Introduction
Hepatocellular carcinoma (HCC) is the most common
primary liver cancer and one of the most common malignancies in the
world, accounting for approximately one million deaths per year
(1). Although liver resection and
local ablation are regarded as potentially curative treatment
(2), its prognosis is poor. Most of
the patients are diagnosed with advanced disease at presentation
for which palliative therapy forms the mainstay of treatment
(3). To improve this situation, the
development of novel molecular therapies against effective targets
is an urgent issue.
Toward this direction, we previously cloned a novel
tumor associated gene HTA (4). To
further elucidate its mechanism in hepatoma carcinogenesis and its
potential role as a cancer biomarker, research was carried out. The
results showed that knockdown of endogenous HTA expression in the
malignant hepatic cell line HepG2 (HepG2/si-HTA) by small
interfering RNA (siRNA), attenuated cell growth, weakened its tumor
formation ability in nude mice (5)
while overexpression of the HTA gene in the hepatic cell line
QSG-7701 (QSG-7701/HTA) via stable transfection promoted its
proliferation rate, colony forming ability and altered the cell
cycle distribution of the cell lines (6). Furthermore, in a previously study, a
cDNA microarray analysis identified that many genes were
differentially expressed in HepG2/si-HTA and QSG-7701/HTA cells.
Following the knockdown and overexpression of HTA, the expression
level of HOXA7 was changed (Li et al, unpublished data),
which suggests that HTA and HOXA7 may be important in HCC
development and progression.
Mammals have 39 HOX genes split between four groups
(A-D) of linked genes on different chromosomes, which are thought
to have arisen from a series of duplication events (7). HOX genes encode morphoregulatory
transcription factors that specify positional identity during
development and regulate differentiation, motility, adhesion and
proliferation in adult tissues (8).
Accumulating evidence suggests that aberrantly expressed HOX genes
may play important roles in the biology of a variety of solid
tumors and hematological malignancies (9–11).
Although numerous studies have documented differences in HOX gene
expression between normal and neoplastic tissues, relatively few
studies have defined specific functional roles and mechanisms of
action for HOX genes in human HCC.
The expression of several HOX genes was found to be
altered in human HCC compared to that in normal tissue (12,13).
Overexpression of HOXA13 in human HCC cells was associated with the
poorest prognosis of hepatocarcinomas (14). Cell biological approach demonstrated
that misexpression of some HOX genes altered the malignant
potential of human tumor cells in culture. Enforced expression of
HOXA1 in human breast cancer cells resulted in enhancement of cell
proliferation (15). Knockdown of
HOXC10 expression by shRNA decreased the invasiveness of cervical
cancer cells (16). Overexpression
of HOXB7 in immortalized ovarian surface epithelial cells was found
to enhance cellular proliferation (17), whereas overexpression of antisense
to HOXB7 and HOXB13 suppressed Transwell invasion of SKOV-3 ovarian
cancer cells (18). Forced
overexpression of HOXD3 enhanced the invasive and metastatic
abilities of lung cancer cells through the activation of integrin
αvβ3 and TGF-β pathways (19–21).Thus, accumulating evidence suggests
that HOX genes are involved in oncogenesis and malignant
progression.
In the present study, through in vitro and
in vivo expriments we investigated the role of HOXA7 in the
regulation of proliferation in human HCC. The results revealed an
important role for HOXA7 in promoting human HCC progression,
mediated by cyclin E1 and CDK2.
Materials and methods
Cell culture
HCC cell lines, HepG2, Hep3B-2 and QGY-7703, and
normal liver cell line, QSG-7701, were maintained in our laboratory
and cultured in RPMI-1640 medium supplemented with 10% fetal bovine
serum (FBS) (both from Gibco). Cells were maintained at 37°C in an
atmosphere of humidified air with 5% CO2.
Reverse transcription-PCR (RT-PCR)
RNA isolated from the cells was reverse-transcribed
and amplified using the One-Step RT-PCR System (Fermentas, Vilnius,
Lithuania). The sets of primers for HOXA7 were: sense,
5′-CTTATACAATGTCAA CAGCC-3′ and antisense,
5′-TCCTTATGCTCTTTCTTCC-3′, 414 bp; sense,
5′-AATCCCATCACCATCTTCCA-3′ and antisense,
5′-CCTGCTTCACCACCTTCTTG-3′ for GAPDH, 580 bp. After heating at 95°C
for 1 min, the samples were exposed to 30 cycles (GAPDH, 25 cycles)
of 95°C for 30 sec, 56°C for 30 sec and 72°C for 1 min 30 sec, with
a final extension at 72°C for 10 min. Reaction products were
separated on 1% agarose gels containing ethidium bromide, and the
level of amplification was analyzed using a Phosphor Imager.
Western blot analysis
The cells were washed with cold phosphate-buffered
saline (PBS) and lysed in Laemmli buffer (62.5 mM Tris-HCl, pH 6.8,
2% SDS, 10% glycerol, 50 mM dithiothreitol and 0.01% bromophenol
blue) for 10 min at 100°C. Cell lysates were analyzed by SDS/PAGE
and transferred electrophoretically to a polyvinylidene difluoride
membrane. The blots were probed with specific antibodies by a
secondary detection step. The immunoreactive proteins were revealed
by an ECL kit. Western blotting was carried out using the following
antibodies: rabbit anti-HOXA7 antibody (Santa Cruz Biotechnology),
rabbit anti-cyclin D1 antibody (Cell Signaling Biotechnology),
rabbit anti-cyclin E1 antibody (Abcam Biotechnology), rabbit
anti-CDK4 antibody, and rabbit anti-CDK2 antibody (both from Santa
Cruz Biotechnology).
Knockdown and overexpression of HOXA7 and
vector construction
To knock down HOXA7 expression, we used the
GV102-U6/Neo/GFP vector encoding a small hairpin RNA directed
against the target gene in the HepG2 and QGY-7703 cells. The target
sequences for HOXA7 were 5′-TCGCCCACACGCTCTGTTT-3′. We used a
negative universal control as a negative control (NC).
For transfection of the plasmid expression vector
encoding human HOXA7, the DNA sequencing containing the full-length
HOXA7 (917 bp) open reading frame flanked by BamHI (sense)
and EcoRI (antisense) restriction sites was PCR amplified
from the HepG2 cells. The primer sequences used were sense,
5′-TCATTCCTCCTCGTCC-3′ and antisense,
5′-ATGAGTTCTTCGTATTATGAACG-3′. The resulting fragment was inserted
into pcDNA3.1(+) to generate pcDNA3.1-HOXA7. The desired sequence
was confirmed by direct DNA sequencing.
Cell transfection
For transfection, the HCC and normal liver cell
lines were grown to 70% confluency and transfected in serum-free
medium for 6 h with FuGENE6 (Roche) and the vector. After 48 h, the
cells were subjected to G418 selection (600 μg/ml), followed by
limited dilution in 96-well plates for the generation of individual
cell clones. The cells were harvested 15 days later to analyze the
expression of HOXA7 by RT-PCR using the primers listed above and by
western blotting using the rabbit antibodies listed above.
MTT analysis
For the MTT analysis, cells were seeded with
RPMI-1640 medium with 1% FBS at a density of 103
cells/well in 96-well plates (n=6), grown overnight, washed in PBS
and incubated in RPMI-1640 with 10% FBS at 37°C in 5%
CO2 for varying periods and exposed to fresh media every
other day. During the last 4 h of each day of culture, the cells
were treated with methyl thiazolyl tetrazolium (MTT; 50 μg/well;
Sigma, USA). The generated formazan was dissolved in dimethyl
sulfoxide (DMSO) and measured as OD (490 nm) for detecting the cell
viability.
Colony formation analysis
For the colony formation analysis, the cells at
1,000 cells/well in 6-cm plates were incubated with RPMI-1640
medium with 1% FBS for 24 h, and then cultured in RPMI-1640 medium
with 10% FBS at 37°C at 5% CO2 for 2 weeks. The cell
colonies were then washed twice with PBS, fixed with 4%
paraformaldehyde for 15 min and stained with Giemsa for 30 min.
Individual clones consisting of >50 cells were counted. Clone
forming efficiency for each individual type of cells was
calculated, according to the number of colonies/the number of
inoculated cells × 100%.
Flow cytometric analysis
For the flow cytometric analysis, the cells were
incubated with RPMI-1640 medium with 1% FBS for 24 h, and then
cultured in RPMI-1640 medium with 10% FBS for 24 h (n=3). The cells
were harvested and resuspended in fixation fluid at a density of
106/ml. Propidium iodide (PI) (1,500 μl) solution was
added, and the cell cycle was detected by FACSCalibur
(Becton-Dickinson).
Tumor formation in the nude mice
To test the influence of HOXA7 on the development of
HCC, the tumor formation in nude mice was examined. Briefly,
HepG2/si-HOXA7, HepG2/NC, QGY-7703/si-HOXA7 and QGY-7703/NC cells
(5×106) were injected subcutaneously into 4-week-old
BALB/c nude mice (n=4/group; Shanghai Laboratory Animal Center,
Shanghai, China). The experimental pairs (HepG2/si-HOXA7, HepG2/NC,
QGY-7703/si-HOXA7 and QGY-7703/NC) were carried out in the
different mice. The development and growth of solid tumors were
monitored by measuring the tumor size using a vernier caliper in a
blinded manner every 5 days for a 40-day period: Tumor volume =
width2 × length × 0.5. At the end of the experiment, all
mice were sacrificed, and individual tumor weights were
measured.
Statistical analysis
All data are expressed as means ± standard
deviation. Difference among groups were determined by ANOVA, and
the comparison between two groups was analyzed by the Student’s
t-test using GraphPad Prism software version 4.0 (GraphPad
Software, Inc., San Diego, CA, USA). A probability value of
P<0.05 was considered to indicate a statistically significant
result.
Results
mRNA and protein expression of HOXA7 in
the HCC and normal liver cell lines
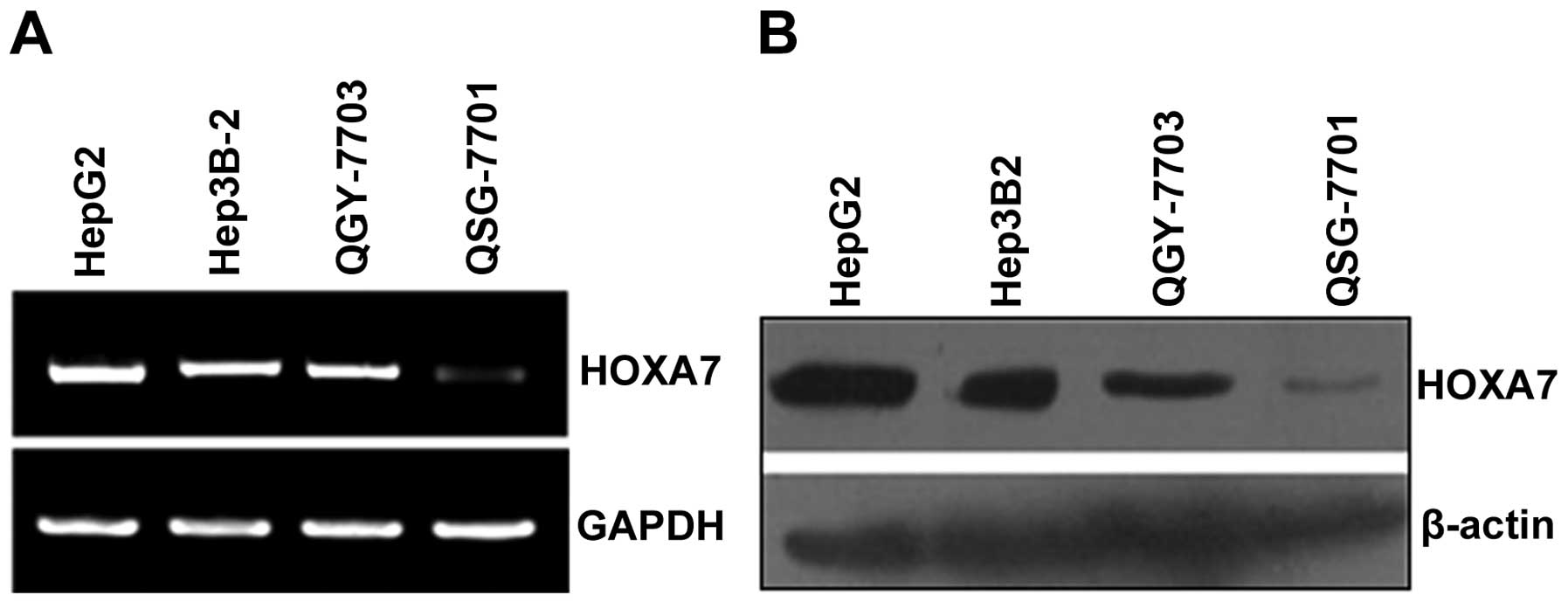
We assessed HOXA7 mRNA and protein expression in the
HCC HepG2, Hep3B-2 and QGY-7703 cell lines and in the normal liver
QSG-7701 cell line. The results of the semi-quantitative RT-PCR and
western blotting showed that HOXA7 mRNA was detected in the HepG2,
Hep3B-2, QGY-7703 and QSG-7701 cells, while HOXA7 protein was
mainly expressed in the HepG2, Hep3B-2 and QGY-7703 cells (Fig. 1A and B).
Impact of the knockdown of HOXA7 on the
proliferation of HCC cells
To study the biological function of HOXA7, we
generated the RNAi vector containing siRNA specifically targeting
HOXA7 to stably knock down the endogenous expression of HOXA7 in
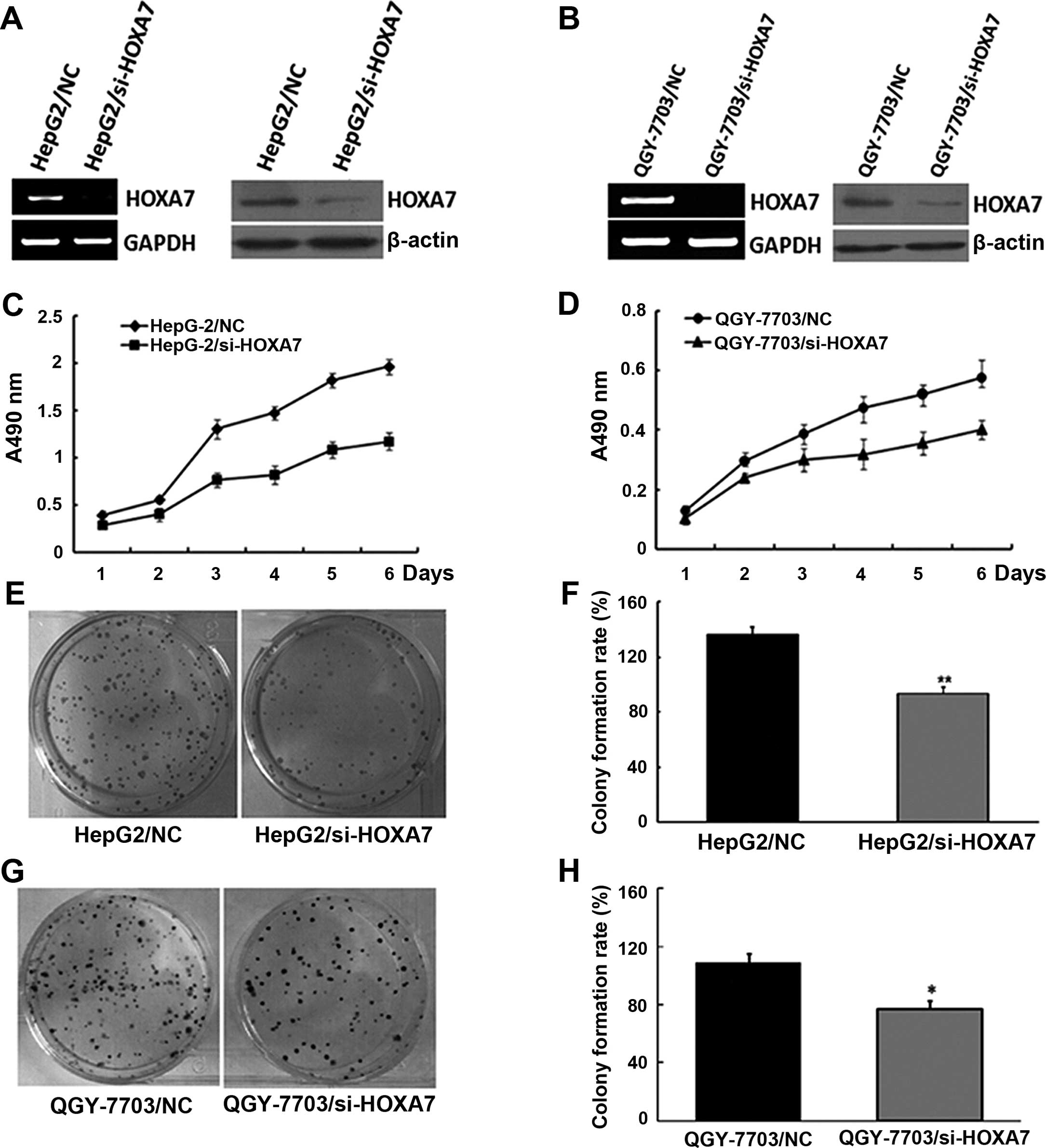
the HepG2 and QGY-7703 cells. As shown in Fig. 2A and B, compared to the control
(HepG2/NC and QGY-7703/NC), cells transfected with si-HOXA7 had
significantly decreased levels of HOXA7 mRNA and protein.
To study the impact of si-HOXA7 on the proliferation
of HCC cells, we examined the proliferative efficiency of decreased
HOXA7 in the HepG2 and QGY-7703 cells by MTT analysis. Following a
6-day period, the proliferation of the HepG2/si-HOXA7 and
QGY-7703/si-HOXA7 cells was much slower than that of the HepG2/NC
and QGY-7703/NC cells, and significantly high numbers of HepG2/NC
and QGY-7703/NC cells were observed from day 3 (Fig. 2C and D). A similar pattern of
inhibition following reduced HOXA7 expression in HepG2 and QGY-7703
cells was achieved in the colony formation assay (Fig. 2E–H). Therefore, the low
proliferative activity and the low number of cell colonies in the
HepG2/si-HOXA7 and QGY-7703/si-HOXA7 cells demonstrated that
downregulation of HOXA7 expression inhibited cell proliferation
in vitro.
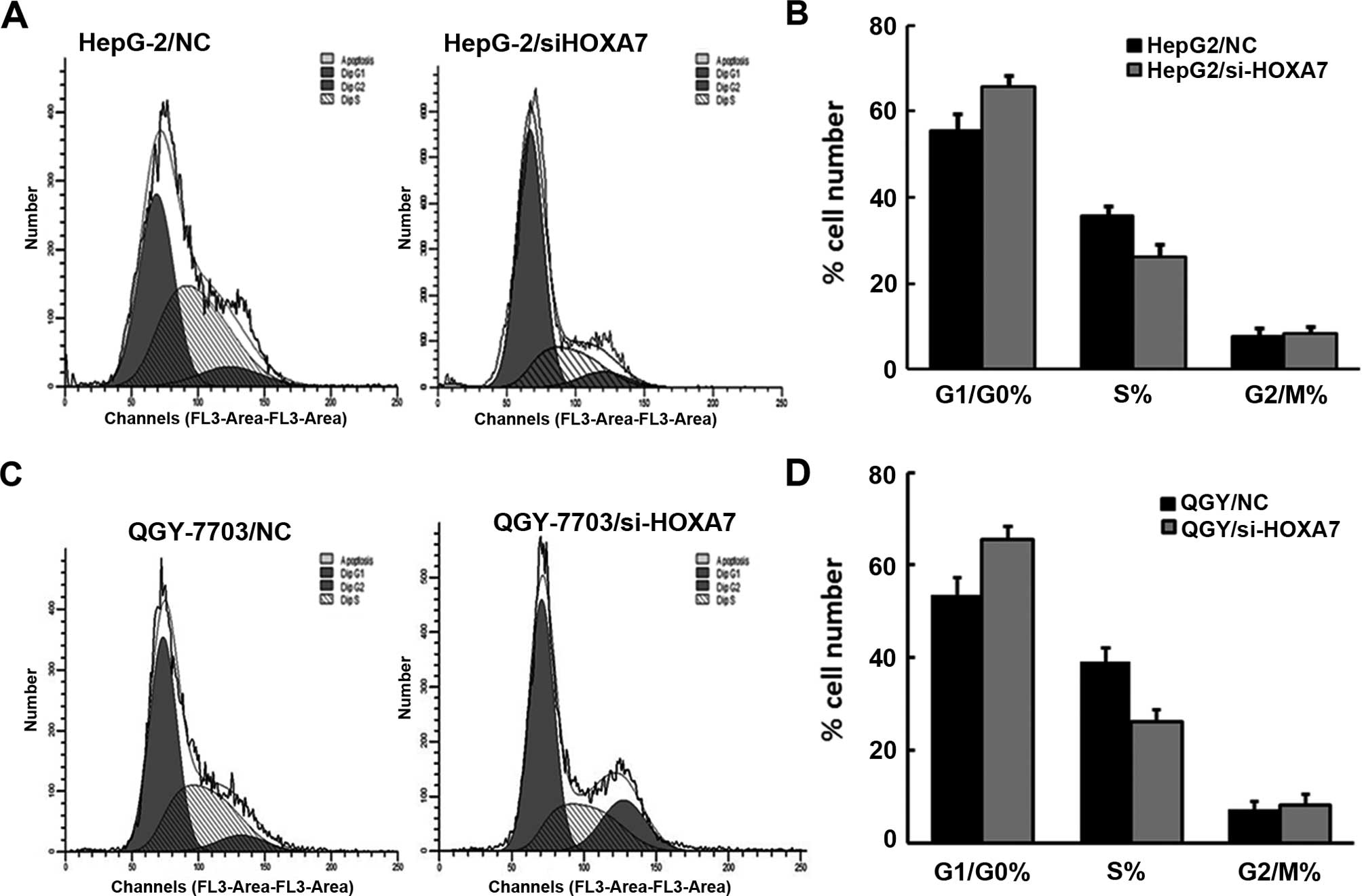
Cell proliferation was also detected by flow
cytometry. The results showed that HepG2 and QGY-7703 cells with
si-HOXA7 exhibited cell cycle arrest in the G1 phase, which
inhibited the proliferation of HepG2 and QGY-7703 cells (Fig. 3A–D). This result was consistent with
the MTT analysis.
Overexpression of HOXA7 promotes QSG-7701
cell proliferation
Given that downregulation of HOXA7 inhibited HepG2
and QGY-7703 cell proliferation in vitro, we hypothesized
that HOXA7 promotes QSG-7701 cell proliferation. To test this,
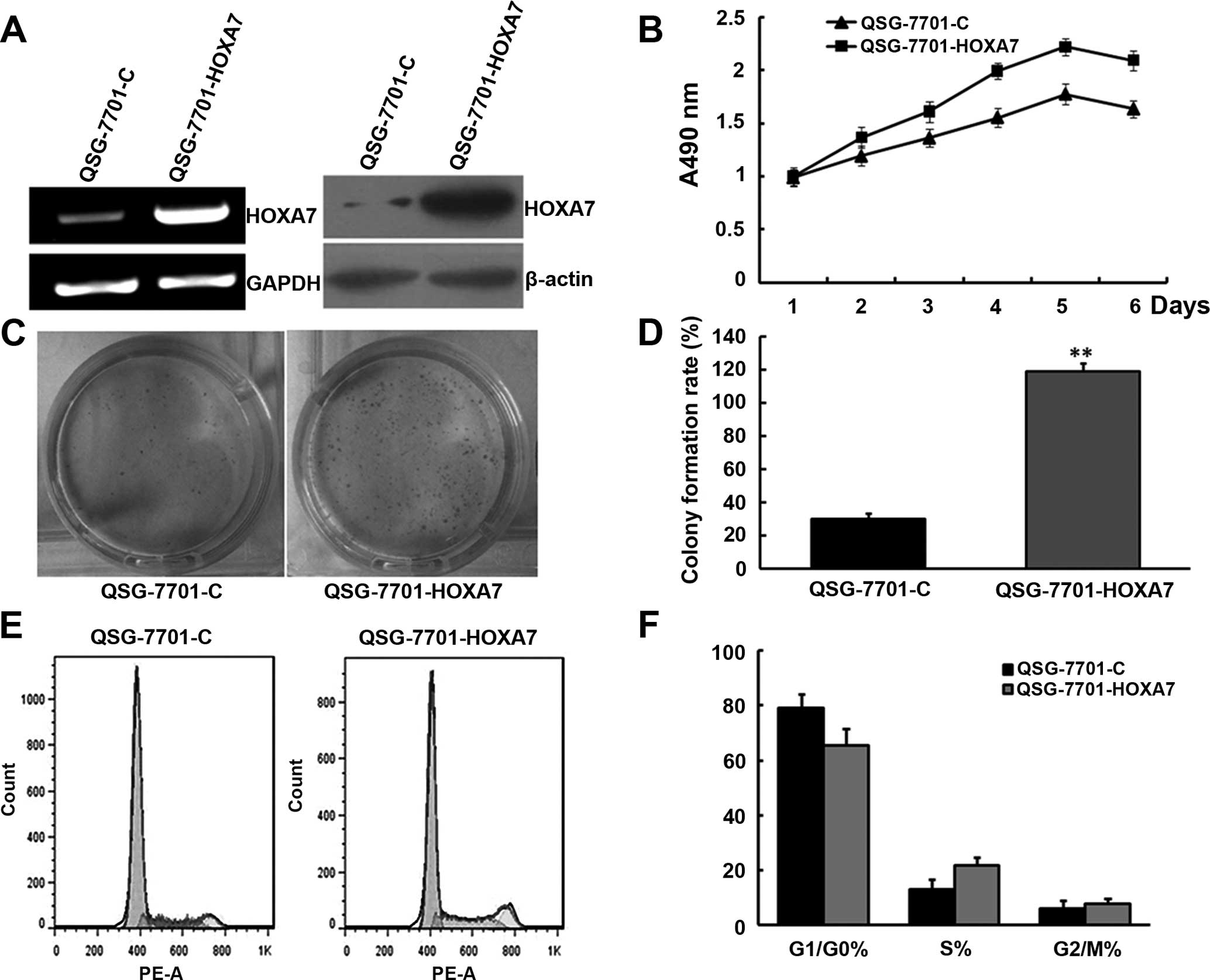
QSG-7701 cells were transfected with a plasmid encoding HOXA7.
Compared to the control cells (QSG-7701-C), cells transfected with
the plasmid encoding HOXA7 (QSG-7701-HOXA7) had increased levels of
HOXA7 mRNA and protein (Fig. 4A).
MTT analysis showed that the proliferation rate of the
QSG-7701-HOXA7 cells was much higher than that in the QSG-7701-C
cells (Fig. 4B). Colony formation
analysis revealed a larger number of cell colonies in the
QSG-7701-HOXA7 cells. The results demonstrated that upregulation of
HOXA7 expression promoted cell proliferation in vitro
(Fig. 4C and D).
Cell proliferation was also detected by flow
cytometry. The results showed that QSG-7701/HOXA7 cells underwent
cell cy arrested the cell cycle in the G1 phase and an increased
number of cells were in the S phase of the cell cycle, which
promoted the proliferation of QSG-7701 cells (Fig. 4E and F). This result is in line with
the above analysis.
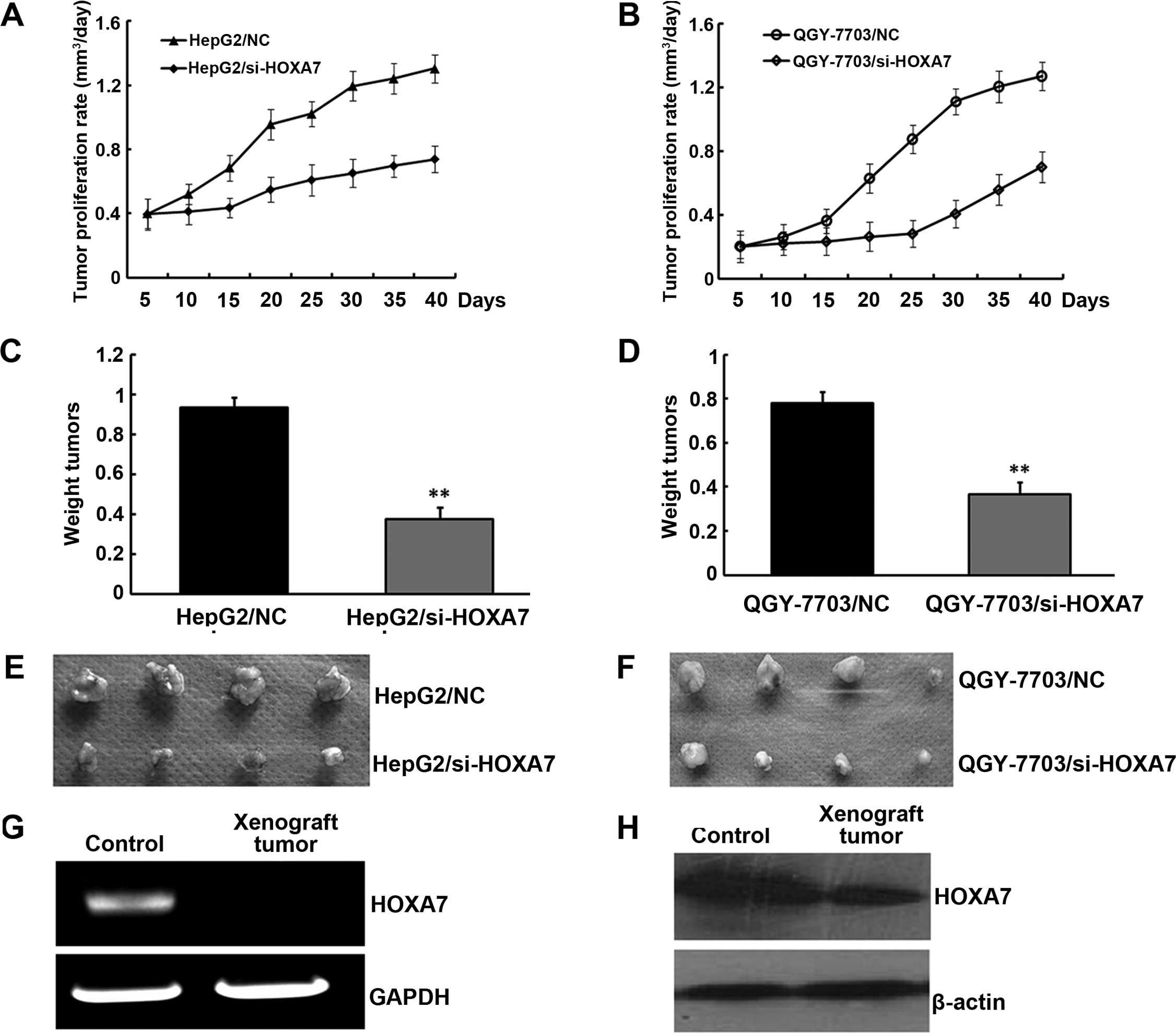
Tumor formation in nude mice
Given that downregulation of HOXA7 inhibited HCC
cell proliferation and upregulation of HOXA7 promoted HCC cell
proliferation in vitro, we hypothesized that HOXA7 promotes
HCC development in vivo. To further determine the role of
HOXA7 in tumorigenesis and development of HCC, HepG2/si-HOXA7 and
QGY-7703/si-HOXA7 or HepG2/NC and QGY-7703/NC cells were injected
subcutaneously into nude mice. The development of the tumors was
monitored for 40 days. As shown in Fig.
5A and B, HOXA7-knockdown tumors emerged later and grew more
slowly compared to the control tumors. At the end of the
experimental period, the final weights of the HOXA7-knockdown
tumors (0.376±0.951 and 0.368±0.093 g) were found to be markedly
lighter than the tumor weights in the controls (0.936±0.98 and
0.736±0.895 g) (Fig. 5C–F). RT-PCR
and western blotting of HOXA7 in the xenograft tumors indicated
that increased HOXA7 expression had been maintained throughout the
experimental time course (Fig. 5G and
H). Collectively, these data indicate that HOXA7 promoted
xenograft tumor development in vivo.
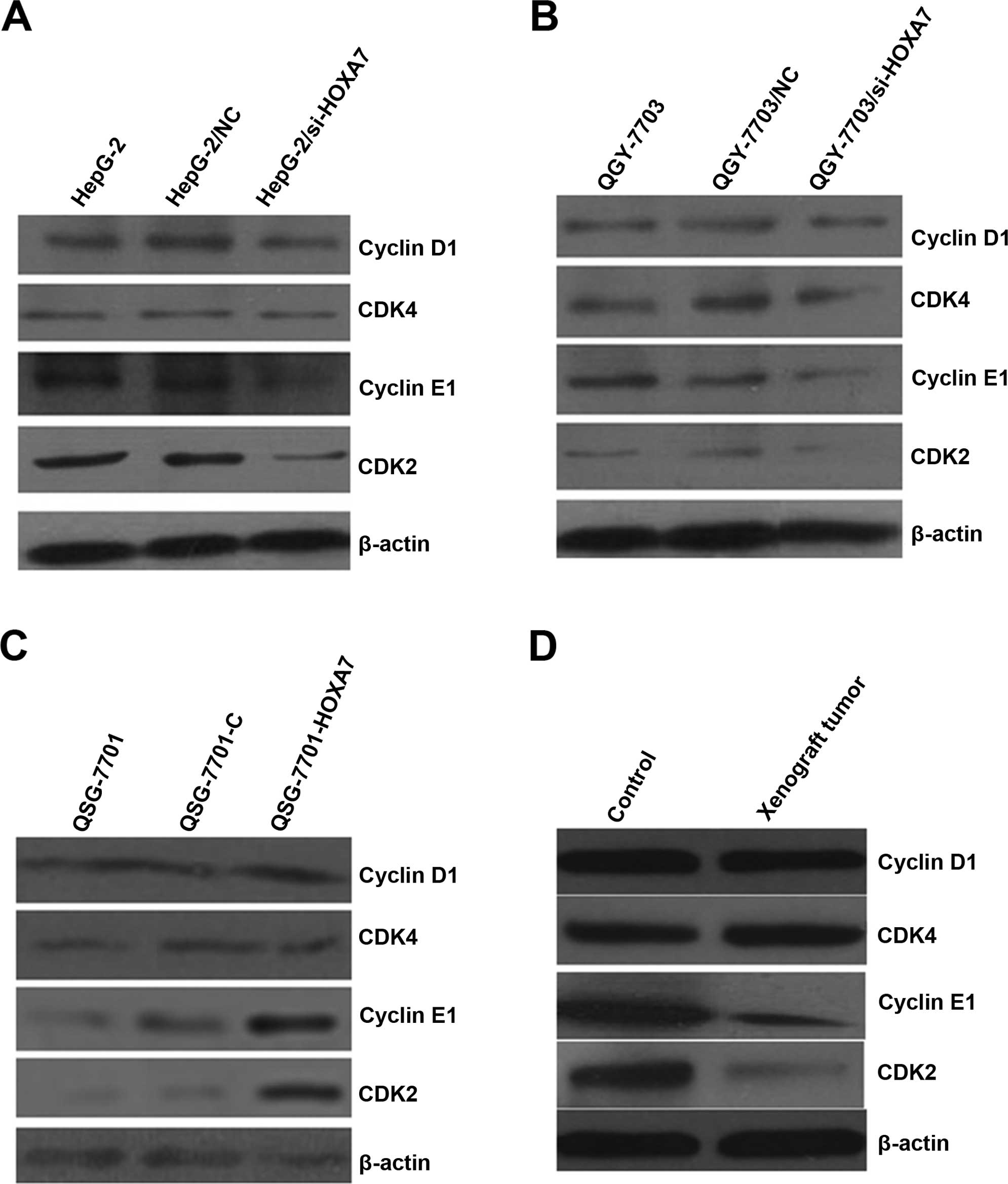
HOXA7 affects the expression of cell
cycle-related proteins in the HCC cells, normal hepatic cells and
xenograft tumors
Cyclin D/CDK4 and cyclin E/CDK2 are important for
cell cycle progression in metazoans and are frequently
overexpressed in cancer cells. In view of the effects of HOXA7 on
cell proliferation, we investigated whether knockdown of HOXA7
influences cyclin D1/CDK4 and cyclin E1/CDK2 expression in the HCC
cells, normal hepatic cells and xenograft tumors. The total
cellular levels of cyclin D1/CDK4, and cyclin E1/CDK2 were analyzed
by western blotting. The results showed that HOXA7 had no
conclusive effect on cyclin D1/CDK4 protein levels. However,
downregulation of endogenous HOXA7 reduced cyclin E1/CDK2 protein
levels (Fig. 6A and B). Moreover,
western blotting also confirmed these results in the QSG-7701-HOXA7
cells and xenograft tumors (Fig. 6C and
D).
Discussion
HOX genes are regulatory genes encoding nuclear
proteins that act as transcription factors, which regulate aspects
of morphogenesis and cell differentiation. There is accumulating
evidence that HOX genes play important roles in oncogenesis,
including ovarian, colon, lung and prostate cancer progression
(22,23). However, the specific role of the HOX
genes in HCC cell function and oncogenesis has not been extensively
explored. In the present study, we showed for the first time a
significant role of HOXA7 in the regulation of HCC cell
proliferation and cyclin E1/CDk2 expression. These findings
indicate a new mechanism for HOX-mediated cell proliferation
through the regulation of cyclin E1/CDK2 expression.
We first demonstrated that all three types of HCC
cells express HOXA7. The expression of HOXA7 in the HCC cells
indicated that HOXA7 exerted a potential role in cell regulation.
In the present study, HepG-2 and QGY-7703 cells were used as an HCC
tumor cell model, and QSG-7701 cells were used as a normal hepatic
cell model. Quantitative analysis showed that the expression level
of HOXA7 was significantly higher in the HCC tumor cells. It has
been reported that many cancers, including leukemia, colon, skin,
prostate, breast and ovarian cancers, have shown alterations in the
expression patterns of HOX genes (24). The overexpression of HOX genes has
been widely associated with a variety of carcinomas (25). Therefore, our results are consistent
with previous observations that HOX genes are highly expressed in a
variety of carcinomas. HOXA7, in particular, plays a role in HCC
cell proliferation. In the present study, we performed
siRNA-mediated knockdown of HOXA7 in a high HOXA7-expressing HCC
cell line. Conversely, we overexpressed HOXA7 in a low
HOXA7-expressing QSG-7701 cell line to determine how HOXA7
regulates hepatic cell growth. The results showed that HOXA7
exerted a specific effect on HCC cell proliferation; knockdown of
HOXA7 induced a decrease in HCC proliferation, while overexpression
of HOXA7 in QSG-7701 cells significantly promoted cell growth. A
previous study indicated that HOXA7 expression was increased during
mitosis in cultured granulosa cells. Moreover, growth
differentiation factor-9 (GDF-9), which enhances early follicular
growth and differentiation, increased HOXA7 protein expression and
regulated the expression of HOXA7 cofactors in granulosa cells
(26,27). These results support the hypothesis
that HOXA7 modulates the growth and oncogenesis of human HCC
cells.
Since cell proliferation is controlled by
progression through the cell cycle, which is regulated by many
proliferative signaling cascades, we carried out flow cytometric
analysis to assess cell cycle distribution. The normal cell cycle
follows ordinary steps, while cancer cells grow without regulation.
The rate of progression in the cell cycle is decided by cyclins and
cyclin-dependent kinases (CDKs). The entering of each phase is
controlled by a specific cyclin-CDK complex. CDK is a member of the
serine-threonine kinase family since a cyclin binds to a CDK and
starts the phosphorylation of its serine and threonine site
(28,29). It has been reported that the G1-S
checkpoint is mainly regulated by a series of cyclins and CDK,
which are predominantly cyclin D/CDK4, CDK6, and cyclin E/CDK2
(30). In HCC, cyclin E1 is an
ideal candidate to be suppressed as an anti-HCC strategy as it is
overexpressed in ~70% of HCC patients (31); overexpression was found to be
directly correlated with HCC grade and a poor prognosis (32). The cyclin E-CDK2 complex guides the
process from G1 to S phase, while the cyclin A-CDK2 complex is
required to pass through the S phase (33,34).
In the present study, we showed that HOXA7 triggered cell cycle
arrest in the G0/G1 phase. Cyclin E1/CDK2 is known to be involved
in numerous cellular processes such as growth, motility and
differentiation (35). The
expression levels of cyclin E1/CDK2 have been correlated with the
pathogenesis of a broad range of human types of cancers (36). In the present study, we examined the
effect of cyclin E1/CDK2 on HCC cell growth. As observed in
multiple cell types (37), cyclin
E1/CDK2 activation was found to promote cell proliferation and
stimulate the replication of cells. In the present study, the
further identified cyclin E1/CDK2 as a novel downstream target of
HOXA7.
In summary, our present data support a novel
mechanistic role for the HOXA7 modulation of HCC cell proliferation
via cyclin E1/CDK2. This finding indicates the effect of HOXA7 on
HCC cell growth, motility and cellular proliferation, thus yielding
new potential targets for the treatment of HCC tumors, and other
cell proliferation dysfunctions of the cancers that are involved in
the aberrant expression of HOX genes.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (81201903), the Open-End Fund for the
Valuable and Precision Instruments of Central South University
(CSU2C2013048), Hunan, China.
References
|
1
|
Hao K, Luk JM, Lee NP, et al: Predicting
prognosis in hepatocellular carcinoma after curative surgery with
common clinicopathologic parameters. BMC Cancer. 9:3892009.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Song TJ, Ip EW and Fong Y: Hepatocellular
carcinoma: current surgical management. Gastroenterology. 127(Suppl
1): S248–S260. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Paul SB, Gamanagatti SR, Mukund A, Abbas
SZ and Acharya SK: Transarterial chemoembolization for
hepatocellular carcinoma: significance of extrahepatic collateral
supply. Indian J Cancer. 48:339–344. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu Y, Zhao Y, Fang S, Li Y and Li G:
Molecular cloning and alternative splicing analysis of hepatoma
associated gene HTA. Zhong Nan Da Xue Xue Bao Yi Xue Ban.
38:869–875. 2013.(In Chinese). PubMed/NCBI
|
|
5
|
Liu Y, Li Y, Guo F, et al: Identification
of HTA as a novel-specific marker for human hepatocellular
carcinoma. J Cancer Res Clin Oncol. 136:1187–1192. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu Y, Zhao Y, Ju Q, et al: Molecular
clone and functional study of a novel hepatoma associated gene. Int
J Oncol. 42:1105–1112. 2013.PubMed/NCBI
|
|
7
|
Scott MP: A rational nomenclature for
vertebrate homeobox (HOX) genes. Nucleic Acids Res. 21:1687–1688.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kawazoe Y, Sekimoto T, Araki M, Takagi K,
Araki K and Yamamura K: Region-specific gastrointestinal Hox code
during murine embryonal gut development. Dev Growth Differ.
44:77–84. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Abate-Shen C: Deregulated homeobox gene
expression in cancer: cause or consequence? Nat Rev Cancer.
2:777–785. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Samuel S and Naora H: Homeobox gene
expression in cancer: insights from developmental regulation and
deregulation. Eur J Cancer. 41:2428–2437. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Argiropoulos B and Humphries RK: Hox genes
in hematopoiesis and leukemogenesis. Oncogene. 26:6766–6776. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kanai M, Hamada J, Takada M, et al:
Aberrant expressions of HOX genes in colorectal and hepatocellular
carcinomas. Oncol Rep. 23:843–851. 2010.PubMed/NCBI
|
|
13
|
Cillo C, Schiavo G, Cantile M, et al: The
HOX gene network in hepatocellular carcinoma. Int J Cancer.
129:2577–2587. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Boyault S, Rickman DS, de Reynies A, et
al: Transcriptome classification of HCC is related to gene
alterations and to new therapeutic targets. Hepatology. 45:42–52.
2007. View Article : Google Scholar
|
|
15
|
Zhang X, Zhu T, Chen Y, Mertani HC, Lee KO
and Lobie PE: Human growth hormone-regulated HOXA1 is a human
mammary epithelial oncogene. J Biol Chem. 278:7580–7590. 2003.
View Article : Google Scholar
|
|
16
|
Zhai Y, Kuick R, Nan B, et al: Gene
expression analysis of preinvasive and invasive cervical squamous
cell carcinomas identifies HOXC10 as a key mediator of invasion.
Cancer Res. 67:10163–10172. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Naora H, Yang YQ, Montz FJ, Seidman JD,
Kurman RJ and Roden RB: A serologically identified tumor antigen
encoded by a homeobox gene promotes growth of ovarian epithelial
cells. Proc Natl Acad Sci USA. 98:4060–4065. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yamashita T, Tazawa S, Yawei Z, et al:
Suppression of invasive characteristics by antisense introduction
of overexpressed HOX genes in ovarian cancer cells. Int J Oncol.
28:931–938. 2006.PubMed/NCBI
|
|
19
|
Hamada J, Omatsu T, Okada F, et al:
Overexpression of homeobox gene HOXD3 induces coordinate expression
of metastasis-related genes in human lung cancer cells. Int J
Cancer. 93:516–525. 2001. View
Article : Google Scholar
|
|
20
|
Miyazaki YJ, Hamada J, Tada M, et al:
HOXD3 enhances motility and invasiveness through the
TGF-β-dependent and -independent pathways in A549 cells. Oncogene.
21:798–808. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ohta H, Hamada J, Tada M, et al:
HOXD3-overexpression increases integrin αvβ3 expression and
deprives E-cadherin while it enhances cell motility in A549 cells.
Clin Exp Metastasis. 23:381–390. 2006. View Article : Google Scholar
|
|
22
|
Grier DG, Thompson A, Kwasniewska A,
McGonigle GJ, Halliday HL and Lappin TR: The pathophysiology of HOX
genes and their role in cancer. J Pathol. 205:154–171. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bhatlekar S, Fields JZ and Boman BM: HOX
genes and their role in the development of human cancers. J Mol
Med. 92:811–823. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang Y, Huang Q, Cheng JC, et al:
Homeobox A7 increases cell proliferation by up-regulation of
epidermal growth factor receptor expression in human granulosa
cells. Reprod Biol Endocrinol. 8:612010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nunes FD, de Almeida FC, Tucci R and de
Sousa SC: Homeobox genes: a molecular link between development and
cancer. Pesqui Odontol Bras. 17:94–98. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ota T, Choi KB, Gilks CB, Leung PC and
Auersperg N: Cell type- and stage-specific changes in HOXA7 protein
expression in human ovarian folliculogenesis: possible role of
GDF-9. Differentiation. 74:1–10. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ota T, Asahina H, Park SH, et al: HOX
cofactors expression and regulation in the human ovary. Reprod Biol
Endocrinol. 6:492008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hartwell LH and Weinert TA: Checkpoints:
controls that ensure the order of cell cycle events. Science.
246:629–634. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kamb A, Gruis NA, Weaver-Feldhaus J, et
al: A cell cycle regulator potentially involved in genesis of many
tumor types. Science. 264:436–440. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pines J: Protein kinases and cell cycle
control. Semin Cell Biol. 5:399–408. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jung YJ, Lee KH, Choi DW, et al:
Reciprocal expressions of cyclin E and cyclin D1 in hepatocellular
carcinoma. Cancer Lett. 168:57–63. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Masaki T, Shiratori Y, Rengifo W, et al:
Cyclins and cyclin-dependent kinases: comparative study of
hepatocellular carcinoma versus cirrhosis. Hepatology. 37:534–543.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dehay C and Kennedy H: Cell-cycle control
and cortical development. Nat Rev Neurosci. 8:438–450. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
van den Heuvel S and Harlow E: Distinct
roles for cyclin-dependent kinases in cell cycle control. Science.
262:2050–2054. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu QX, Wang XF, Ikeo K, Hirose S, Gehring
WJ and Gojobori T: Evolutionarily conserved transcription factor
Apontic controls the G1/S progression by inducing cyclin E during
eye development. Proc Natl Acad Sci USA. 111:9497–9502. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gladden AB and Diehl JA: Cell cycle
progression without cyclin E/CDK2: breaking down the walls of
dogma. Cancer Cell. 4:160–162. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rath SL and Senapati S: Why are the
truncated cyclin Es more effective CDK2 activators than the
full-length isoforms? Biochemistry. 53:4612–4624. 2014. View Article : Google Scholar : PubMed/NCBI
|