Introduction
The invasion and metastasis of cancer cells are
known to be primary causes of cancer progression (1). When tumor cells metastasize, a number
of proteolytic enzymes contribute to the degradation of ECM
components and the basement membrane (2,3).
Metalloproteinases (MMPs) play an essential role in the process and
promotion of tumor invasion and metastasis in many types of cancer
(4).
Among the previously reported MMPs, MMP-9 and MMP-2
are key enzymes for degrading ECM and type IV collagen (5,6). MMP-9
correlates with malignant phenotypes in various types of cancer and
can be activated by a variety of stimuli such as cytokines and
phorbol myristate acetate (PMA) during varied pathological
processes (7,8). The MMP-9 promoter region contains one
NF-κB and two AP-1 binding sites (9–11),
which are absent in the MMP-2 promoter region. Therefore, the
activation of MMP-9 in cancer progression may be derived in part
from its modulation by AP-1 and NF-κB transcription factors in
response to extracellular stimuli.
Ginseng has become one of the most commonly used
alternative herbal medicines, and ginsenoside Rg1 is one of its
most active and abundant components. Ginsenoside Rg1 has
pharmacological effects in the central nervous, cardiovascular, and
immune systems, and also exerts anticancer properties (12–20).
However, the effect of ginsenoside Rg1 on cancer metastasis remains
to be investigated. In the present study, we demonstrated the
effects of ginsenoside Rg1 on PMA-induced invasion and migration in
breast cancer and examine the potential mechanism involved in these
effects. Our results support a model by which ginsenoside Rg1
inhibits MMP-9 expression by suppressing NF-κB activation to
inhibit PMA-induced invasion and migration in MCF-7 cells.
Materials and methods
Materials
Ginsenoside Rg1 was purchased from Shanghai Yaji
(Group) Co., Ltd. (Shanghai, China). Sulforhodamine B (SRB),
trichloroacetic acid (TCA), acetic acid, anti-β-actin and dimethyl
sulfoxide (DMSO) were obtained from Sigma-Aldrich (St. Louis, MO,
USA). TRIzol and cell culture reagents were purchased from
Invitrogen Life Technologies, (Carlsbad, CA, USA). Antibodies for
phospho-p65, phospho-c-jun and MMP-9 were obtained from Cell
Signaling. Secondary antibodies for western blotting were obtained
from Amersham Biosciences Corporation (Piscataway, NJ, USA). Other
reagents were obtained from Sigma-Aldrich unless stated
otherwise.
Sulforhodamine B (SRB) assay
Cytotoxicity was determined by the SRB assay. Cells
were seeded into 96-well plates and exposed to different
concentrations (50, 100, 200 and 400 μM) of ginsenoside Rg1. After
48 h of incubation, the cells were fixed with TCA for 1 h at 4°C,
air-dried, and then stained with 0.4% SRB solution for 30 min at
room temperature. After staining, the SRB solution was removed, and
the cells were subsequently washed five times with 1% acetic acid.
Then, 10 mM Tris base solution (pH 10.5) was added to dissolve the
protein-bound dye, and plates were incubated on a plate shaker for
10 min. The OD570 nm was determined using a 96-well plate reader
(MRX; Dynex Technologies, Chantilly, VA, USA).
Western blotting
MCF-7 cells were seeded in 6-well plates and exposed
to the indicated concentrations of ginsenoside Rg1 with or without
PMA. After treatment, the cells were harvested using lysis buffer
(pH 7.4, 20 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1%
Triton, 2.5 mM sodium pyrophosphate, 1 mM
Na3VO4, 1 mM β-glycerophosphate, 1:1,000
protease inhibitors). Protein concentrations were determined by the
BCA method. Total protein (25 μg) was then separated using 8–12%
sodium dodecyl sulfate-polyacrylamide gels and transferred to
nitrocellulose blotting membranes. The membranes were probed with
monoclonal antibodies against MMP-9, MMP-2, TIMP-1 or β-actin
(1:8,000), and immunopositive bands were visualized using the
Amersham ECL™ Plus Western Blotting Detection kit (GE Healthcare,
Piscataway, NJ, USA).
Electrophoretic mobility shift assay
(EMSA) (21,22)
Biotin 3′ end-labeled DNA probes containing the
NF-κB consensus site: sense 3′-TCAACTCCCCTGAAAGGGTCCG-5′ and
antisense 5′-AGTTGAGGGGACTTTCCCAGGC-3′, were purchased from
Invitrogen (Shanghai, China). EMSA was performed using the light
shift chemiluminescent EMSA kit (Pierce, Rockford, IL, USA).
Briefly, the nuclear proteins were incubated in 1X binding buffer,
50 ng/μl poly (dI•dC), 0.05% NP-40, 5 mM MgCl2, 50 mM
KCl, 2.5% glycerol and ddH2O for 20 min at room
temperature in a total volume of 20 μl. The reaction mixture was
separated on a 6% non-denaturing polyacrylamide gel and transferred
to a positively charged nylon membrane. The membrane was
cross-linked, and the biotin-labeled DNA was detected by
chemiluminescence.
Wound-healing assay
MCF-7 cells were seeded in a 6-well plate and
incubated until they reached 80% confluence. A 200 μl pipette tip
was used to create a wound, and the cells were washed twice with
serum-free culture medium to remove floating cells and then
replaced with fresh medium without serum. The cells were subjected
to the indicated treatment for 24 h, and cells migrating from the
leading edge were photographed at 0 and 24 h.
Matrigel invasion assay
The invasion of MCF-7 cells was performed in a
24-well Transwell unit (8-μM pore size), which was coated with 1
mg/ml Matrigel matrix as previously described (23). Briefly, MCF-7 cells were placed on
the Matrigel-coated Transwell (the upper compartment of the
invasion chamber) in the presence or absence of Rg1 and PMA.
Conditioned medium (500 μl) was added to the lower compartment of
the invasion chamber. After incubation at 37°C for 48 h, the cells
that had invaded the lower surface of the membrane were fixed with
methanol and stained with hematoxylin and eosin. Random fields were
counted by light microscopy.
RNA extraction and reverse transcription
polymerase chain reaction (RT-PCR)
TRIzol reagent was used to extract total RNA
according to the manufacturer’s instructions. Complementary DNA
(cDNA) was created from 1 μg RNA using standard procedures with
avian myeloblastosis virus reverse transcriptase (Promega). For
polymerase chain reaction (PCR) quantification, 2 μl of cDNA was
amplified in a 20 μl standard PCR reaction. PCR was carried out by
initial denaturation at 94°C for 3 min; 36 cycles of 94°C for 45
sec, 55°C for 45 sec, 72°C for 45 sec; and a final extension for 10
min at 72°C, followed by termination at 4°C. RT-PCR was performed
using the following primer pairs (Invitrogen) for semi-quantitative
assessment: sense 5′-TCCCTGGAGACC TGAGAA-3′ and antisense
5′-CGGCAAGTCTTCCGA GTAGTT-3′ for MMP-9; sense 5′-TGAGCTCCCGGAAAA
GATTG-3′ and antisense 5′-TCA GCAGCCTAGCCAGTCG-3′ for MMP-2; sense
5′-GGGGCTTCA CCAAGACCTACAC-3′ and antisense
5′-AAGAAAGATGGGAGTGGGAACA-3′ for TIMP-1; sense
5′-CGTGGACATCCGCAAAGAC-3′ and anti-sense 5′-GCATTTGCGGTGGACGAT-3′
for β-actin. β-actin transcript served as an internal control for
standardising the quantity of input cDNA. The PCR products were
separated by electrophoresis on a 1.5% agarose gel and visualised
under UV light using a gel documentation system (Bio-Rad, Hercules,
CA, USA).
Transient transfection and luciferase
reporter assay
To determine promoter activity, MCF-7 cells were
seeded in 6-well plates. At 70–80% confluence, the cells were
cotransfected with pCMA-β-galactosidase plasmid along with
pGL2-MMP-WT, pGL2-MMP-9-mAP-1–2 or pGL2-MMP-9-mNF-κB. The
transfected cells were subsequently treated with ginsenoside Rg1
and stimulated with PMA. Following incubation for 2 h, the cells
were lysed and luciferase activity was measured using a luminometer
(Luminoscan Ascent; Thermo Electon Co., Germany).
Zymography assay
Zymography was performed as previously described
(23,24). Briefly, cells were incubated in
serum-free RPMI-1640 and the supernatants were collected after
incubation for 24 h. Conditioned media were collected, centrifuged
and electrophoresed at 4°C on 10% SDS-PAGE containing 1 mg/ml
gelatin. The gels were washed with 2.5% Triton X-100 and then
incubated at 37°C for 24 h in a buffer containing 5 mM
CaCl2, 50 mM Tris-HCl and 1 μM ZnCl2. The
gels were stained with 0.05%Coomassie Brilliant Blue R-250 for 30
min at room temperature. The gelatinolytic activity of MMP-9 was
observed as a white zone in a dark blue field.
Transfection of siRNA
Human breast cancer MCF-7 cells were obtained from
ATCC (Manassas, VA, USA) and seeded at 2×105 cells/ml in
6-well plates. The cell plates were grown to 50% confluency and
transfected with double-stranded siRNA for NF-κB p65 target
sequence: (sense, 5′-CUUCCAAGUUCCUAUAGAAdTdT-3′ and antisense,
3′-dTdTGAAGGUU CAAGGAUAUCUU-5′) or with a siRNA non-specific
control (Guangzhou RiboBio Co., Ltd., Guangzhou, China). Silencing
was confirmed by western blotting and EMSA.
Statistical analysis
Statistical analysis between groups was performed
using an unpaired Student’s t-test with SigmaPlot 10.0 software
(Jandel Scientific, San Rafael, CA, USA). Data are presented as
means ± SEM. P<0.05 was considered to indicate a statistically
significant result.
Results
Ginsenoside Rg1 inhibits cell invasion
and migration of MCF-7 cells
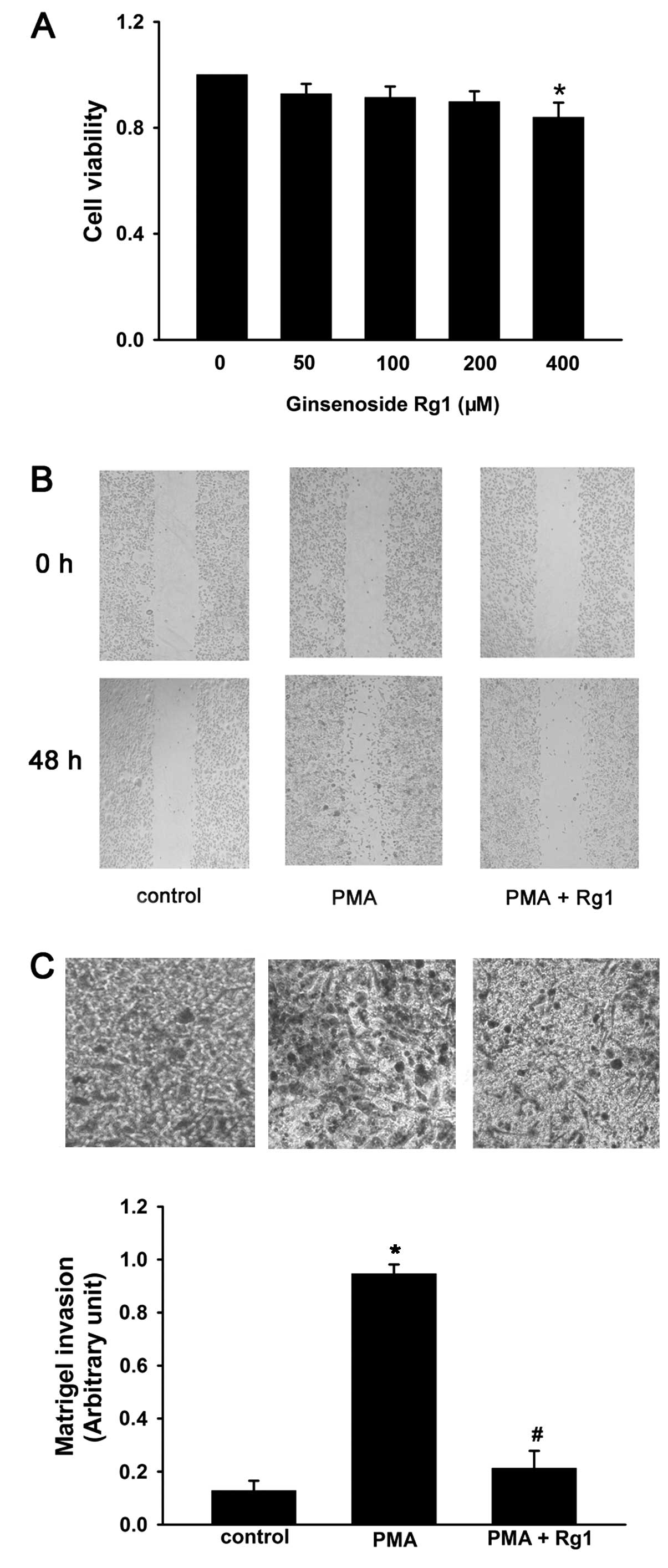
To observe the effect of ginsenoside Rg1 on cell
viability in MCF-7 cells, the cells were treated with increasing
concentrations of ginsenoside Rg1 (50–400 μM) for 48 h and then
assessed by SRB assay. Ginsenoside Rg1 had no effect on cell
viability up to a concentration of 400 μM. Thus, the non-cytotoxic
concentrations of ginsenoside Rg1, from 50 to 200 μM were used in
subsequent experiments (Fig.
1A).
To investigate the effects of ginsenoside Rg1 on
PMA-induced cell invasion and migration, Transwell and
wound-healing assays were performed. The cells were pretreated with
50 nM PMA for 24 h and then exposed to ginsenoside Rg1 (50–200 μM)
for 48 h. PMA significantly increased the invasion and migration of
MCF-7 cells as compared with PMA-untreated control cells, while
ginsenoside Rg1 reversed this effect (Fig. 1B and C). These results suggested
that ginsenoside Rg1 may constitute an effective inhibitor of the
invasion and migration in MCF-7 cells.
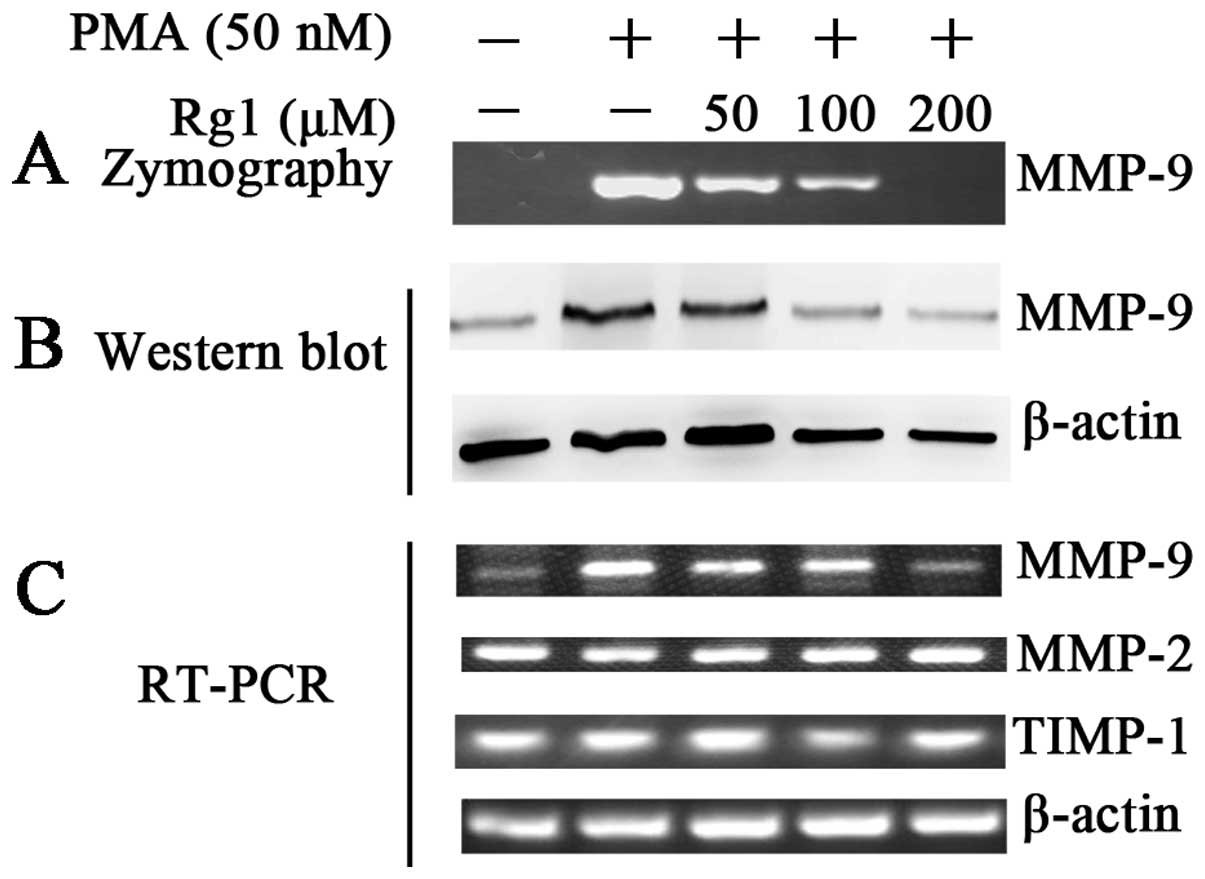
Ginsenoside Rg1 suppresses MMP-9
secretion by inhibiting its protein and mRNA expression
MMP-9 and MMP-2 are important ECM-degrading enzymes
that are involved in cancer invasion and metastasis (5–8).
Zymography assay, western blotting and RT-PCR were used to
determine the effects of ginsenoside Rg1 on PMA-induced MMP-9
activity, and the protein and mRNA expression (Fig. 2). The results showed that
ginsenoside Rg1 suppressed PMA-induced MMP-9 activity in a
dose-dependent manner, which may be explained by decreased levels
of MMP-9 protein and mRNA. Ginsenoside Rg1 did not affect MMP-2
mRNA expression suggesting specificity for MMP-9. Since the
activity of MMPs is regulated by the endogenous inhibitor TIMP-1,
RT-PCR was used to determine the mRNA levels of TIMP-1. However,
the mRNA levels of TIMP-1 were not significantly affected by
ginsenoside Rg1 (Fig. 2C). This
suggested that ginsenoside Rg1 suppressed PMA-induced MMP-9
secretion by affecting its transcription levels. However, the
regulation of MMP-9 was not likely to occur at the level of
regulation of TIMP-1 mRNA.
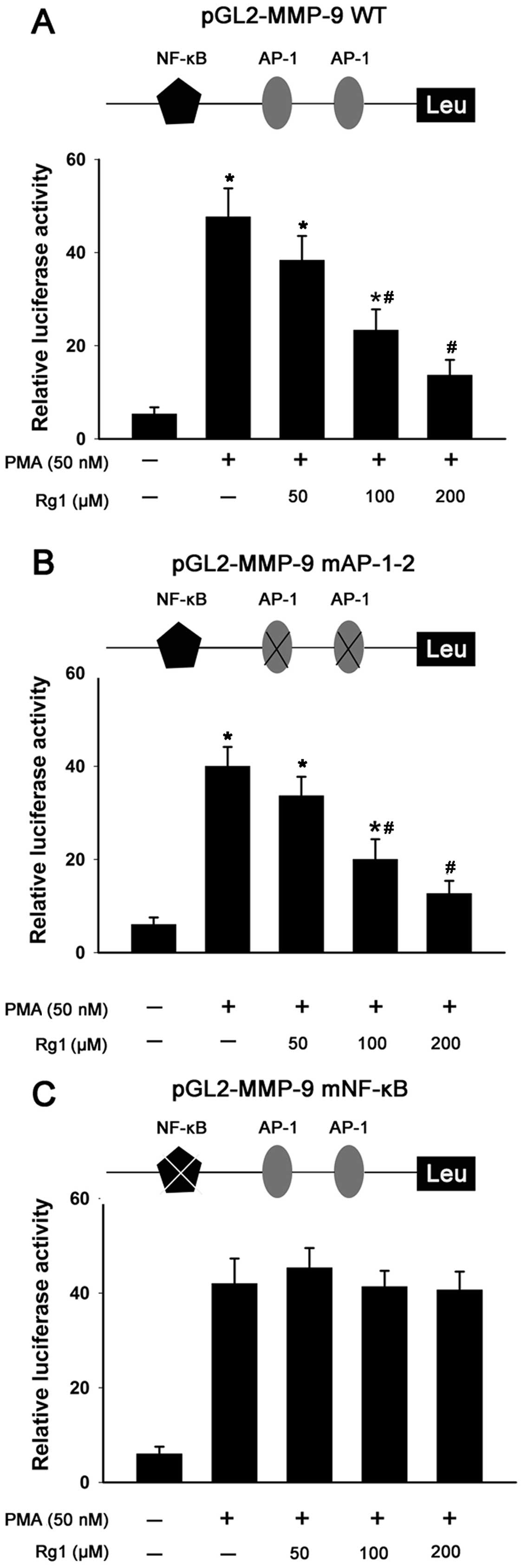
Ginsenoside Rg1 inhibits PMA-induced
MMP-9 activity by blocking its promoter activity
AP-1 and NF-κB transcriptional elements in the MMP-9
promoter are important in the regulation of its expression. To
investigate the effect of ginsenoside Rg1 on PMA-induced MMP-9
promoter activity and the possible role of these elements, MCF-7
cells were transiently transfected with a wt-MMP-9 promoter
luciferase reporter construct and reporter constructs with
mutations in the AP-1 and NF-κB sites (MMP-9-mAP-1–2 or
MMP-9-mNF-κB). The transcriptional activity of the wt-MMP-9
promoter reporter gene was activated up to 14-fold in cells by PMA.
However, ginsenoside Rg1 significantly decreased the PMA-induced
transcriptional activity of the reporter gene (Fig. 3A). Consistent with this result, the
cells that received MMP-9-mAP-1–2 also demonstrated abrogated
PMA-induced MMP-9 activity with ginsenoside Rg1 treatment (Fig. 3B). In contrast to the wt-MMP-9 and
MMP-9-mAP-1–2, when cells were transfected with reporter plasmid
containing the MMP-9-mNF-κB promoter, the luciferase activity of
the cells induced by PMA was not affected by ginsenoside Rg1
(Fig. 3C). These results suggested
that ginsenoside Rg1 inhibited PMA-induced MMP-9 activity by
suppressing the transcriptional activity of NF-κB.
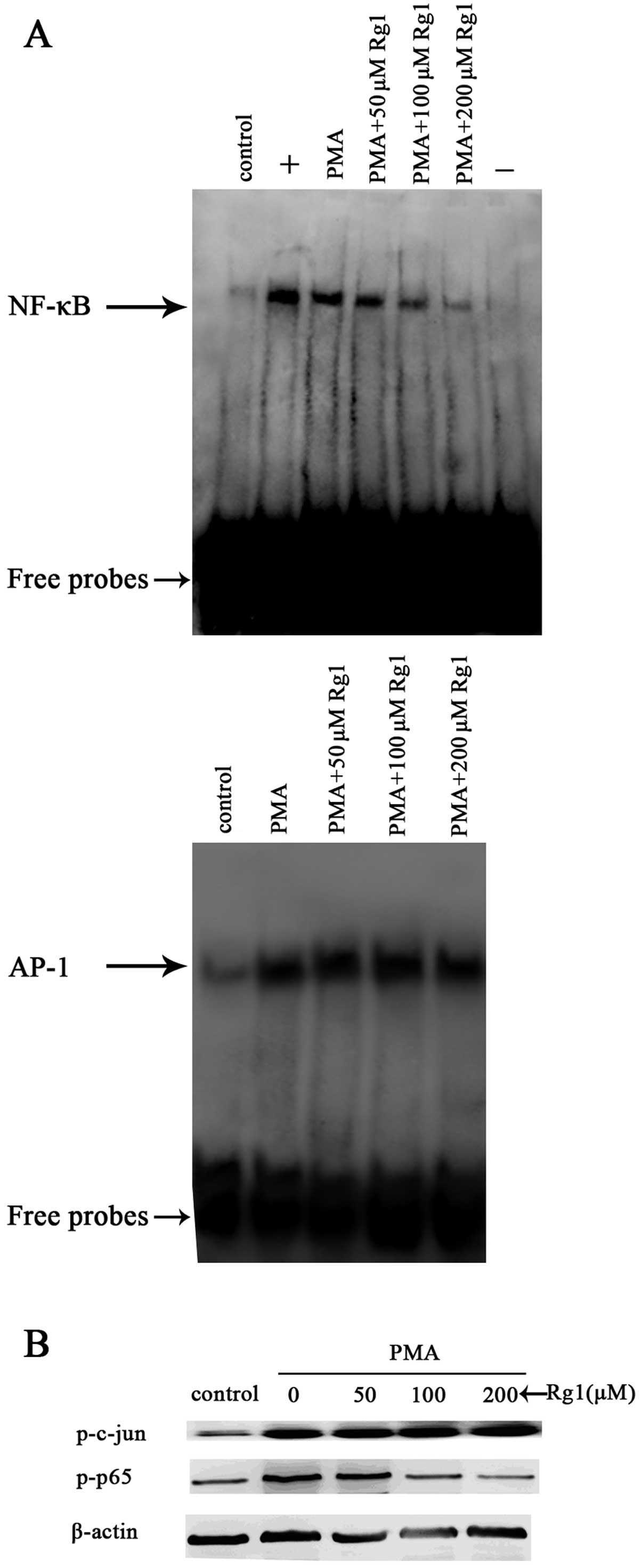
Ginsenoside Rg1 inhibits NF-κB activation
induced by PMA
To investigate the effect of ginsenoside Rg1 on
PMA-induced NF-κB and AP-1 activity, we performed EMSA using
binding elements from the MMP-9 promoter. MCF-7 cells were
pretreated with ginsenoside Rg1 for 48 h and then exposed to 50 nM
PMA for 2 h. PMA caused a modest, yet reproducible enhancement of
the DNA binding activity of NF-κB, whereas ginsenoside Rg1
suppressed this activity in a dose-dependent manner (Fig. 4A, top panel). Conversely, although
AP-1 was induced by PMA, ginsenoside Rg1 had no effect on
PMA-induced AP-1 activity. These results suggested that ginsenoside
Rg1 regulates the transcriptional activity of MMP-9 by inhibiting
PMA-induced NF-κB, but not AP-1.
To confirm these results, the effect of ginsenoside
Rg1 on the PMA-induced phosphorylation of p65, a major subunit of
NF-κB; and c-jun, a major subunit of AP-1, were investigated by
western blotting. Consistent with the EMSA data, the two proteins
were phosphorylated in response to PMA. Furthermore, ginsenoside
Rg1 suppressed the PMA-induced phosphorylation of p65 expression in
a dose-dependent manner. However, it did not affect the
phosphorylation of c-jun (Fig. 4B).
These results are consistent with the possibility that NF-κB
functions as a target of the inhibitory effects of ginsenoside Rg1
on MMP-9 expression.
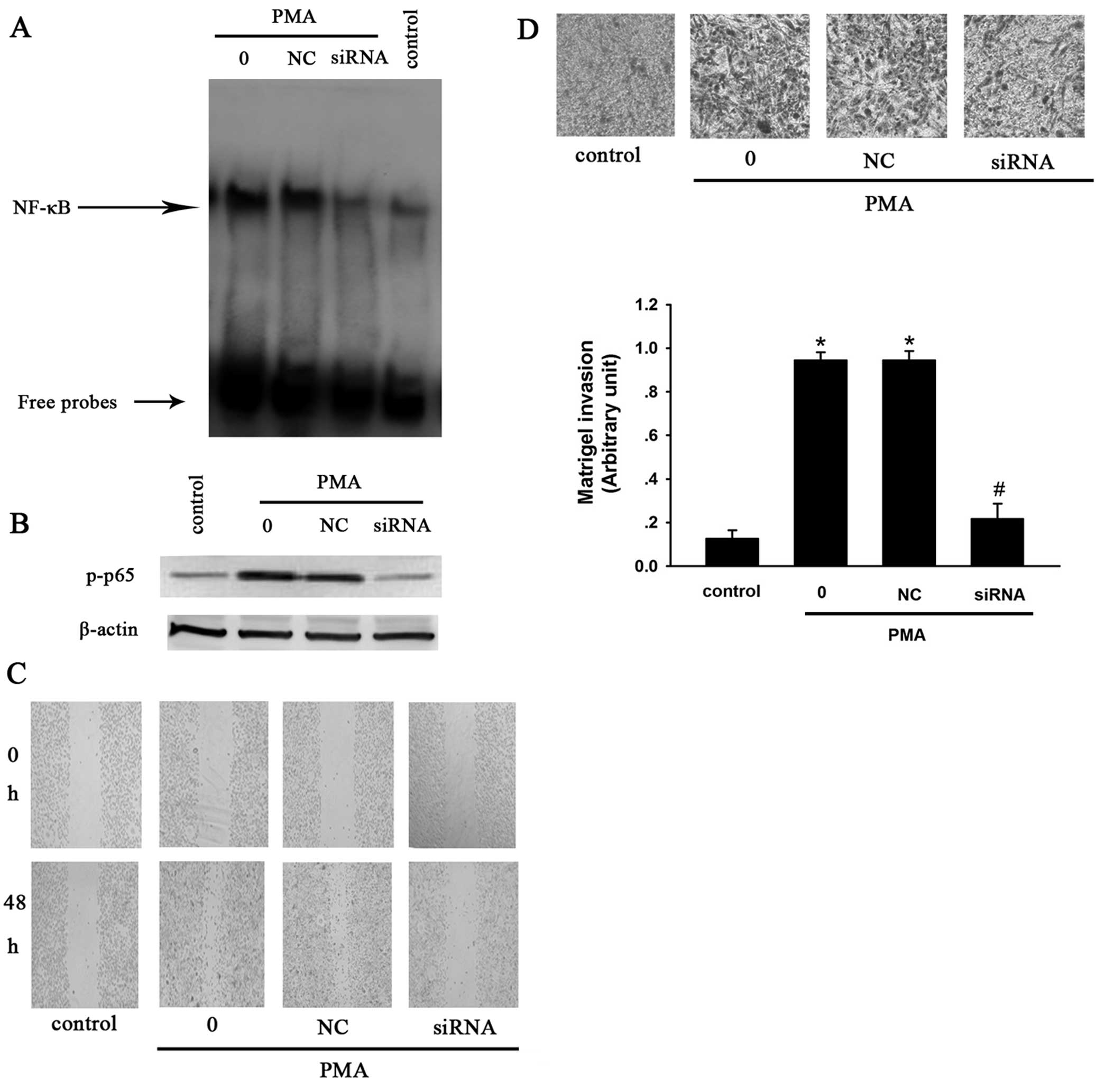
P65 suppresses siRNA cell invasion and
migration of MCF-7 cells
To confirm that PMA-induced MMP-9 expression is
NF-κB-dependent, p65 siRNA was used to block the activation of
NF-κB in the present study. MCF-7 cells were transfected with p65
siRNA for 48 h, followed by treatment with 50 nM PMA for 2 h.
Western blotting and EMSA were used to investigate the expression
of p-p65 and the activity of NF-κB. The results of the western
blotting showed that PMA enhanced the expression of p-p65, while
p65 siRNA reversed this effect. Consistent with the downregulation
of p-p65 expression, NF-κB activation was shown by EMSA to be
significantly inhibited by the siRNAs (Fig. 5A).
Transwell and wound-healing assays were subsequently
performed to observe the effect of p65 siRNA on cell invasion and
migration. The results demonstrated that PMA significantly promoted
the invasion and migration of MCF-7 cells. However, p65 siRNA
suppressed PMA-induced cell invasion and migration relative to the
siRNA-negative controls (Fig.
5).
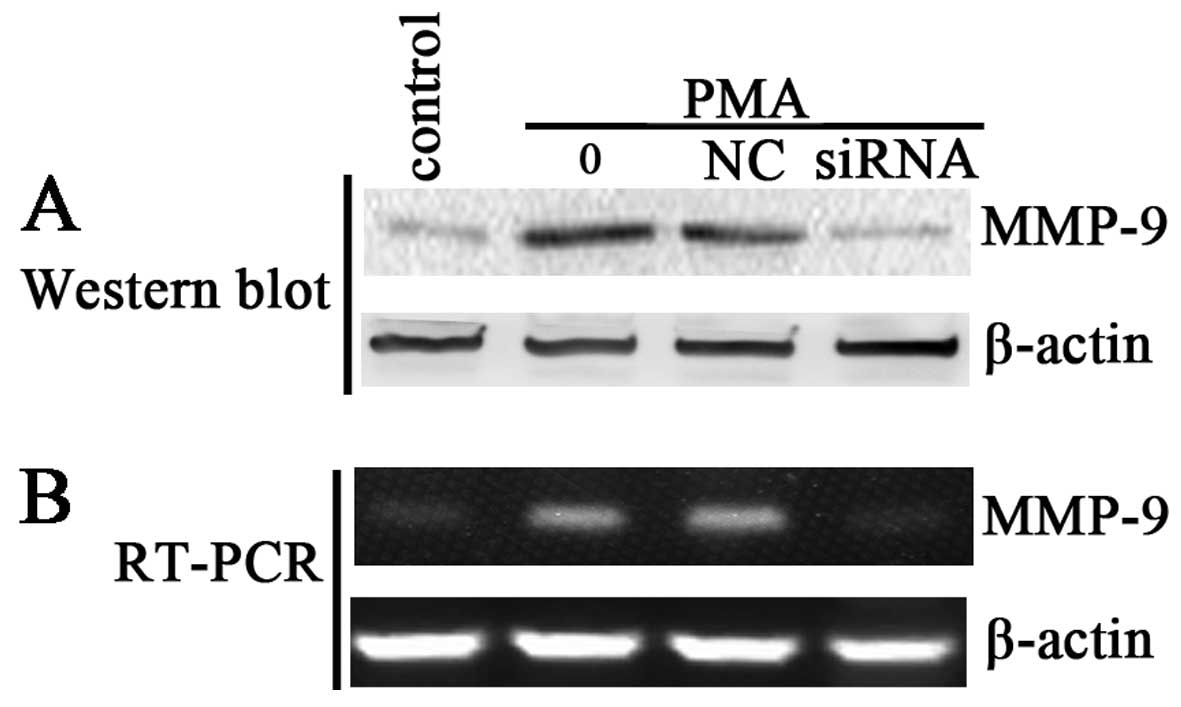
P65 siRNA inhibits MMP-9 protein and mRNA
expression by blocking its promoter activity
The above results have demonstrated that ginsenoside
Rg1 may suppress PMA-induced invasion and migration by inhibiting
NF-κB-dependent MMP-9 expression. Thus, in the present study we
detected the effect of p65 siRNA on the protein and mRNA expression
of MMP-9. Results of western blotting and RT-PCR showed that p65
siRNA significantly inhibited the PMA-induced levels of MMP-9
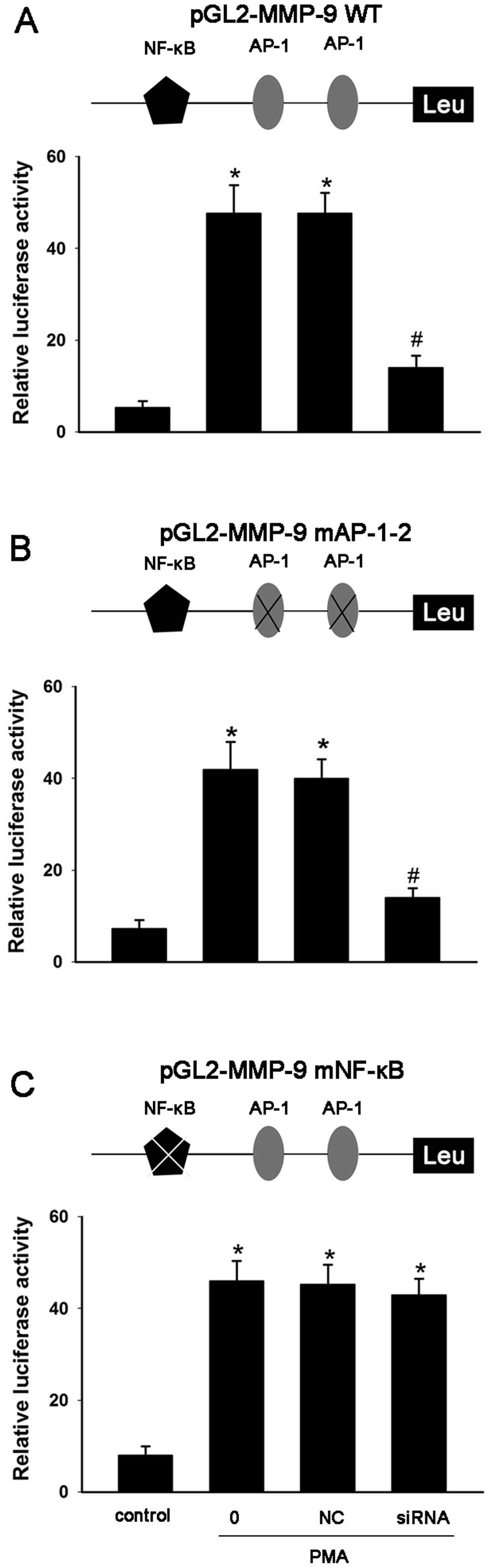
protein and mRNA (Fig. 6). The
effect of p65 siRNA on the promoter activity of MMP-9 was also
investigated. MCF-7 cells were transiently transfected with a
wt-MMP-9 promoter luciferase reporter construct and reporter
constructs with mutations in the AP-1 and NF-κB sites
(MMP-9-mAP-1–2 or MMP-9-mNF-κB). Consistent with the effect of
ginsenoside Rg1, the results showed that p65 siRNA significantly
inhibited PMA-induced MMP-9 activity by suppressing the
transcriptional activity of NF-κB, but had no effect on the
transcriptional activity of AP-1 (Fig.
7). Thus, the siRNA studies showed that PMA-induced MMP-9
expression is NF-κB-dependent. These results confirmed that
ginsenoside Rg1 suppresses PMA-induced tumor cell invasion and
migration by inhibiting NF-κB-dependent MMP-9 expression.
Discussion
In the present study, we have demonstrated that
ginsenoside Rg1 effectively suppresses PMA-induced invasion and
migration. Ginsenoside Rg1 inhibits MMP-9 secretion by inhibiting
its protein and mRNA expression, and since MMP-9 is known to act as
a key regulator of metastatic function, it is likely that the
suppressive effects of ginsenoside Rg1 on invasion and metastasis
are mediated through its inhibition of MMP-9. Our results also show
that ginsenoside Rg1 inhibits PMA-induced NF-κB activation in a
dose-dependent manner, which may explain the transcriptional
effects on MMP-9 mRNA. The siRNA studies show that PMA-induced
MMP-9 expression is NF-κB-dependent. These results suggest that
ginsenoside Rg1 serves as a potential inhibitor in preventing the
invasion and metastasis of human breast cancers.
Invasion and metastasis are fundamental properties
of cancer cells, and their control is therefore an important
therapeutic goal. The identification of novel candidate agents that
inhibit these processes is essential for preventing the progression
of malignant tumors. In the present study, we investigated the
effect of ginsenoside Rg1 on invasion and migration in MCF-7 breast
cancer cells. PMA was used to induce cell invasion and migration
based on its well-characterized role as a tumor promoter in
chemical-induced carcinogenesis in many cancer cells, including
hepatoma, colon, glioma and breast cancer cells (25–27).
The Transwell and wound-healing assays demonstrated that PMA
significantly induces invasion and migration in MCF-7 cells, while
ginsenoside Rg1 suppresses these processes. These results suggest
that ginsenoside Rg1 constitutes an effective inhibitor of cancer
cell progression.
The enhanced expression of MMP-9 and/or MMP-2 has
been shown to be responsible for PMA-induced cell invasion and
migration (5–8). To investigate how ginsenoside Rg1
suppresses PMA-induced cell invasion and migration, we assessed the
effects of ginsenoside Rg1 on PMA-induced MMP-9 by western blotting
and RT-PCR. Ginsenoside Rg1 suppressed the PMA-induced MMP-9
protein and mRNA expression in a dose-dependent manner. However,
ginsenoside Rg1 did not affect MMP-2 mRNA expression, suggesting a
specificity of this response. The mRNA levels of TIMP-1 were also
not affected by ginsenoside Rg1, suggesting that the suppression of
PMA-induced MMP-9 is not mediated through TIMP-1.
However, one potential pathway of the regulation of
MMP-9 is suggested by the results of the EMSA and western blot
analyses. The results show that ginsenoside Rg1 suppresses
PMA-induced NF-κB DNA-binding activity and the phosphorylation of
p65, a major component of NF-κB that is known to translocate to the
nucleus following phosphorylation and transactivate the promoters
of multiple cancer-related genes, including MMP-9. Conversely, AP-1
DNA-binding activity and the PMA-induced phosphorylation of its
c-jun subunit were not inhibited by ginsenoside Rg1, suggesting
that the effects of ginsenoside Rg1 are limited to NF-κB. PMA is
known to enhance the expression of MMP-9 through one NF-κB and two
AP-1 binding sites within its promoter region (9–11). In
the present study, the mutational analysis of the minimal MMP-9
promoter demonstrated a requirement for NF-κB, but not AP-1 for the
suppression by ginsenoside Rg1. The siRNA studies also verified
that PMA-induced MMP-9 expression is NF-κB-dependent. The
above-mentioned results suggest a potential model whereby
ginsenoside Rg1 inhibits PMA-induced MMP-9 activity through NF-κB
to suppress breast cancer cell migration and invasion. Thus,
ginsenoside Rg1 constitutes a potential antimetastatic and
anti-invasive agent that may be useful in future clinical studies
against cancer.
Acknowledgements
This study is supported by the China Postdoctoral
Science Foundation (no. 20090461139), the National Natural Science
Foundation of China (no. 81001457), and the Foundation of Bengbu
Medical College (nos. Bykf13A11 and Byycx1329).
References
|
1
|
Weng CJ, Chau CF, Hsieh YS, Yang SF and
Yen GC: Lucidenic acid inhibits PMA-induced invasion of human
hepatoma cells through inactivating MAPK/ERK signal transduction
pathway and reducing binding activities of NF-κB and AP-1.
Carcinogenesis. 29:147–156. 2008.PubMed/NCBI
|
|
2
|
Kato Y, Yamashita T and Ishikawa M:
Relationship between expression of matrix metalloproteinase-2 and
matrix metalloproteinase-9 and invasion ability of cervical cancer
cells. Oncol Rep. 9:565–569. 2002.PubMed/NCBI
|
|
3
|
Liotta LA, Steeg PS and Stetler-Stevenson
WG: Cancer metastasis and angiogenesis: an imbalance of positive
and negative regulation. Cell. 64:327–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rao JS: Moleculae mechanisms of glioma
invasiveness: the role of proteases. Nat Rev Cancer. 3:489–501.
2003. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chambers AF and Matrisian LM: Changing
views of the role of matrix metalloproteinases in metastasis. J
Natl Cancer Inst. 89:1260–1270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nabeshima K, Inoue T, Shimao Y and
Sameshima T: Matrix metalloproteinases in tumor invasion: Role for
cell migration. Pathol Int. 52:255–264. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yan C and Boyd DD: Regulation of matrix
metalloproteinase gene expression. J Cell Physiol. 211:19–26. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cho HJ, Kang JH, Kwak JY, Lee TS, Lee IS,
Park NG, Nakajima H, Magae J and Chang YC: Ascofuranone suppresses
PMA-mediated matrix metalloproteinase-9 gene activation through the
Ras/Raf/MEK/ERK-and Ap1-dependent mechanisms. Carcinogenesis.
28:1104–1110. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sato H and Seiki M: Regulatory mechanism
of 92 kDa type IV collagenase gene expression which is associated
with invasiveness of tumor cells. Oncogene. 8:395–405.
1993.PubMed/NCBI
|
|
10
|
Sato H, Kita M and Seiki M: v-Src
activates the expression of 92-kDa type IV collagenase gene through
the AP-1 site and the GT box homologous to retinoblastoma control
elements. A mechanism regulating gene expression independent of
that by inflammatory cytokines. J Biol Chem. 268:23460–23468.
1993.
|
|
11
|
Takahra T, Smart DE, Oakley F and Mann DA:
Induction of myofibroblast MMP-9 transcription in three-dimensional
collagen I gel cultures: regulation by NF-κB, AP-1 and Sp1. Int J
Biochem Cell Biol. 36:353–363. 2004.PubMed/NCBI
|
|
12
|
Attele AS, Wu JA and Yuan CS: Ginseng
pharmacology: multiple constituents and multiple actions. Biochem
Pharmacol. 58:1685–1693. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xu L, Chen WF and Wong MS: Ginsenoside Rg1
protects dopaminergic neurons in a rat model of Parkinson’s disease
through the IGF-I receptor signalling pathway. Br J Pharmacol.
158:738–748. 2009.PubMed/NCBI
|
|
14
|
Liu Q, Kou JP and Yu BY: Ginsenoside Rg1
protects against hydrogen peroxide-induced cell death in PC12 cells
via inhibiting NF-κB activation. Neurochem Int. 58:119–125.
2011.PubMed/NCBI
|
|
15
|
Li CY, Deng W, Liao XQ, Deng J, Zhang YK
and Wang DX: The effects and mechanism of ginsenoside Rg1 on
myocardial remodeling in an animal model of chronic thromboembolic
pulmonary hypertension. Eur J Med Res. 18:162013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yin H, Liu Z, Li F, Ni M, Wang B, Qiao Y,
Xu X, Zhang M, Zhang J, Lu H and Zhang Y: Retraction note to:
Ginsenoside-Rg1 enhances angiogenesis and ameliorates ventricular
remodeling in a rat model of myocardial infarction. J Mol Med.
91:6452013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee EJ, Ko E, Lee J, Rho S, Ko S, Shin MK,
Min BI, Hong MC, Kim SY and Bae H: Ginsenoside Rg1 enhances
CD4+ T-cell activities and modulates Th1/Th2
differentiation. Int Immunopharmacol. 4:235–244. 2004. View Article : Google Scholar
|
|
18
|
Qu DF, Yu HJ, Liu Z, Zhang DF, Zhou QJ,
Zhang HL and Du AF: Ginsenoside Rg1 enhances immune response
induced by recombinant Toxoplasma gondii SAG1 antigen. Vet
Parasitol. 179:28–34. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li QF, Shi SL, Liu QR, Tang J, Song J and
Liang Y: Anticancer effects of ginsenoside Rg1, cinnamic acid, and
tanshinone IIA in osteosarcoma MG-63 cells: nuclear matrix
downregulation and cytoplasmic trafficking of nucleophosmin. Int J
Biochem Cell Biol. 40:1918–1929. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu J, Cai SZ, Zhou Y, Zhang XP, Liu DF,
Jiang R and Wang YP: Senescence as a consequence of ginsenoside rg1
response on k562 human leukemia cell line. Asian Pac J Cancer Prev.
13:6191–6196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hussain AR, Ahmed SO, Ahmed M, Khan OS, Al
Abdulmohsen S, Platanias LC, Al-Kuraya KS and Uddin S: Cross-talk
between NFκB and the PI3-kinase/AKT pathway can be targeted in
primary effusion lymphoma (PEL) cell lines for efficient apoptosis.
PLos One. 7:e399452012.
|
|
22
|
Hussain AR, Ahmed M, Al-Jomah NA, Khan AS,
Manogaran P, Sultana M, Abubaker J, Platanias LC, Al-Kuraya KS and
Uddin S: Curcumin suppresses constitutive activation of nuclear
factor-κB and requires functional Bax to induce apoptosis in
Burkitt’s lymphoma cell lines. Mol Cancer Ther. 7:3318–3329.
2008.PubMed/NCBI
|
|
23
|
Kim KS, Yao L, Lee YC, Chung E, Kim KM,
Kwak YJ, Kim SJ, Cui Z and Lee JH: Hyul-Tong-Ryung suppresses
PMA-induced MMP-9 expression by inhibiting AP-1-mediated gene
expression via ERK1/2 signaling pathway in MCF-7 human breast
cancer cells. Immunopharmacol Immunotoxicol. 32:600–606. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yao L, Kim KS and Kang NY: Inhibitory
effect of a traditional Chinese formulation, Hyul-Tong-Ryung, on
PMA-induced MMP-9 expression in MCF-7 human breast carcinoma cells.
J Trad Med. 26:25–34. 2009.
|
|
25
|
Han SY, Lee MS, Kim HR, Baek SH, Ahn DH,
Chae HS, Erickson RH, Sleisenger MH and Kim YS: Phorbol
12-myristate 13-acetate induces alteration in mucin gene expression
and biological properties of colon cancer cells. Int J Oncol.
17:487–494. 2000.PubMed/NCBI
|
|
26
|
Lin CW, Shen SC, Hou WC, Yang L and Chen
YC: Heme oxygenase-1 inhibits breast cancer invasion via
suppressing the expression of matrix metalloproteinase-9. Mol
Cancer Ther. 7:1195–1206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scorilas A, Karameris A, Arnogiannaki N,
Ardavanis A, Bassilopoulos P, Trangas T and Talieri M:
Overexpression of matrix-metalloproteinase-9 in human breast
cancer: a potential favourable indicator in node-negative patients.
Br J Cancer. 84:1488–1496. 2001. View Article : Google Scholar
|