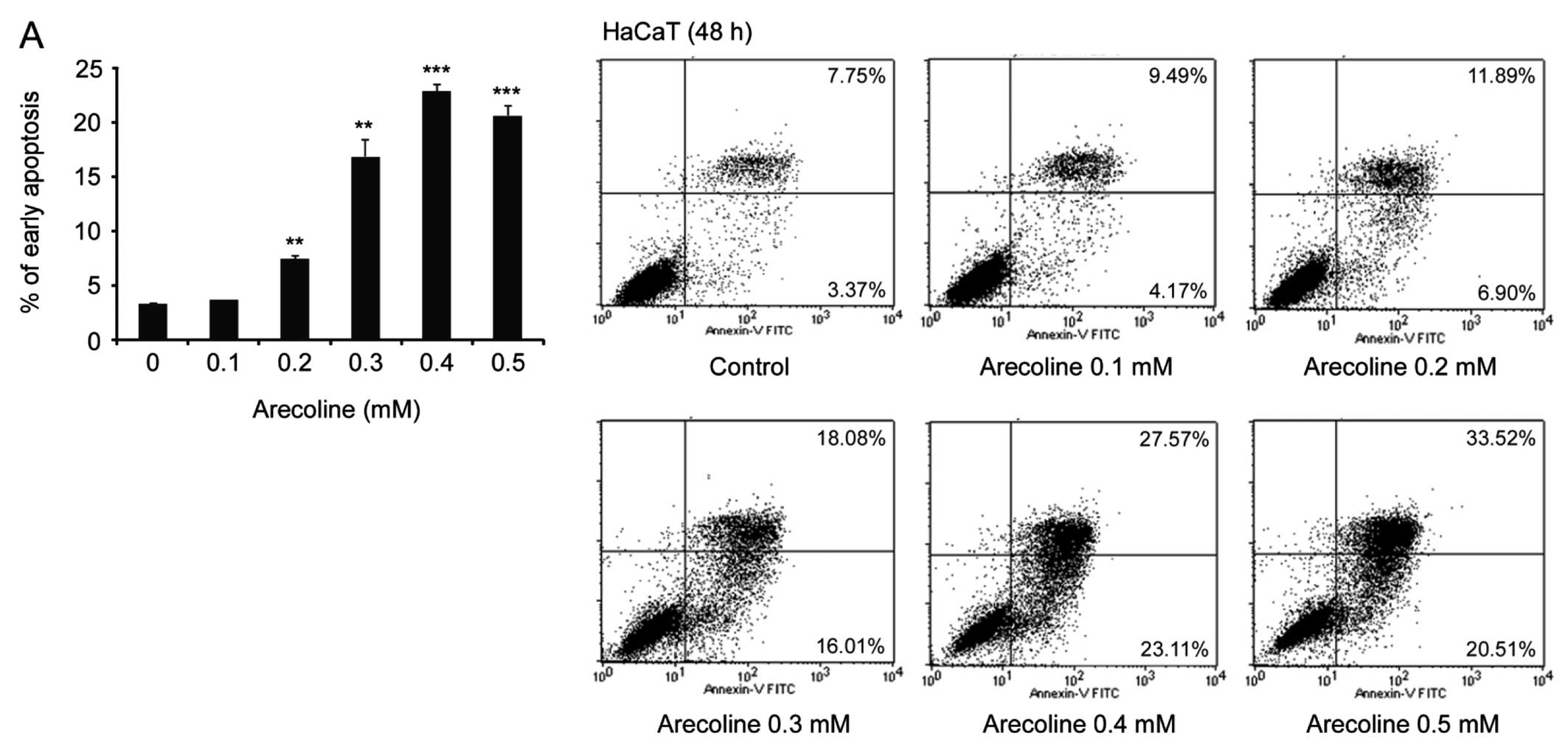
Arecoline induces HaCaT cell
apoptosis
To further investigate whether arecoline-induced
HaCaT cell morphological change is due to apoptosis, we monitored
the impact of different concentrations of arecoline on cell
apoptosis by flow cytometric analysis. Our results demonstrated
that exposure to 0.1 mM arecoline did not result in HaCaT cell
apoptosis, consistent with our previous data that a low
concentration of arecoline had no obvious effect on HaCaT cell
morphology. However, in comparison to the control cells without
exposure to arecoline, exposure to ≥0.2 mM arecoline markedly
induced HaCaT cell apoptosis (Fig.
4A). Of note, Hel cells displayed much more tolerance to high
levels of arecoline-mediated cytotoxicity, since arecoline did not
significantly induce Hel cell apoptosis (Fig. 4B). These results indicate that HaCaT
epithelial cells are much more sensitive to arecoline induced
apoptosis than Hel fibroblasts in vitro.
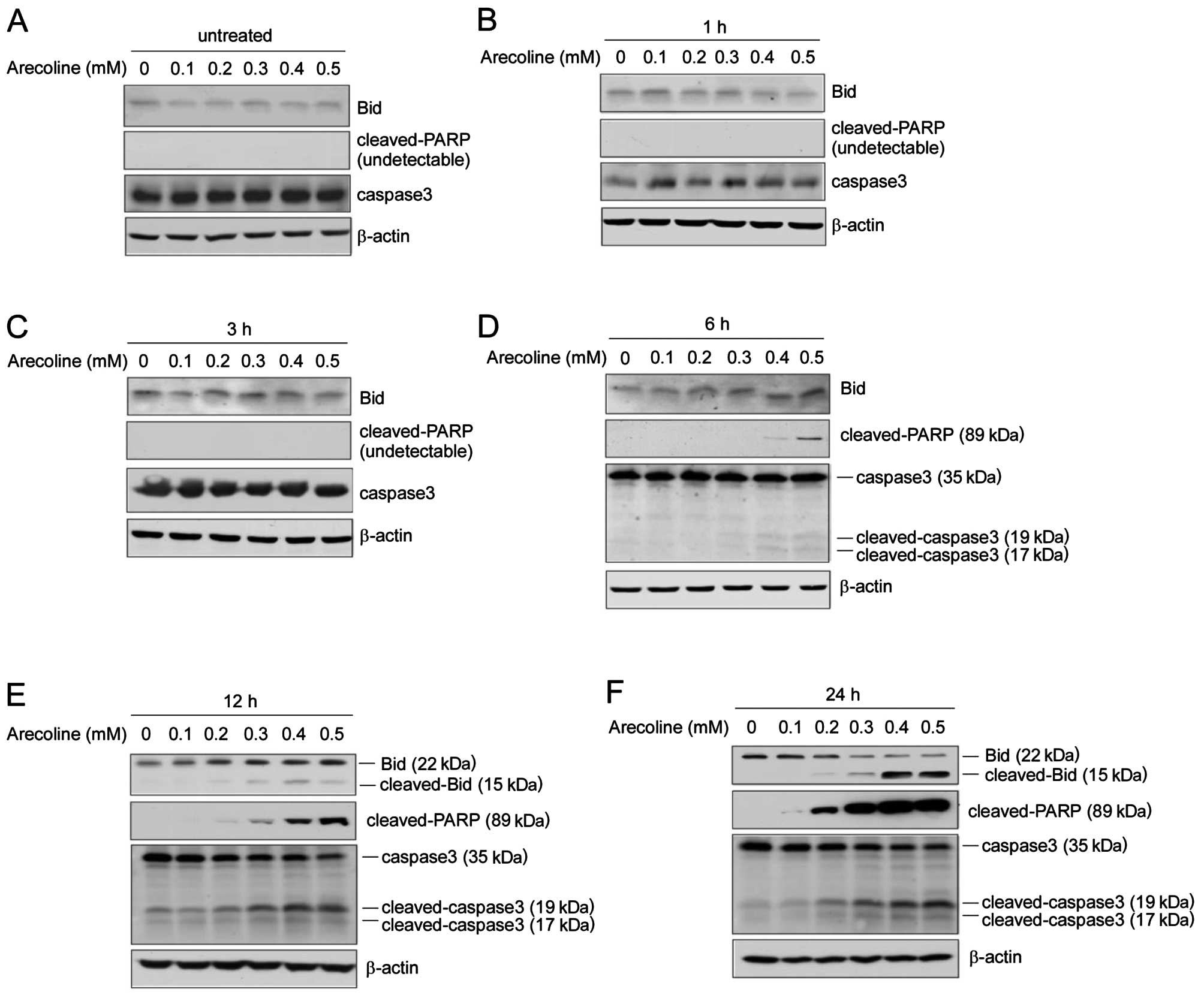
Arecoline induces apoptosis-related
protein expression and caspase activity
Caspase-3, a member of the caspase family, is
expressed in cells as an inactive 32-kDa proenzyme, procaspase-3.
During apoptosis, procaspase-3 is activated by cleavage at specific
Asp residues to generate active caspase-3, consisting of 17- and
19-kDa subunits. Caspase-3 then cleaves its substrate,
poly(ADP-ribose) polymerase (PARP), into 89- and 24-kDa fragments.
The BH3 domain-only protein, Bid, is activated upon proteolytic
cleavage by caspase-8 and relays an apoptotic signal from the cell
surface to mitochondria. We further detected the expression of
caspase-3, cleaved-PARP and Bid by western blot analysis. We found
that after exposure to different concentrations of arecoline for 3
h, the expression of caspase-3, cleaved-PARP and Bid showed no
obvious change (Fig. 5A–C). As
shown in Fig. 5D, a 6-h exposure of
HaCaT cells to 0.4/0.5 mM of arecoline resulted in marked
activation of caspase-3 and cleaved-PARP. However, truncated Bid
(tBid) was only detected following arecoline exposure for >12 h.
As shown in Fig. 5E and F,
arecoline at 0.2–0.5 mM induced upregulation of cleaved-caspase-3,
Bid and PARP in a dose- and time-dependent manner. Our data imply
that exposure to high concentrations of arecolin or long-term
treatment of arecoline confers a significant cytotoxic effect, and
arecoline-induced HaCaT epithelial cell apoptosis may involve both
the death receptor and mitochondrial signaling pathways.
Discussion
Data from recent epidemiological studies provide
overwhelming evidence that the areca nut is the main etiological
factor for OSF (16,28–30).
Increased collagen synthesis and/or reduced collagen degradation
may be involved in the development of this disease. Betel quid
chewing affects the wound healing and fibrotic processes in OSF via
inducing buccal mucosa fibroblast contraction (31). Areca nut extract (ANE) or arecoline
stimulates collagen (32),
plasminogen activator inhibitor-1 (PAR-1) (33), cyclooxygenase-2 (34), tissue inhibitor of
metalloproteinase-1 (35)
expression and decreases collagen phagocytosis (36) in human fibroblasts. Several
signaling pathways are deregulated in OSF, such as TGF-β (37,38),
NF-κB, JNK, P38 and ERK MAPK (39,40),
which may contribute to areca quid chewing-associated OSF.
In order to further investigate the mechanism
underlying ANE- or arecoline-induced epithelial atrophy and
progressive accumulation of collagen fibers in the lamina propria
in OSF, we first assessed the sensitivities of the HaCaT epithelial
cells and the Hel fibroblast cells to arecoline exposure. The
salivary concentration of arecoline during BQ chewing has usually
been observed to be ~0.3 mM, and sometimes has been detected to
reach ~0.9 mM (41–43). In addition, Sundqvist et al
found that the areca alkaloids, such as arecoline, at a millimolar
concentration range is significant for the long-term effects of BQ
chewing on buccal mucosa and OSF (44). Therefore, we tested the effects of
arecoline on human HaCaT epithelial cells and Hel fibroblast cells
at concentrations ranging from 0.1 to 0.5 mM. Our data demonstrated
that arecoline dose-dependently promoted morphological changes in
HaCaT cells (Fig. 1), but had no
obvious effect on Hel cells (Fig.
2). We found that continued exposure to over 0.3 mM arecoline
for 48 h markedly induced HaCaT cell detachment and rounding.
Moreover, a significant proportion of HaCaT cells displayed
contraction and membrane blebbing, which is indicative of cell
apoptosis (Fig 1). MTS assay also
demonstrated that arecoline inhibited HaCaT cell, but not Hel cell
viability in a dose- and time-dependent manner (Fig 3). Our results imply that a high
concentration of arecoline has substantial cytotoxicity and that
betel quid chewing can directly induce epithelial cell apoptosis.
However, the Hel fibroblast cells were more tolerant to
arecoline-mediated cytotoxicity and apoptosis induction.
Cell death is a crucial event for a variety of
physiological processes. Its dysfunction leads to severe human
diseases that ensue from excessive cell accumulation (e.g.,
neoplasia and autoimmune disorders) or excessive cell loss (e.g.,
neurodegenerative diseases) (21).
Previous studies have indicated that arecoline-induced human oral
epithelial and endothelial cell cycle arrest or apoptosis is
involved in the pathogenesis of OSF (45–47).
However, the molecular mechanism has not been adequately
elucidated. In the present study, we first assessed
arecoline-induced apoptosis by flow cytometric analysis. Our
results showed that exposure to arecoline at concentrations >0.2
mM induced HaCaT cell apoptosis in a dose-dependent manner.
Notably, the Hel cells were much more resistant to
arecoline-induced apoptosis; 0.5 mM arecoline only slightly
enhanced Hel cell apoptosis (Fig
4).
To further investigate the mechanisms of
arecoline-induced apoptosis in HaCaT epithelial cells, we examined
the protein levels of cleaved-PARP, Bid and caspase-3 by western
blot analysis. We clearly detected caspsae-3 and PARP cleavage
following exposure to >0.3 mM arecoline for 6 h (Fig 5D). It has been demonstrated that
arecoline can activate the intrinsic apoptosis pathway, and that
arecoline treatment increases the cleavage of procaspase-9
(48). Moreover, exposure to ANE or
arecoline directly deregulated the expression of Bcl-2 family
proteins, such as Bcl-2, Bcl-Xl and Bax (49), which disequilibrated mitochondrial
controlled apoptosis. Unfortunately, no evidence has illustrated
that the extrinsic apoptosis pathway is upregulated in
arecoline-induced apoptosis. In this study, we did not detect
significant caspase-8 cleavage in HaCaT cell (data not shown).
However, the downstream target protein of cleaved-caspase-8, Bid,
was markedly cleaved in a dose- and time-dependent manner following
exposure to arecoline (Fig 5E and
F), indicating the possibility that other factors or pathways
are involved in arecoline-induced Bid activation. Further
investigation is warranted to determine whether arecoline directly
activates other cell-death modes to induce Bid cleavage and link
with the mitochondrial pathway, regulating cell apoptosis.
Overall, the present study demonstrated that
arecoline inhibits HaCaT epithelial cell viability and induces
apoptosis in a dose- and time-dependent manner, while this
observation was not obvious in Hel fibroblast cells. These results
suggest that epithelial cells are more sensitive to
arecoline-mediated cytotoxicity than fibroblast cells, which may
contribute to BQ chewing-induced epithelial atrophy and progressive
accumulation of collagen fibers in OSF. Our findings may aid in the
further understanding of the pathogenesis of arecoline-associated
OSF, and may facilitate the future development of preventive and
therapeutic strategies.
Acknowledgements
The study was supported by the National Natural
Science Foundation of China (81371690) and the International
Cooperation Program Funds of the China Hunan Provincial Science and
Technology Department (project no. 2012WK4005 and 2013
2013FJ6009).
Abbreviations:
|
OSF
|
oral submucous fibrosis
|
|
BQ
|
betel quid
|
|
OSCC
|
oral squamous cell carcinoma
|
|
ANE
|
areca nut extract
|
|
PARP
|
poly(ADP-ribose) polymerase
|
References
|
1
|
Khan S, Chatra L, Prashanth SK, et al:
Pathogenesis of oral submucous fibrosis. J Cancer Res Ther.
8:199–203. 2012. View Article : Google Scholar
|
|
2
|
Arakeri G and Brennan PA: Oral submucous
fibrosis: an overview of the aetiology, pathogenesis,
classification, and principles of management. Br J Oral
Maxillofacial Surg. 51:587–593. 2013. View Article : Google Scholar
|
|
3
|
Cox SC and Walker DM: Oral submucous
fibrosis. A review. Aust Dent J. 41:294–299. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mahomed F: Oral submucous fibrosis - a
potentially malignant condition of growing concern. SADJ.
67:562–565. 2012.PubMed/NCBI
|
|
5
|
Chole RH, Gondivkar SM, Gadbail AR, et al:
Review of drug treatment of oral submucous fibrosis. Oral Oncol.
48:393–398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paissat DK: Oral submucous fibrosis. Int J
Oral Surg. 10:307–312. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Aziz SR: Oral submucous fibrosis: an
unusual disease. J N J Dent Assoc. 68:17–19. 1997.PubMed/NCBI
|
|
8
|
Warnakulasuriya S, Johnson NW and van der
Waal I: Nomenclature and classification of potentially malignant
disorders of the oral mucosa. J Oral Pathol Med. 36:575–580. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tilakaratne WM, Klinikowski MF, Saku T, et
al: Oral submucous fibrosis: review on aetiology and pathogenesis.
Oral Oncol. 42:561–568. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Walvekar RR, Chaukar DA, Deshpande MS, et
al: Verrucous carcinoma of the oral cavity: a clinical and
pathological study of 101 cases. Oral Oncol. 45:47–51. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thomas SJ, Bain CJ, Battistutta D, et al:
Betel quid not containing tobacco and oral cancer: a report on a
case-control study in Papua New Guinea and a meta-analysis of
current evidence. Int J Cancer. 120:1318–1323. 2007. View Article : Google Scholar
|
|
12
|
Ko YC, Huang YL, Lee CH, et al: Betel quid
chewing, cigarette smoking and alcohol consumption related to oral
cancer in Taiwan. J Oral Pathol Med. 24:450–453. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Panigrahi GB and Rao AR:
Chromosome-breaking ability of arecoline, a major betel-nut
alkaloid, in mouse bone-marrow cells in vivo. Mutat Res.
103:197–204. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chou WW, Guh JY, Tsai JF, et al:
Arecoline-induced phosphorylated p53 and p21(WAF1) protein
expression is dependent on ATM/ATR and
phosphatidylinositol-3-kinase in clone-9 cells. J Cell Biochem.
107:408–417. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Stich HF, Stich W and Lam PP: Potentiation
of genotoxicity by concurrent application of compounds found in
betel quid: arecoline, eugenol, quercetin, chlorogenic acid and
Mn2+. Mutat Res. 90:355–363. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jeng JH, Chang MC and Hahn LJ: Role of
areca nut in betel quid-associated chemical carcinogenesis: current
awareness and future perspectives. Oral Oncol. 37:477–492. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Dasgupta R, Saha I, Pal S, et al:
Immunosuppression, hepatotoxicity and depression of antioxidant
status by arecoline in albino mice. Toxicology. 227:94–104. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Grutter MG: Caspases: key players in
programmed cell death. Curr Opin Struct Biol. 10:649–655. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Crowder RN and El-Deiry WS: Caspase-8
regulation of TRAIL-mediated cell death. Exp Oncol. 34:160–164.
2012.PubMed/NCBI
|
|
20
|
Cory S and Adams JM: The Bcl2 family:
regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ghiotto F, Fais F and Bruno S: BH3-only
proteins: the death-puppeteer’s wires. Cytometry A. 77:11–21.
2010.PubMed/NCBI
|
|
22
|
Tait SW and Green DR: Mitochondria and
cell death: outer membrane permeabilization and beyond. Nat Rev Mol
Cell Biol. 11:621–632. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kantari C and Walczak H: Caspase-8 and
bid: caught in the act between death receptors and mitochondria.
Biochim Biophys Acta. 1813:558–563. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang K, Yin XM, Chao DT, et al: BID: a
novel BH3 domain-only death agonist. Genes Dev. 10:2859–2869. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li H, Zhu H, Xu CJ, et al: Cleavage of BID
by caspase 8 mediates the mitochondrial damage in the Fas pathway
of apoptosis. Cell. 94:491–501. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Luo X, Budihardjo I, Zou H, et al: Bid, a
Bcl2 interacting protein, mediates cytochrome c release from
mitochondria in response to activation of cell surface death
receptors. Cell. 94:481–490. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gross A, Yin XM, Wang K, et al: Caspase
cleaved BID targets mitochondria and is required for cytochrome
c release, while BCL-XL prevents this release but not tumor
necrosis factor-R1/Fas death. J Biol Chem. 274:1156–1163.
1999.PubMed/NCBI
|
|
28
|
Reichart PA: Oral cancer and precancer
related to betel and miang chewing in Thailand: a review. J Oral
Pathol Med. 24:241–243. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Trivedy CR, Craig G and Warnakulasuriya S:
The oral health consequences of chewing areca nut. Addict Biol.
7:115–125. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
IARC Working Group on the Evaluation of
Carcinogenic Risks to Humans. Betel-quid and areca-nut chewing and
some areca-nut derived nitrosamines. IARC Monogr Eval Carcinog
Risks Hum. 85:1–334. 2004.PubMed/NCBI
|
|
31
|
Chang MC, Lin LD, Wu HL, et al: Areca
nut-induced buccal mucosa fibroblast contraction and its signaling:
a potential role in oral submucous fibrosis - a precancer
condition. Carcinogenesis. 34:1096–1104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li X, Ling TY, Gao YJ, Tang DS and Li WH:
Arecoline and oral keratinocytes may affect the collagen metabolism
of fibroblasts. J Oral Pathol Med. 38:422–426. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yang SF, Hsieh YS, Tsai CH, et al:
Increased plasminogen activator inhibitor-1/tissue type plasminogen
activator ratio in oral submucous fibrosis. Oral Dis. 13:234–238.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chiang SL, Chen PH, Lee CH, et al:
Up-regulation of inflammatory signalings by areca nut extract and
role of cyclooxygenase-2-1195G>a polymorphism reveal risk of
oral cancer. Cancer Res. 68:8489–8498. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shieh DH, Chiang LC and Shieh TY:
Augmented mRNA expression of tissue inhibitor of
metalloproteinase-1 in buccal mucosal fibroblasts by arecoline and
safrole as a possible pathogenesis for oral submucous fibrosis.
Oral Oncol. 39:728–735. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shieh DH, Chiang LC, Lee CH, et al:
Effects of arecoline, safrole, and nicotine on collagen
phagocytosis by human buccal mucosal fibroblasts as a possible
mechanism for oral submucous fibrosis in Taiwan. J Oral Pathol Med.
33:581–587. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Khan I, Kumar N, Pant I, et al: Activation
of TGF-beta pathway by areca nut constituents: a possible cause of
oral submucous fibrosis. PloS One. 7:e518062012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kale AD, Mane DR and Shukla D: Expression
of transforming growth factor beta and its correlation with
lipodystrophy in oral submucous fibrosis: an immunohistochemical
study. Med Oral Patol Oral Cir Bucal. 18:e12–e18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yu CC, Tsai CH, Hsu HI, et al: Elevation
of S100A4 expression in buccal mucosal fibroblasts by arecoline:
involvement in the pathogenesis of oral submucous fibrosis. PloS
One. 8:e551222013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Deng YT, Chen HM, Cheng SJ, et al:
Arecoline-stimulated connective tissue growth factor production in
human buccal mucosal fibroblasts: modulation by curcumin. Oral
Oncol. 45:e99–e105. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Nair J, Ohshima H, Friesen M, et al:
Tobacco-specific and betel nut-specific N-nitroso compounds:
occurrence in saliva and urine of betel quid chewers and formation
in vitro by nitrosation of betel quid. Carcinogenesis. 6:295–303.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Shirname LP, Menon MM, Nair J, et al:
Correlation of mutagenicity and tumorigenicity of betel quid and
its ingredients. Nutr Cancer. 5:87–91. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cox S, Vickers ER, Ghu S and Zoellner H:
Salivary arecoline levels during areca nut chewing in human
volunteers. J Oral Pathol Med. 39:465–469. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Sundqvist K, Liu Y, Nair J, et al:
Cytotoxic and genotoxic effects of areca nut-related compounds in
cultured human buccal epithelial cells. Cancer Res. 49:5294–5298.
1989.PubMed/NCBI
|
|
45
|
Lee PH, Chang MC, Chang WH, et al:
Prolonged exposure to arecoline arrested human KB epithelial cell
growth: regulatory mechanisms of cell cycle and apoptosis.
Toxicology. 220:81–89. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tseng SK, Chang MC, Su CY, et al:
Arecoline induced cell cycle arrest, apoptosis, and cytotoxicity to
human endothelial cells. Clin Oral Investig. 16:1267–1273. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou ZS, Li M, Gao F, et al: Arecoline
suppresses HaCaT cell proliferation through cell cycle regulatory
molecules. Oncol Rep. 29:2438–2444. 2013.PubMed/NCBI
|
|
48
|
Shih YT, Chen PS, Wu CH, et al: Arecoline,
a major alkaloid of the areca nut, causes neurotoxicity through
enhancement of oxidative stress and suppression of the antioxidant
protective system. Free Radic Biol Med. 49:1471–1479. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cheng HL, Su SJ, Huang LW, et al:
Arecoline induces HA22T/VGH hepatoma cells to undergo anoikis -
involvement of STAT3 and RhoA activation. Mol Cancer. 9:1262010.
View Article : Google Scholar : PubMed/NCBI
|