Introduction
As one of the most common malignant tumors in the
urinary system, renal cancer accounts for more than 270,000 new
cases worldwide each year (1).
Clear cell renal cell carcinoma (ccRCC) is the most common
histological subtype, accounting for 75–85% of all RCC cases. The
incidence of RCC has steadily increased in many countries (2). However, the treatment options for RCC
are limited, and the mechanisms of the pathogenesis of renal
carcinoma are not completely defined. Therefore, new treatment
strategies for patients with RCC are urgently needed. With
continued research on the mechanisms of tumors, targeted therapy
has made remarkable progress (3).
In recent years, novel markers and therapeutic targets of RCC have
continued to emerge. Nevertheless, to date, no biomarkers have been
used in RCC in the routine clinic (4).
The Hippo signaling pathway, initially discovered in
Drosophila, is a potent regulator of cell growth and
apoptosis (5). A series of
biochemical and genetic studies identified this pathway whose
components are highly conserved between Drosophila and
mammals (6,7). In mammals, the components of the Hippo
pathway include MST1/2, Sav1, LATS1/2, YAP, TAZ and TEAD (8,9).
MST1/2, Sav1 and LATS1/2 are regarded as upstream kinases of the
mammalian Hippo pathway. It is presumed that through
phosphorylation of YAP, the upstream kinases LATS1/2 inhibit YAP
transcription activity (10). YAP,
TAZ and TEAD were identified as downstream transcription factors of
this pathway (11).
Yes-associated protein (YAP), the mammalian homolog
of Drosophila Yorkie (Yki), is a 65-kDa protein which was
originally identified due to its interaction with the Src family
tyrosine kinase c-Yes (12). Recent
studies have found that the transcriptional co-activator YAP is a
key nuclear effector of the Hippo pathway which regulates organ
size by governing cell proliferation and apoptosis (13,14).
The TEAD family, located in the downstream of the Hippo pathway,
are major mediators of YAP transcriptional activity (15,16).
The domain of YAP which interacts with the TEAD family of
transcription factors is essential for YAP-mediated tumor growth
and metastasis (17). Recently, YAP
has been shown to be a candidate oncogene in recurrent
amplification at human chromosome 11q22 (18). Overexpression of YAP protein and its
nuclear localization were observed in colon, lung, pancreatic,
hepatocellular, ovarian and prostate carcinomas (16,19,20).
Despite of the growing evidence of YAP as a crucial
regulator of human types of cancers, its involvement in ccRCC is
still obscure. In the present study, we attempted to identify the
role of YAP in ccRCC. We evaluated the expression levels and
clinical significance of YAP in ccRCC tissues. In addition, we
investigated the effects of YAP silencing via YAP-shRNA lentiviral
vectors transfected into ccRCC cell lines. These findings suggest
that YAP may serve as a potential target in the treatment of
ccRCC.
Materials and methods
Tissue samples and cell culture
Tissue samples from radical nephrectomy were
obtained from the First Affiliated Hospital of Chongqing Medical
University from March 2010 to November 2011. The tissue samples
consisted of 30 cases of ccRCC tissues and 30 cases of adjacent
normal renal tissues (4 cm or more from the tumor). Samples were
confirmed by a pathologist and stored at −80°C. The present study
was conducted with approval from the Ethics Committee of the First
Affiliated Hospital of Chongqing Medical University. Informed
consent was obtained from all patients. Human ccRCC cell lines
786-0 and ACHN and the human HEK-293 cell line were purchased from
ATCC. ACHN and 786-0 cells were cultured in RPMI-1640 medium, and
HEK-293 in DMEM. All medium contained 10% fetal bovine serum (FBS)
(Gibco-BRL, Carlsbad, CA, USA). All cells were maintained in an
incubator with 5% CO2 at 37°C.
Immunohistochemistry
The tissues were fixed with 10% formaldehyde
(ZSGB-Bio, Beijing, China) embedded in paraffin, sliced into 4-μm
sections and used for staining. In brief, the sections were
rehydrated, and antigen retrieval was performed by a microwave
vacuum histoprocessor (RHS-1; Bergamo, Italy) at 121°C for 15 min.
After being blocked with goat serum (Gibco-BRL), the primary
antibody against YAP (Santa Cruz, Dallas, TX, USA) was diluted
(1:200) and incubated with the tissue sections overnight in a
humidity chamber at 4°C. The sections were treated with horseradish
peroxidase (HRP)-labeled secondary antibody (ZSGB-Bio) and DAB, and
the results were observed with a microscope. The tissues were
scored according to the percentage of the positively stained area
and staining intensity. The percentage of the positive area was
graded as 0 (≤5%), 1 (6–25%), 2 (26–50%), 3 (≥51%), and the
staining intensity was graded as 0–2 (i.e., negative, 0; weak, 1
and strong, 2). The two grades were multiplied and tissues were
assigned to one of 3 levels: 0, negative; 1–4, weak positive; and
5–6, strong positive.
Construction of the lentiviral
interference vectors and transfection
According to the YAP gene sequence (NM_006106) in
the GenBank, three sequences of the shRNA targeting YAP were
designed as follows: 5′-GCTCATTCCTCTCCAGCTT-3′,
5′-CCTTAACAGTGGCACCTAT-3′ and 5′-CCGTTTCCCAG ACTACCTT-3′.
Non-silencing shRNA (5′-TTCTCCGAAC GTGTCACGT-3′) was synthesized as
the negative control (NC shRNA). The YAP lentiviral interference
vector was designed and constructed by Shanghai SBO Medical
Biotechnology. Briefly, for generation of shRNA-YAP or control
lentiviral vector, the lentivirus expressing plasmids pLTR-G and
pNLEGFP/CMV/WPREdU3 which were carrying shRNA, along with
lentivirus packing plasmids pCD/NL-BH*DDD (both from
Addgene, USA) were added into 293T cells for lentiviral vector
packaging. Eventually, the lentivirus was extracted and lentivirus
concentrations were verified. For lentivirus transfection, 786-0
cells were grown to 30–50% confluency and transfected with three
pairs of YAP-shRNAs and the non-silencing control shRNA at an
optimized multiplicity of infection (MOI) of 20 with Polybrene at a
concentration of 5 μg/ml. The successful transfection of YAP-shRNA
lentiviral vectors was observed by fluorescence microscope 96 h
after transfected. The YAP-shRNA infected cells were detected as
GFP-positive cells under fluorescence microscopy at a magnification
of ×200 (Olympus, Tokyo, Japan). To detect the transfection
efficiency, the mRNA and protein levels of YAP were respectively
assessed by RT-PCR and western blotting 96 h after
transfection.
RT-PCR
The tissues and the cultured cells were collected.
Total RNA were extracted using the TRIzol kit (Takara, Osaka,
Japan). The total RNA quality was detected by a UV
spectrophotometer (Bio-Rad, Hercules, CA, USA), and 1 μg total RNA
was transcribed reversely for the cDNA and PCR reaction
amplification of YAP and TEAD1 genes, and GAPDH as an internal
reference. Specific primers were designed: YAP-F,
5-TGAACAAACGTCCAGCAAGATAC-3 and YAP-R, 5-CAGCCCCCAAAATGAACAGTAG-3;
TEAD1-F, 5-TGAATCAGTGGACATTCGTCA-3 and TEAD1-R, 5-GC
CATTCTCAAACCTTGCATA-3; and GAPDH-F, 5-ACCAC CATGGAGAAGGCTGG-3 and
GAPDH-R, 5-CTCAGTTAG CCCAGGATGC-3. PCR products were subjected to
agarose gel electrophoresis (SABC, China), UV light observation and
photography. A Bio-Rad gel formatter was used to analyze the
original band.
Western blotting
The YAP antibody was purchased from Santa Cruz
Biotechnology (Santa Cruz, CA, USA) and the TEAD1 antibody was
purchased from Proteintech Group, Inc. (Chicago, IL, USA). Total
protein was extracted from the tissue homogenates and the cell
lysate by RIPA buffer (50 mM Tris, pH 7.4, 150 mM NaCl, 1% Triton
X-100, 1% Nonidet-P40, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM
EDTA and PMSF) (Beyotime, Shanghai, China). The lysate containing
50 μg protein quantified by the BCA protein assay kit for each
sample was subjected to SDS-PAGE, and was then transferred to a
0.45-μm polyvinylidene fluoride (PVDF) membrane (both from
Beyotime). The membrane was blocked in Tris-buffered saline-Tween
0.1% (TBST) with 5% skimmed milk powder for 2 h at 37°C. The
primary antibodies were diluted (YAP 1:200, TEAD1 1:200 and β-actin
1:1,000). The proper species and diluted HRP-labeled secondary
antibodies were added, and Western blot results were analyzed by
Bio-Rad software.
Proliferation assay
The 786-0 cells were seeded in 96-well plates for 12
h at a concentration of 5×103 cells/well. Cells were
transfected with shYAP-3 or NC shRNA lentiviral vectors. After
treatment for 48, 96 and 144 h, 10 μl of Cell Counting Kit-8
(CCK-8) reagent (Sigma, USA) in 90 μl RPMI-1640/well was added to
the cell culture medium and then incubated at 37°C for 1 h. The
absorbance was measured with a microplate reader (Spectramax M2;
MD, USA) at 450 nm.
Cell cycle assay by flow cytometry
(FCM)
The 786-0 cells were transfected with the shYAP-3
lentiviral vector which was the most efficient inhibition
transfected group or NC shRNA for 96 h, and then harvested
abundantly. The cells were washed with PBS twice, and fixed with
70% ethanol overnight at 4°C. Fixed cells were stained with 50
μg/μl propidium iodide (PI) (Bioscience, Shanghai, China) and 100
μg/μl RNase (Sigma). Cell cycle distribution of each group was
measured with a flow cytometer (Beckman Coulter, USA).
Apoptosis assay
The 786-0 cells were transfected with shYAP-3 or the
NC shRNA lentiviral vector for 96 h, then trypsinized and washed
with PBS, resuspended in 100 μl binding buffer, and stained with
7-aminoactinomycin D (7-AAD) and PE-conjugated Annexin V
(Bioscience). The apoptosis rate of each group was measured with a
flow cytometer (Beckman Coulter).
Statistical analysis
Statistical analysis was performed with SPSS 19.0
software. Data are presented as the means ± standard deviation. The
Chi-square trend test was used to assess the correlation between
YAP protein expression and clinicopathologic factors of the renal
cancer cases. A Student’s two-tailed t-test was used to compare two
groups. One-way ANOVA was used in comparison of means among
multiple groups. p<0.05 was considered to indicate a
statistically significant result.
Results
Expression of YAP in ccRCC tissues and
correlation with clinicopathological factors
As determined by RT-PCR, the expression level of YAP
mRNA in the ccRCC tissues was 0.569±0.066, while that in adjacent
normal renal tissues was 0.515±0.068. The expression level of YAP
mRNA in ccRCC tissues was significantly higher than that in
adjacent normal renal tissues (p<0.05) (Fig. 1).
The expression of YAP protein in 30 cases of ccRCC
tissues and 30 cases of adjacent normal renal tissues was detected
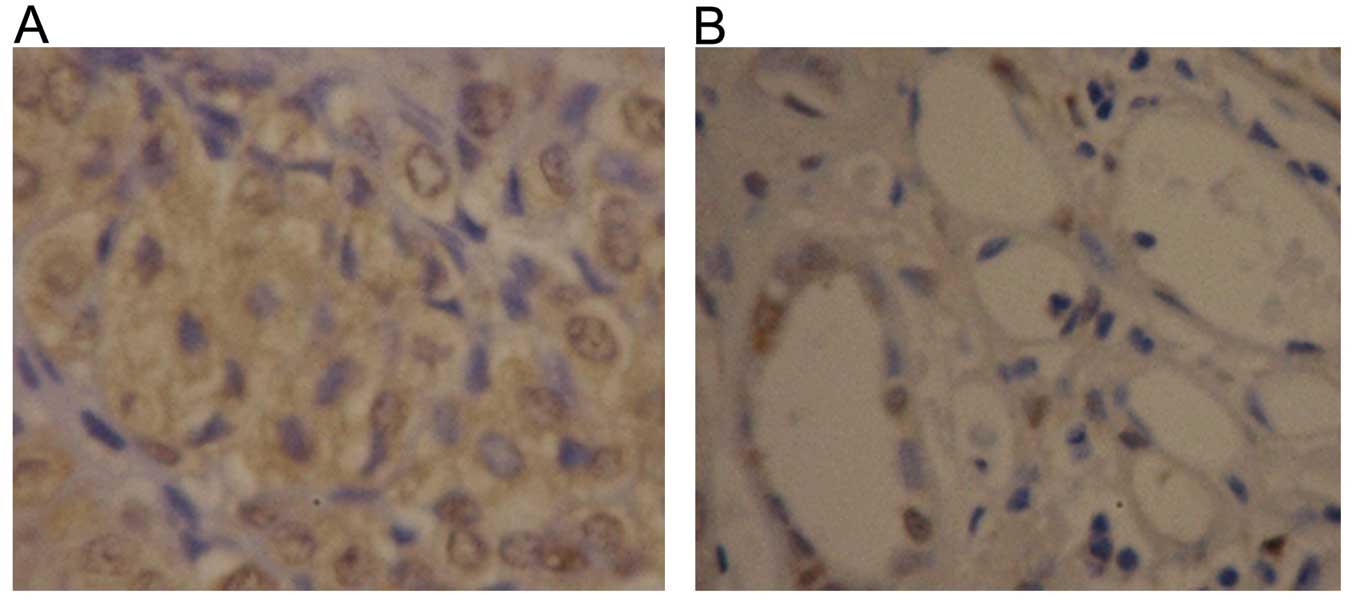
by immunohistochemistry. In the ccRCC tissues, 63.3% (19/30) of the
samples exhibited positive expression, and the positive areas were
primarily located in the nucleus and cytoplasm (Fig. 2A). In contrast, the positive rate of
YAP protein expression in the normal tissues was 33.3% (10/30);
most cases showed weak positive expression and the YAP protein was
confined to the renal collecting duct system (Fig. 2B). The positive rate of YAP protein
in the ccRCC tissues was obviously higher than that in the adjacent
normal renal tissues (p<0.05).
We further analyzed the correlation of YAP protein
expression with clinicopathological factors of ccRCC. As shown in
Table I, there was no obvious
relationship between YAP protein expression and gender, age, renal
vein metastasis or tumor size. However, the positive rate of YAP
protein expression was 33.3% (3/8) in well differentiated, 61.5%
(8/13) in moderately differentiated and 88.9% (8/9) in low
differentiated tumors (p=0.018). In addition, the positive rate of
YAP protein expression was 47.1% (8/17) in stage I–II and 84.6%
(11/13) in stage III–IV cases (p=0.034). These results showed that
the expression level of YAP protein was closely correlated to
clinical stage and differentiation in the ccRCC tissues.
 | Table IAssociation of YAP protein expression
with clinicopathological factors of the ccRCC cases
(χ2-test). |
Table I
Association of YAP protein expression
with clinicopathological factors of the ccRCC cases
(χ2-test).
| Factors | Patients | YAP expression | (−) | YAP-positive rate
(%) | P-value |
|---|
|
|---|
| (+) |
|---|
| Gender | | | | | 0.705 |
| Male | 15 | 10 | 5 | 66.7 | |
| Female | 15 | 9 | 6 | 60.0 | |
| Age (years) | | | | | 0.757 |
| <60 | 18 | 11 | 7 | 61.1 | |
| ≥60 | 12 | 8 | 4 | 66.7 | |
| Renal vein
metastasis | | | | | 0.265 |
| Present | 2 | 2 | 0 | 100.0 | |
| None | 28 | 17 | 11 | 60.7 | |
| Tumor size
(cm) | | | | | 0.424 |
| >5 | 8 | 6 | 2 | 75.0 | |
| ≤5 | 22 | 13 | 9 | 59.1 | |
|
Differentiation | | | | | 0.018 |
| Well | 8 | 3 | 5 | 33.3 | |
| Moderate | 13 | 8 | 5 | 61.5 | |
| Poor | 9 | 8 | 1 | 88.9 | |
| Clinical stage | | | | | 0.034 |
| I–II | 17 | 8 | 9 | 47.1 | |
| III–IV | 13 | 11 | 2 | 84.6 | |
Expression of YAP in 786-0 cells is
suppressed by shRNA lentiviral vectors
We examined the expression level of YAP protein in
ccRCC cell lines 786-0 and ACHN and in the human embryonic kidney
HEK-293 cells using western blotting. As shown in Fig. 3A, the YAP protein levels in the two
ccRCC cell lines 786-0 and ACHN were significantly higher than that
in the HEK-293 cells (p<0.05).
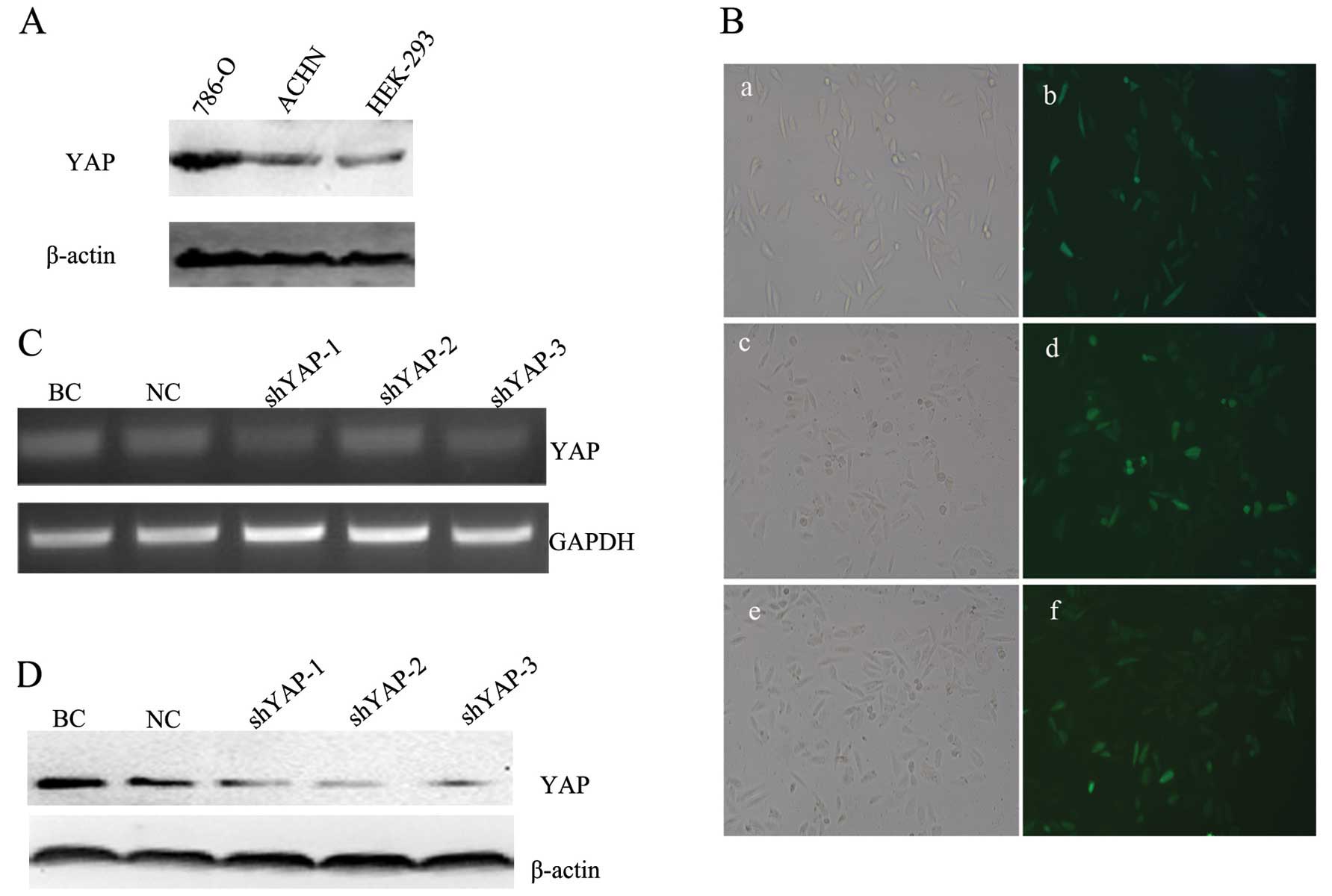 | Figure 3YAP-shRNA lentiviral vector was
transfected into 786-0 cells and suppressed the expression levels
of YAP mRNA and protein. (A) The YAP protein levels in two ccRCC
cell lines and HEK-293 cells. (B) GFP expression in 786-0 cells at
96 h after transfection with the YAP-shRNA lentiviral vector as
shown by fluorescence microscopy (magnification, ×200). a, c and e
images, respectively, represent shYAP-1, the shYAP-2 and shYAP-3
transfected group cells under light microscopy; b, d and f images,
respectively, represent shYAP-1, shYAP-2 and shYAP-3 transfected
group cells under fluorescence microscopy. (C) The 786-0 cells were
transfected with shYAP-1, shYAP-2, shYAP-3 and NC lentiviral
vector, and the BC group was used as a blank control. YAP mRNA
expression was assessed by RT-PCR. Expression of GAPDH is shown for
comparison. (D) Expression of YAP protein in the different groups
was confirmed by western blotting. Expression of β-actin is shown
for comparison. BC, blank control group, cells only; NC, negative
control group, negative control vector; YAP, Yes-associated
protein; ccRCC, clear cell renal cell carcinoma. |
To determine whether YAP plays a role in
tumorigenesis of renal cancer, three specific YAP-shRNA lentiviral
vectors, shYAP-1, shYAP-2 and shYAP-3, were successfully
constructed and transfected into 786-0 cells. After 96 h of
transfection, as shown in Fig. 3B,
786-0 cells were successfully transfected with the YAP-shRNA
lentiviral vectors. Three YAP interference groups, as well as a
blank control group (BC group, cells only) and negative control
group (NC group) were established from the 786-0 cells.
Furthermore, the efficient knockdown of YAP expression was detected
by RT-PCR and Western blot analysis. RT-PCR analysis showed that
the relative mRNA expression level of YAP in the YAP-shRNA
transfection cells was markedly lower than levels in the BC and NC
groups (p<0.05) (Fig. 3C).
Similarly, the expression of YAP protein was also significantly
inhibited as confirmed by western blotting (p<0.05) (Fig. 3D). The lowest level of YAP
expression was observed in the cells transfected with the shYAP-3
lentiviral vectors. These results indicated that the shYAP-3
lentiviral vector had the highest inhibition rate of the three
specific YAP-shRNA lentiviral vectors. Therefore, the shYAP-3
lentiviral vector was chosen for subsequent experiments.
Knockdown of YAP inhibits expression of
TEAD1
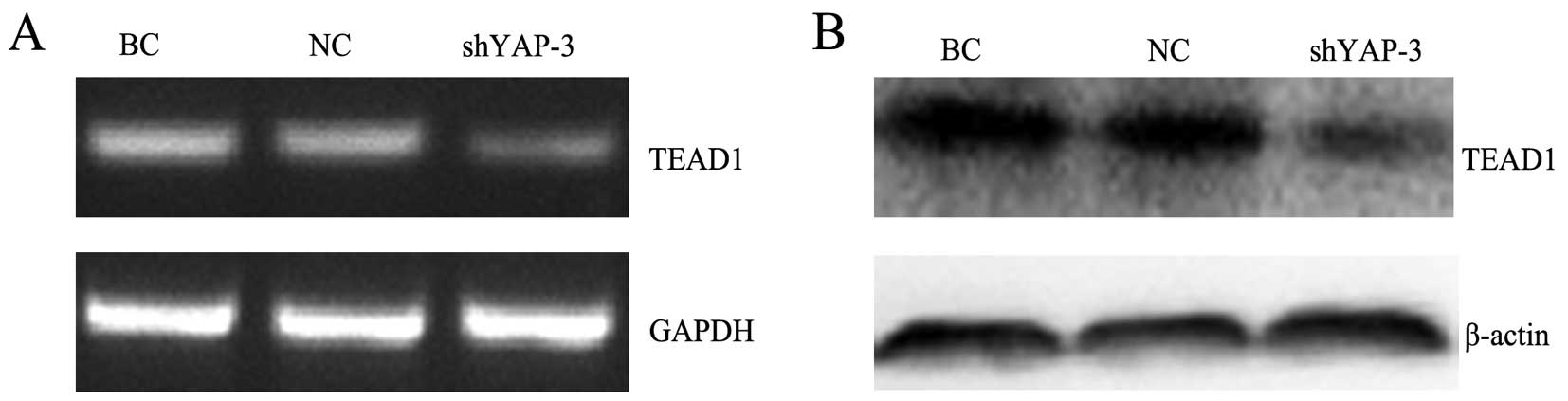
RT-PCR analysis showed that the relative mRNA
expression level of TEAD1 in the shYAP-3-transfected cells was
markedly lower than levels in the BC and NC group cells (p<0.05)
(Fig. 4A). As shown in the western
blot analysis (Fig. 4B), we also
observed that the protein expression level of TEAD1 was
significantly decreased compared with the levels in the BC and NC
groups (p<0.05), indicating that silencing of YAP inhibited the
expression level of TEAD1, which is a downstream gene in the Hippo
pathway.
YAP knockdown suppresses cell
proliferation
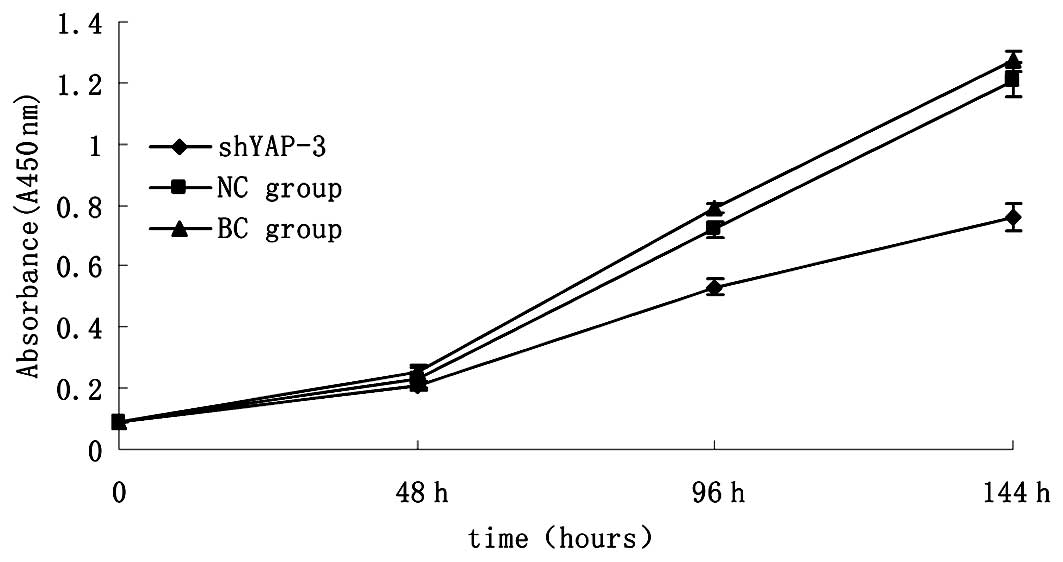
To determine whether the YAP-shRNA lentiviral vector
actually affects the proliferation of 786-0 cells, we examined the
proliferation of the three 786-0 cell groups including the
shYAP-3-transfected group, the blank group and negative control
group. As shown in Fig. 5, after
silencing of YAP in the 786-0 cells for 144 h, we found that there
was a significant reduction in cell proliferation (p<0.05). In
addition, the NC group presented a minimal effect on the
proliferation of 786-0 cells compared with the blank control group
(p>0.05). The results demonstrated that YAP plays an important
role in cell survival, and the knockdown of the YAP gene leads to
suppression of renal cancer cell proliferation.
YAP knockdown arrests the cell cycle and
induces cell apoptosis
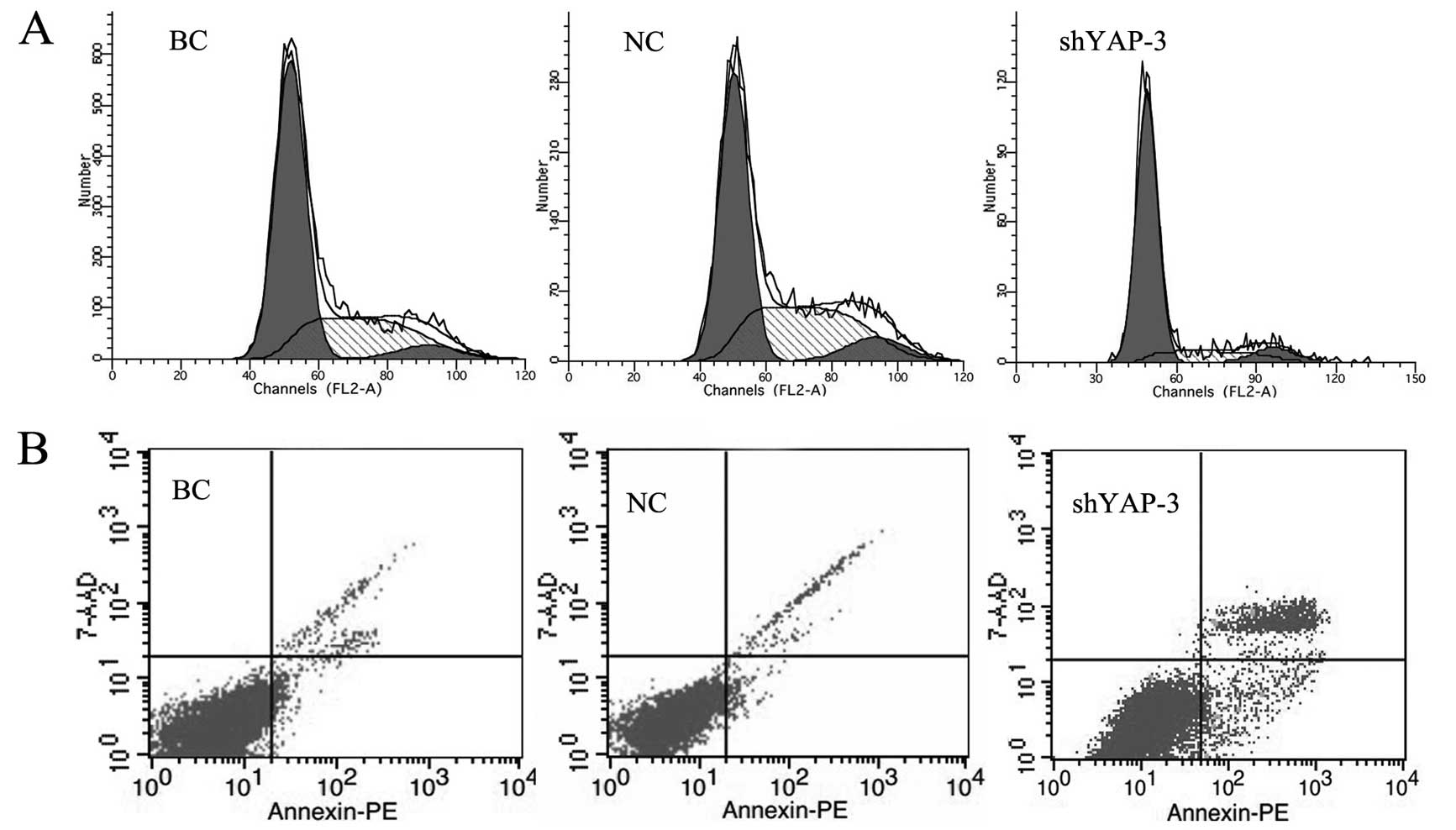
Ninety-six hours after shYAP-3 lentiviral vector
transfection, the effect of YAP on the cell cycle progression was
analyzed by FCM. In the shYAP-3-transfected group, we found that
61.4% of cells were in the G1 phase and 29.4% were in the S phase,
compared with 51.4% of cells in the G1 phase and 39.4% in the S
phase in the blank control group and 51.0% of cells in G1 phase and
39.8% in S phase in the NC group (p<0.05) (Fig. 6A). No significant difference was
observed in cell cycle progression between the blank control and
the NC group (p>0.05). Furthermore, the apoptosis rate of cells
with YAP knockdown was analyzed by FACS. The results showed that
the apoptosis rate in the 786-0 cells with YAP knockdown was
significantly increased as compared with the rates in the control
groups (p<0.05) (Fig. 6B). The
difference between the blank control group and the NC group was not
obvious (p=0.582). This finding suggests that knockdown of YAP
induced 786-0 cell apoptosis and G1 phase arrest.
Discussion
The Hippo pathway has been recognized as an
important regulator of both organ size control and tumorigenesis
(6). In mammals, the Hippo pathway
negatively regulates cell growth through a kinase cascade and
results in the inactivation of YAP. Hippo core components form
protein kinase complexes acting in a cascade to phosphorylate YAP
and induce its cytoplasmic translocation (21). YAP expression has been significantly
associated with tumor metastasis, grade and stage (22). A study in lung cancer showed that
YAP expression was significantly correlated with TNM stage
(23).
In the present study, YAP mRNA and protein
expression levels in 30 cases of ccRCC tissues and 30 cases of
adjacent normal renal tissues were first detected. The results
indicated that expression levels of YAP mRNA and protein in ccRCC
tissues were significantly higher than those in adjacent normal
renal tissues. ccRCC tissues had a high frequency of YAP-positive
cells, whereas the adjacent normal tissues had a lower frequency
(63.3% vs. 33.3%). Although a significant percentage of adjacent
normal cases stained positive for YAP, the overall expression level
was significantly higher in the tumor tissues. We then found that
the expression level of YAP protein in ccRCC tissues was
significantly correlated with differentiation and clinical stage.
In our immunohistochemistry study, the staining intensity of YAP
was tightly linked with progression from well-differentiated to
poorly differentiated tumors. Furthermore, YAP protein expression
was correlated with clinical stage which may serve as an
independent predictor for renal cancer. In the lower stages of
ccRCC, there were low levels of expression of YAP, and higher
expression of YAP was accompanied by higher stages of malignancy.
In previous studies, YAP was found to play an important role in
tumorigenesis and to be as an independent prognostic marker in HCC
and ESCC (24,25). Consistent with these research
findings, the present study showed that the YAP gene is a
tumor-specific gene, which is involved in the progression of
ccRCC.
In previous studies, overexpression of YAP has been
found in several human cancer cell types, and the oncogenic
activity of YAP has been clearly demonstrated (16,25,26).
However, another study also reported that YAP may act as a tumor
suppressor in certain breast carcinomas (27). The expression and activity of YAP in
renal cancer cells are currently unknown. The present report showed
that there was a significantly higher expression of YAP protein in
ccRCC cell lines, 786-0 and ACHN, when compared with that in the
HEK-293 cells. Specific shRNA lentiviral vectors targeting YAP were
successfully constructed and transfected into 786-0 cells. We found
that YAP mRNA and protein were effectively inhibited in the 786-0
cells. Furthermore, functional experiments were performed. The
results indicated that silencing of YAP in 786-0 cells
significantly inhibited cell proliferation, induced G1 phase cell
cycle arrest and induced 786-0 cell apoptosis. Therefore, it is
evident from the results that YAP plays an important role in the
development of ccRCC.
In our initial experiment, we synthesized an siRNA
targeting YAP and transfected it into 786-0 cells using
Lipofectamine 2000, but it did not achieve the expected results;
the effect of the suppression of the expression of the YAP gene was
poor. Then, lentiviral vectors were engineered to knock down the
YAP gene in 786-0 cells. All of the three YAP-shRNA interference
groups were able to suppress the mRNA and protein expression levels
of YAP. Further statistical analysis found that the lowest level of
YAP mRNA expression in the 786-0 cells was in the shYAP-3
interference group, and the expression level of YAP protein in the
shYAP-3 interference group cells was in accordance with the
expression level of YAP mRNA. We chose lentiviral vectors for gene
silencing due to their attractive properties, including high
efficiency, persistent gene silencing and safety for humans
(28,29). Lentiviral vectors are a powerful
tool for gene transfer into a broad range of cell types (30). Lentiviral vectors producing shRNAs
have widespread use in the knockdown of gene expression both in
vitro and in vivo.
Acting as downstream gene of the Hippo pathway, YAP
is a transcription co-activator which interacts with the
PPXY-motif- containing transcription factors, including TEAD,
ErbB4, P73 and Runx2 (31,32). It has been reported that YAP and
TEAD1 genetically interact to promote tissue growth (33). TEAD as a new component in the Hippo
pathway plays an essential role in mediating the biological
functions of YAP. In humans, there are four TEAD family members.
YAP and TEAD1 bind to a common set of promoters in MCF10A cells
(33). Furthermore, TEAD1 has been
shown to be linked to the regulation of YAP in Drosophila
and mammalian epithelial cell lines (34). A recent study has provided proof of
principle that inhibiting TEAD-YAP interactions is a
pharmacologically viable strategy against the YAP oncoprotein
(35). TEAD has been demonstrated
to be important for the growth-promoting function of YAP (36). The present study showed that
silencing of YAP led to the decreased expression of TEAD1 and
resulted in the growth inhibition of 786-0 cells. Thus, we
speculated that the relevant mechanism involved the suppression of
TEAD1 expression by the knockdown of YAP which consequently affects
the expression of downstream cell growth factors and thus plays
functional roles in the regulation of 786-0 cell proliferation. Yet
this may not be the sole mechanism, and further research on the
molecular functions of the YAP gene in renal cancer is
warranted.
In conclusion, these findings demonstrated that YAP
is an oncogene overexpressed in ccRCC, and the expression level of
YAP protein is closely correlated with clinical stage and
differentiation of ccRCC tissues. Moreover, suppression of YAP
expression by YAP-shRNA lentiviral vectors reduced 786-0 cell
growth and arrested the cell cycle at the G1 phase and also led to
induced apoptosis. Therefore, YAP could be a potential novel
therapeutic target for the treatment of ccRCC.
Acknowledgements
The present study was supported by the Natural
Science Foundation of China (grant no. 30972999) and the Nature
Science Foundation of Chongqing (no. cqpc2012jja1698).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Quivy A, Daste A, Harbaoui A, Duc S,
Bernhard JC, Gross-Goupil M and Ravaud A: Optimal management of
renal cell carcinoma in the elderly: a review. Clin Interv Aging.
8:433–442. 2013.PubMed/NCBI
|
|
3
|
Rathmell WK and Godley PA: Recent updates
in renal cell carcinoma. Curr Opin Oncol. 22:250–256. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Vasudev NS, Selby PJ and Banks RE: Renal
cancer biomarkers: the promise of personalized care. BMC Med.
10:1122012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Edgar BA: From cell structure to
transcription: Hippo forges a new path. Cell. 124:267–273. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhao B, Li L, Lei Q and Guan KL: The
Hippo-YAP pathway in organ size control and tumorigenesis: an
updated version. Genes Dev. 24:862–874. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pan D: The hippo signaling pathway in
development and cancer. Dev Cell. 19:491–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhao B, Wei X, Li W, et al: Inactivation
of YAP oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stanger BZ: Quit your YAPing: a new target
for cancer therapy. Genes Dev. 26:1263–1267. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang J, Ji JY, Yu M, et al: YAP-dependent
induction of amphiregulin identifies a non-cell-autonomous
component of the Hippo pathway. Nat Cell Biol. 11:1444–1450. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bao Y, Hata Y, Ikeda M and Withanage K:
Mammalian Hippo pathway: from development to cancer and beyond. J
Biochem. 149:361–379. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dong J, Feldmann G, Huang J, et al:
Elucidation of a universal size-control mechanism in
Drosophila and mammals. Cell. 130:1120–1133. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhao B, Tumaneng K and Guan KL: The Hippo
pathway in organ size control, tissue regeneration and stem cell
self-renewal. Nat Cell Biol. 13:877–883. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Harvey K and Tapon N: The
Salvador-Warts-Hippo pathway - an emerging tumour-suppressor
network. Nat Rev Cancer. 7:182–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sawada A, Kiyonari H, Ukita K, Nishioka N,
Imuta Y and Sasaki H: Redundant roles of Tead1 and
Tead2 in notochord development and the regulation of cell
proliferation and survival. Mol Cell Biol. 28:3177–3189. 2008.
|
|
16
|
Chan SW, Lim CJ, Chen L, Chong YF, Huang
C, Song H and Hong W: The Hippo pathway in biological control and
cancer development. J Cell Physiol. 226:928–939. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lamar JM, Stern P, Liu H, Schindler JW,
Jiang ZG and Hynes RO: The Hippo pathway target, YAP, promotes
metastasis through its TEAD-interaction domain. Proc Natl Acad Sci
USA. 109:E2441–E2450. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zender L, Spector MS, Xue W, et al:
Identification and validation of oncogenes in liver cancer using an
integrative oncogenomic approach. Cell. 125:1253–1267. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li H, Wolfe A, Septer S, et al:
Deregulation of Hippo kinase signalling in human hepatic
malignancies. Liver Int. 32:38–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Steinhardt AA, Gayyed MF, Klein AP, et al:
Expression of Yes-associated protein in common solid tumors. Hum
Pathol. 39:1582–1589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang J, Smolen GA and Haber DA: Negative
regulation of YAP by LATS1 underscores evolutionary conservation of
the Drosophila Hippo pathway. Cancer Res. 68:2789–2794.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao B, Li L, Wang L, Wang CY, Yu J and
Guan KL: Cell detachment activates the Hippo pathway via
cytoskeleton reorganization to induce anoikis. Genes Dev. 26:54–68.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang Y, Dong Q, Zhang Q, Li Z, Wang E and
Qiu X: Overexpression of yes-associated protein contributes to
progression and poor prognosis of non-small-cell lung cancer.
Cancer Sci. 101:1279–1285. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu MZ, Yao TJ, Lee NP, et al:
Yes-associated protein is an independent prognostic marker in
hepatocellular carcinoma. Cancer. 115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Muramatsu T, Imoto I, Matsui T, et al: YAP
is a candidate oncogene for esophageal squamous cell carcinoma.
Carcinogenesis. 32:389–398. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Diep CH, Zucker KM, Hostetter G, et al:
Down-regulation of Yes associated protein 1 expression reduces cell
proliferation and clonogenicity of pancreatic cancer cells. PLoS
One. 7:e327832012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tufail R, Jorda M, Zhao W, Reis I and
Nawaz Z: Loss of Yes-associated protein (YAP) expression is
associated with estrogen and progesterone receptors negativity in
invasive breast carcinomas. Breast Cancer Res Treat. 131:743–750.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sakuma T, Barry MA and Ikeda Y: Lentiviral
vectors: basic to translational. Biochem J. 443:603–618. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Abbas-Terki T, Blanco-Bose W, Déglon N,
Pralong W and Aebischer P: Lentiviral-mediated RNA interference.
Hum Gene Ther. 13:2197–2201. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Santamaria J, Khalfallah O, Sauty C, et
al: Silencing of choline acetyltransferase expression by
lentivirus-mediated RNA interference in cultured cells and in the
adult rodent brain. J Neurosci Res. 87:532–544. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Strano S, Monti O, Pediconi N, et al: The
transcriptional coactivator Yes-associated protein drives p73
gene-target specificity in response to DNA damage. Mol Cell.
18:447–459. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li Z, Zhao B, Wang P, et al: Structural
insights into the YAP and TEAD complex. Genes Dev. 24:235–240.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhao B, Ye X, Yu J, et al: TEAD mediates
YAP-dependent gene induction and growth control. Genes Dev.
22:1962–1971. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cao X, Pfaff SL and Gage FH: YAP regulates
neural progenitor cell number via the TEA domain transcription
factor. Genes Dev. 22:3320–3334. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu-Chittenden Y, Huang B, Shim JS, et al:
Genetic and pharmacological disruption of the TEAD-YAP complex
suppresses the oncogenic activity of YAP. Genes Dev. 26:1300–1305.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ota M and Sasaki H: Mammalian Tead
proteins regulate cell proliferation and contact inhibition as
transcriptional mediators of Hippo signaling. Development.
135:4059–4069. 2008. View Article : Google Scholar : PubMed/NCBI
|