Introduction
Prostate cancer (PCa) is the most common malignancy
and the second leading cause of cancer-related death in men in
Western countries. Advanced and metastatic stages of the disease
are found in 35% of patients with PCa diagnosed at autopsy
(1). Among patients with localized
cancer who are eligible for radical prostatectomy, ~35% will
develop recurrence (metastatic disease) within 10 years of surgery
(2,3).
Androgen deprivation therapy (ADT) can be effective
in patients who present with or progress to advanced or meta-static
disease. Unfortunately, the median duration of response to ADT is
limited to between 8 months and 3 years (4), and these patients will eventually
become castration resistant. Chemotherapy is an effective treatment
for castration-resistant PCa, but the median duration of response
is only 10.3 months (5). There is
clearly an urgent need to develop additional systemic interventions
for patients with progressive PCa. Angiogenesis plays a crucial
role in PCa progression and metastasis. Microvessel density (MVD)
has been found to be more prominent in PCa than in benign prostatic
hyperplasia (BPH) and normal tissue (6,7). It
has been reported that MVD increases with increased Gleason’s
score, particularly in poorly differentiated PCa (8). MVD was also significantly correlated
with cancer-specific survival in 221 patients with PCa followed up
for a median of 15 years (9).
Vascular endothelial growth factor (VEGF) is the
most prominent regulator of physiological angiogenesis and has been
correlated with increased levels of angiogenesis in clinical
studies comparing PCa with BPH (7).
Higher VEGF expression and serum levels have also been found in
patients with metastasis or poorly differentiated tumors, as well
as in those with a poor prognosis (10–13).
However, it has become increasingly apparent that current
anti-angiogenic therapy targeting VEGF has only a modest effect in
the clinical setting.
RhoA and its downstream effector, Rho-associated
protein kinase (ROCK), serve as key regulators of extracellular
stimulus-mediated signaling networks that are involved in various
cellular processes, including motility, mitosis, proliferation and
apoptosis (14). Suppression of the
RhoA/ROCK signaling pathway with the ROCK inhibitor, Y-27632, was
found to inhibit VEGF-induced angiogenesis in vitro
(15). Another ROCK inhibitor,
fasudil, has been shown to inhibit VEGF-induced angiogenesis in
vitro and in vivo (16).
A study carried out on endothelial cells from transgenic
adenocarcinoma of the mouse prostate (TRAMP) mice revealed that
their behavior correlated with a constitutively high level of
baseline activity of Rho GTPase and ROCK (17). This suggests that the RhoA/ROCK
pathway has an important role in PCa angiogenesis. However, the
anti-angiogenic effects of ROCK inhibitors in PCa are unclear. We
investigated the role of fadusil, a ROCK inhibitor, that has been
approved for clinical use for pulmonary arterial hypertension, on
PCa-induced angiogenesis in vitro.
Materials and methods
Cell culture
Human umbilical vein endothelial cells (HUVECs) were
purchased from PromoCell (C-12200; Heidelberg, Germany) and
cultured in endothelial cell growth medium (C-22010; PromoCell).
Cultures were maintained at 37°C in a humidified atmosphere
containing 5% CO2. Subcultures were obtained by
trypsinization and were used for experiments at passages 3 to 9.
Before performing the experiments, the cells were made quiescent by
incubating overnight in endothelial cell basal medium (C-22210)
containing 0.5% (w/v) fetal bovine serum (FBS). The PCa cell line,
PC-3, was purchased from The European Collection of Cell Cultures
and grown in F-12K (Gibco-Invitrogen, Carlsbad, CA, USA) containing
10% (w/v) FBS. PC-3 cells were seeded at a concentration of
6×106 cells/T75 flask. On the following day, the medium
was replaced with basal medium without FBS, and the supernatants
were harvested after a 24-h incubation to serve as conditioned
medium (PC3CM). Recombinant human VEGF 165 was purchased from
R&D Systems (293-VE; Minneapolis, MN, USA). HUVECs were
cultured in endothelial cell basal medium plus 2% (w/v) FBS
(control group), in PC3CM plus 2% (w/v) FBS (PC3CM group), or in
basal medium plus 2% (w/v) FBS and 30 ng/ml VEGF (VEGF group).
Cell proliferation assay
HUVEC proliferation was evaluated using a BrdU
incorporation assay kit (Amersham; Cell Proliferation Biotrak ELISA
System; RPN250; GE Healthcare, Little Chalfont, UK), according to
the manufacturers’ instructions. In brief, HUVECs were plated in
96-well microculture plates (3×103 cells/well). After a
48-h incubation at 37°C in a 5% CO2 atmosphere, with or
without fasudil (1–100 μM), 10 μl BrdU labeling reagent was added,
and the cells were cultured for a further 2 h. Cells were washed
twice with Dulbecco’s PBS (D8537; Sigma-Aldrich, St. Louis, MO,
USA), fixed with fixative solution and then blocked with blocking
buffer. BrdU incorporation was revealed by incubation with 100
μl/well horseradish peroxidase (HRP)-labeled anti-BrdU working
solution for ~ 90 min. Tetramethylbenzidine (TMB) substrate at room
temperature was added at 100 μl/well for 20 min. Absorbance was
measured at 450 nm using a microplate reader. All determinations
were performed in octuplicate, and each experiment was repeated
three times.
Cell migration assay
Cell motility was assessed using a wound-healing
migration assay. HUVECs were seeded to full confluency in 6-well
plates. The following day, a uniform scratch was made down the
centre of the well using a 100-μl micropipette tip, and the cells
were washed twice with PBS. After incubation for 24 h with or
without 30 μM fasudil in the control, PC3CM and VEGF groups, the
cells were fixed and photographed. Photographic imaging was
performed using a Leica inverted microscope. Cell migration was
quantified by measuring the ratio of the migration area to the
total area of the wound gap. Each experiment was repeated three
times.
Tube formation assay
Ninety-six-well plates were chilled to 4°C and
coated with 50 μl of Matrigel (354234; BD Biosciences, Oxford, UK)
per well. Freshly passaged HUVECs were seeded onto the gel.
Endothelial tube morphogenesis was carried out in the presence or
absence of fasudil (3–30 μM). Endothelial tube formation was
observed after 16 h and photographed under phase contrast
microscopy using a Leica inverted microscope. Quantification of the
digital images was performed by counting the total number of tubes
in five 40× fields, and total tube length was quantified using
ImageJ™ software (NIH, Bethesda, MD, USA). Tube formation was
expressed as fold change or percentage, compared to the controls.
All determinations were performed three times, and each experiment
was repeated three times.
Spheroid sprouting assay
HUVECs were suspended in culture medium containing
0.2% (w/v) methylcellulose (Sigma-Aldrich) and seeded in
non-adherent round-bottom 96-well plates (Greiner, Frickenhausen,
Germany). All suspended cells formed a single spheroid in each well
of defined size and cell number (~400 cells/spheroid). Spheroids
were left to form for 24 h and then embedded in 1.5 mg/ml collagen
gel. The spheroid-containing gel was rapidly transferred to
pre-warmed 24-well plates and allowed to polymerize for 30 min.
Endothelial basal medium or PC3CM with or without fasudil (1–100
μM) was then added to the surface of the gel (500 μl/well). After
16 h, images were captured using a Leica inverted microscope.
Sprouting was quantified using NIH ImageJ software by measuring the
cumulative sprout length, which consisted of every sprout from 10
spheroids in each group.
Western blot assay
Protein was extracted on ice from the cultured
HUVECs with cold RIPA lysis buffer (9806; Cell Signaling
Technology, Boston, MA, USA) containing Pierce™ Protease and
Phosphatase Inhibitor (88669; Thermo Scientific, Rockford, IL,
USA). Lysates were centrifuged at 12,000 × g for 20 min at 4°C, and
the supernatant was collected. Total protein concentrations were
determined using a bicinchoninic acid assay (BCA) protein assay kit
(23250; Thermo Scientific). Equal amounts of protein were subjected
to sodium dodecyl sulfate polyacrylamide gel electrophoresis and
then electrically transferred onto nitrocellulose membranes. The
membranes were blocked for 1 h with 5% (w/v) non-fat milk in
PBS-0.1% (v/v) Tween-20 (PBST) and incubated with primary
antibodies against MYPT-1 (1:1,000; sc-25618; Santa Cruz
Biotechnology, Dallas, TX, USA), phospho-MYPT-1 (1:500; ABS45;
Millipore, Billerica, MA, USA), anti-ROCK1 (1:500; sc-6055),
anti-ROCK2 (1:1,000; sc-1851; both from Santa Cruz Biotechnology)
and β-actin (1:500; ab8229; Abcam, Cambridge, UK) overnight at 4°C.
Finally, the membrane was incubated with HRP-conjugated secondary
antibodies as follows: goat anti-mouse IgG-HRP (1:5,000; sc-2005;
Santa Cruz Biotechnology), rabbit anti-goat IgG-HRP (1:10,000;
sc-2768; Santa Cruz Biotechnology), goat anti-rabbit IgG-HRP
(1:10,000; sc-2004; Santa Cruz Biotechnology) for 1 h at room
temperature. After washing three times with PBST, proteins were
visualized using an ECL Prime Western blotting detection kit (GE
Healthcare). Photographs of the protein bands were captured using a
digital imaging system (ImageQuant LAS; GE Healthcare), and
densitometric measurements of band intensity in the western
blotting were performed using NIH ImageJ software. The results
shown are representative of three or more independent
experiments.
Statistical analysis
Data are expressed as means ± standard deviation.
Significance of differences was determined by the two-tailed
Student’s t-test or the analysis of variance least significant
difference (ANOVA LSD) test. A P-value <0.05 was considered to
indicate a statistically significant difference.
Results
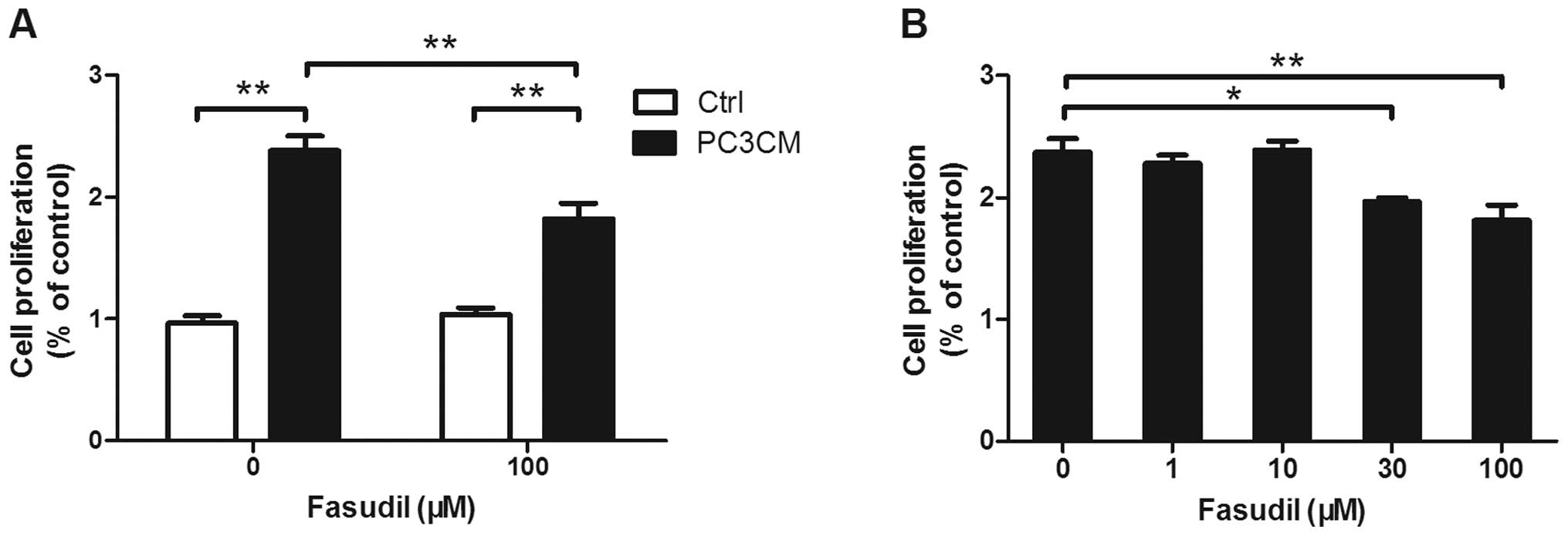
Fasudil inhibits PC3CM-induced HUVEC
proliferation
Endothelial cell proliferation is crucial for
angiogenesis. PC3CM-treated HUVECs were exposed to fasudil
concentrations ranging from 1 to 100 μM, and HUVEC proliferation
was examined using a BrdU assay. Fasudil concentrations of ≥ 30 μM
had a significant inhibitory effect on PC3CM-induced cell
proliferation, while proliferation in the control group was
unchanged (Fig. 1).
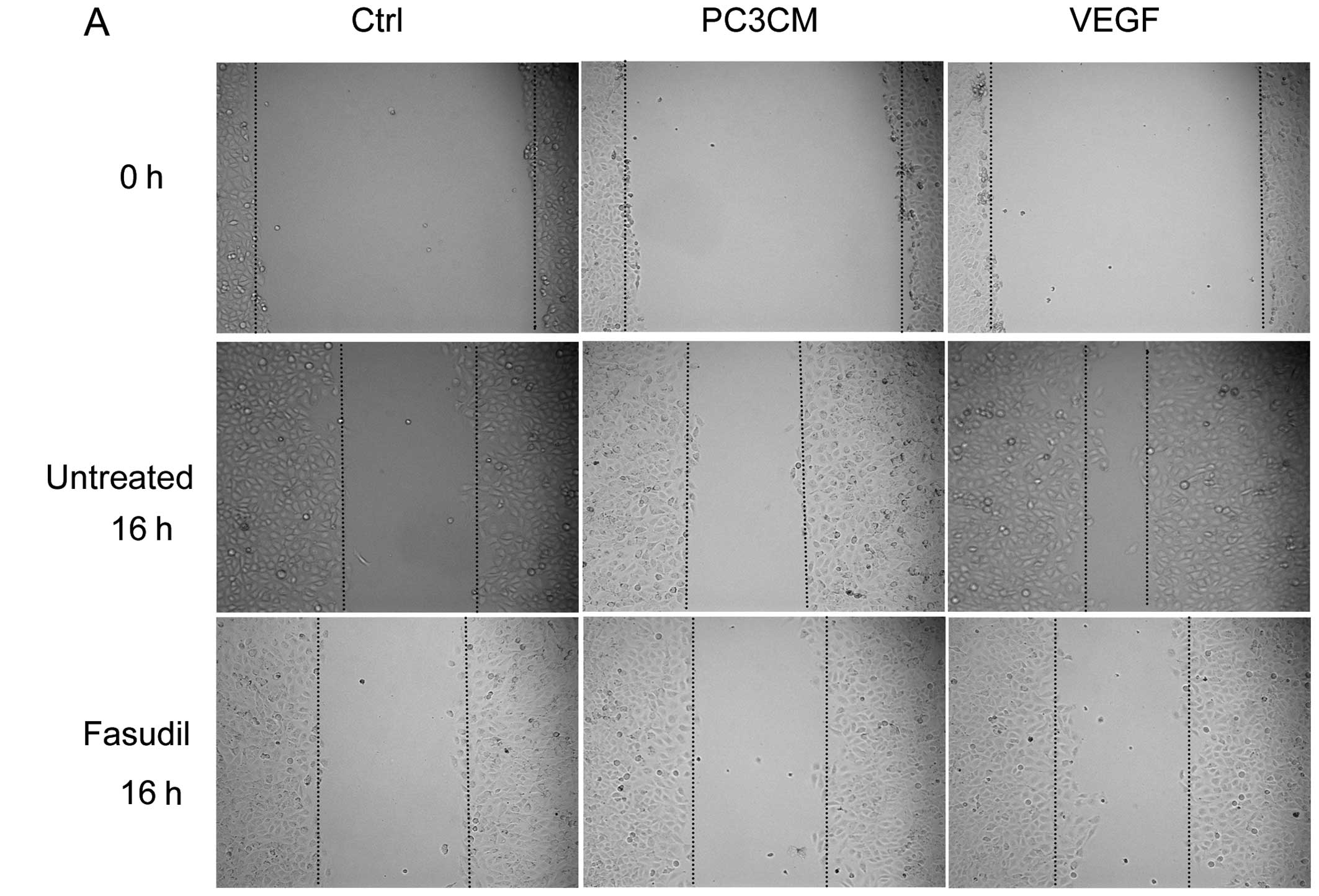
Fasudil inhibits PC3CM-induced HUVEC
migration
The inhibitory effects of fasudil on endothelial
cell motility were assessed using a wound-healing migration assay.
Fasudil (30 μM) significantly decreased the number of cells
migrating into the scratched gap in the control, PC3CM and VEGF
groups, indicating the potent inhibitory effect of fasudil on HUVEC
movement and migration. VEGF increased HUVEC migration
significantly more than PC3CM-induced HUVEC migration. After
treatment with 30 μM fasudil, all migrations decreased to similar
levels (Fig. 2).
Fasudil inhibits PC3CM-induced HUVEC tube
formation
The effect of fasudil on capillary-like structure
formation in vitro was examined using a 3-dimensional (3D)
Matrigel assay. When seeded onto Matrigel, HUVECs form tube
structures and connect with each other, mimicking the in
vivo process of angiogenesis. Sixteen hours after seeding,
untreated HUVECs exhibited a clear capillary-like network
formation. However, fasudil treatment dramatically decreased the
capillary-like network formation in a dose-dependent manner. As
fasudil concentration increased, total tube length gradually
decreased (Fig. 3).
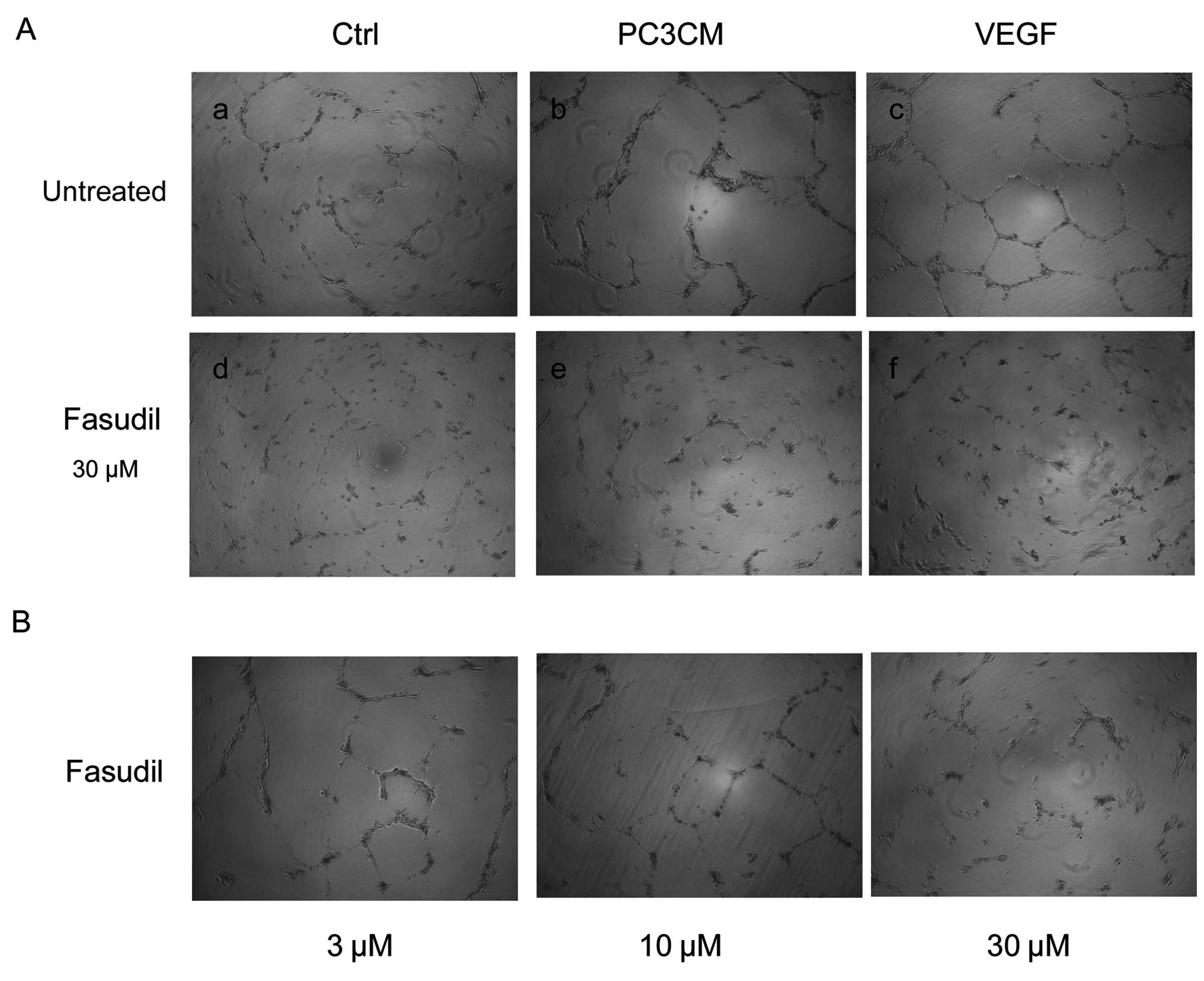 | Figure 3PC3CM-induced capillary-like tube
formation is reduced by fasudil. (A) HUVECs were cultured on
Matrigel with basal medium (a), PC3CM (b), or basal medium
containing 30 ng/ml VEGF (c) either in the absence (a–c), or
presence of 30 μM fasudil (d–f). Images were captured after 16 h
under a Leica inverted phase-contrast microscope. Representative
images are shown above. PC3CM (b) increased the total tube length
compared to the control (a), whereas fasudil strongly inhibited
tube formation (e). (B) Dose-dependent effects of fasudil on total
tube length in PC3CM-cultured HUVECs. As the fasudil concentration
increased, the total tube length gradually decreased. (C and D)
Total tube length was quantified by evaluating five fields in each
experiment, and data were verified by three independent
experiments. Values are expressed as the mean ± SEM. HUVECs, human
umbilical vein endothelial cells; PCa, prostate cancer; PC3CM, PCa
cell line PC3-conditioned media; SEM, standard error of the mean;
VEGF, vascular endothelial growth factor. |
Fasudil inhibits PC3CM-induced HUVEC
spheroid sprouting
In the sprout formation assay, HUVECs seeded in
non-adhesive conditions in round bottom 96-well plates contributed
to the formation of a single spheroid with a quiescent,
non-proliferating surface monolayer within 24 h. The spheroids were
then embedded in a 3D collagen matrix. In the untreated control
group, baseline sprouting was low (Fig.
4Aa). When cultured with PC3CM (Fig. 4Ab), baseline sprouting increased
dramatically, although it was still less than that in the cells
cultured with basal medium containing 30 ng/ml VEGF (Fig. 4Ac). Sprouting was almost completely
inhibited by treatment with 100 μM fasudil (Fig. 4Ad–f). We then examined the
dose-dependent response of fasudil on PCa-induced HUVEC sprouting.
As shown in Fig. 4B, fasudil
decreased HUVEC sprouting in a dose-dependent manner and 100 μM
fasudil again inhibited sprouting almost completely.
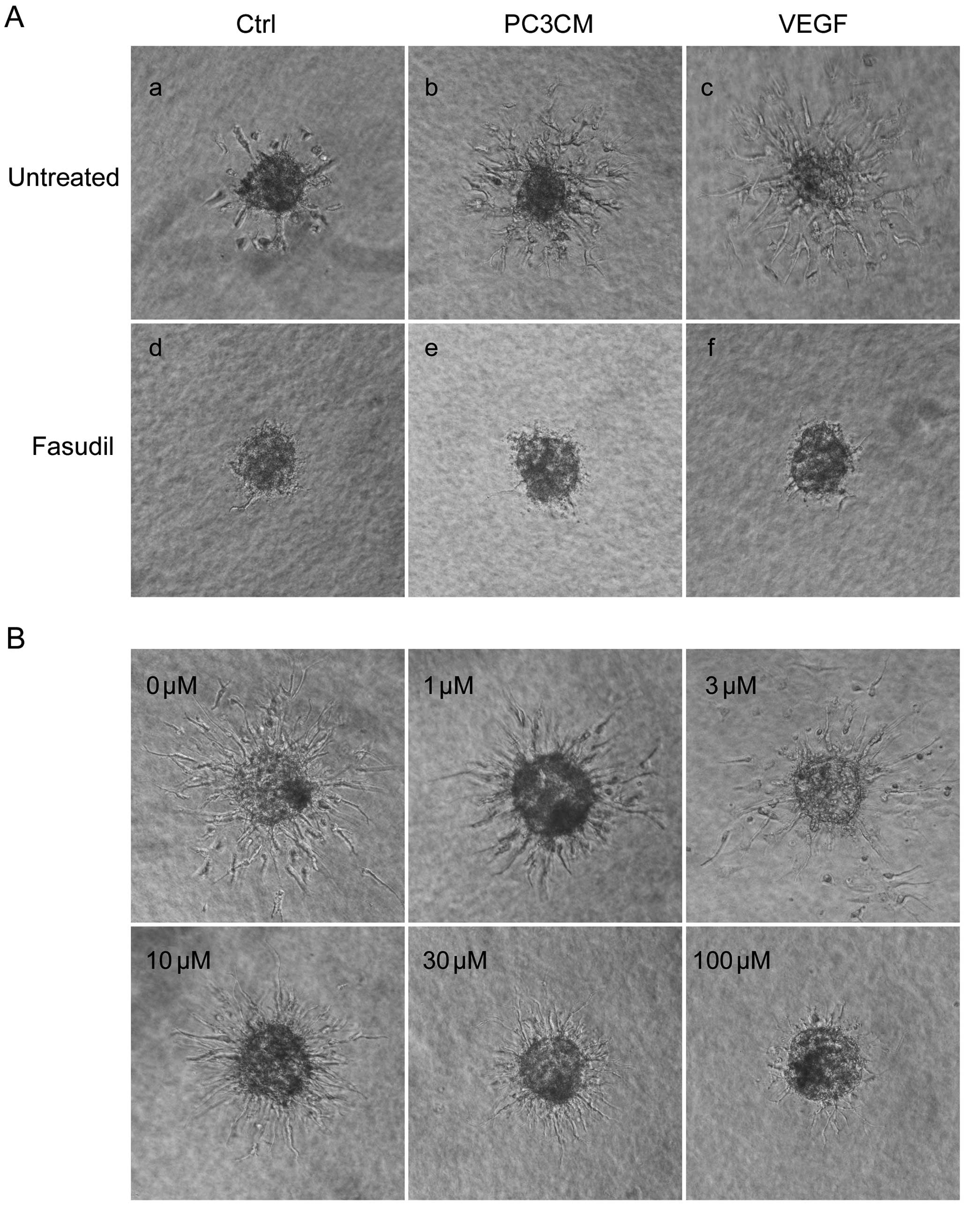 | Figure 4PC3CM-induced HUVEC spheroid sprouting
is decreased by fasudil. (A) The sprouting of capillary-like
structures from collagen-embedded HUVEC spheroids treated with
fasudil (100 μM) or left untreated was determined in basal medium
(Ctrl), PC3CM, or basal medium containing 30 ng/ml VEGF. After 16
h, sprouting was digitally recorded using a phase-contrast
microscope. Sprouting from HUVEC spheroids stimulated with PC3CM
(b) was almost completely inhibited by fasudil (e). (B) Fasudil
decreased HUVEC sprouting in a dose-dependent manner, and 100 μM
fasudil inhibited sprouting almost completely. With increasing
concentration of fasudil from 10–30 μM, the sprouts became thinner
and more abundant compared with the ordered architecture of the
untreated HUVEC spheroid sprouts. These sprouts were more like cell
protrusions. (C and D) Quantification of sprouts from 10 spheroids
(mean ± SEM) was assessed by evaluating the cumulative sprout
length per spheroid, and data were verified by three independent
experiments. *P<0.05; **P<0.01. HUVECs,
human umbilical vein endothelial cells; PCa, prostate cancer;
PC3CM, PCa cell line PC3-conditioned media; SEM, standard error of
the mean; VEGF, vascular endothelial growth factor. |
Furthermore, when treated with increasing
concentrations of fasudil, the sprouts became thinner and the HUVEC
nucleus seldom emerged from the spheroids. These sprouts resembled
cell protrusions, were markedly thinner compared with the untreated
HUVEC sprouts, and were more abundant compared with the ordered
architecture of the single HUVEC spheroid sprouts (Fig. 4B).
Fasudil inhibits PC3CM-induced HUVEC ROCK
activation
MYPT-1 is one of the most crucial downstream
effectors of ROCK. As fasudil is a ROCK inhibitor, we examined the
inhibitory effects of fasudil on ROCK by measuring phospho-MYPT-1
(pMYPT-1), the active form of MYPT-1.
As shown in Fig. 5,
when cultured with PC3CM or basal medium containing VEGF,
expression of both MYPT-1 and pMYPT-1 was increased in the HUVECs,
resulting in a moderate increase in the pMYPT-1/MYPT-1 ratio,
indicating ROCK activation. Fasudil treatment lead to a significant
decrease in pMYPT-1 and a slight decrease in MYPT-1, resulting in a
significant decrease in the pMYPT-1/MYPT-1 ratio (Fig. 5A–D). A moderate increase in ROCK1
and ROCK2 expression was also detected, but ROCK expression was not
altered significantly by fasudil treatment (Fig. 5E and F).
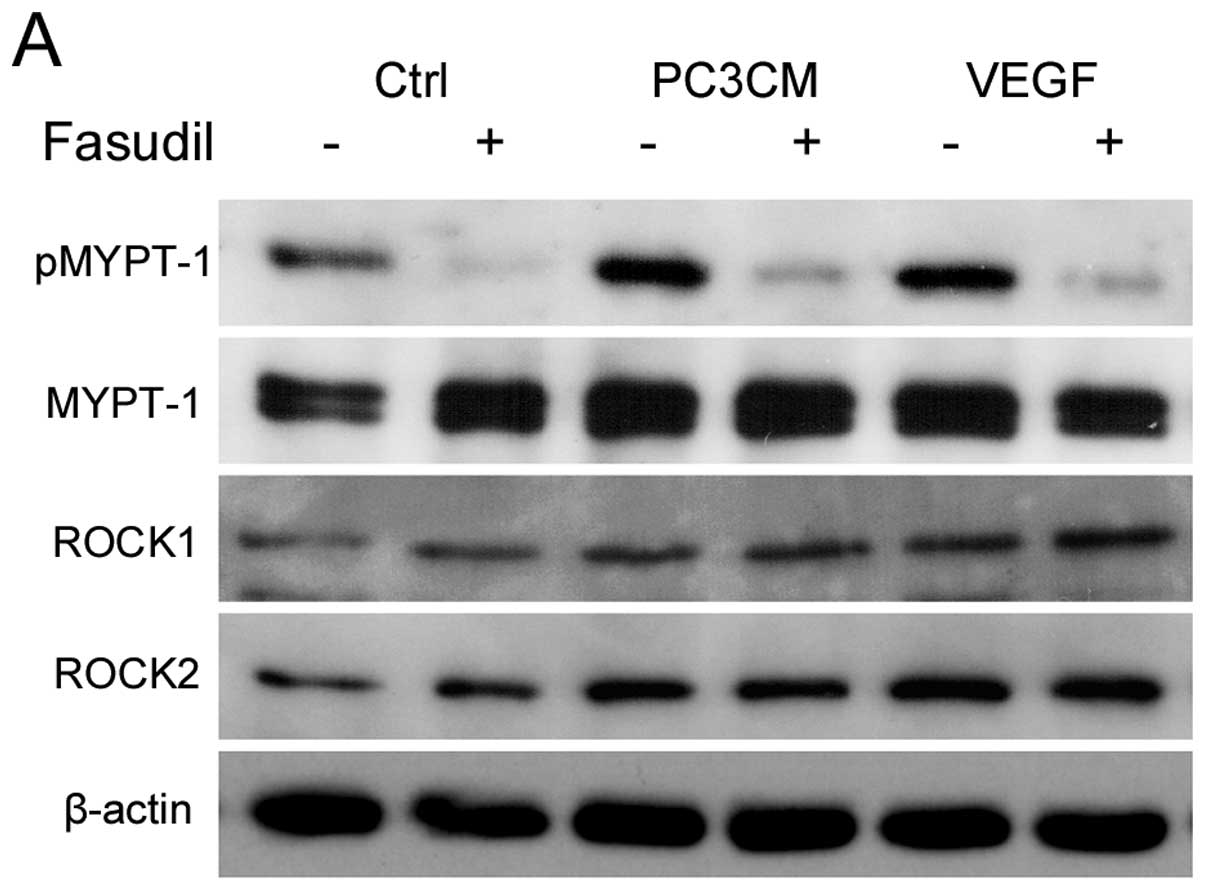 | Figure 5Fasudil inhibits PC3CM-induced ROCK
activity. HUVECs were cultured in either basal medium (Ctrl),
PC3CM, or basal medium containing VEGF (30 ng/ml) and treated with
30 μM fasudil or left untreated. ROCK expression was detected by
immunoblotting with anti-ROCK1 and anti-ROCK2 antibodies. ROCK
activity was detected by phosphorylation of the downstream
effector, MYPT-1, with anti-phospho-MYPT-1 and anti-MYPT-1
antibodies. (A) Representative images of western blots are shown.
The relative density of each blot was quantified as fold-expression
relative to the control. The data shown are the mean ± SEM of 6
independent experiments. PC3CM increased pMYPT-1 expression and
fasudil decreased pMYPT-1 expression (B) without any significant
changes in MYPT-1 (C), resulting in an increase in the
pMYPT-1/MYPT-1 ratio (D) in the PC3CM group and a decrease in this
ratio after fasudil treatment. (E and F) PC3CM increased ROCK1 and
ROCK2 expression while fasudil had no effect on ROCK expression.
HUVECs, human umbilical vein endothelial cell; MYPT, myosin
phosphatase target subunit 1; PCa, prostate cancer; PC3CM, PCa cell
line PC3-conditioned media; ROCK, Rho-associated protein kinase;
SEM, standard error of the mean; VEGF, vascular endothelial growth
factor. |
Discussion
To our knowledge, there have been no previous
reports on the effects of fasudil on PCa-induced angiogenesis. In
this study, HUVECs were cultured with the PCa cell line PC3CM to
mimic endothelial cells in PCa tissue. Fasudil was then added to
examine its effects on PC3CM-induced HUVECs using in vitro
angiogenesis assays.
When cultured with PC3CM, ROCK1 and ROCK2 expression
increased in the HUVECs, as did pMYPT-1 and total MYPT-1
expression. The pMYPT-1/MYPT-1 ratio was also increased. This
indicates activation of the RhoA/ROCK pathway in PC3CM-stimulated
HUVECs. It has been reported that endothelial cells in PCa tissue
from TRAMP mice, a spontaneous PCa mouse model, have a
constitutively high baseline level of activity of Rho GTPase and
its downstream effector ROCK (17).
This suggests that the RhoA/ROCK pathway plays a crucial role in
PCa angiogenesis. HUVECs cultured in PC3CM share some of the
characteristics of PCa endothelium and can therefore be used to
represent it.
Angiogenesis involves a complex series of events
that take place in a multi-step process. Endothelial cells migrate
through the basement membrane toward an angiogenic stimulus. The
leading front of migrating cells is driven by enhanced
proliferation of endothelial cells, followed by the formation of
capillary tubes via endothelial cell organization. The RhoA/ROCK
pathway plays a role in each of these steps.
We evaluated the effects of fasudil on each of these
steps in PCa-induced HUVECs. Fasudil was found to inhibit
PC3CM-induced HUVEC proliferation, migration, tube formation and
spheroid sprouting. This is in accordance with previous studies on
VEGF-induced endothelial cell proliferation, migration and tube
formation after treatment with the RhoA inhibitor, C3, or ROCK
inhibitors, Y-27632 and fasudil (15,16,18).
It is interesting to note the morphological changes
that occurred in the spheroid sprouting assay after treatment with
fasudil. After treatment with 10 μM fasudil, the sprouts were much
thinner than those on untreated cells. However, the HUVEC nucleus
was observed less frequently moving out of the spheroids than in
the controls. The movement of the nucleus decreased as the fasudil
concentration increased, whereas sprouting was not affected until
the concentration of fasudil exceeded 30 μM. These sprouts were
more akin to cell protrusions, were markedly thinner compared with
PC3CM-induced HUVEC sprouts, and were more abundant and
disorganized compared with the ordered architecture of single HUVEC
spheroid sprouting.
In conclusion, fasudil significantly inhibits the
key steps of endothelial cell angiogenesis, including
proliferation, migration and capillary tube formation, in a
dose-dependent manner. These effects may be due to inhibition of
ROCK activity induced by PCa cell secretions. Fasudil may be a
useful anti-angiogenic agent and should be investigated further for
its potential role in the anti-angiogenic treatment of PCa.
Acknowledgements
This study was supported by a grant from The Program
of International Science and Technology Cooperation (grant no.
2012DFG31440), awarded by The Ministry of Science and Technology,
P.R. China. The authors are grateful to NewMed Publishing Services
for providing final editing services.
References
|
1
|
Bubendorf L, Schöpfer A, Wagner U, et al:
Metastatic patterns of prostate cancer: An autopsy study of 1,589
patients. Hum Pathol. 31:578–583. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Roehl KA, Han M, Ramos CG, Antenor JA and
Catalona WJ: Cancer progression and survival rates following
anatomical radical retropubic prostatectomy in 3,478 consecutive
patients: long-term results. J Urol. 172:910–914. 2004. View Article : Google Scholar
|
|
3
|
Hull GW, Rabbani F, Abbas F, Wheeler TM,
Kattan MW and Scardino PT: Cancer control with radical
prostatectomy alone in 1,000 consecutive patients. J Urol.
167:528–534. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Daneshgari F and Crawford ED: Endocrine
therapy of advanced carcinoma of the prostate. Cancer.
71:1089–1097. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Eymard JC, Oudard S, Gravis G, et al:
Docetaxel reintroduction in patients with metastatic
castration-resistant docetaxel-sensitive prostate cancer: a
retrospective multicentre study. BJU Int. 106:974–978. 2010.
View Article : Google Scholar
|
|
6
|
Bigler SA, Deering RE and Brawer MK:
Comparison of microscopic vascularity in benign and malignant
prostate tissue. Hum Pathol. 24:220–226. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stefanou D, Batistatou A, Kamina S,
Arkoumani E, Papachristou DJ and Agnantis NJ: Expression of
vascular endothelial growth factor (VEGF) and association with
microvessel density in benign prostatic hyperplasia and prostate
cancer. In Vivo. 18:155–160. 2004.PubMed/NCBI
|
|
8
|
Weidner N, Carroll PR, Flax J, Blumenfeld
W and Folkman J: Tumor angiogenesis correlates with metastasis in
invasive prostate carcinoma. Am J Pathol. 143:401–409.
1993.PubMed/NCBI
|
|
9
|
Borre M, Offersen BV, Nerstrom B and
Overgaard J: Microvessel density predicts survival in prostate
cancer patients subjected to watchful waiting. Br J Cancer.
78:940–944. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Green MM, Hiley CT, Shanks JH, et al:
Expression of vascular endothelial growth factor (VEGF) in locally
invasive prostate cancer is prognostic for radiotherapy outcome.
Int J Radiat Oncol Biol Phys. 67:84–90. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bok RA, Halabi S, Fei DT, et al: Vascular
endothelial growth factor and basic fibroblast growth factor urine
levels as predictors of outcome in hormone-refractory prostate
cancer patients: a cancer and leukemia group B study. Cancer Res.
61:2533–2536. 2001.
|
|
12
|
Peyromaure M, Camparo P, Badoual C,
Descazeaud A and Dinh-Xuan AT: The expression of vascular
endothelial growth factor is associated with the risk of cancer
progression after radical prostatectomy. BJU Int. 99:1150–1153.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
George DJ, Halabi S, Shepard TF, et al:
Prognostic significance of plasma vascular endothelial growth
factor levels in patients with hormone-refractory prostate cancer
treated on Cancer and Leukemia Group B 9480. Clin Cancer Res.
7:1932–1936. 2001.
|
|
14
|
Etienne-Manneville S and Hall A: Rho
GTPases in cell biology. Nature. 420:629–635. 2002. View Article : Google Scholar
|
|
15
|
van Nieuw Amerongen GP, Koolwijk P,
Versteilen A and van Hinsbergh VW: Involvement of RhoA/Rho kinase
signaling in VEGF-induced endothelial cell migration and
angiogenesis in vitro. Arterioscler Thromb Vasc Biol. 23:211–217.
2002.PubMed/NCBI
|
|
16
|
Yin L, Morishige K, Takahashi T, et al:
Fasudil inhibits vascular endothelial growth factor-induced
angiogenesis in vitro and in vivo. Mol Cancer Ther. 6:1517–1525.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ghosh K, Thodeti CK, Dudley AC, Mammoto A,
Klagsbrun M and Ingber DE: Tumor-derived endothelial cells exhibit
aberrant Rho-mediated mechanosensing and abnormal angiogenesis in
vitro. Proc Natl Acad Sci USA. 105:11305–11310. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bryan BA, Dennstedt E, Mitchell DC, et al:
RhoA/ROCK signaling is essential for multiple aspects of
VEGF-mediated angiogenesis. FASEB J. 24:3186–3195. 2010. View Article : Google Scholar : PubMed/NCBI
|