Introduction
Prostate cancer (PCa) is the most common solid
cancer and the second leading cause of cancer-related deaths in men
(1). It is a clinically
heterogeneous-multifocal disease and the incidents are continuously
rising (2). With the wide usage of
the prostate-specific antigen (PSA)-based screening program, the
majority of PCas is at a localized stage at diagnosis. Although
patients with localized PCa can often be successfully treated with
radical prostatectomy or radiotherapy, an increasing number of PCa
patients may die of hormone refractory and metastatic disease.
Carcinogenesis and the mechanisms influencing the progression and
prognosis of PCa is a multistep process, which is likely a
reflection of the underlying genomic diversity (3). Recent studies of PCa have indicated
significant heterogeneity in the gene expression profiles, genetic
and epigenetic alterations involving tumorigenesis and tumor
progression of human PCa (4–6).
Although genetic alterations are implicated in the activation or
inactivation of cancer-associated genes, several types of
epigenetic alterations have been identified in PCa, such as DNA
methylation, loss of imprinting, and altered histone modification
patterns (7–9).
DNA methylation is the most frequently studied
epigenetic modification in PCa. It is involved not only in the
initiation process of PCa, but also in the progression phase of the
disease (10). Since the high
stability of DNA, the ease of analysis with the present techniques
available and the ability to assess the biomarker in body fluids
such as blood, urine and saliva, an increasing number of studies
has focused on the identification of the DNA methylation-based
biomarkers for PCa (11). With
respect to DNA hypermethylation, this implies that several
tumor-suppressor genes become hypermethylated during the initiation
phase of PCa in a very consistent trend. In contrast, DNA
hypomethylation, which is a decrease in the genomic DNA methylation
level, is commonly linked to activation of oncogenes and
chromosomal instability (12).
Especially in PCa, recent studies have indicated that this
particular epigenetic alteration may be associated with the
advanced metastatic stage. Several genes, including those that
encode glutathione S-transferase π 1 (GSTP1), adenomatous polyposis
coli (APC), Ras association domain-containing protein 1 (RASSF1A)
and prostaglandin-endoperoxide synthase 2 (PTGS2), have been
demonstrated to be hypermethylated in PCa, but not in the normal
prostate tissues (6,13,14).
Similarly, many hypomethylated genes have been identified and
demonstrated to be associated with the recurrent status of PCa
patients (15–17). These findings suggest that the DNA
methylation of these loci may improve the diagnostic efficiency for
PCa. Thus, it is reasonable to hypothesize that the investigation
on the methylation status of certain genes is of great significance
to elucidate the molecular mechanisms involved in the regulation of
tumorigenesis and tumor progression in PCa.
Extracellular matrix metalloproteinase (MMP) inducer
EMMPRIN (also known as CD147), a member of the immunoglobulin
family, is a glycoprotein enriched on the surface of many types of
tumor cells (18). CD147 has been
demonstrated to be involved in tumor invasion and metastasis via
stimulating MMPs synthesis, in neighboring fibroblasts and
malignant cell proliferation via the activation of ERK1/2 and p38
mitogen-activated protein kinases, in enhancing angiogenesis via
vascular endothelial growth factor, in inducing chemoresistant
tumor cells via the production of hyaluronan, and in the resistance
of cancer cells to anoikis through inhibition of Bim (19). The roles of CD147 and MMPs in tumor
invasiveness have been confirmed immunohistochemically in several
types of cancer cells. Especially in PCa, Wang et al
(20) found that CD147
downregulation by RNAi technology could decrease the invasive
capability of the PCa cells; data of our research group (21) and Madigan et al (22) both showed that CD147 could regulate
MMPs and was involved in PCa progression. Hao et al
(23) also reported that CD147
expression may be correlated with drug resistance during PCa
metastasis and may be a useful potential therapeutic target in
advanced disease; our research group further identified the
potential role of CD147 as an independent predictor of biochemical
recurrence, development of metastasis and reduced overall survival
in PCa (24,25). However, the mechanisms that result
in the aberrant expression of CD147 in PCa are not fully
elucidated. Thus, the aim of the present study was to investigate
the CD147 promoter methylation status and the correlation with the
tumorigenicity in PCa cells.
Materials and methods
Cell culture and epigenetic drug
treatment
All the PCa cell lines (PC3, DU145, 22RV1 and Lncap)
and the non-tumorigenic benign human prostatic epithelial cell
lines (P69 and RWPE-1) were provided by Dr Chin-Lee Wu
(Massachusetts General Hospital, Boston, MA, USA). The cells were
grown in RPMI-1640 medium (Cellgro Mediatech, Inc., Manassas, VA,
USA) with the necessary supplements. P69 and PC3 cell lines which
were respectively treated with demethylating agent
5-aza-2′-deoxycytidine (Sigma-Aldrich Corp., St. Louis, MO, USA)
were maintained in a RPMI-1640 medium with 1% fetal bovine serum,
1% 100 U/ml penicillin streptomycin mixture. P69 and PC3 cell lines
were starved for 24 h before drug treatment. The cells were treated
with 5-aza-2′-deoxycytidine in different doses (0, 1, 2, 3, 4 and 5
μM) for 3 days at different time-points (72, 48, 24, 12, 6 and 0
h), respectively. These concentrations are safe for cell culture
without obvious toxicity. The growth medium was changed daily and
the cells were harvested on day 4. All the experiments were
performed in duplicate and repeated twice.
Clinical samples and tissue
processing
The study was approved by the Research Ethics
Committee of Guangzhou First People’s Hospital, Guangzhou Medical
College, China. An informed consent was obtained from all the
patients. All the specimens were handled and made anonymous
according to the ethical and legal standards.
Thirty-one PCa samples obtained from 31 PCa patients
who underwent radical prostatectomy or transurethral resection of
the prostate (TURP) at the Guangzhou First People’s Hospital
(Guangzhou, China) were collected. None of the patients recruited
in this study received chemotherapy or radiotherapy prior to the
surgery. In addition, 10 adjacent benign prostate tissues were
collected as control tissue samples. All the tissues were frozen by
liquid nitrogen and stored at −80°C. All the tissues were checked
by H&E staining before DNA/RNA extraction.
Real-time quantitative reverse
transcriptase PCR
Total RNA was isolated from the cultured cells and
the prostate tissues using TRIzol reagent (Invitrogen, Carlsbad,
CA, USA) according to the manufacturer instructions. Reverse
transcription was carried out using a ReverTra Ace reagent kit
(Toyobo, Osaka, Japan). Regular PCR was used to amplify CD147 and
glyceraldehyde-3-phosphate (GADPH) (internal control). In addition,
real-time quantitative PCR was also performed with a MiniOpticon
Real-Time PCR detection system (Bio-Rad, Hercules, CA, USA) in a
SYBR-Green master mix (Takara, Otsu, Japan). The thermal cycling
conditions comprised 1 cycle at 94°C for 4 min, and 40 cycles at
94°C for 15 sec, 60°C for 15 sec, and 72°C for 15 sec. All the data
were analyzed using Opticon Monitor software (version 3.1; Bio-Rad)
and the expression of CD147 was calculated as relative expression
level to GAPDH using the comparative cycle threshold (CT) method.
The sequences of PCR primers were forward, TGTACATTTTTAAAGGCAG
AGATG and reverse, TTTCTTCCTTCCTGCTTTCAAA.
Western blot analysis
PCa cells and non-tumorigenic benign human prostatic
epithelial cells were lysed in lysis buffer containing M-PER
Mammalian Protein Extraction reagent and Protease Inhibitor
Cocktail kit (both from Thermo Scientific, Rockford, IL, USA).
Protein concentration of the cell lysates was determined by using
the Synergy 2 Multi-Mode microplate reader (BioTek) according to
the instructions of the manufacturer. Equal amounts of protein were
loaded onto 4–20% Tris-glycine gel (Invitrogen) following 130 v for
90 min. Then resolved proteins were electrophoretically transferred
to the Immobilon-P transfer membranes (Millipore, Bedford, MA,
USA). The membranes were blocked with 5% fetal bovine serum in TBST
(100 ml 10X TBST mixed with 900 ml ddH2O and 1 ml
Tween-20) for 1 h at room temperature followed by incubation with
the anti-CD147 and anti-MMP2 rabbit monoclonal antibody (Abcam,
Cambridge, MA, USA) at 4°C overnight. Blots were extensively washed
with TBST 3 times and incubated with 1:3,000 dilution of goat
against rabbit antibody diluted in TBST for 2 h at room
temperature. Bound antibodies were detected with an enhanced
chemiluminescence system (ECL, Amersham).
DNA extraction and sodium bisulfite
genomic sequencing
Genomic DNA from cultured cells and prostate tissues
were extracted with the QIAamp DNA mini kit (Qiagen, Valencia, CA,
USA). All the genomic DNA was purified with the QIAquick PCR
purification kit (Qiagen). Genomic DNA was treated with sodium
bisulfite as described by EpiTect Bisulfite kit (Qiagen) as
described by the instructions of the manufacturer. The sodium
bisulfate-treated DNA was amplified by PyroMark PCR kit (Qiagen)
for region (−940 to −670) of CD147 gene. The primers were (5′-TTT
GGG TGA GTT GAG TTT AGT AGG-3′) and (5′-TAC CAC AAA CTA CTA ACA TCA
CTA AAC-3′) designed using Pyrosequencing Assay Design software,
version 1.0. The PCR conditions were as follows: 95°C for 15 min,
45 cycles at 94°C for 45 sec, 56°C for 30 sec, 72°C for 30 sec and
72°C for 10 min. The PCR products were then subcloned and sequenced
(Life Technologies, Guangzhou, China).
Statistical analysis
SPSS version 13.0 for Windows (SPSS Inc., Chicago,
IL, USA) and SAS 9.1 software (SAS Institute, Cary, NC, USA) was
used for statistical analysis. All the results were confirmed in at
least three separate experiments and expressed as mean ± SD. Data
were analyzed for statistical significance by the Student’s t-test
or the independent samples t-test. The Spearman correlation was
calculated between methylation intensity and mRNA levels of CD147.
Differences were considered to be statistically significant when
the P-value was <0.05.
Results
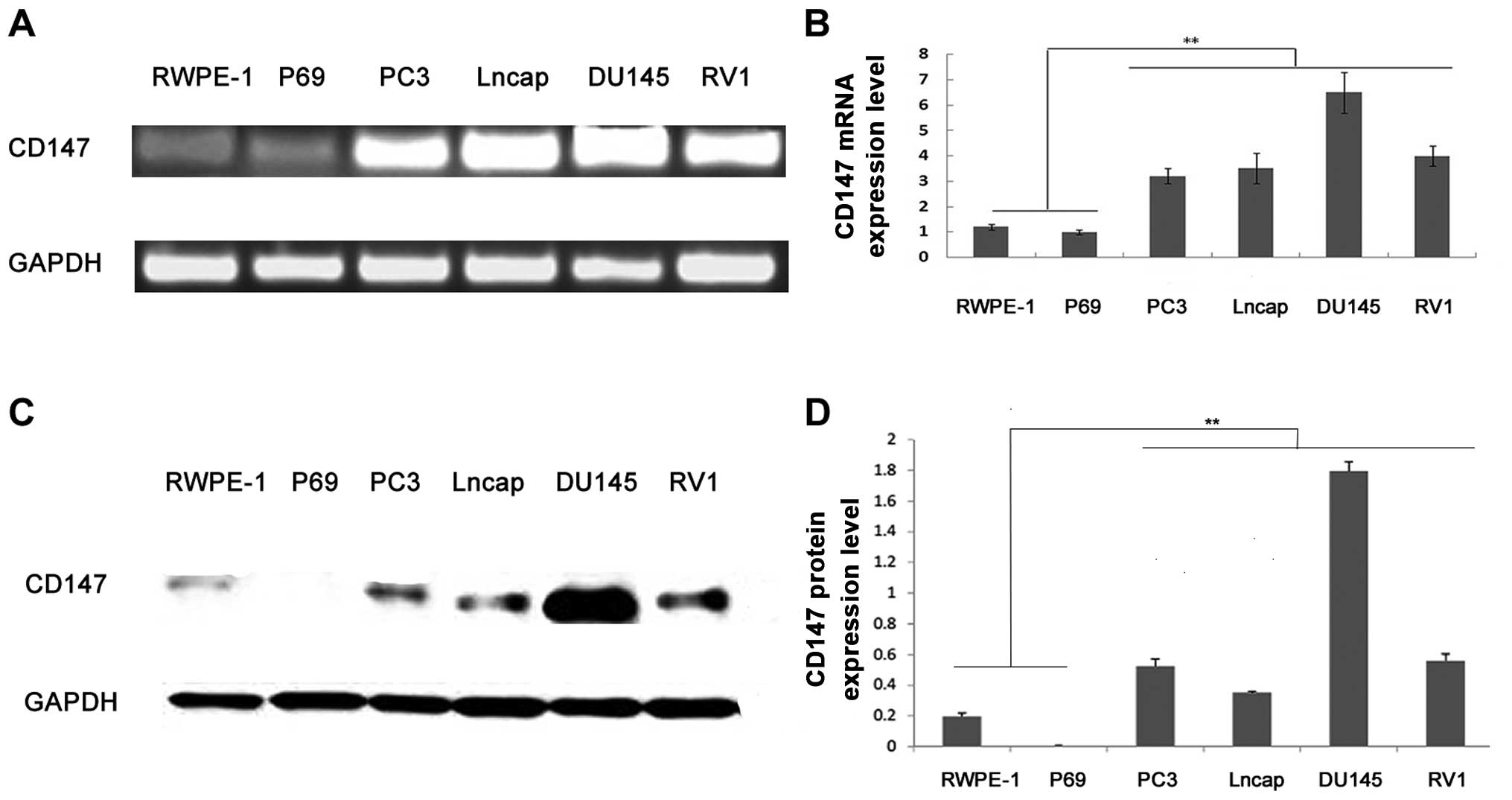
Overexpression of CD147 in PCa cell
lines
Real-time quantitative reverse transcriptase PCR and
western blot analyses were, respectively, performed to detect the
expression of CD147 at mRNA and protein levels in 4 PCa and 2
non-tumorigenic benign human prostatic epithelial cell lines. As
shown in Fig. 1, both CD147 mRNA
and protein were highly expressed in PCa cell lines compared to
non-tumorigenic benign human prostatic epithelial cell lines (all
P<0.01), which were consistent with our previous data in human
PCa and non-cancerous prostate tissues (21,24,25).
DNA promoter hypomethylation of CD147 in
PCa cell lines
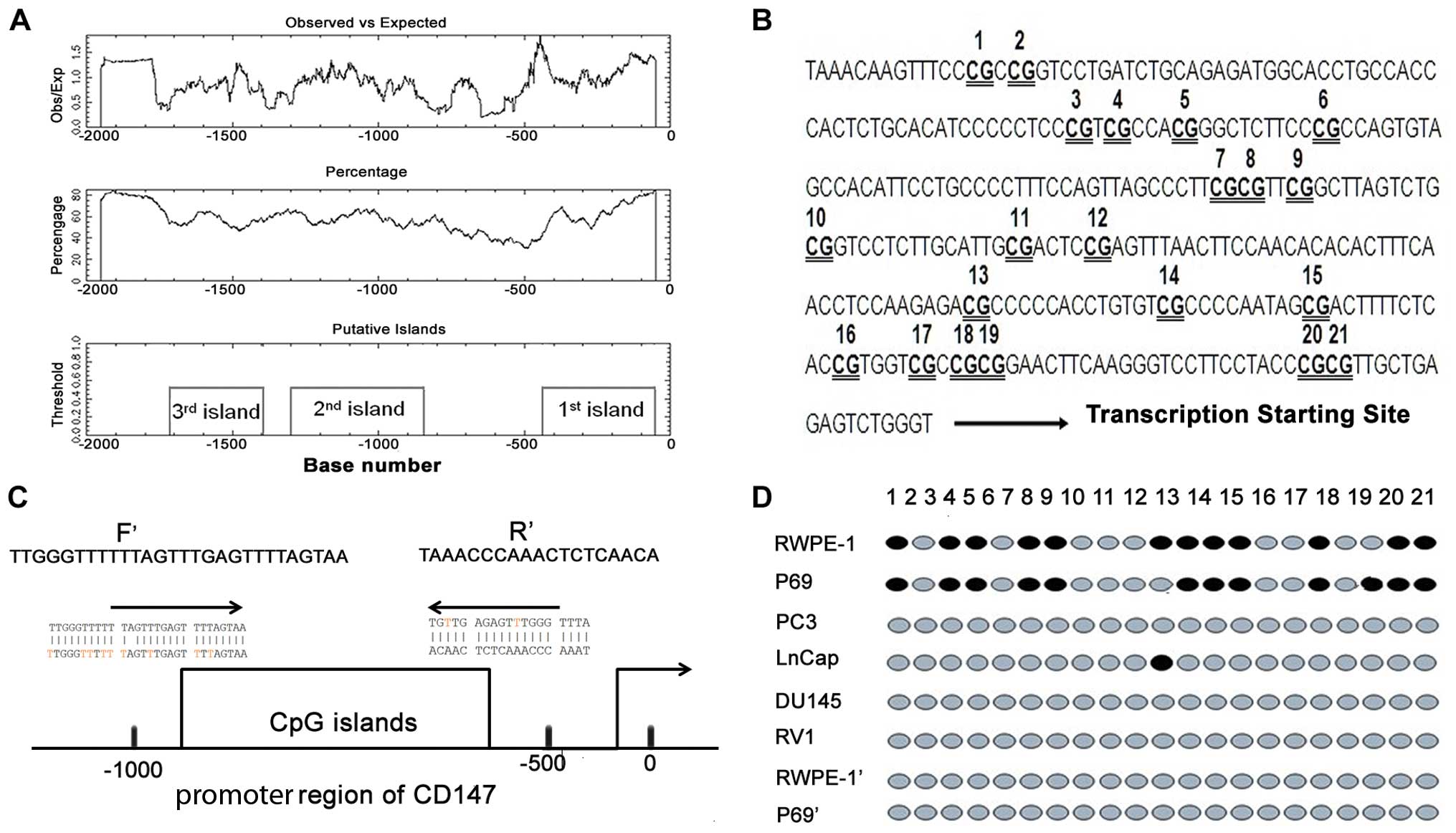
To evaluate whether CD147 promoter region contains
an area rich with CpG dinucleotides, we used the EMBOSS CpG plot
(EMBL-EBI: http://www.ebi.ac.uk/Tools/emboss/cpgplot/index.html).
The ‘cpgplot’ identifies the CpG islands in one or more nucleotide
sequences. The ratio of the observed to the expected number of GC
dinucleotide patterns was calculated over a sequence region. The
calculated ratios were plotted graphically, together with the
regions matching the definition of the program ‘CpG island’ (a CG
dinucleotide rich area). An area was considered a CpG island if an
average of 10 nucleotide sequences which were at least 200 bases,
contained over 50% G and C nucleotides, and the calculated
observed/expected ratio was over 0.6. We analyzed the DNA sequence
of the CD147 promoter region (NG_007468.1) using the EMBOSS tool,
the result of evaluation for the CpG islands is shown in Fig. 2A, where 3 CpG islands are predicted.
Among them, the sequence of -500 has been reported by Sharma et
al (5). According to the
results of Liang et al (26), a fragment -1024 to -59 bp upstream
of the CD147 coding region was sufficient to promote transcription.
In the present study, we chose 1–2 CpG islands located in this
fragment to further study. To investigate the correlation between
DNA methylation status and CD147 expression level, promoter
methylation was analyzed for CD147 by genomic bisulfite sequencing
in 4 PCa (PC3, Lncap, DU145 and RV1) and 2 non-tumorigenic benign
human prostatic epithelial cell lines (RWPE-1 and P69). However,
our data showed that no methylated CpG spot in typical 46 CpG sites
was detected in this region of the 1st CpG island in either PCa or
non-tumorigenic benign human prostatic epithelial cells. As shown
in Fig. 2B and C, a total of 21 CpG
sites in the 2nd CpG island were assessed by PCR and sequencing.
The non-tumorigenic benign human prostatic epithelial cell lines
exhibited a denser methylation pattern at the CD147 promoter region
than the corresponding PCa cell lines (Fig. 2D), suggesting that the
hypomethylation of the CD147 promoter region may contribute to the
increased CD147 expression in the PCa cells. We also found that the
non-tumorigenic benign human prostatic epithelial cell lines
treated with 5-aza-2′-deoxycytidine may exhibit the same
methylation pattern as the PCa cell lines (Fig. 2D).
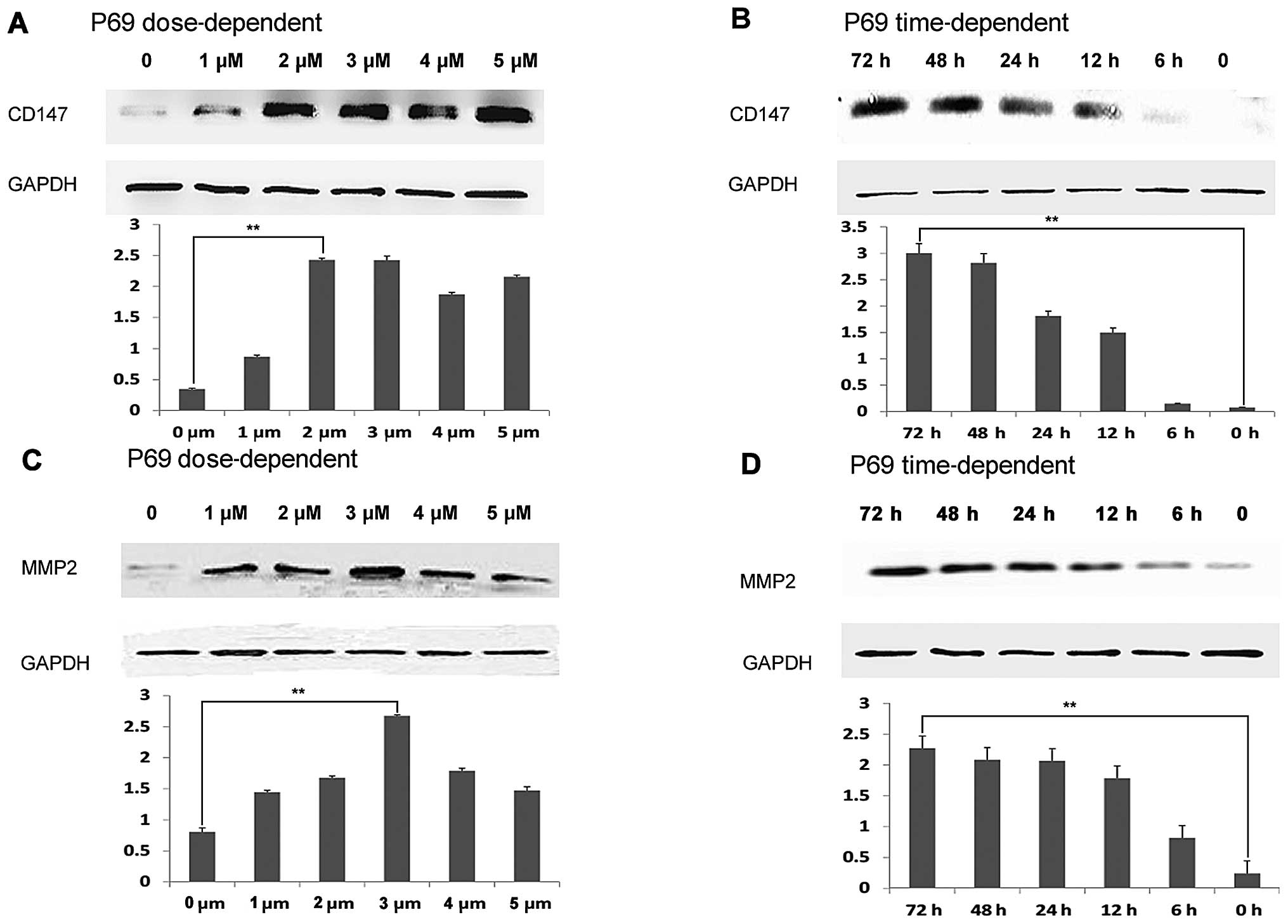
Re-expression of CD147 induced by
5-aza-2′-deoxycytidine treatment in non-tumorigenic benign human
prostatic epithelial cell line
To examine whether the CD147 expression in the
non-tumorigenic benign human prostatic epithelial cell line P69
could be restored by the treatment with DNA methylation inhibitors,
we treated the P69 cells with 5-aza-2′-deoxycytidine and then
analyzed for CD147 protein expression by western blot analysis. As
shown in Fig. 3A and B, the
treatment of 5-aza-2′-deoxycytidine is able to induce CD147
expression in P69 cells in a dose- and time-dependent manner. Since
MMP-2 has been demonstrated to be one of the most important
downstream targets of CD147, we further examined whether MMP-2
expression in P69 cells could be activated by the treatment with
DNA methylation inhibitors. As shown in Fig. 3C and D, the treatment of
5-aza-2′-deoxycytidine is also able to activate MMP-2 expression in
P69 cells in a dose- and time-dependent manner.
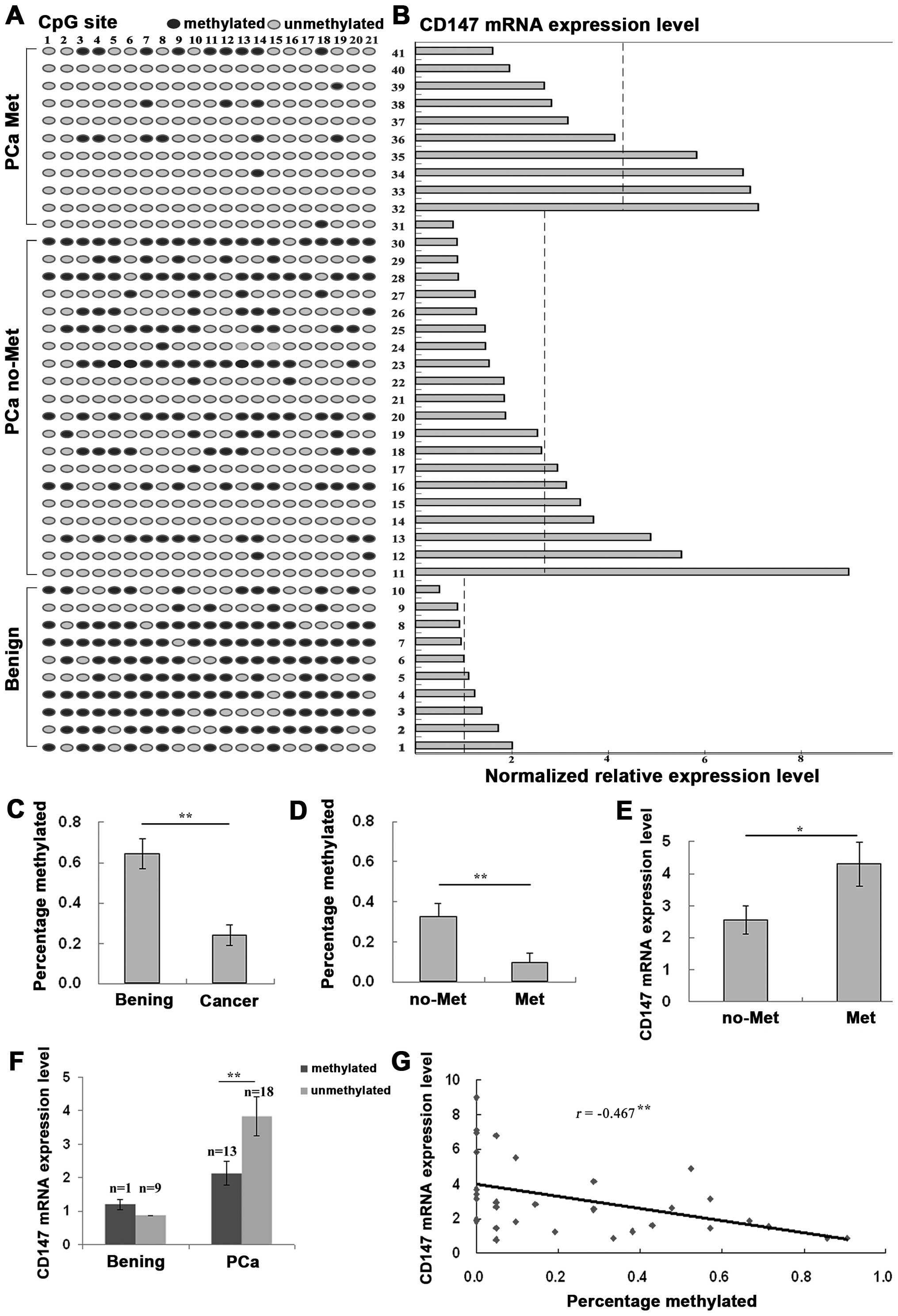
Hypomethylation affects CD147 expression
in clinical PCa tissues and is related to the metastasis of
cancer
To investigate the epigenetic mechanism responsible
for CD147 transcription in human PCa, the DNA methylation status of
the CD147 promoter region (2nd CpG islands) was determined in PCa
and corresponding non-cancerous prostate tissues by sodium
bisulfite genomic sequencing and the expression levels of CD147
mRNA were also detected by real-time quantitative RT-PCR using the
same clinical samples. As shown in Fig.
4A and C, the CpG sites in the island exhibited significant
hypomethylation in PCa tissues compared to corresponding benign
prostate tissues (ratio of methylation PCa vs. benign: 0.24 vs.
0.64, P<0.01). Importantly, CpG sites in the island exhibited
significant hypomethylation in metastatic PCa tissues compared to
non-metastatic PCa tissues (ratio of methylation metastatic PCa vs.
non-metastatic PCa: 0.10 vs. 0.32, P=0.009, Fig. 4D). Similarly, the expression levels
of CD147 mRNA in PCa tissues were significantly higher than those
in corresponding benign prostate tissues (PCa vs. benign: 3.12 vs.
1.18, P<0.01, Fig. 4B) and the
expression levels of CD147 mRNA in metastatic PCa tissues were also
significantly higher than those in non-metastatic PCa tissues (PCa
vs. benign: 4.30 vs. 2.55, P=0.046, Fig. 4E). In addition, methylation
intensity was significantly correlated inversely with CD147 mRNA
levels in PCa tissues (Fig. 4B).
CD147 mRNA levels in PCa tissues with DNA hypomethylation were
significantly higher than those with DNA methylation (methylated
PCa vs. unmethylated PCa: 2.12 vs. 3.83, P=0.017), but no
statistical difference was found between mythylated and
hypomethylated benign prostate tissues (P>0.05, Fig. 4F). There was a significant negative
correlation between CD147 mRNA levels and the number of methylated
sites in PCa tissues (Pearson correlation r=−0.467, P=0.008,
Fig. 4G).
Discussion
Many studies have emphasized that epigenetic
alterations play an important role in the initiation of various
human cancers. Aberrant expression of cancer-related genes may
benefit the expansion of the cells in the early abnormal cloning
and ‘addict’ cancer cells to the subsequent genetic and epigenetic
alterations that further promote tumor progression (27). According to our previous data, as
well as that of other research groups, CD147 functions as an
oncogene and is a good target for diagnosis, prediction for disease
progression and therapy of PCa (20–25).
In the present study, we observed the overexpression of CD147 mRNA
and protein in several PCa cell lines, consistent with our previous
studies on PCa tissues (21,24,25).
Notably, we found that overexpression was associated with
hypomethylation of CD147 promoter in PCa cell lines. In addition,
re-expression of CD147 and its downstream target MMP-2 are able to
be induced by demethylation agent 5-aza-2′-deoxycytidine in
low-expressing non-tumorigenic benign human prostatic epithelial
cell lines. Furthermore, PCa tissues displayed decreased DNA
methylation in the promoter region of CD147 compared to
corresponding non-cancerous prostate tissues, and methylation
intensity correlated inversely with CD147 mRNA levels. There was a
significantly negative correlation between CD147 mRNA levels and
the number of methylated sites in the PCa tissues. Although still
leaving much to investigate, to our knowledge, this is the first
attempt to explore a possible mechanism leading to the abnormal
CD147 expression levels in PCa.
Despite the comprehensive investigation of CD147
expression by previous studies (20–25),
the regulatory mechanisms underlying aberrant CD147 expression in
various human cancers are not yet fully elucidated. Since the
aberrant methylation of promoter CpG islands has been widely
considered as a kind of regulatory mechanism of silencing tumor
suppressors or activating oncogenes, it is reasonable to assume the
possible role of aberrant methylation on CD147 expression levels.
To validate this hypothesis, we firstly investigated the
methylation status of CD147 promoter in PCa and non-tumorigenic
benign human prostatic epithelial cell lines. The results showed a
significantly lower level of methylation status of CD147 promoter
in the PCa cells when compared with the benign controls. There was
a correlation between the promoter hypomethylation and the CD147
overexpression, suggesting that the promoter demethylation
increased the expression of CD147 in the PCa cells, which was
further validated by our subsequent experiments in cell lines P69
and PC3 treated with the demethylating agent. Then, the present
study also showed that human PCa tissues displayed a relatively
hypomethylated CD147 promoter, compared to corresponding
non-cancerous prostate tissues. The above findings confirm our
initial hypothesis that DNA methylation may play a crucial role in
regulating the expression of CD147 in PCa.
In conclusion, our data offer convincing evidence
for the first time that DNA promoter hypomethylation of CD147 may
be one of the regulatory mechanisms involved in the cancer-related
overexpression of CD147 and may play a crucial role in the
tumorigenesis of human PCa.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (nos. 81200550 and 81170699),
the Medical Research Fund of Guangdong Province (no. A2012489) and
the Science and Technology Project of Bureau of Health in Guangzhou
Municipality (no. 20121A011004).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Molitierno J, Evans A, Mohler JL, Wallen
E, Moore D and Pruthi RS: Characterization of biochemical
recurrence after radical prostatectomy. Urol Int. 77:130–134. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Klotz L: Hormone therapy for patients with
prostate carcinoma. Cancer. 88(Suppl 12): 3009–3014. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Schoenborn JR, Nelson P and Fang M:
Genomic profiling defines subtypes of prostate cancer with the
potential for therapeutic stratification. Clin Cancer Res.
19:4058–4066. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sharma S and Watabe K: Biomarkers and
mechanisms associated with recurrent prostate cancer. Front Biosci
(Landmark Ed). 19:339–351. 2014. View
Article : Google Scholar
|
|
6
|
Devaney JM, Wang S, Funda S, Long J,
Taghipour DJ, Tbaishat R, Furbert-Harris P, Ittmann M and
Kwabi-Addo B: Identification of novel DNA-methylated genes that
correlate with human prostate cancer and high-grade prostatic
intraepithelial neoplasia. Prostate Cancer Prostatic Dis.
16:292–300. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yang B, Bhusari S, Kueck J, Weeratunga P,
Wagner J, Leverson G, Huang W and Jarrard DF: Methylation profiling
defines an extensive field defect in histologically normal prostate
tissues associated with prostate cancer. Neoplasia. 15:399–408.
2013.PubMed/NCBI
|
|
8
|
Lin PC, Giannopoulou EG, Park K, Mosquera
JM, Sboner A, Tewari AK, Garraway LA, Beltran H, Rubin MA and
Elemento O: Epigenomic alterations in localized and advanced
prostate cancer. Neoplasia. 15:373–383. 2013.PubMed/NCBI
|
|
9
|
Aryee MJ, Liu W, Engelmann JC, Nuhn P,
Gurel M, Haffner MC, Esopi D, Irizarry RA, Getzenberg RH, Nelson
WG, et al: DNA methylation alterations exhibit intraindividual
stability and interindividual heterogeneity in prostate cancer
metastases. Sci Transl Med. 5:169ra102013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Majumdar S, Buckles E, Estrada J and
Koochekpour S: Aberrant DNA methylation and prostate cancer. Curr
Genomics. 12:486–505. 2011. View Article : Google Scholar :
|
|
11
|
Dimitriadis E, Kalogeropoulos T, Velaeti
S, Sotiriou S, Vassiliou E, Fasoulis L, Klapsas V, Synesiou M,
Apostolaki A, Trangas T, et al: Study of genetic and epigenetic
alterations in urine samples as diagnostic markers for prostate
cancer. Anticancer Res. 33:191–197. 2013.
|
|
12
|
Yu YP, Ding Y, Chen R, Liao SG, Ren BG,
Michalopoulos A, Michalopoulos G, Nelson J, Tseng GC and Luo JH:
Whole-genome methylation sequencing reveals distinct impact of
differential methylations on gene transcription in prostate cancer.
Am J Pathol. 183:1960–1970. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Haldrup C, Mundbjerg K, Vestergaard EM,
Lamy P, Wild P, Schulz WA, Arsov C, Visakorpi T, Borre M, Høyer S,
et al: DNA methylation signatures for prediction of biochemical
recurrence after radical prostatectomy of clinically localized
prostate cancer. J Clin Oncol. 31:3250–3258. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chao C, Chi M, Preciado M and Black MH:
Methylation markers for prostate cancer prognosis: A systematic
review. Cancer Causes Control. 24:1615–1641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dietrich D, Hasinger O, Bañez LL, Sun L,
van Leenders GJ, Wheeler TM, Bangma CH, Wernert N, Perner S,
Freedland SJ, et al: Development and clinical validation of a
real-time PCR assay for PITX2 DNA methylation to predict
prostate-specific antigen recurrence in prostate cancer patients
following radical prostatectomy. J Mol Diagn. 15:270–279. 2013.
View Article : Google Scholar
|
|
16
|
Kron K, Trudel D, Pethe V, Briollais L,
Fleshner N, van der Kwast T and Bapat B: Altered DNA methylation
landscapes of polycomb-repressed loci are associated with prostate
cancer progression and ERG oncogene expression in prostate cancer.
Clin Cancer Res. 19:3450–3461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vasiljević N, Ahmad AS, Beesley C, Thorat
MA, Fisher G, Berney DM, Møller H, Yu Y, Lu YJ, Cuzick J, et al:
Association between DNA methylation of HSPB1 and death in low
Gleason score prostate cancer. Prostate Cancer Prostatic Dis.
16:35–40. 2013. View Article : Google Scholar
|
|
18
|
Gabison EE, Mourah S, Steinfels E, Yan L,
Hoang-Xuan T, Watsky MA, De Wever B, Calvo F, Mauviel A and Menashi
S: Differential expression of extracellular matrix
metalloproteinase inducer (CD147) in normal and ulcerated corneas:
role in epitheliostromal interactions and matrix metalloproteinase
induction. Am J Pathol. 166:209–219. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun J and Hemler ME: Regulation of MMP-1
and MMP-2 production through CD147/extracellular matrix
metalloproteinase inducer interactions. Cancer Res. 61:2276–2281.
2001.PubMed/NCBI
|
|
20
|
Wang L, Wu G, Yu L, Yuan J, Fang F, Zhai
Z, Wang F and Wang H: Inhibition of CD147 expression reduces tumor
cell invasion in human prostate cancer cell line via RNA
interference. Cancer Biol Ther. 5:608–614. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhong WD, Han ZD, He HC, Bi XC, Dai QS,
Zhu G, Ye YK, Liang YX, Qin WJ, Zhang Z, et al: CD147, MMP-1, MMP-2
and MMP-9 protein expression as significant prognostic factors in
human prostate cancer. Oncology. 75:230–236. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Madigan MC, Kingsley EA, Cozzi PJ,
Delprado WJ, Russell PJ and Li Y: The role of extracellular matrix
metalloproteinase inducer protein in prostate cancer progression.
Cancer Immunol Immunother. 57:1367–1379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hao JL, Cozzi PJ, Khatri A, Power CA and
Li Y: CD147/EMMPRIN and CD44 are potential therapeutic targets for
metastatic prostate cancer. Curr Cancer Drug Targets. 10:287–306.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Han ZD, Bi XC, Qin WJ, He HC, Dai QS, Zou
J, Ye YK, Liang YX, Zeng GH, Chen ZN, et al: CD147 expression
indicates unfavourable prognosis in prostate cancer. Pathol Oncol
Res. 15:369–374. 2009. View Article : Google Scholar
|
|
25
|
Zhong WD, Liang YX, Lin SX, Li L, He HC,
Bi XC, Han ZD, Dai QS, Ye YK, Chen QB, et al: Expression of CD147
is associated with prostate cancer progression. Int J Cancer.
130:300–308. 2012. View Article : Google Scholar
|
|
26
|
Liang L, Major T and Bocan T:
Characterization of the promoter of human extracellular matrix
metalloproteinase inducer (EMMPRIN). Gene. 282:75–86. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Truong M, Yang B, Livermore A, Wagner J,
Weeratunga P, Huang W, Dhir R, Nelson J, Lin DW and Jarrard DF:
Using the epigenetic field defect to detect prostate cancer in
biopsy negative patients. J Urol. 189:2335–2341. 2013. View Article : Google Scholar
|