Introduction
Lung cancer is the leading cause of cancer-related
deaths, and non-small cell lung cancer (NSCLC) accounts for most
cases of lung cancer (1). It has
been estimated that NSCLC may occupy the leading position for the
next decade (2). Obviously, early
treatment is extremely important as the 5-year survival rate is
~80% in stage I/II but declines significantly to about 14% in stage
III/IV (1). In fact, only 25% of
NSCLC patients are treated in early stages and this leads to a high
mortality rate (2). In spite of the
development in the fields of oncology and surgery, the prognosis of
lung cancer remains unsatisfactory, and lung cancer often reoccurs
within one year (3).
microRNAs (miRNAs) are characterized by endogenous,
single-stranded, non-coding RNA consisting of fewer than 22
nucleotides (4). As known, miRNAs
play a role in negatively regulating targeted gene translation
post-transcriptionally and specifically without changing DNA
sequences by pairing with the 3′-untranslated region of the target
mRNAs, leading to translation inhibition or mRNA degradation
(5,6), consequently controlling cell
proliferation and apoptosis, and stem cell differentiation
(4,7,8).
Moreover, accumulating evidence has shown that miRNAs act as
oncogenes or inhibit cancer development and progression according
to the roles of their target genes (9). To date, miRNAs are believed to
participate in the regulation of the expression of more than half
of the human genes (7,10,11)
and a considerable number have been found to be deregulated in
cancer cells compared with normal cells (12,13).
In other words, miRNAs play a key role in regulating tumorigenesis,
which is characterized by its aberrant expression (14,15).
There are relevant studies concerning both miRNAs
and NSCLC. For example, it has been reported that miR-449a
expression is decreased in NSCLC, which could promote cell
proliferation and inhibit apoptosis (16). Similarly, clinical data analysis has
shown that decreased miR-224 and increased miRNA-96 expression in
NSCLC tissues are closely related to clinicopathologic stage and a
poor prognosis (17,18). Furthermore, other miRNAs, including
miR-30b/c, miR-99a, miR-493 and miR-27b, have all exhibited
inhibitory effects on both the proliferation and invasion of NSCLC
cells (19–22). These previous studies and results
indicate that miRNAs may act as potential therapeutic targets of
human NSCLC.
Another cancer-related miRNA is microRNA-370
(miR-370). As reported, Lo et al demonstrated that miR-370
was overexpressed in gastric carcinoma, which indicated a more
advanced nodal metastasis and a higher clinical stage (23). Notably, miR-370 demostrated opposite
effects in laryngeal squamous cell carcinoma. The results reported
by Yungang et al showed that miR-370 was downregulated in
human laryngeal squamous cell carcinoma tissues, suggesting that
miR-370 may function as a tumor suppressor (24). We focused on lung cancer research
and aimed to ascertain whether, how and in which manner miR-370
functions in NSCLC. However, studies on the expression and
functions of miR-370 in NSCLC are sparse. Therefore, we compared
the expression of miR-370 in NSCLC and normal tissues. Next, with
the aid of two cell models, we evaluated the effects of high
expression of miR-370 on the proliferation and apoptosis of NSCLC
cells. Furthermore, we assessed the tumor formation ability in
vivo under conditions of high and low expression of
miR-370.
The tumor necrosis factor receptor-associated factor
(TRAF) family consists of 7 members (TRAF1-7) (25), which have been demonstrated to
participate in human diseases, including tumors and cancer, immune
system disorders and neurodiseases (26,27).
Therefore, targeting these molecules may contribute to the
treatment of TRAF-mediated human diseases. Among the seven TRAF
family members, TRAF4 is the first member that has been found to be
overexpressed in cancer and is now considered as an oncogene
(28,29). TRAF4 has been reported to promote
breast cancer progression (30);
however, the underlying mechanism of TRAF4 in NSCLC has not been
fully investigated or determined.
Based on the present experiments, we confirmed a
close connection between TRAF4 and miR-370 in NSCLC cell lines,
demonstrating that TRAF4 may be a negative downstream effector of
miR-370. In contrast, TRAF4 overexpression had a promotive effect
on NSCLC cells. In conclusion, our results showed a growth
inhibitory effect of miR-370 on NSCLC and provides an experimental
basis for its potential therapeutic use.
Materials and methods
Sample collection and animals
All of the samples were collected from the First
Affiliated Hospital of Xi’an Jiaotong University. We obtained
informed consents from all of the participants following the
approval of the Ethics Committee of Xi’an Jiaotong University. The
samples were collected before any other therapeutic methods were
employed on the patients. In total, 25 early-stage NSCLC and 23
adjacent non-tumor samples were collected. The samples were
immediately stored in liquid nitrogen for further analysis. Female
BALB/c mice were purchased from the Animal Experimental Center of
the College of Medicine, Xi’an Jiaotong University and raised in
sterilized mouse cages with free access to water and food. All the
experiments on mice complied with the requirements of the
Institutional Animal Care and Use Committee of Xi’an Jiaotong
University.
Cell culture
Normal human lung epithelium cells (BEAS-2B) and two
human NSCLC cell lines (A549 and H358) were purchased from the
American Type Culture Collection (ATCC; Manassas, VA, USA). The
cells were cultured in RPMI-1640 medium (Invitrogen, Carlsbad, CA,
USA) supplemented with 10% fetal bovine serum (FBS; HyClone, Logan,
UT, USA), 100 U/ml penicillin and 100 μg/ml streptomycin in
a humidified atmosphere at 37°C with 5% CO2.
Lentivirus construction and
transfection
The lentivirus gene transfer vectors of hsa-miR-370
and TRAF4 were constructed by GenePharma Co., Ltd. (Shanghai,
China). Scramble lentivirus vectors served as negative controls.
A549 and H358 cells were plated in 6-well plates (5×104
cells/well) in preparation for use. The lentiviruses were diluted
in 0.2 ml complete medium containing Polybrene (8 mg/ml) and added
to the cells for 12 h of incubation at 37°C, followed by incubation
in 0.3 ml of freshly prepared Polybrene-DMEM for another 24 h,
which was replaced with fresh DMEM and the cells were cultured for
another 3 days. Subsequently, miR-370 and TRAF4 expression was
measured by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR), respectively.
RT-qPCR
Total RNA was isolated from the samples and cells
using TRIzol reagent (Invitrogen). The reverse transcription
reaction was performed with 5 μg of RNA using the
Transcriptor First Strand cDNA Synthesis kit (Roche, Basel,
Switzerland) following the manufacturer’s instructions. RT-qPCR was
conducted using FastStart Universal Real-Time PCR Master Mix
(Roche) on the PikoReal Real-Time PCR Detection system (Thermo
Fisher Scientific, Waltham, MA, USA) with cycling conditions of
95°C for 10 min, followed by 45 cycles of 95°C for 15 sec and 60°C
for 60 sec. β-actin served as an internal control for TRAF4 and U6
small nuclear RNA (snRNA) for miR-370. The primer sequences were as
follows: miR-370 forward, 5′-GTA GGC GAT ATC GTC TGC TAC-3′ and
reverse, 5′-TAG AAG GTA GCA CCC GAT G-3′; U6 forward, 5′-GTA GAT
ACT GCA GTA CG-3′ and reverse, 5′-ATC GCA TGA CGT ACC TGA GC-3′;
TRAF4 forward, 5′-CTG GAC ATT TGA TCA TGC AG-3′ and reverse, 5′-ATT
TGA TCC AAT AGT TGC TCG A-3′; and β-actin forward, 5′-CCA AGT TGC
ATT TGA CCT GG-3′ and reverse, 5′-GAC CTG CGA GAT GCC TCA CC-3′.
The threshold cycle (Ct) value was recorded. Each sample was
measured in triplicate, and the relative expression of miR-370 to
U6 and TRAF4 to β-actin was calculated using the 2−ΔΔCt
method.
Western blotting
Proteins were extracted using RIPA buffer
supplemented with 1 mM phenylmethanesulfonyl fluoride (PMSF) (both
from Sigma-Aldrich, St. Louis, MO, USA). The protein concentration
was measured by the BCA protein assay kit (CWBio, Beijing, China).
A total of 20 μg of protein was isolated using 12% sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to a polyvinylidene fluoride (PVDF) membrane, followed
by the removal of non-specific binding in the membrane with 2.5%
skimmed milk. Subsequently, the membrane was incubated with primary
antibodies for 4 h at 37°C, including mouse anti-TRAF4, -caspase-3
and -Bax (all from Santa Cruz Biotechnology, Santa Cruz, CA, USA),
followed by washing 3 times with Tris-buffered saline and Tween-20
(TBST) and incubation with a secondary antibody (Santa Cruz
Biotechnology) conjugated with horseradish peroxidase (HRP) for 2 h
at room temperature. Finally, the protein band was detected by an
enhanced chemiluminescence (ECL) detection system (Amersham, Little
Chalfont, UK).
MTT and BrdU assays
For the MTT assay, A549 and H358 cells were seeded
in 96-well plates and reached a confluency of 80%. Subsequently,
the cells were transfected with the lentivirus gene transfer
vectors of hsa-miR-370 and TRAF4, as previously described. Next, 20
μl of MTT was added to each well and incubated for 4 h at
37°C with 5% CO2. Dimethyl sulfoxide (DMSO;
Sigma-Aldrich) was added to dissolve the formazan crystals and the
absorbance was measured at 490 nm.
For the BrdU assay, when the A549 and H358 cells
grew to a confluency of 50% in 96-well plates, the medium was
replaced with fresh BrdU medium and the cells were incubated in 5%
CO2 at 37°C for 1 h. After being washed with
phosphate-buffered saline (PBS) and fixed with 70% ethanol, the
cell slides were incubated with the primary human anti-BrdU
antibody and secondary FITC-conjugated-anti-human antibody. The
cells were observed and counted in randomly selected fields under a
fluorescence microscope (Olympus). Data are presented as
percentages of BrdU-positive cells vs. total cells.
Tumor formation assay
A549 and H358 cells (2×106 cells)
transfected with the lentivirus vectors overexpressing miR-370 and
scramble lentivirus vectors were diluted in 200 μl of PBS
followed by injection subcutaneously into the right flank of BALB/c
mice. The tumor volume was measured every 2 days with a Vernier
caliper and presented as Volume = length × width2 × π/6.
Five weeks later, the mice were sacrificed by subcutaneous
injection with excessive sodium pentobarbital (40 mg/kg), and the
tumors were removed and measured.
Dual-luciferase reporter assay
The possible miR-370 binding sites in the TRAF4 gene
3′-untranslated regions (3′-UTR) were predicted via microRNA.org.
The cDNA fragments containing the predicted miR-370 binding sites
in TRAF4 were amplified and subcloned into pmirGLO Dual-Luciferase
miRNA Target Expression Vector (named pmirGLO-TRAF4; Promega,
Madison, WI, USA). As a control, the pmirGLO-mutTRAF4 plasmids were
also constructed using cDNA fragments containing corresponding
mutated nucleotides for miR-370. The pre-miR-370 and
pre-miR-scramble plasmids were constructed and amplified in
preparation for use. Subsequently, HEK-293T cells, cultured in
6-well plates with a confluency of 80%, were co-transfected with
100 ng of pmirGLO-TRAF4 or pmirGLO-mutTRAF4 vectors in the presence
of 40 nM pre-miR-370 or pre-miR-scramble using Lipofectamine
transfection reagent (Invitrogen) and incubated for 48 h. The cells
were harvested and the luciferase activities were measured using
the Dual-Luciferase Reporter Assay kit (Promega) according to the
manufacturer’s instructions.
Statistical analysis
Data are presented as mean ± standard deviation
(SD). The differences were analyzed by the Student’s t-test or
one-way ANOVA analysis. A p<0.05 was considered to be
statistically significant and a p<0.01 extremely
significant.
Results
miR-370 expression is downregulated in
NSCLC samples and cells
To investigate the expression of miR-370 in the
NSCLC patient tissues and cell lines and compare its expression
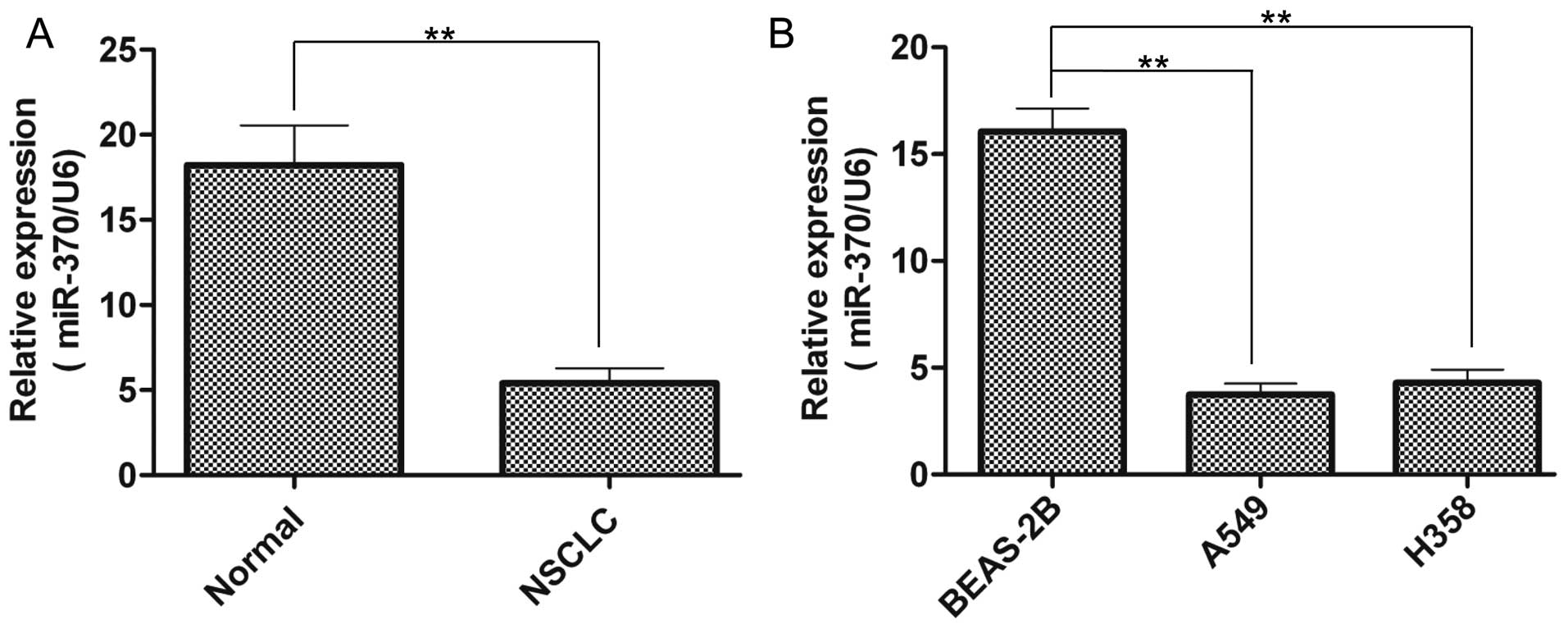
with normal samples or cells, RT-qPCR was performed. As shown in
Fig. 1A, miR-370 expression was
significantly inhibited in the NSCLC tissues compared with that in
the normal tissues. Furthermore, miR-370 expression was
significantly downregulated in the NSCLC cell lines (Fig. 1B), including A549 and H358 cells, in
contrast to the normal lung epithelium BEAS-2B cells. These results
suggest that miR-370 may play a role in the tumorigenesis of
NSCLC.
miR-370 is overexpressed in A549 and H358
cells via lentivirus transfection
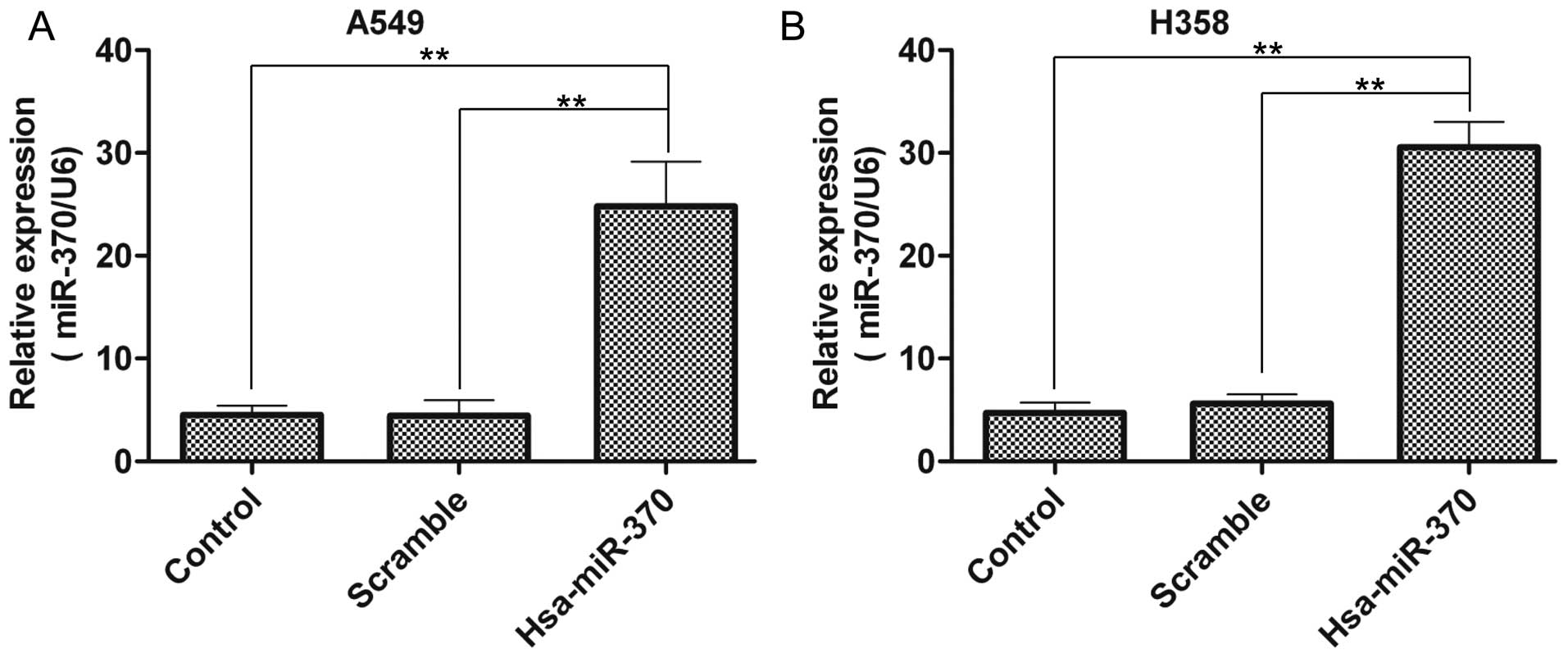
Next, miR-370 was overexpressed via lentivirus
transfection in the A549 and H358 cells. The miR-370 expression had
a 5.5-fold and 5.6-fold increase in the A549 cells, when compared
with the level in the control group and scramble vector group,
respectively (Fig. 2A). Similarly,
in the H358 cells, the data were 6.5-fold and 5.4-fold,
respectively (Fig. 2B). These
results confirmed the overexpression of miR-370 in the A549 and
H358 cells via lentivirus transfection.
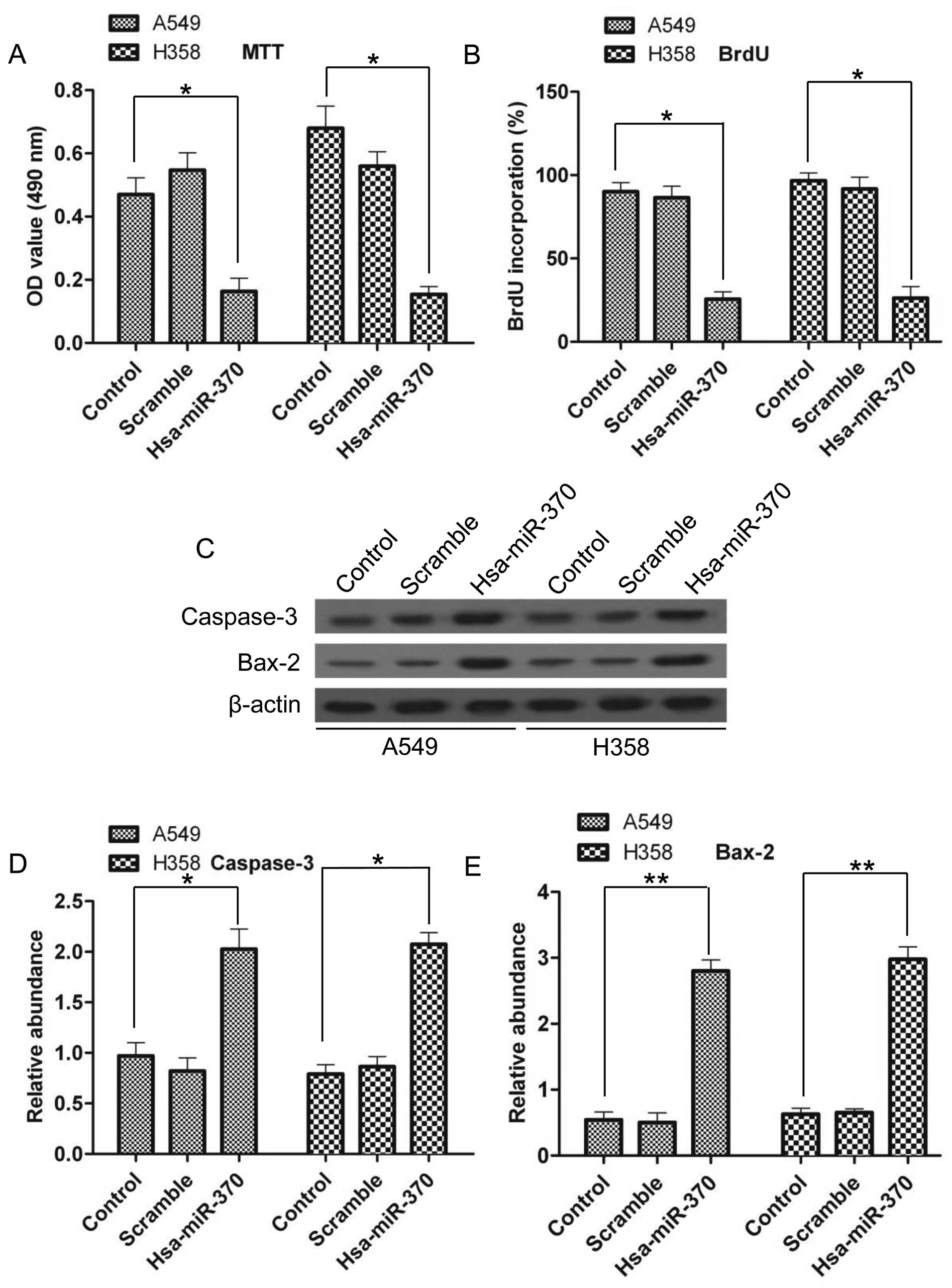
miR-370 overexpression inhibits
proliferation and induces the apoptosis of A549 and H358 cells
Since we know that miR-370 is a unique molecule in
NSCLC tissues, we next investigated its effects on NSCLC cell
lines. Cell proliferation was evaluated by MTT and BrdU assays. As
shown in Fig. 3A, there was a sharp
decrease in cell proliferation following miR-370 overexpression; in
A549 cells, cell proliferation had a 66% decrease and in H358
cells, cell proliferation had a 78% decrease compared with the
control group. The BrdU results also showed a reduced percentage of
BrdU incorporation after miR-370 overexpression (Fig. 3B), suggesting that miR-370 could
inhibit the proliferation of NSCLC cells. Cell apoptosis was
evaluated by western blotting. Upregulated expression of
apoptosis-related genes, including caspase-3 and Bax-2, at the
protein level was noted (Fig. 3C)
The densitometry analysis demonstrated a 2.1-fold and 2.6-fold
increase in caspase-3 in the A549 and H358 cells, respectively, and
a 5.2-fold and 4.7-fold increase in Bax-2 compared with the control
group, respectively (Fig. 3D and
E). These results confirmed that miR-370 had a positive effect
on NSCLC cell apoptosis.
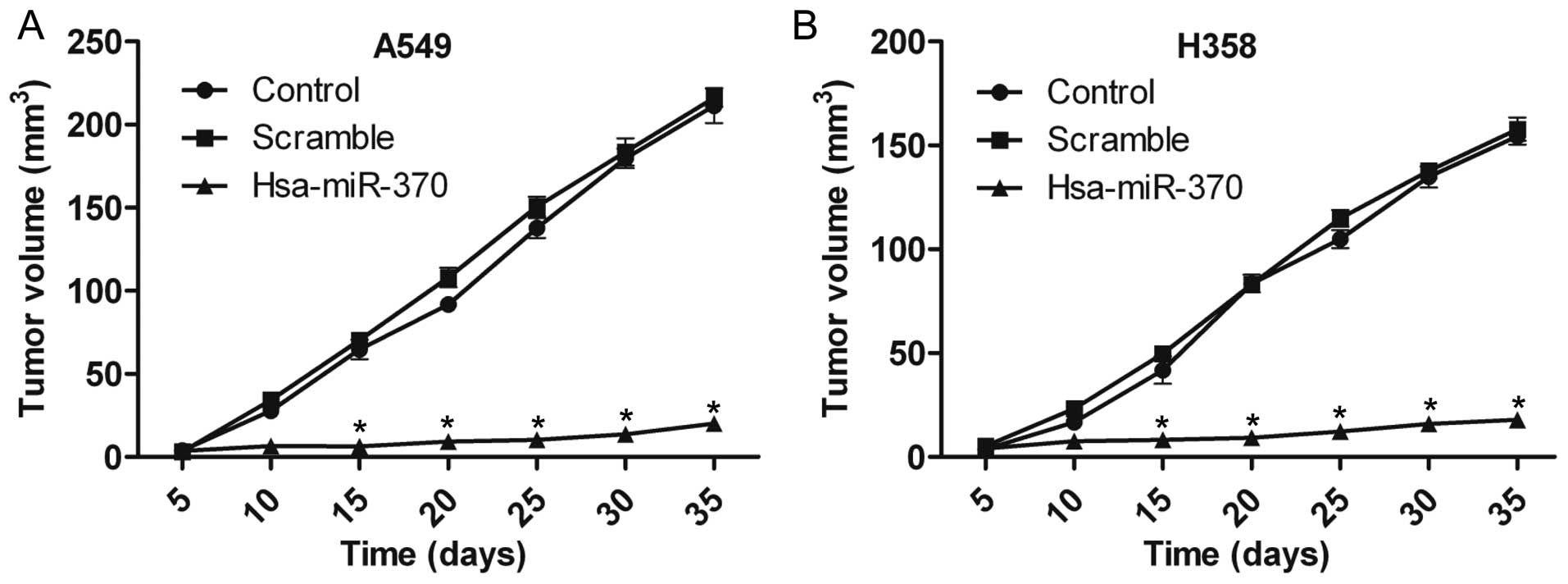
miR-370 overexpression inhibits tumor
formation in vivo
To evaluate the effects of miR-370 on NSCLC cell
growth in vivo, A549 and H358 cells transfected with
hsa-miR-370 vectors were injected into BALB/c mice. As shown in
Fig. 4A, tumor formation was
significantly inhibited 15 days after injection with the A549
hsa-miR-370 vector-transfected cells. The H358 hsa-miR-370
vector-transfected cells also had a lower decrease in tumor
formation ability (Fig. 4B). At 35
days post-injection, the tumor volume was 20.00±3.05 and 17.97±0.85
mm3 in the A549 and H358 cells transfected with
hsa-miR-370, respectively, compared with 216.23±5.51 and
157.80±5.55 mm3 in the scramble vector groups (Fig. 4A and B).
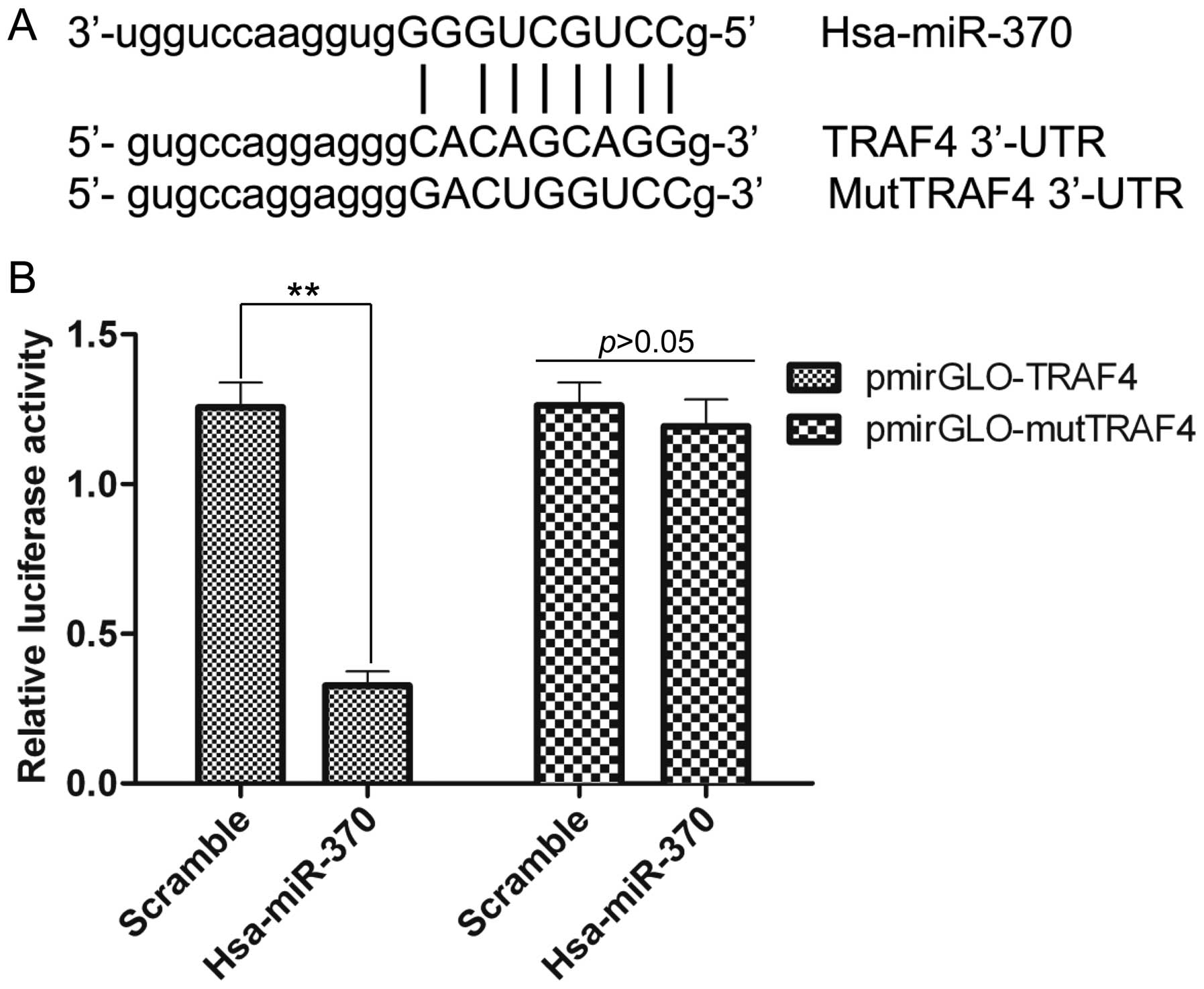
TRAF4 is a target gene of miR-370
To explore the potential mechanism of miR-370 in
regulating NSCLC, the predicted target genes of miR-370 were
screened by bioinformatic analysis. As shown in Fig. 5A, TRAF4, which is an important
oncogene, was shown to bind with hsa-miR-370 within the 3′-UTR, but
mut-TRAF4 had no binding sites with hsa-miR-370 within the 3′-UTR.
To confirm whether the predicted binding sites for miR-370 within
the 3′-UTR of TRAF4 were correct, dual-luciferase reporter assay
was performed. The results demonstrated that hsa-miR-370
significantly reduced the luciferase activity in the presence of
pmirGLO-TRAF4, whereas the luciferase activity was not affected by
hsa-miR-370 in the presence of pmirGLO-mutTRAF4 compared with the
scramble group (Fig. 5B). The above
results were consistent with the predicted analysis shown in
Fig. 5A. The data imply that
miR-370 directly binds to the 3′-UTR of TRAF4, in other words,
TRAF4 is a target gene of miR-370.
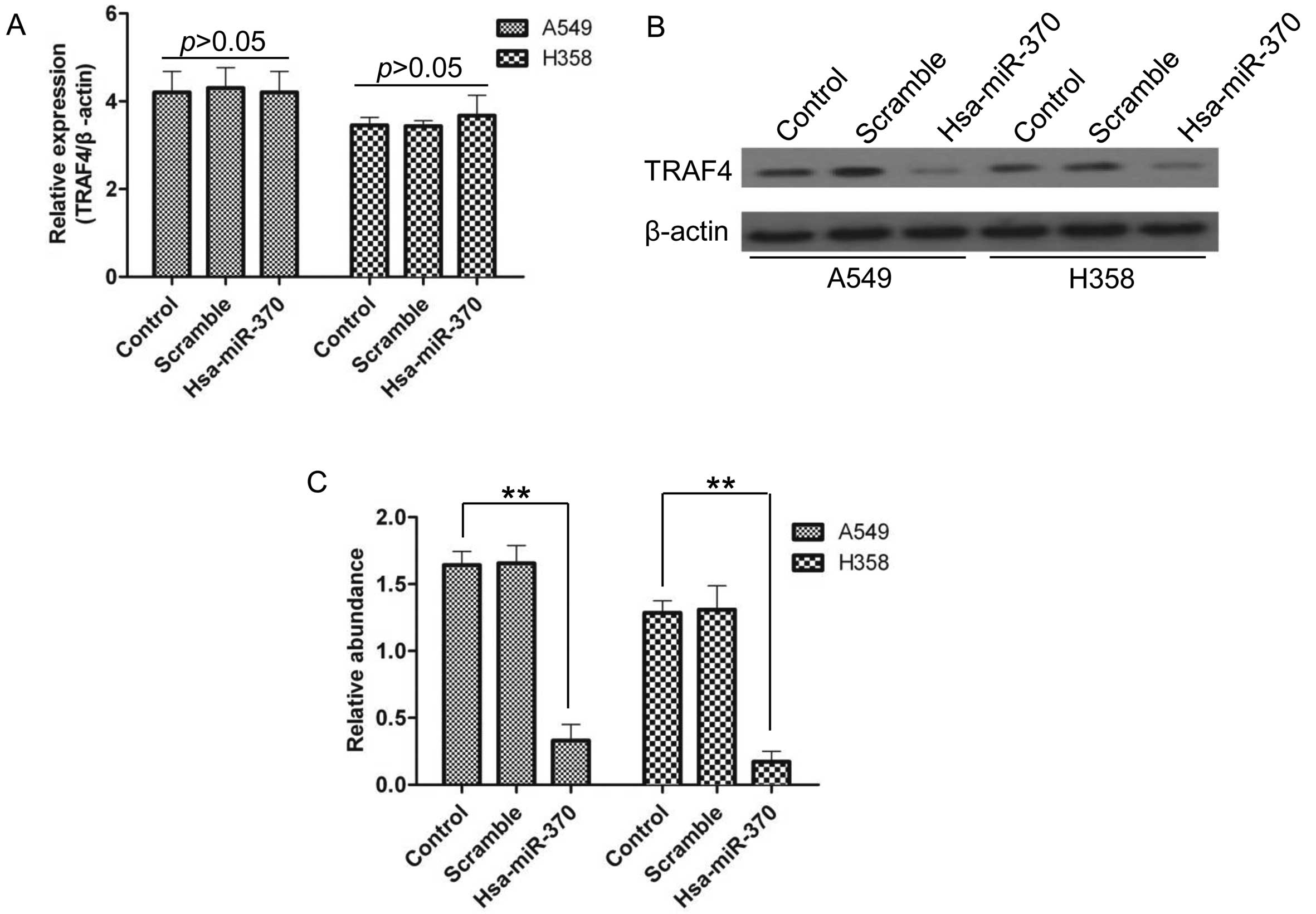
TRAF4 expression is regulated by miR-370
at the protein level but not at the mRNA level
To identify the effects of miR-370 on TRAF4
expression, RT-qPCR and western blotting were performed. Notably,
transfection of A549 and H358 cells with hsa-miR-370 had no effect
on the expression of TRAF4 at the mRNA level (Fig. 6A), whereas at the protein level
TRAF4 was significantly inhibited (Fig.
6B and C), suggesting that miR-370 prevented TRAF4 expression
at the post-transcriptional translation level.
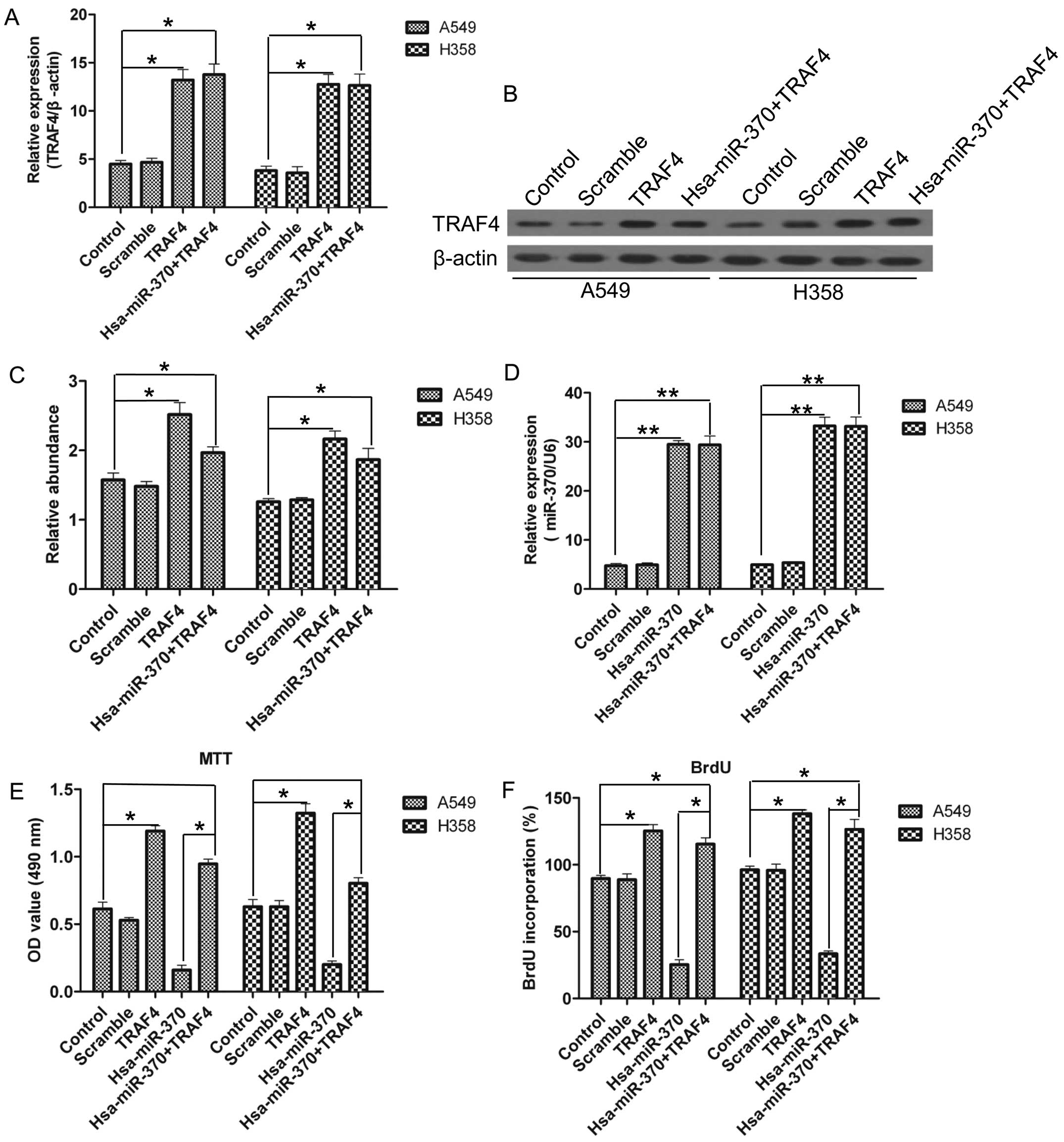
TRAF4 overexpression abolishes the
inhibitory growth effects on NSCLC cells induced by miR-370
TRAF4 expression was inhibited by miR-370, and we
aimed to ascertain whether miR-370 suppresses the proliferation of
NSCLC cells via downregulation of TRAF4. We co-transfected
hsa-miR-370 with TRAF4 vectors harboring no specific-binding
sequences of miR-370 in the 3′-UTR for both overexpression or
single overexpression in the A549 and H358 cells. As shown in
Fig. 7A–C, TRAF4 was overexpressed
at both the mRNA and the protein level, and miR-370 was
overexpressed at the mRNA level (Fig.
7D). Notably, TRAF4 transfection restored its expression at the
protein level in the presence of miR-370 overexpression (Fig. 7C). The MTT and BrdU results clearly
showed that the overexpression of TRAF4 promoted the proliferation
of NSCLC cells and counteracted the inhibition of the proliferation
of NSCLC cells induced by miR-370 over-expression (Fig. 7E and F).
Discussion
Lung cancer is the leading cause of cancer-related
deaths worldwide (31), 80% of
which are due to NSCLC (32). Great
progress have been achieved in elucidating the mechanisms and in
developing therapeutic methods for NSCLC in recent years, yet an
effective target is urgently needed for the treatment of NSCLC. In
the present study, we found that miR-370 expression was
downregulated in the NSCLC tissues and cells, which indicated that
miR-370 plays an important role in NSCLC. Upon this discovery,
further experiments were performed. miR-370 was overexpressed in
NSCLC cell lines, A549 and H358, via lentivirus transfection. The
results showed that miR-370 overexpression inhibited the
proliferation and promoted the apoptosis of NSCLC cells.
Furthermore, miR-370 overexpression suppressed the ability of tumor
formation of NSCLC cells in vivo. Next, TRAF4 was confirmed
to be a target gene of miR-370 in humans, and TRAF4 negatively
regulated the effects on NSCLC cells induced by miR-370, implying
that miR-370 possibly functions through TRAF4. In conclusion,
miR-370 may serve as an effective target for the treatment of
NSCLC.
miRNAs are a group of single-stranded, non-coding
molecules, participating in the regulation of their target genes at
the level of translation (4,33).
There are various studies on miRNAs and NSCLC. For example, Geng
et al identified five miRNAs as biomarkers for NSCLC
(34). Tafsiri et al
investigated miRNAs and their association with clinicopathological
features in NSCLC (35). miR-370
was also reported. Lo et al investigated miR-370 and its
novel target TGFb-RII in the progression of gastric carcinoma
(23). The authors demonstrated
that miR-370 was overexpressed in gastric carcinoma. However,
Yungang et al reported that miR-370 was downregulated in
human laryngeal squamous cell carcinoma by targeting FoxM1 as a
tumor suppressor (24). The results
showed that miR-370 may have various functions in different types
of cancers. Herein, we aimed to ascertain the functions of miR-370
in NSCLC and the potential mechanism. Our data showed that miR-370
suppressed the development of NSCLC. We used bioinformatic analysis
to predict the binding sequences of miR-370 and 3′-UTR of TRAF4.
Next, the dual-luciferase reporter assay was employed to verify our
hypothesis. The results showed that TRAF4 was indeed a target gene
of miR-370 in humans, which was consistent with our hypothesis.
As reported, TRAF4 was found to be overexpressed in
various cancer cells (28),
implying that TRAF4 overexpression is common in the majority of
cancer cells, whereas TRAF4 also has an antitumor effect (36,37).
Therefore, TRAF4 has different functions in different cancer cells,
dependent on the microenvironment and stimulation. Our data showed
that TRAF4 was highly expressed in NSCLC cells, and miR-370
overexpression significantly inhibited the expression of TRAF4.
These results suggest that TRAF4 serves as an oncogene in NSCLC.
Notably, inhibition of TRAF4 expression was only shown at the
protein level, namely, at the translational level, since miRNAs
function via binding with the 3′-UTR of mRNAs. It is worth noting
that the co-overexpression of miR-370 and TRAF4 demonstrated an
interesting phenomenon. TRAF4 overexpression diminished the
inhibitory effects on NSCLC cell proliferation caused by miR-370
overexpression, verifying their mutual crosstalk.
In conclusion, our research demonstrated that
miR-370 negatively regulates oncogene TRAF4 and inhibits the
proliferation of NSCLC in vitro and in vivo. Taken
together, our data provide clear evidence that miR-370 functions as
a tumor suppressor and may serve as an effective target for the
treatment of NSCLC. Therefore, further studies are necessary to
confirm the effects and investigate the underlying mechanisms of
miR-370 to provide a firm foundation for the use of miR-370 in the
treatment of NSCLC.
Acknowledgments
This study was supported by a grant from the
Scientific and Technological Research Project of Shaanxi Province
(2011K12-70).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Foss KM, Sima C, Ugolini D, Neri M, Allen
KE and Weiss GJ: miR-1254 and miR-574-5p: serum-based microRNA
biomarkers for early-stage non-small cell lung cancer. J Thorac
Oncol. 6:482–488. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Verdecchia A, Francisci S, Brenner H,
Gatta G, Micheli A, Mangone L and Kunkler I; EUROCARE-4 Working
Group: Recent cancer survival in Europe: a 2000–02 period analysis
of EUROCARE-4 data. Lancet Oncol. 8:784–796. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao G, Cai C, Yang T, Qiu X, Liao B, Li
W, Ji Z, Zhao J, Zhao H, Guo M, et al: MicroRNA-221 induces cell
survival and cisplatin resistance through PI3K/Akt pathway in human
osteosarcoma. PLoS One. 8:e539062013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mendell JT and Olson EN: MicroRNAs in
stress signaling and human disease. Cell. 148:1172–1187. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pillai RS: MicroRNA function: Multiple
mechanisms for a tiny RNA? RNA. 11:1753–1761. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shimono Y, Zabala M, Cho RW, Lobo N,
Dalerba P, Qian D, Diehn M, Liu H, Panula SP, Chiao E, et al:
Downregulation of miRNA-200c links breast cancer stem cells with
normal stem cells. Cell. 138:592–603. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sassen S, Miska EA and Caldas C: MicroRNA:
implications for cancer. Virchows Arch. 452:1–10. 2008. View Article : Google Scholar
|
|
10
|
Engels BM and Hutvagner G: Principles and
effects of microRNA-mediated post-transcriptional gene regulation.
Oncogene. 25:6163–6169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rodriguez A, Griffiths-Jones S, Ashurst JL
and Bradley A: Identification of mammalian microRNA host genes and
transcription units. Genome Res. 14:1902–1910. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol
Med. 20:460–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mishra PJ and Merlino G: MicroRNA
reexpression as differentiation therapy in cancer. J Clin Invest.
119:2119–2123. 2009.PubMed/NCBI
|
|
15
|
Schmittgen TD: Regulation of microRNA
processing in development, differentiation and cancer. J Cell Mol
Med. 12:1811–1819. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ding M, Qiu TF and Zhou PG: microRNA-449a
suppresses non-small cell lung cancer. Cell Biochem Biophys.
71:1255–1259. 2015. View Article : Google Scholar
|
|
17
|
Li J, Li P, Chen T, Gao G, Chen X, Du Y,
Zhang R, Yang R, Zhao W, Dun S, et al: Expression of microRNA-96
and its potential functions by targeting FOXO3 in non-small cell
lung cancer. Tumour Biol. 36:685–692. 2015. View Article : Google Scholar
|
|
18
|
Zhu D, Chen H, Yang X, Chen W, Wang L, Xu
J and Yu L: Decreased microRNA-224 and its clinical significance in
non-small cell lung cancer patients. Diagn Pathol. 9:1982014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu Y, Cheng Y, Song Y, Zhang Z, Deng M,
Wang C, Zheng G and He Z: MicroRNA-493 suppresses tumor growth,
invasion and metastasis of lung cancer by regulating E2F1. PLoS
One. 9:e1026022014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Jiang J, Lv X, Fan L, Huang G, Zhan Y,
Wang M and Lu H: MicroRNA-27b suppresses growth and invasion of
NSCLC cells by targeting Sp1. Tumour Biol. 35:10019–10023. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu SH, Zhang CL, Dong FS and Zhang YM:
miR-99a suppresses the metastasis of human non-small cell lung
cancer cells by targeting AKT1 signaling pathway. J Cell Biochem.
116:268–276. 2015. View Article : Google Scholar
|
|
22
|
Zhong K, Chen K, Han L and Li B:
microRNA-30b/c inhibits non-small cell lung cancer cell
proliferation by targeting Rab18. BMC Cancer. 14:7032014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lo SS, Hung PS, Chen JH, Tu HF, Fang WL,
Chen CY, Chen WT, Gong NR and Wu CW: Overexpression of miR-370 and
downregulation of its novel target TGFβ-RII contribute to the
progression of gastric carcinoma. Oncogene. 31:226–237. 2012.
View Article : Google Scholar
|
|
24
|
Yungang W, Xiaoyu L, Pang T, Wenming L and
Pan X: miR-370 targeted FoxM1 functions as a tumor suppressor in
laryngeal squamous cell carcinoma (LSCC). Biomed Pharmacother.
68:149–154. 2014. View Article : Google Scholar
|
|
25
|
Inoue Ji, Ishida T, Tsukamoto N, Kobayashi
N, Naito A, Azuma S and Yamamoto T: Tumor necrosis factor
receptor-associated factor (TRAF) family: adapter proteins that
mediate cytokine signaling. Exp Cell Res. 254:14–24. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Namjou B, Choi CB, Harley IT,
Alarcón-Riquelme ME, Kelly JA, Glenn SB, Ojwang JO, Adler A, Kim K,
Gallant CJ, et al: Evaluation of TRAF6 in a large multiancestral
lupus cohort. Arthritis Rheum. 64:1960–1969. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xie P: TRAF molecules in cell signaling
and in human diseases. J Mol Signal. 8:72013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Camilleri-Broët S, Cremer I, Marmey B,
Comperat E, Viguié F, Audouin J, Rio MC, Fridman WH, Sautès-Fridman
C and Régnier CH: TRAF4 overexpression is a common characteristic
of human carcinomas. Oncogene. 26:142–147. 2007. View Article : Google Scholar
|
|
29
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
Large-scale meta-analysis of cancer microarray data identifies
common transcriptional profiles of neoplastic transformation and
progression. Proc Natl Acad Sci USA. 101:9309–9314. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang J, Li X, Yang W, Jiang X and Li N:
TRAF4 promotes tumorigenesis of breast cancer through activation of
Akt. Oncol Rep. 32:1312–1318. 2014.PubMed/NCBI
|
|
31
|
Rivas-Perez H and Nana-Sinkam P:
Integrating pulmonary rehabilitation into the multidisciplinary
management of lung cancer: A review. Respir Med. 109:437–442. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Stinchcombe TE: Recent advances in the
treatment of non-small cell and small cell lung cancer. F1000Prime
Rep. 6:1172014. View
Article : Google Scholar
|
|
33
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: microRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Geng Q, Fan T, Zhang B, Wang W, Xu Y and
Hu H: Five microRNAs in plasma as novel biomarkers for screening of
early-stage non-small cell lung cancer. Respir Res. 15:1492014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tafsiri E, Darbouy M, Shadmehr MB,
Zagryazhskaya A, Alizadeh J and Karimipoor M: Expression of miRNAs
in non-small-cell lung carcinomas and their association with
clinicopathological features. Tumour Biol. 36:1603–1612. 2015.
View Article : Google Scholar
|
|
36
|
Gu X, Coates PJ, MacCallum SF, Boldrup L,
Sjöström B and Nylander K: TRAF4 is potently induced by TAp63
isoforms and localised according to differentiation in SCCHN.
Cancer Biol Ther. 6:1986–1990. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Rozan LM and El-Deiry WS: Identification
and characterization of proteins interacting with Traf4, an
enigmatic p53 target. Cancer Biol Ther. 5:1228–1235. 2006.
View Article : Google Scholar : PubMed/NCBI
|