Introduction
Lung cancer is the leading cause of cancer-related
death in the world and non-small cell lung cancer (NSCLC) accounts
for more than 85% of all diagnosed lung cancers (1). Although advancements have been made in
early detection, prevention and treatment options (such as
molecular targeting therapy), the overall survival of lung cancer
has not significantly improved. Advanced lung cancers are
associated with a high rate of metastasis and limited treatment
options, all of which contribute to the low 5-year survival rate.
Importantly, a large number of patients are still being diagnosed
at advanced stages of the disease, which excludes surgery as an
option for their treatment. Thus, novel approaches to facilitate
early detection and identification and evaluation of prognostic
markers could help physicians to more efficiently manage lung
cancer patients. At the molecular level, development of lung
cancer, similar to most other cancers, involves the activation of
oncogenes and inactivation of tumor-suppressor genes and to date, a
great number of gene alterations have been identified as being
associated with lung carcinogenesis.
One of the molecules involved in tumorigenesis is
liver kinase B1 (LKB1, also known as serine/threonine kinase 11,
STK11), which is a member of the serine/threonine kinase family and
functions to regulate cell polarity. LKB1 was first reported as the
causative gene of Peutz-Jeghers syndrome (2,3)
characterized by multiple gastrointestinal polyps, mucocutaneous
hyperpigmentation and increased cancer risk. The LKB1 protein
contains 433 amino acids, a nuclear location signal in the
N-terminal non-catalytic domain and the central catalytic domain
and a C-terminal non-catalytic domain (4,5).
Previous studies have shown that LKB1 functions as a
tumor-suppressor gene by activating a group of other kinases,
including AMP-activated protein kinase (AMPK) (6,7) or
AMPK-related kinases, which in turn regulate cell cycle (8), cell apoptosis, cell polarity and
metabolism (9,10). In lung cancer, particularly NSCLC,
LKB1 was shown to be frequently mutated or inactivated, and
loss of LKB1 function was found to be associated with lung cancer
development, tumor de-differentiation, progression and metastasis
(11).
Alteration of autophagy has been shown to play an
important role in human tumorigenesis. Although autophagy is
considered as an adaptive response to integrated stress, including
nutrient deprivation, reactive oxygen species, or endoplasmic
reticulum stress (12), it acts as
a double-edged sword in cancer. On the one hand, autophagy has
tumor-suppressor effects, whereas on the other hand, it can promote
tumor survival in response to cellular stress (13). There is preclinical evidence for the
improved antitumor effect of chemotherapy when combined with
autophagy inhibitors (14–17). In this regard, Beclin1 is a crucial
modifier of autophagy and a component of the class III
phosphatidylinositol 3-kinase (PI3K) complex that is involved in
the activation of autophagy and formation of autophagosomes
(18,19). Previous studies have shown aberrant
expression of Beclin1 in breast, esophageal, gastric and colorectal
cancers (20–22). LKB1 has been demonstrated to be
involved in autophagy and regulates the expression of
autophagy-related molecules in vitro. A number of
preclinical studies have been conducted that demonstrate the
utility of targeting LKB1 and autophagy as a therapeutic strategy
for several human cancers (23–25).
However, the clinicopathological significance of LKB1 and
autophagy-related gene Beclin1 and their relationship in
vivo remain to be determined. In the present study, we
quantified expression of LKB1 and Beclin1 in NSCLC tissues and
defined their association with clinicopathological features and
survival of the patients.
Materials and methods
Patients
We collected tissue samples from 142 NSCLC patients
who underwent surgery between January 2008 and October 2012 at The
First Affiliated Hospital of Xi’an Jiaotong University. The
patients were diagnosed with NSCLC based on pathology and did not
receive any chemotherapy and/or radiotherapy before surgery. The
patient cohort consisted of 82 male and 60 female patients with an
average age of 58.2 years (range, 31–84 years). Tumor tissues and
matched surrounding normal tissues (at least 5 cm away from the
tumor lesions) were obtained from the Pathology Department. All of
the specimens were examined and evaluated by two independent
pathologists. Clinicopathological data were collected from the
patient medical records and are shown in Table I. Tumor staging was evaluated
according to the 7th edition of the AJCC cancer staging manual
(26). The last follow-up date of
all patients was recorded in February 2014. For survival analysis,
the survival data were censored on the date of the last follow-up
or death from causes other than NSCLC. This study was approved by
the Medical Ethics Committee of The First Affiliated Hospital of
Xi’an Jiaotong University, and informed consent was obtained from
each patient.
 | Table IAssociation of LKB1 and Beclin1
protein expression with clinicopathological parameters of the NSCLC
patients. |
Table I
Association of LKB1 and Beclin1
protein expression with clinicopathological parameters of the NSCLC
patients.
| | LKB1
expression | | Beclin1
expression | |
|---|
| |
| |
| |
|---|
|
Characteristics | n | Low | High | P-value | Low | High | P-value |
|---|
| All cases | 142 | | | | | | |
| Gender | | | | 0.408 | | | 0.666 |
| Male | 82 | 17 | 65 | | 33 | 49 | |
| Female | 60 | 16 | 44 | | 22 | 38 | |
| Age (years) | | | | 0.721 | | | 0.186 |
| <60 | 77 | 17 | 60 | | 26 | 51 | |
| ≥60 | 65 | 16 | 49 | | 29 | 36 | |
| Histologic
type | | | | <0.001 | | | 0.032 |
| ADC | 83 | 28 | 55 | | 26 | 57 | |
| SQCC | 59 | 5 | 54 | | 29 | 30 | |
|
Differentiation | | | | <0.001 | | | 0.015 |
| Poor | 57 | 23 | 34 | | 29 | 28 | |
| Moderate to
well | 85 | 10 | 75 | | 26 | 59 | |
| Lymph node
metastasis | | | | 0.150 | | | 0.166 |
| No | 80 | 15 | 65 | | 27 | 53 | |
| Yes | 62 | 18 | 44 | | 28 | 34 | |
| TNM stage | | | | 0.075 | | | 0.076 |
| I–II | 107 | 21 | 86 | | 37 | 70 | |
| III–IV | 35 | 12 | 23 | | 18 | 17 | |
Immunohistochemistry
Formalin-fixed paraffin-embedded blocks from normal
and cancerous tissues were cut into 5-μm sections, deparaffinized
and rehydrated in xylene and graded alcohols, respectively. After
antigen retrieval by microwave heating in citric acid buffer, the
sections were incubated with 3% H2O2 for 30
min at room temperature to block endogenous peroxidase activity.
Sections were then incubated with normal serum for 30 min at room
temperature and then further incubated with the primary antibody
against LKB1 (D60C5F10; Cell Signaling Technology, Danvers, MA,
USA) at a dilution of 1:250 and Beclin1 (EPR1733Y; Abcam,
Cambridge, UK) at a dilution of 1:50 overnight at 4°C. On the
following day, the sections were washed three times with
phosphate-buffered saline (PBS) and then incubated with a
biotin-labeled secondary antibody. Sections were washed three times
with PBS and incubated with 3,3′-diaminobenzidine
tetrahydrochloride (DAB) and counterstained with hematoxylin.
Breast cancer tissue sections were used as positive controls, and
the negative controls were incubated with PBS instead of the
primary antibody.
Immunostained tissue sections were then
independently reviewed and scored by two clinical pathologists. The
scores were based on the staining intensity (0–3) and the
percentage of positive-staining tumor cells (0–100%). The staining
intensity was defined as negative (0), weak (1), moderate (2) or
strong (3), and the percentage of positive tumor cells was scored
as 0 (0%), 1 (1–25%), 2 (26–50%), 3 (51–75%) and 4 (76–100%). The
staining intensity and the percentage of positive tumor cells were
multiplied to reach a final score for each case, which ranged
between 0 and 12, with a score of 0–4 being considered as low
expression of either LKB1 or Beclin1 and a score of 5–12 as high
expression of these proteins.
qRT-PCR analysis
Total RNA was extracted from fresh tissues using
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and then reversely
transcribed into cDNA using the SYBR ExScript RT-PCR kit (Takara,
Dalian, China) according to the manufacturers’ instructions. qPCR
amplification was then performed using the iQ5 Multicolor Real-Time
PCR Detection System (Bio-Rad, Hercules, CA, USA) with SYBR Premix
Ex Taq™ II (Takara). The PCR primers for LKB1 and
Beclin1 were designed and synthesized by Takara
Biotechnology Co. and their sequences were LKB1,
5′-AGCATGACTGTGGTG CCGTACT-3′ and 5′-TCCATTGTGACTGGCCTCCTC-3′;
Beclin1, 5′-AACCAACGTCTTTAATGCAACCTTC-3′ and
5′-AGCAGCATTAATCTCATTCCATTCC-3′. Glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) was used as the reference control and
the primers were 5′-ATGGGGAAGGT GAAGGTCG-3′ and
5′-GGGTCATTGATGGCAACAATA TC-3′. All of the samples were performed
in triplicate, and melting curve analysis was conducted to evaluate
the specificity of the amplification. The 2−ΔΔCt method
was adopted to quantify levels of the target genes vs. the
GAPDH gene. The relative expression 2−ΔΔCt was
calculated using the following formula: ΔCt = Ct (target) − Ct
(reference); ΔΔCt = ΔCt (tumor) − ΔCt (normal).
Protein extraction and western
blotting
Total cellular protein for western blotting was
extracted from the fresh tissues using RIPA lysis buffer (Beyotime
Chemical Co., Beijing, China). The protein samples were separated
on 6–12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and transferred onto PVDF membranes (Millipore Corp.,
Billerica, MA, USA). For western blotting, the PVDF membranes were
blocked in 5% non-fat milk at room temperature for 1 h and then
incubated with primary antibodies (anti-LKB1 antibody, 1:800,
27D10, Cell Signaling Technology; anti-Beclin1 antibody, 1:800,
EPR1733Y, Abcam and anti-β-actin antibody, 1:1000, sc-130301, Santa
Cruz Biotechnology Inc., Santa Cruz, CA, USA) at 4°C overnight. On
the following day, the membranes were washed with PBS-Tween-20
(PBS-T) three times and then incubated with the secondary antibody
(Santa Cruz Biotechnology) and the immunoreaction was detected
using the ECL reagent (Millipore Corp.). Each experiment was
repeated twice, and the band intensities were quantified by
densitometry.
Statistical analysis
Pearson’s χ2 test was used to determine
an association between expression of LKB1 or Beclin1 and the
clinicopathological parameters. Independent t-test was adopted to
assess the statistical significance of differences between cancer
and the matched surrounding normal tissues. Spearman’s test was
used to assess the correlation between LKB1 and Beclin1 expression.
Survival curves were calculated using the Kaplan-Meier method and
statistically analyzed using the log-rank test. For univariate and
multivariate analyses, Cox regression model was carried out to
estimate the prognostic value of the clinicopathological factors
and expression of LKB1 and Beclin1. Statistical analyses were
carried out using SPSS 17.0 software (SPSS, Chicago, IL, USA).
P<0.05 was defined as statistically significant.
Results
Expression of LKB1 in NSCLC and paired
surrounding normal tissues
We performed qRT-PCR, western blotting and
immunohistochemistry to assess the expression of LKB1 and Beclin1
in 142 paired normal and NSCLC tissue specimens. The data showed
that the level of LKB1 mRNA was significantly lower in the
NSCLC tissues than that in the normal lung tissues (P<0.001).
Reduced levels of LKB1 mRNA were associated with tumor
histology (P=0.041) and de-differentiation (P=0.006) (Table II). Immunohistochemical staining
revealed that LKB1 protein was moderately or strongly expressed in
the cytoplasm (Fig. 1). One hundred
and nine (76.8%) cases of NSCLC tissues expressed LKB1 protein but
the expression level was significantly lower than that in the
surrounding normal lung tissues (P<0.001). Reduced expression of
LKBI protein was associated with tumor histologic type (P<0.001)
and de-differentiation (P<0.001), but not with lymph node
metastasis and advanced TNM stage or with patient gender and age
(Table I). Similarly, western
blotting was consistent with the immunohistochemical data, showing
that LKB1 was expressed in 71.8% (102/142) of NSCLC tissues and in
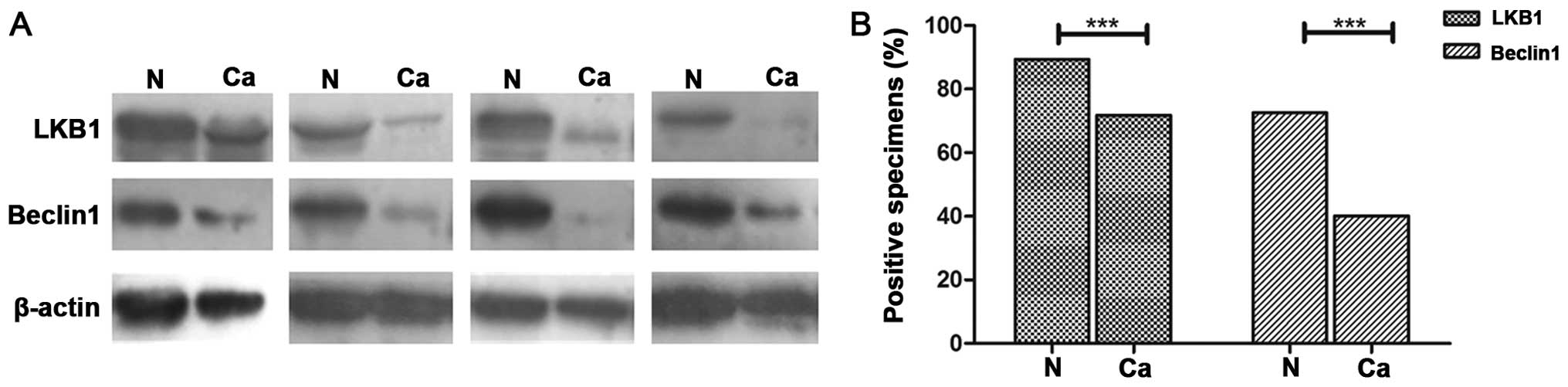
89.4% (127/142) of surrounding normal lung tissues (Fig. 2). LKB1 expression was significantly
lower in the NSCLC than that in paired normal lung tissues.
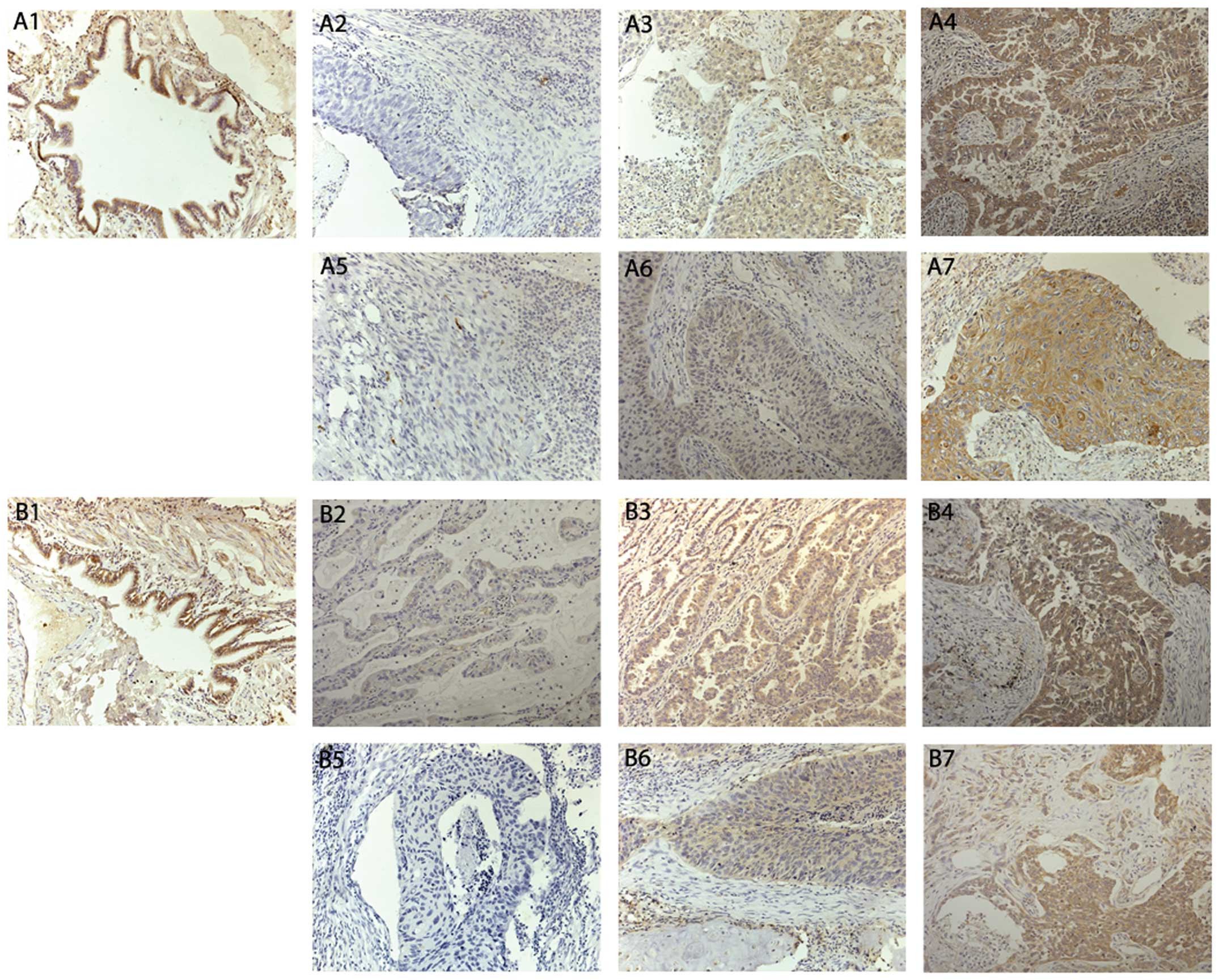 | Figure 1Immunohistochemical staining of LKB1
and Beclin1 proteins in NSCLC tissues (magnification, ×200).
Immunohistochemical expression of LKB1 in paired normal lung
tissues (A1), lung adenocarcinoma (A2–A4) and lung squamous cell
carcinoma (A5–A7). Immunohistochemical expression of Beclin1 in
paired normal lung tissues (B1), lung adenocarcinoma (B2–B4) and
lung squamous cell carcinoma (B5–B7). (A2, A5, B2, B5) weak
expression, (A3, A6, B3, B6) moderate expression and (A4, A7, B4,
B7) strong expression. LKB1, liver kinase B1; NSCLC, non-small cell
lung cancer. |
| Table IIAssociation of LKB1 and Beclin1 mRNA
levels with the clinicopathological parameters of the NSCLC
patients. |
Table II
Association of LKB1 and Beclin1 mRNA
levels with the clinicopathological parameters of the NSCLC
patients.
| | LKB1 mRNA
expression | | Beclin1 mRNA
expression | |
|---|
| |
| |
| |
|---|
|
Characteristics | n | Low | High | P-value | Low | High | P-value |
|---|
| All cases | 142 | | | | | | |
| Gender | | | | 0.419 | | | 0.927 |
| Male | 82 | 37 | 45 | | 43 | 39 | |
| Female | 60 | 23 | 37 | | 31 | 29 | |
| Age (years) | | | | 0.601 | | | 0.704 |
| <60 | 77 | 31 | 46 | | 39 | 38 | |
| ≥60 | 65 | 29 | 36 | | 35 | 30 | |
| Histologic
type | | | | 0.041 | | | 0.005 |
| ADC | 83 | 41 | 42 | | 35 | 48 | |
| SQCC | 59 | 19 | 40 | | 39 | 20 | |
|
Differentiation | | | | 0.006 | | | 0.031 |
| Poor | 57 | 32 | 25 | | 36 | 21 | |
| Moderate to
well | 85 | 28 | 57 | | 38 | 47 | |
| Lymph node
metastasis | | | | 0.193 | | | 0.112 |
| No | 80 | 30 | 50 | | 37 | 43 | |
| Yes | 62 | 30 | 32 | | 37 | 25 | |
| TNM stage | | | | 0.097 | | | 0.282 |
| I–II | 107 | 41 | 66 | | 53 | 54 | |
| III–IV | 35 | 19 | 16 | | 21 | 14 | |
Expression of Beclin1 in NSCLC and paired
surrounding normal tissues
Beclin1 protein was predominantly expressed in the
cytoplasm of both normal and tumor lung tissues. In NSCLC tissues,
Beclin1 protein expression was observed in 87 (61.3%) cases, which
was lower than that in the normal lung tissues (Fig. 1). Reduced expression of Beclin1 was
significantly associated with tumor histologic type (P=0.032) and
de-differentiation (P=0.015). However, there was no significant
association with patient gender and age, tumor lymph node
metastasis or TNM stage (Table I).
Similar results were observed in data from the western blot
analysis. Beclin1 protein was expressed in 40.1% (57/142) of NSCLC
tissues, which was significantly lower than that of the paired
surrounding normal lung tissues (72.5%, 103/142). Furthermore,
levels of Beclin1 mRNA were also significantly lower in the
NSCLC tissues than that in the paired normal lung tissues. The
reduced Beclin1 mRNA level was also associated with tumor
histology (P=0.005) and de-differentiation (P=0.031).
Correlation of LKB1 and Beclin1
expression in NSCLC tissue specimens
Since LKB1 and Beclin1 are being targeted for
clinical control of different human cancers, we examined the
possible correlation between these two proteins in terms of their
expression in NSCLC tissue samples using Spearman’s rank test. We
found that LKB1 protein expression was statistically correlated
(r=0.247, P=0.003) with Beclin1 expression (Table III). However, there was no
correlation between levels of LKB1 and Beclin1 mRNA
(r=0.135, P=0.109).
| Table IIICorrelation of LKB1 with Beclin1
expression in the NSCLC tissues. |
Table III
Correlation of LKB1 with Beclin1
expression in the NSCLC tissues.
| Beclin1
expression | |
|---|
|
| |
|---|
| Low | High | P-value |
|---|
| LKB1
expression |
| Low | 20 | 13 | |
| High | 35 | 74 | 0.003 |
Association of LKB1 and Beclin1 protein
expression with overall survival of NSCLC patients
All 142 patients were followed up until February
2014. We found that the median survival time of these patients was
31 months (range 3–71 months). The Kaplan-Meier curves were then
plotted after stratification based on LKB1 and Beclin1 protein
expression and analyzed using the log-rank test. The overall
survival of NSCLC patients with low LKB1 or Beclin1 expression was
significantly shorter than that of patients with high LKB1 or
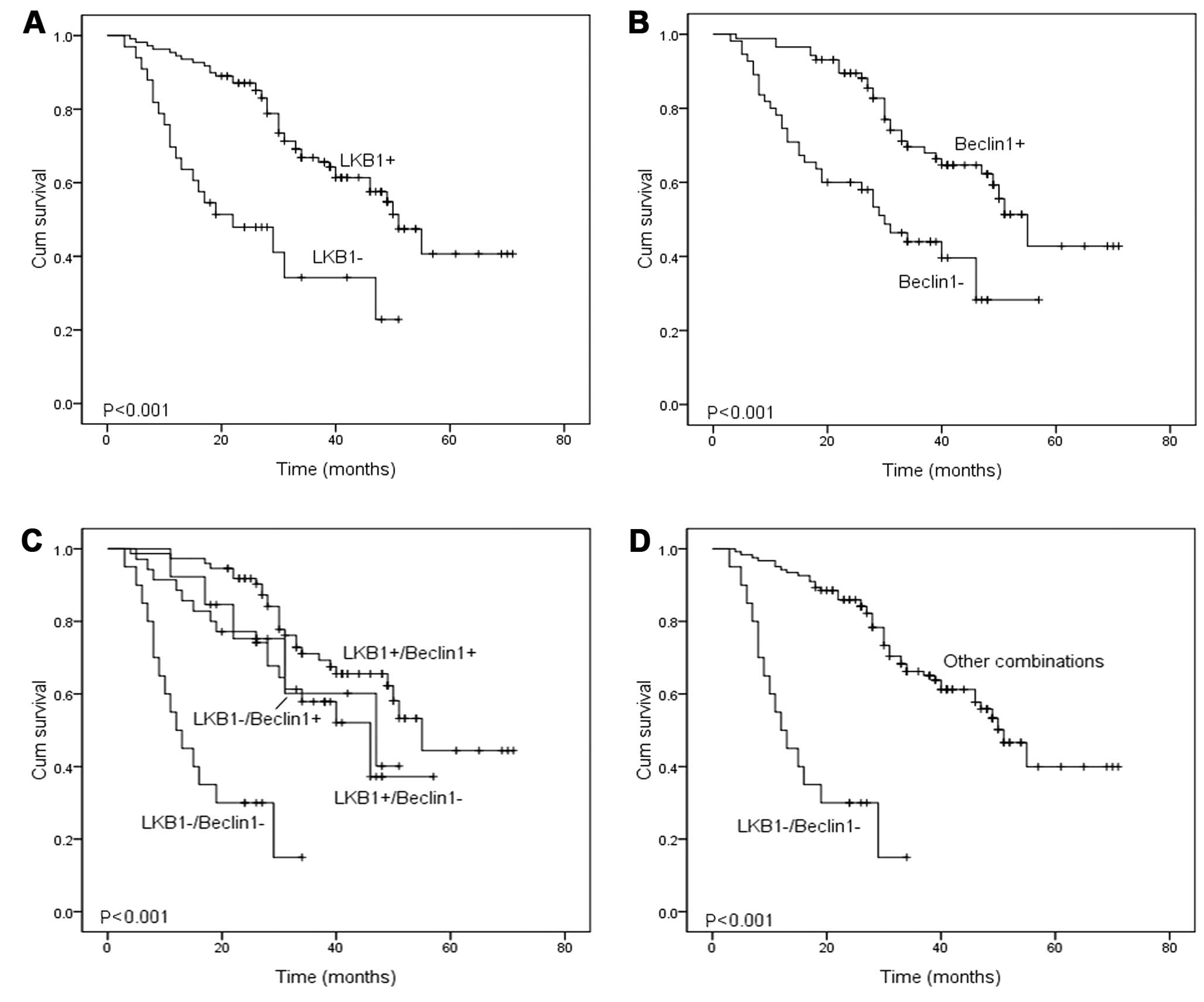
Beclin1 expression (Fig. 3). In
order to evaluate the prognostic significance of the combined LKB1
and Beclin1 expression, NSCLC patients were classified into four
subgroups: LKB1+/Beclin1+,
LKB1+/Beclin1-,
LKB1-/Beclin1- and
LKB1-/Beclin1+. We found that patients
without LKB1 and Beclin1 expression had a poorer overall survival
than the other three groups of patients (Fig. 3). Univariate and multivariate Cox
proportional hazards models (Table
IV) were established to assess prognostic indicators using
clinicopathological data and LKB1 and Beclin1 expression.
Specifically, univariate analysis showed that adenocarcinoma
(P=0.011), lymph node metastasis (P<0.001), advanced TNM stage
(P<0.001) and reduced expression of LKB1 [Exp (B), 0.31; 95% CI,
0.18–0.54; P<0.001] and Beclin1 [Exp (B), 0.38; 95% CI,
0.23–0.63; P<0.001] all predicted shorter overall survival of
NSCLC patients. Multivariate analysis further showed that
adenocarcinoma, lymph node metastasis, advanced TNM stage and
reduced expression of LKB1 and Beclin1 are all independent
indicators for overall survival of the NSCLC patients.
| Table IVUnivariate and multivariate analyses
of the overall survival of 142 NSCLC patients. |
Table IV
Univariate and multivariate analyses
of the overall survival of 142 NSCLC patients.
| | Overall
survival/univariate | Overall
survival/multivariate |
|---|
| |
|
|
|---|
|
Characteristics | n | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Gender |
| Male | 82 | 1 | | | |
| Female | 60 | 1.45
(0.88–2.37) | 0.145 | | |
| Age (years) |
| <60 | 77 | 1 | | | |
| ≥60 | 65 | 1.13
(0.69–1.85) | 0.637 | | |
| Histologic
type |
| ADC | 83 | 1 | | 1 | |
| SQCC | 59 | 0.50
(0.29–0.85) | 0.011 | 0.37
(0.20–0.67) | 0.001 |
|
Differentiation |
| Poor | 57 | 1 | | | |
| Moderate to
well | 85 | 0.84
(0.51–1.36) | 0.490 | | |
| Lymph node
metastasis |
| No | 80 | 1 | | 1 | |
| Yes | 62 | 4.34
(2.54–7.42) | <0.001 | 2.21
(1.17–4.17) | 0.015 |
| TNM stage |
| I-II | 107 | 1 | | 1 | |
| III-IV | 35 | 5.97
(3.56–10.01) | <0.001 | 4.80
(2.51–9.17) | <0.001 |
| LKB1 |
| Low | 33 | 1 | | 1 | |
| High | 109 | 0.31
(0.18–0.54) | <0.001 | 0.47
(0.25–0.88) | 0.018 |
| Beclin1 |
| Low | 55 | 1 | | 1 | |
| High | 87 | 0.38
(0.23–0.63) | <0.001 | 0.26
(0.15–0.48) | <0.001 |
Discussion
In the present study, we detected expression of LKB1
and Beclin1 mRNA and protein in 142 NSCLC and paired surrounding
normal lung tissues. We found that expression of both LKB1 and
Beclin1 mRNA and protein was reduced significantly in the NSCLC
tissues compared to levels in the normal tissues. Expression of
LKB1 protein was correlated with Beclin1 expression in the NSCLC
tissues and the reduced expression of LKB1 and
Beclin1 mRNA and proteins was associated with tumor
histology and de-differentiation. Furthermore, the reduced
expression of both LKB1 and Beclin1 proteins was associated with
poor overall survival of the NSCLC patients. Patients without LKB1
and Beclin1 expression had a poorer overall survival than the other
three groups of patients. Multivariate analysis confirmed that
expression of both LKB1 and Beclin1 proteins, together with
adenocarcinoma, lymph node metastasis and advanced TNM stage, are
independent indicators for the overall survival of NSCLC patients.
Thus, future studies are required to further evaluate these
molecules as biomarkers for NSCLC prognosis.
Our current data are consistent with previous
studies. LKB1 inactivation is common in lung adenocarcinoma
(27,28) and plays a role as a tumor biomarker
in mouse and human tissues (29).
Furthermore, inactivation of LKB1 via somatic mutations
commonly occurs in NSCLC, and loss of function mutations of
LKB1 have been reported in ~20–30% of NSCLC patients
(30). A mutation analysis also
identified LKB1 mutations in other cancers, such as
melanoma, cervical, pancreatic and prostate cancers (31–33).
Ji et al (11) provided
strong evidence that LKB1 plays a key role in lung cancer
differentiation and metastasis. In addition, mouse models of LKB1
heterozygous deletion showed that the LKB1 deletion is
sufficient to induce cancer development in mice (34–36).
However, Onozato et al (37)
showed that there was no statistically significant correlation
between patient survival curves and expression of LKB1
mRNA.
At the cellular level, LKB1 can induce or prevent
apoptosis, depending on the cell context. This is due to the fact
that LKB1 is also involved in autophagy, considered as type II
programmed cell death. Autophagy is a self-digesting process that
degrades and recycles aged protein and organelles engulfed into
autophagosomes through the lysosomal pathway (38). Autophagy is a highly conserved
degradation process regulated by autophagy-related genes. Several
diseases, such as neurodegenerative, cancer, ageing and certain
liver and heart diseases are related to dysfunctional autophagy
(39). The current theory suggests
that autophagy can act as a tumor suppressor or cell apoptosis
protector. A previous study showed that LKB1 activated autophagy
through AMPK activation to inhibit mTOR and protected from cell
death (6). The LKB1-AMPK pathway
can be activated by energy stress and ROS. Moreover, Beclin1
directly participates in autophagy regulation, and altered Beclin1
expression is associated with human tumorigenesis. Indeed, the
present data showed that expression of Beclin1 mRNA and protein was
significantly reduced in NSCLC tissues compared to that in the
paired surrounding normal lung tissues, which is consistent with
previous studies of NSCLC (40,41).
However, Ahn et al (22)
showed that Beclin1 expression in gastric and colorectal cancer was
notably higher than that in paired normal tissues. The reason for
this discrepancy is not known, but it may be tissue-specific. In
addition, our present data showed that Beclin1 expression was
associated with tumor histology and differentiation and with
overall survival of NSCLC patients. We also found that low LKB1
expression was significantly correlated with low Beclin1
expression. We therefore propose that LKB1 may influence tumor
differentiation and metastasis through autophagy. However, the
exact molecular mechanisms require further investigation.
In conclusion, the present study demonstrated that
expression of LKB1 and Beclin1 was reduced in NSCLC tissues, and
the reduced expression of these proteins, together with
adenocarcinoma, lymph node metastasis and advanced TNM stage, are
independent indicators for the overall survival of NSCLC patients.
Further studies to verify our data are required before translation
into clinical practice.
Acknowledgements
This study was supported in part by a grant from the
National Natural Science Foundation of China (nos. 81101777 and
8130190). The authors would like to thank Professsor Chen Huang of
Xi’an Jiaotong University for the technical support.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar
|
|
2
|
Hemminki A, Markie D, Tomlinson I, et al:
A serine/threonine kinase gene defective in Peutz-Jeghers syndrome.
Nature. 391:184–187. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jenne DE, Reomann H, Nezu J, et al:
Peutz-Jeghers syndrome is caused by mutations in a novel serine
threonine kinase. Nat Genet. 18:38–43. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Smith DP, Spicer J, Smith A, Swift S and
Ashworth A: The mouse Peutz-Jeghers syndrome gene Lkbl
encodes a nuclear protein kinase. Hum Mol Genet. 8:1479–1485. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Boudeau J, Kieloch A, Alessi DR, Stella A,
Guanti G and Resta N: Functional analysis of LKB1/STK11 mutants and
two aberrant isoforms found in Peutz-Jeghers syndrome patients. Hum
Mutat. 21:1722003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shackelford DB and Shaw RJ: The LKB1-AMPK
pathway: metabolism and growth control in tumour suppression. Nat
Rev Cancer. 9:563–575. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kahn BB, Alquier T, Carling D and Hardie
DG: AMP-activated protein kinase: ancient energy gauge provides
clues to modern understanding of metabolism. Cell Metab. 1:15–25.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baas AF, Boudeau J, Sapkota GP, et al:
Activation of the tumour suppressor kinase LKB1 by the STE20-like
pseudokinase STRAD. EMBO J. 22:3062–3072. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Nakada D, Saunders TL and Morrison SJ:
Lkb1 regulates cell cycle and energy metabolism in haematopoietic
stem cells. Nature. 468:653–658. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jansen M, Ten Klooster JP, Offerhaus GJ
and Clevers H: LKB1 and AMPK family signaling: the intimate link
between cell polarity and energy metabolism. Physiol Rev.
89:777–798. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ji H, Ramsey MR, Hayes DN, et al: LKB1
modulates lung cancer differentiation and metastasis. Nature.
448:807–810. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kroemer G, Mariño G and Levine B:
Autophagy and the integrated stress response. Mol Cell. 40:280–293.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
White E and DiPaola RS: The double-edged
sword of autophagy modulation in cancer. Clin Cancer Res.
15:5308–5316. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Carew JS, Nawrocki ST, Kahue CN, et al:
Targeting autophagy augments the anticancer activity of the histone
deacetylase inhibitor SAHA to overcome Bcr-Abl-mediated drug
resistance. Blood. 110:313–322. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Michaud M, Martins I, Sukkurwala AQ, et
al: Autophagy-dependent anticancer immune responses induced by
chemotherapeutic agents in mice. Science. 334:1573–1577. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Moretti L, Yang ES, Kim KW and Lu B:
Autophagy signaling in cancer and its potential as novel target to
improve anticancer therapy. Drug Resist Updat. 10:135–143. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu YL, Yang PM, Shun CT, Wu MS, Weng JR
and Chen CC: Autophagy potentiates the anti-cancer effects of the
histone deacetylase inhibitors in hepatocellular carcinoma.
Autophagy. 6:1057–1065. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi AM, Ryter SW and Levine B: Autophagy
in human health and disease. N Engl J Med. 368:651–662. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wrighton KH: Autophagy: kinase crosstalk
through beclin 1. Nat Rev Mol Cell Biol. 14:402–403. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Won KY, Kim GY, Kim YW, Song JY and Lim
SJ: Clinicopathologic correlation of beclin-1 and bcl-2 expression
in human breast cancer. Hum Pathol. 41:107–112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Y, Lu Y, Lu C and Zhang L: Beclin-1
expression is a predictor of clinical outcome in patients with
esophageal squamous cell carcinoma and correlated to
hypoxia-inducible factor (HIF)-1α expression. Pathol Oncol Res.
15:487–493. 2009.PubMed/NCBI
|
|
22
|
Ahn CH, Jeong EG, Lee JW, et al:
Expression of beclin-1, an autophagy-related protein, in gastric
and colorectal cancers. APMIS. 115:1344–1349. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen N and Karantza V: Autophagy as a
therapeutic target in cancer. Cancer Biol Ther. 11:157–168. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen S, Rehman SK, Zhang W, Wen A, Yao L
and Zhang J: Autophagy is a therapeutic target in anticancer drug
resistance. Biochim Biophys Acta. 1806:220–229. 2010.PubMed/NCBI
|
|
25
|
Alexander A and Walker CL: The role of
LKB1 and AMPK in cellular responses to stress and damage. FEBS
Lett. 585:952–957. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Edge SB and Compton CC: The American Joint
Committee on Cancer: the 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shackelford DB, Abt E, Gerken L, et al:
LKB1 inactivation dictates therapeutic response of non-small cell
lung cancer to the metabolism drug phenformin. Cancer Cell.
23:143–158. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sanchez-Cespedes M, Parrella P, Esteller
M, et al: Inactivation of LKB1/STK11 is a common event in
adenocarcinomas of the lung. Cancer Res. 62:3659–3662.
2002.PubMed/NCBI
|
|
29
|
Nakada Y, Stewart TG, Peña CG, et al: The
LKB1 tumor suppressor as a biomarker in mouse and human tissues.
PLoS One. 8:e734492013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Matsumoto S, Iwakawa R, Takahashi K, et
al: Prevalence and specificity of LKB1 genetic alterations
in lung cancers. Oncogene. 26:5911–5918. 2007.
|
|
31
|
Wingo SN, Gallardo TD, Akbay EA, et al:
Somatic LKB1 mutations promote cervical cancer progression.
PLoS One. 4:e51372009.
|
|
32
|
Su GH, Hruban RH, Bansal RK, et al:
Germline and somatic mutations of the STK11/LKB1
Peutz-Jeghers gene in pancreatic and biliary cancers. Am J Pathol.
154:1835–1840. 1999.PubMed/NCBI
|
|
33
|
Rowan A, Bataille V, MacKie R, et al:
Somatic mutations in the Peutz-Jegners (LKB1/STKII) gene in
sporadic malignant melanomas. J Invest Dermatol. 112:509–511. 1999.
View Article : Google Scholar
|
|
34
|
Nakau M, Miyoshi H, Seldin MF, Imamura M,
Oshima M and Taketo MM: Hepatocellular carcinoma caused by loss of
heterozygosity in Lkb1 gene knockout mice. Cancer Res.
62:4549–4553. 2002.PubMed/NCBI
|
|
35
|
Miyoshi H, Nakau M, Ishikawa TO, Seldin
MF, Oshima M and Taketo MM: Gastrointestinal hamartomatous
polyposis in Lkb1 heterozygous knockout mice. Cancer Res.
62:2261–2266. 2002.PubMed/NCBI
|
|
36
|
McCarthy A, Lord CJ, Savage K, et al:
Conditional deletion of the Lkb1 gene in the mouse mammary
gland induces tumour formation. J Pathol. 219:306–316. 2009.
|
|
37
|
Onozato R, Kosaka T, Achiwa H, et al:
LKB1 gene mutations in Japanese lung cancer patients. Cancer
Sci. 98:1747–1751. 2007. View Article : Google Scholar
|
|
38
|
Klionsky DJ and Emr SD: Autophagy as a
regulated pathway of cellular degradation. Science. 290:1717–1721.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu J, Lin Y, Yang H, Deng Q, Chen G and
He J: The expression of p33(ING1), p53, and autophagy-related gene
Beclin1 in patients with non-small cell lung cancer. Tumour Biol.
32:1113–1121. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kim KM, Yu TK, Chu HH, et al: Expression
of ER stress and autophagy-related molecules in human non-small
cell lung cancer and premalignant lesions. Int J Cancer.
131:E362–E370. 2012. View Article : Google Scholar : PubMed/NCBI
|