Introduction
The abnormalities of various genes, and multiple
signaling pathways were reported to be involved in the gastric
cancer, a type of gastric epithelial malignancy (1–3). When
subjected to various external stimuli, normal gastric mucosa
suffers from gradual and piecemeal changes, including chronic
inflammation, atrophy, intestinal metaplasia, dysplasia and
tumorigenesis (1–3). Early detection, diagnosis and
treatment are crucial for the prognosis of patients with gastric
cancer (1–3). Chronic atrophic gastritis (CAG) with
intestinal metaplasia is an important precancerous lesion of
gastric mucosa (2,3). The main type of gastric cancer is the
intestinal type, which is associated with several factors, such as
Helicobacter pylori (Hp) infection, radiation, and chronic
bile reflux (1–4). Bile acid-containing gastric and
duodenal reflux induces the chronic inflammation of gastric mucosa
and subsequently gastric intestinal metaplasia (4,5).
Deoxycholic acid (DCA), secondary bile acid, is a type of
carcinogenic factor of the gastrointestinal tract and has received
increasing attention (5–7).
Farnesoid X receptor (FXR) is a member of the
nuclear hormone receptor superfamily and is involved in the
homeostasis of bile acid, cholesterol and lipid (8–10).
Bile acid, as an endogenous ligand, can induce FXR activation,
which is involved in the abnormal growth of the digestive tract,
chronic inflammatory disease, metabolic disease and carcinogenesis
(7–12).
Caudal-related homeobox transcription factor 2
(Cdx2) and mucin 2 (MUC2) are regarded as two key modulators of
intestinal metaplasia (7,13–16).
Cdx2, a key intestine-specific nuclear transcription factor,
regulates the expression of genes on nutrient digestion and
absorption, and plays an important role in the intestinal mucosa,
epithelial cell differentiation, proliferation and the maintenance
of early differentiation characteristics (13,15,17).
Cdx2 is expressed from the early stages of embryonic development to
mature individuals. Cdx2 expression was reported to occur in
intestinal epithelium from endodermal, but not normal gastric
mucosa (7,18). However, accumulating evidence
indicated that Cdx2 is also expressed in CAG-associated gastric
intestinal metaplasia, Barrett's esophagus and some sections of
gastric cancer (13,14,19–21).
In a Cdx2 transgenic mouse model, the overexpression of Cdx2
induces morphologically the occurrence of the intestinal metaplasia
in gastric mucosa (19,22,23).
Such ectopic expression of Cdx2 protein is considered an important
initial factor or switching for intestinal metaplasia of gastric
mucosa, and may modulate the differentiation process of intestinal
epithelial cells.
As a specific marker of intestinal epithelial goblet
cell, secreted MUC2 protein is involved in the renewal and
differentiation of epithelial cells and the maintenance of
epithelial integrity (24,25). MUC2 protein is located downstream of
the Cdx2 protein and can be regulated by Cdx2 (26). It was reported that the treatment of
chenodeoxycholate (CDCA) on gastric epithelial RGM-1 cells in mice
induces the ectopic expression of Cdx2 and MUC2 protein (7). However, the role of bile acid-FXR
pathway in normal human gastric mucous epithelial GES-1 cells model
remains to be elucidated.
The present study focused on the DCA-treated normal
human GES-1 gastric mucosa cells at different concentrations for
different time periods, and then analyzed cell proliferation and
the role of FXR in gastric intestinal metaplasia. We found that DCA
stimulated GES-1 cell growth and induced an increase in the
expression levels of intestinal metaplasia genes in a time- and
dose-dependent manner, including FXR, Cdx2 and
MUC2, via nuclear receptor FXR-NF-κB activity.
Materials and methods
Cells and treatments
Normal human gastric epithelial cells (GES-1) were
purchased from the Beijing Institute for Cancer Research (China)
and cultured in Roswell Park Memorial Institute 1640 (RPMI-1640) at
37°C with 5% CO2, supplemented with 10% fetal bovine
serum (FBS), 100 U/ml penicillin and 100 µg/ml streptomycin
(Gibco-BRL, Grand Island, NY, USA) in a humidified incubator. The
GES-1 cells were untreated or treated with DCA (50, 200, 400, 600
and 1,000 µmol/l), 400 µmol/l DCA plus 1.0
µmol/l GW4064 (FXR agonist) or 400 µmol/l DCA plus 50
µmol/l guggulsterone (Gug.) (FXR antagonist) for different
time points, respectively.
3-(4,5-Dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium
(MTS) assay
The MTS assay was performed to measure GES-1 cell
proliferation, using an ELISA microplate reader Multiskan (Thermo
Labsystems, Beverly, MA, USA), according to the manufacturer's
instructions. Briefly, the cells were cultured in 96-well plates
and treated with the indicated dose of DCA. After incubation with
20 µl MTS solution (Promega, Madison, WI, USA) for 4 h at
37°C, the absorbance value at 490 nm was measured.
Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) staining and flow cytometry
The apoptosis of DCA-treated GES-1 cells was
assessed by flow cytometry, after double staining with Annexin
V-FITC and PI using an apoptosis detection kit (Solarbio Life
Sciences, Beijing, China). Briefly, 1×106 GES-1 cells/ml
were washed gently with cold PBS solution, and incubated with 10
µl Annexin V-FITC and 10 µl PI in binding buffer for
15 min, according to the manufacturer's instructions. Flow
cytometry was then performed, using the Guava easyCyte system
(Millipore, Billerica, MA, USA).
Terminal transferase dUTP nick
end-labeling (TUNEL) assay
A TUNEL assay was performed to measure the apoptosis
of simulated GES-1 cells, using a peroxidase In Situ Cell Death
Detection kit (Roche, Germany), according to the manufacturer's
instructions. Briefly, GES-1 cells were cultured on coverslips and
fixed in 4% formaldehyde. The TUNEL reaction mix was then added.
The apoptosis index (AI) was calculated by the ratio of
TUNEL-positive cells to total cells.
Western blotting
Western blotting was performed to detect the
expression levels of FXR, Cdx2 and MUC2 proteins. Briefly, total
cell lysates of GES-1 were harvested using Nonidet P-40 buffer [50
mM Tris-HCl (pH 7.6), 0.5% Nonidet P-40, 0.1 mM EDTA, 300 mM NaCl,
20% glycerol, 1 mM sodium butyrate, 0.1 mM sodium orthovanadate],
supplemented with protease inhibitor mixture (Roche Applied
Science, Indianapolis, IN, USA) for 10 min on ice. The protein
samples were resolved using SDS-PAGE and then transferred to
ImmunoBlot polyvinylidene fluoride membranes (Millipore) by
semi-dry electroblotting. The membranes were blocked with 5%
non-fat dry milk for 2 h and incubated with primary antibodies
overnight at 4°C. The primary antibodies used were: rabbit anti-FXR
(Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA), rabbit
anti-Cdx2 (Cell Signaling Technology, Beverly, MA, USA), mouse
anti-MUC2 (Sigma-Aldrich, St. Louis, MO, USA), mouse anti-GAPDH
(Zhongshan Jinqiao, Beijing, China) antibodies. After washing, the
membranes were visualized with an enhanced chemiluminescence
detection kit (Amersham Biosciences, Pittsburgh, PA, USA) using
horseradish peroxidase-conjugated goat anti-rabbit or anti-mouse
secondary antibodies (Bio-Rad, Hercules, CA, USA). Band density in
the western blot assay was digitized with TotalLab software (Tyne
and Wear, UK) and normalized to GAPDH protein.
Luciferase reporter assay
GES-1 cells were cultured and co-transfected with
pNF-κB-Luc plasmid (Clontech, Palo Alto, CA, USA) and pRL-TK
(Promega), using Lipofectamine 2000. After 18 h, cells were treated
or not treated with 400 µmol/l DCA, 400 µmol/l DCA
plus 1.0 µmol/l GW4064 or 400 µmol/l DCA plus 50
µmol/l Gug. for 6 h, and then luciferase activity was
assessed with a dual luciferase reporter assay kit (Promega),
according to the manufacturer's instructions. Each assay was
performed in triplicate and repeated at least three times.
Chromatin immunoprecipitation (ChIP)
assay
The GES-1 cells were untreated (control group) or
treated with 400 µmol/l DCA, 400 µmol/l DCA plus 1.0
µmol/l GW4064, or 400 µmol/l DCA plus 50
µmol/l Gug. for 6 h, respectively. Formaldehyde (1%) was
added to the treated or untreated GES-1 cells at 37°C for 10 min.
Glycine (125 mM) (pH 7.0) was used to stop the cross-linking. The
nuclear extracts were then prepared and 10% extracts were saved as
input. The expression levels of input for p50 or p65 protein were
detected by western blotting, using rabbit anti-p65 (Cell Signaling
Technology), rabbit anti-p50 (Cell Signaling Technology), or rabbit
anti-histone H3 (Abcam, Cambridge, UK) as a nuclear control.
Additional extracts were precleared with protein G Dynabeads
(Invitrogen-Life Technologies, Carlsbad, CA, USA) and
immunoprecipitated with rabbit anti-p65, anti-p50, or rabbit IgG
(Sigma-Aldrich) antibodies, and then the protein G Dynabeads
(Invitrogen-Life Technologies). Dynabeads were washed three times
with buffer containing 10 mM Tris-HCl (pH 8.0), 140 mM NaCl, 1 mM
EDTA, 0.1% Triton X-100 and once with TE buffer (10 mM Tris-HCl, pH
8.0, 1 mM EDTA). Dynabeads were divided into two portions. In one
portion, the chromatin fragments were eluted from dynabeads with
elution buffer containing 62.5 mM Tris-HCl (pH 6.8), 200 mM NaCl,
2% SDS, 10 mM dithiothreitol and crosslinks were reverted by
heating at 65°C overnight. In the other portion, the amount of
enriched p65 or p50 protein was analyzed by the western blot
analysis and TotalLab software.
DNA was extracted with phenol/chloroform and ethanol
precipitated from the chromatin fragments and input fraction. The
amount of bound Cdx2 promoter fragments was measured with a
FastStart Universal SYBR-Green Master (Rox) (Roche Diagnostics,
Basel, Switzerland) using an Applied Biosystems 7900 Fast Real-Time
PCR system (Applied Biosystems, Foster City, CA, USA). The primer
sequences targeting the promoter region of human Cdx2 gene
(NM_001265, position from −371 to −234) were used:
5′-CCTCGACGTCTCCAACCAT-3′ (forward) and 5′-CCA CTAGGCTGCAGAGGC-3′
(reverse). The ΔCt value of the immunoprecipitated Cdx2
promoter fragment was normalized to the input fraction Ct value in
the same assay. The ΔΔCt value was then determined by subtracting
the ΔCt (normalized anti-IgG group) from the ΔCt (normalized
anti-p65 or p50 group). Relative binding amount of Cdx2
promoter fragment was calculated by raising 2 to the power of the
negative-ΔΔCt and then normalized to the amount of enriched p65 or
p50 protein. The level of control group was set on 1.
Statistical analysis
Data were presented as the means ± SE (standard
error). SPSS 16.0 software was used to perform one-way analysis of
variance (ANOVA) and followed multiple mean comparisons using the
Student-Newman-Keuls test. P<0.05 was considered statistically
significant for all the analyses. The experiments were repeated
three times.
Results
Effect of DCA on the proliferation and
apoptosis of the GES-1 cells
To determine the effect of DCA stimulation on the
proliferation of GES-1, the cells were cultured in vitro and
then treated with DCA (0, 50, 200, 400, 600 and 1,000
µmol/l) for 24 h, respectively. As shown in Fig. 1A, evident proliferation, strong
refraction and exteremely few exfoliated cells were observed in 50,
200 and 400 µmol/l DCA groups. Although the number of
exfoliated cells and intercellular space increased, cell refractive
difference decreased in 600 and 1,000 µmol/l DCA groups.
Notably, cytoplasmic vacuoles increased and the cell membranes
ruptured in some DCA (600 and 1,000 µmol/l)-treated GES-1
cells. The proliferation of cells was then analyzed by an MTS
assay. The data in Fig. 1B show
that DCA stimulation promoted the proliferation of normal human
GES-1 cell in a dose-dependent manner. Compared with the 0
µmol/l control group, the highest reproductive rate of GES-1
cells was detected in the 400 µmol/l group (P<0.01). The
treatment of 1,000 µmol/l DCA for 24 h resulted in a
decrease of GES-1 cell proliferation (P<0.05). Furthermore, the
potential apoptotic mode of DCA-treated GES-1 cells was evaluated
by double staining with FITC-conjugated annexin V/PI, followed by
flow cytometry. As shown in Fig.
1C, after exposure to 1,000 µmol/l DCA, increased GES-1
cells underwent late apoptosis or necrosis (19.76%, Annexin
V-FITC+/PI+), compared with the control group (3.45%). However, the
treatment of 50, 200, 400 and 600 µmol/l DCA failed to
efficiently induce apoptosis of the GES-1 cells.
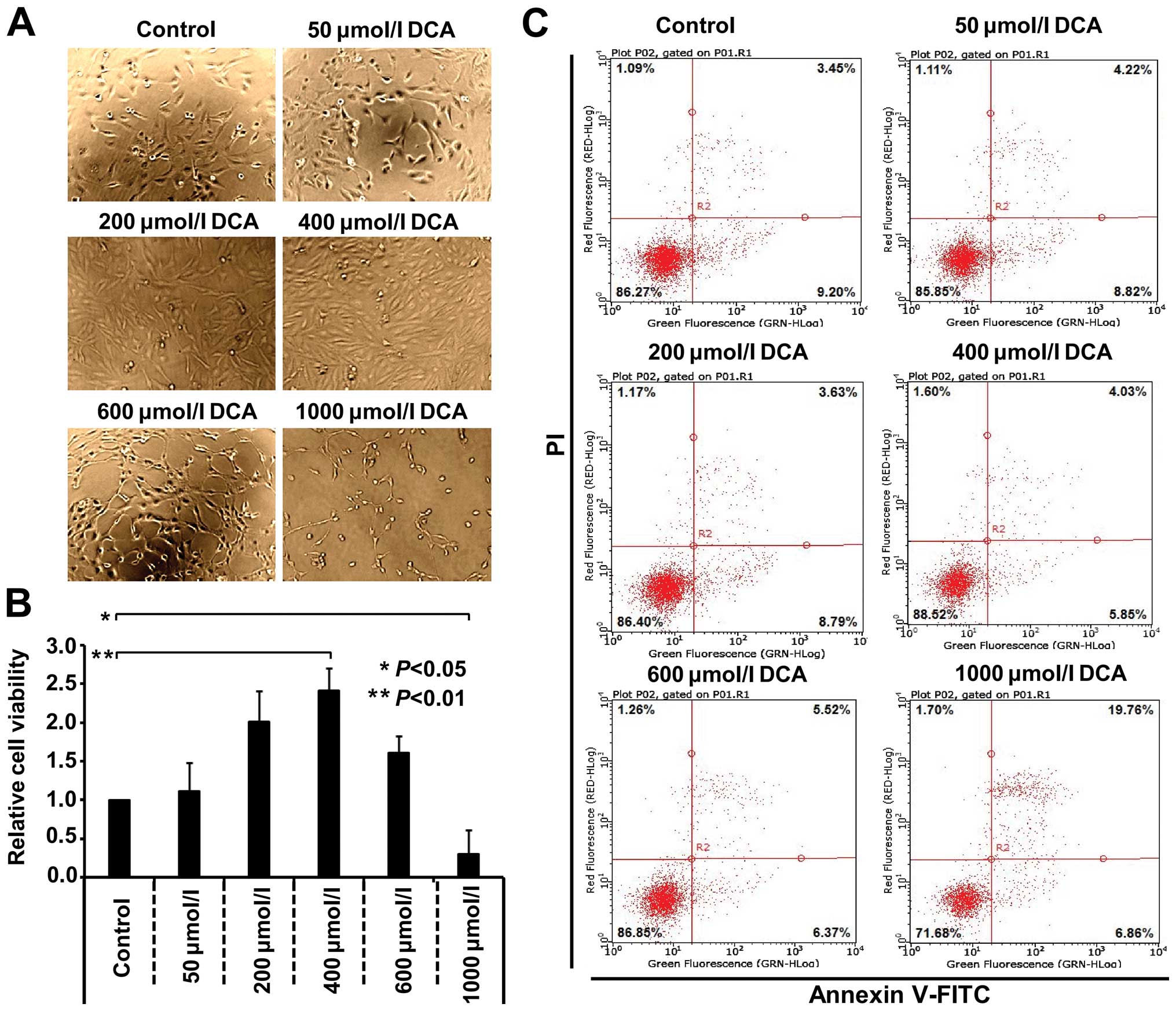 | Figure 1DCA affects the proliferation and
apoptosis of GES-1 cells in a dose-dependent manner. The GES-1
cells were cultured and treated with the DCA agent with the
indicated concentrations (50, 200, 400, 600 and 1,000
µmol/l) or not (control) for 24 h, respectively. (A) Images
of cells in different groups. (B) Cell proliferation was measured
by an MTS assay. One-way analysis of variance (ANOVA) was
performed, and significant differences were indicated as:
*P<0.05, **P<0.01. (C) Cell apoptosis
was also measured by Annexin V-FITC/PI staining, followed by flow
cytometry. DCA, deoxycholic acid; GES-1, gastric epithelial
cells. |
In addition, the MTS and TUNEL assays were performed
in GES-1 cells under the treatment of 400 µmol/l DCA for
different time points (0, 3, 6, 12, 24, 48 and 72 h). As shown in
Fig. 2A, the highest reproductive
rate was detected in the 6 h group (P<0.01), whereas the lowest
reproductive rate was detected in the 72 h group (P<0.05),
compared with the 0 h group. A reverse trend was observed in the
apoptosis level of GES-1 cells. As shown in Fig. 2B, compared with the 0 h group, the
400 µmol/l DCA for 48 and 72 h induced the apoptosis of
GES-1 cells, whereas 400 µmol/l DCA for 6 h reduced cell
apoptosis (P<0.05). The highest apoptotic rates were detectable
in the 72 h group (P<0.01). These observations demonstrated that
the effect of DCA on the growth of GES-1 cells exhibited duality,
i.e., DCA promoted proliferation and attenuated the apoptosis of
GES-1 cells in vitro under a low-moderate dose for short
period of time. Inhibition of the cell proliferation and an
increase of apoptosis under a high dose for a long period of time
may be associated with the toxicity of DCA to GES-1 cells.
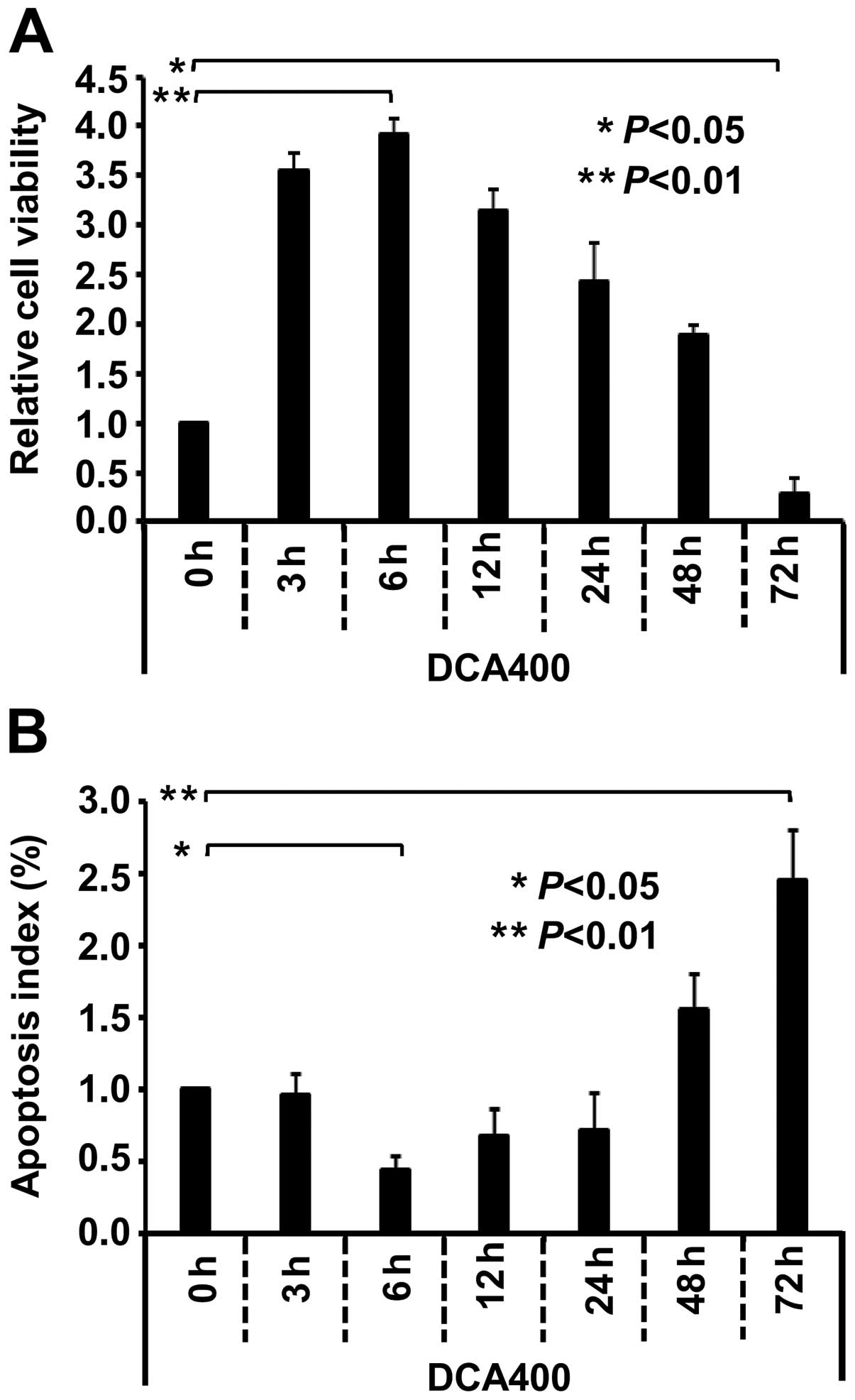 | Figure 2DCA affects the proliferation and
apoptosis of GES-1 cells in a time-dependent manner. The GES-1
cells were cultured and treated with 400 µmol/l DCA for the
indicated time points (0, 3, 6, 12, 24, 48 and 72 h), respectively.
(A) Proliferation or (B) apoptosis of cells was then measured by an
MTS or TUNEL assay. One-way analysis of variance was performed, and
significant differences were indicated as follows:
*P<0.05, **P<0.01. DCA, deoxycholic
acid; GES-1, gastric epithelial cells. |
Effect of DCA on the expression of FXR,
Cdx2 and MUC2 in GES-1 cells
We performed western blotting to detect the
expression levels of FXR, Cdx2 and MUC2, to determine the role of
DCA in the intestinal metaplasia and its expression in GES-1 cells.
As shown in Fig. 3, when the GES-1
cells were treated with DCA (0, 50, 100, 200 and 400 µmol/l)
for 6 h, the expression of endogenous FXR, Cdx2 and MUC2 increased
at the protein levels in a dose-dependent manner. Compared with the
0 µmol/l control group, the highest expression levels of
FXR, Cdx2 and MUC2 protein were observed in the 400 µmol/l
group (P<0.001, P<0.01). Significantly increased expression
levels of FXR, Cdx2 and MUC2 mRNA were also
observed as measured using quantitative PCR (qPCR) in the 400
µmol/l DCA-treated GES-1 cells (data no shown).
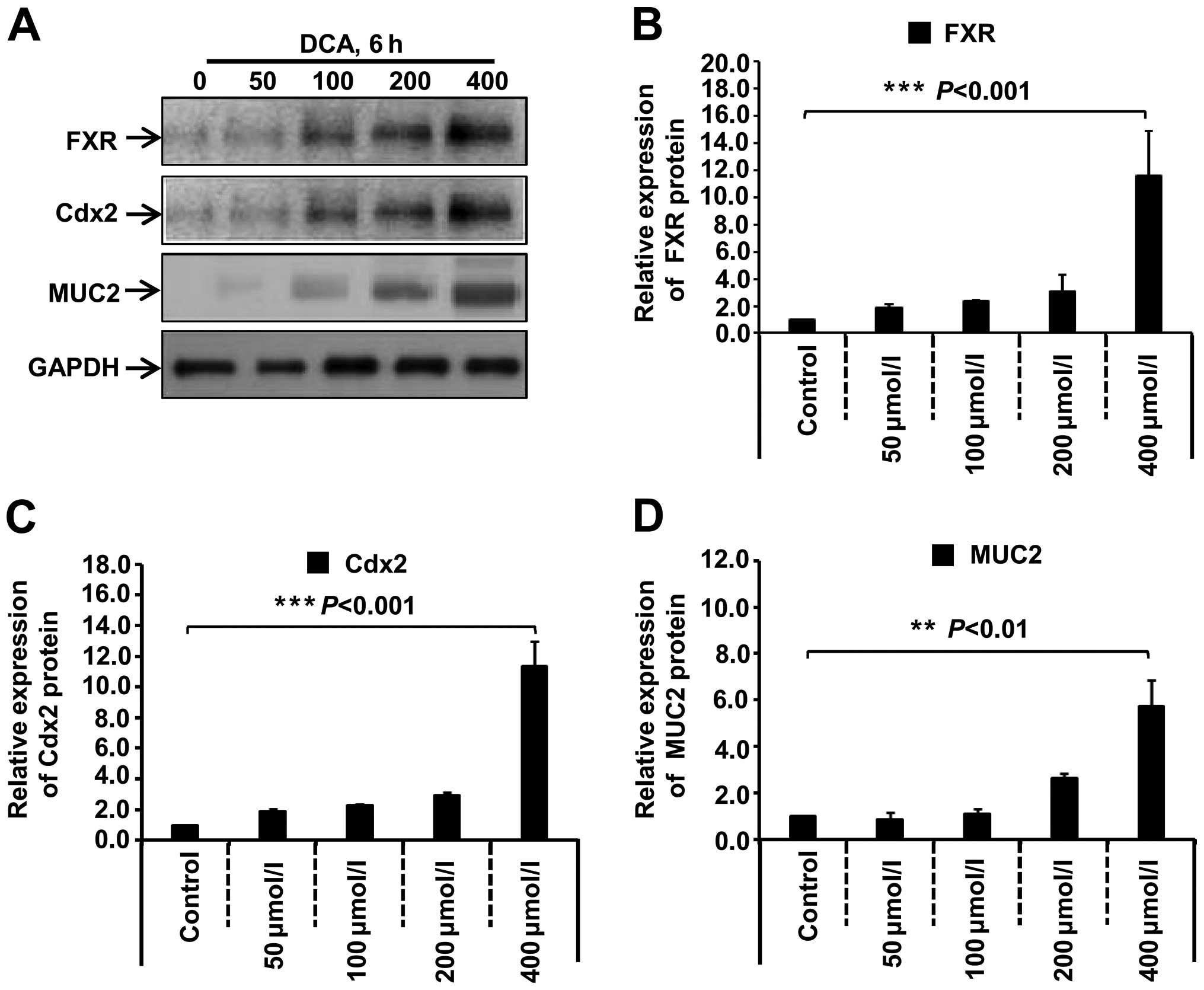 | Figure 3DCA induces the expression of FXR,
Cdx2 and MUC2 protein in GES-1 cells. (A) GES-1 cells were cultured
and treated with the DCA agent with the indicated concentrations
(50, 100, 200 and 400 µmol/l) or not (control),
respectively, for 6 h. The expression levels of FXR, Cdx2 and MUC2
protein were measured by western blotting. (B–D) Band density was
digitized with TotalLab software, and the expression levels of FXR,
Cdx2 and MUC2 protein were normalized against the GAPDH protein.
One-way analysis of variance (ANOVA) was performed, and significant
differences were indicated as follows: ***P<0.001,
**P<0.01. DCA, deoxycholic acid; FXR, farnesoid X
receptor; Cdx2, caudal-related homeobox transcription factor 2;
MUC2, mucin 2; GES-1, gastric epithelial cells. |
Under the treatment of 400 µmol/l DCA, we
also examined the expression level of FXR, Cdx2 and MUC2 in GES-1
cells at different time points (0, 3, 6, 12 or 24 h). As shown in
Fig 4, compared with the 0 h group,
the expression levels of FXR and Cdx2 protein increased at 3 h,
reached a maximum at 6 h (Fig.
4A–C, P<0.001) and then decreased at 12 and 24 h (Fig. 4A–C, P<0.01, P<0.05) while MUC2
expression increased following DCA treatment, showing the highest
signal at 12 h (Fig. 4A and D,
P<0.001), and then decreased at 24 h (Fig. 4A and D, P<0.05). A similar trend
was detectable when FXR, Cdx2 and MUC2 mRNA
levels were measured (data not shown). These results suggested
that, after the stimulation of DCA in GES-1 cells, FXR expression
increased and gradually induced the expression of Cdx2 and
intestinal metaplasia marker MUC2.
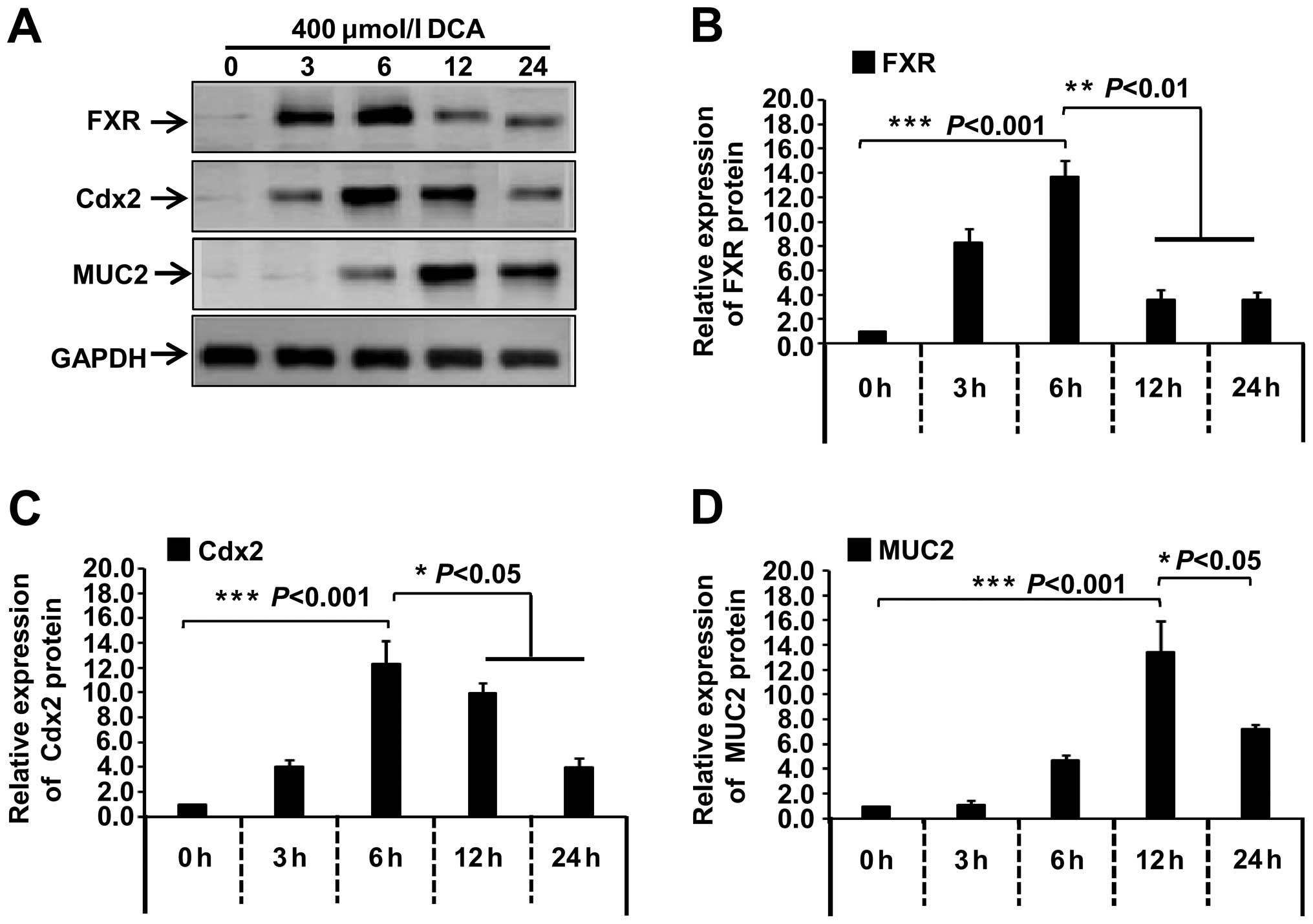 | Figure 4Effect of 400 µmol/l DCA on
the expression of FXR, Cdx2 and MUC2 proteins in GES-1 cells. (A)
GES-1 cells were cultured and treated with the 400 µmol/l
DCA for the indicated time (0, 3, 6, 12 and 24 h), respectively.
The expression levels of FXR, Cdx2 and MUC2 protein were measured
by western blotting. (B–D) Band density was digitized with TotalLab
software, and the expression levels of FXR, Cdx2 and MUC2 protein
were normalized against the GAPDH protein. One-way analysis of
variance (ANOVA) was performed, and significant differences were
indicated as follows: *P<0.05,
**P<0.01, ***P<0.001. DCA, deoxycholic
acid; FXR, farnesoid X receptor; Cdx2, caudal-related homeobox
transcription factor 2; MUC2, mucin 2; GES-1, gastric epithelial
cells. |
GW4064/Guggulsterone affect the
expression of FXR, Cdx2 and MUC2 in DCA-induced GES-1 cells
To provide insight into the role of FXR regulation
and its downstream gene expression in the DCA-induced gastric
mucous membrane intestinal metaplasia, we utilized the synthetic
high-affinity FXR agonist GW4064 and FXR antagonist Gug. As shown
in Fig. 5, compared with the
control group, when GES-1 cells were treated with 400 µmol/l
DCA for 6 h, the expression levels of FXR, Cdx2 and MUC2 protein
increased. However, the treatment of FXR agonist GW4064
significantly increased the expression levels of FXR, Cdx2 and MUC2
protein (P<0.01). By contrast, after FXR antagonist Gug.
stimulation, the upregulation of DCA-induced FXR, Cdx2 and MUC2
expression was attenuated (P<0.05). The similar trend was also
obtained in the measure of FXR, Cdx2 and MUC2
mRNA levels (data not shown). These results demonstrated that DCA
is capable of modulating the expression of Cdx2 and the downstream
MUC2 via the nuclear receptor FXR. Additionally, FXR signaling
pathway plays an important role in DCA-induced gastric intestinal
metaplasia and carcinogenesis.
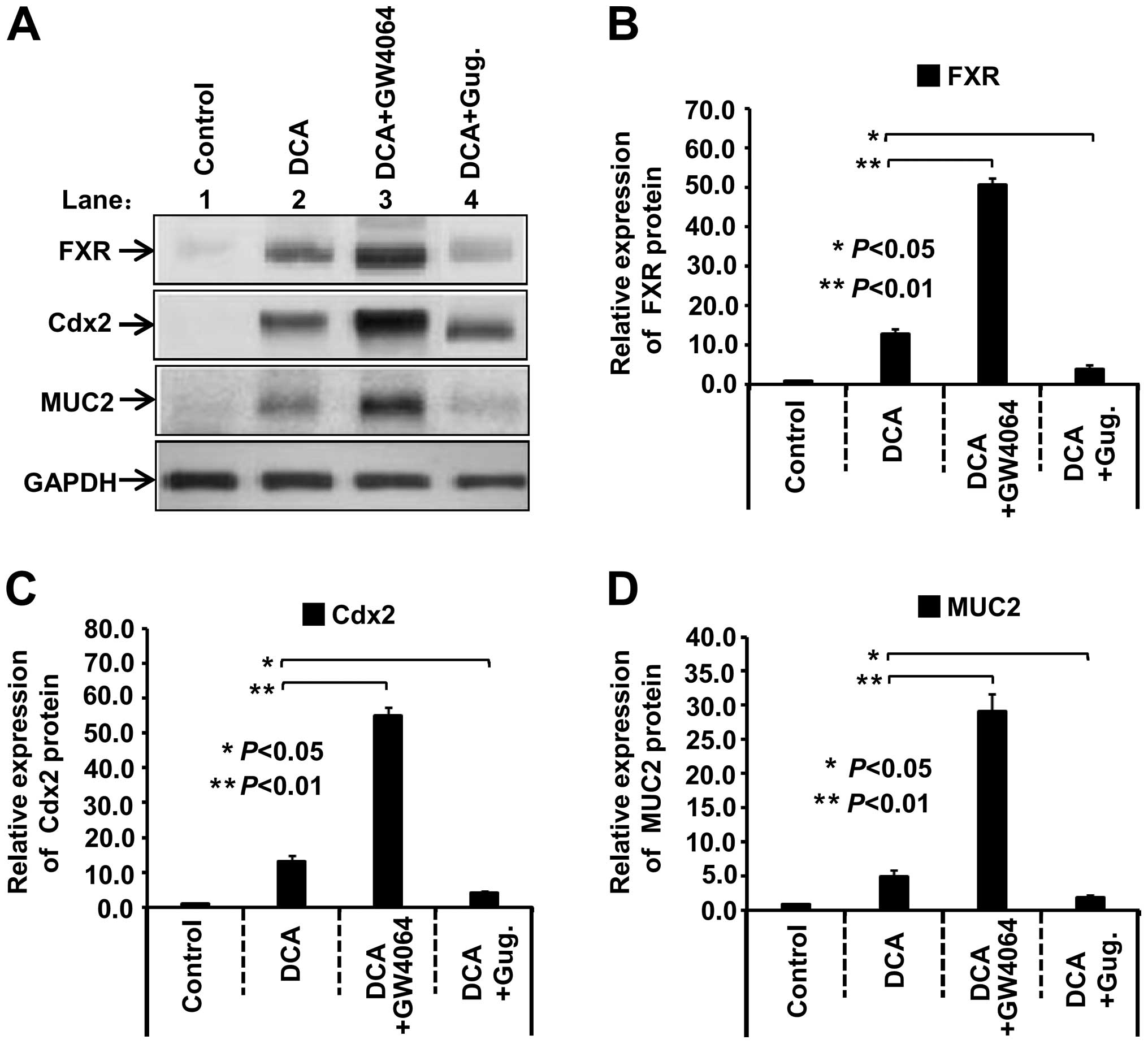 | Figure 5GW4064/guggulsterone (Gug.) affect
the expression of FXR, Cdx2 and MUC2 in 400 µmol/l
DCA-treated GES-1 cells. (A) GES-1 cells were cultured and treated
with 400 µmol/l DCA, 400 µmol/l DCA plus 1.0
µmol/l GW4064 or 400 µmol/l DCA plus 50 µmol/l
Gug. for 6 h, respectively. (B–D) Band density was digitized with
TotalLab software, and the expression levels of FXR, Cdx2 and MUC2
protein were normalized against the GAPDH protein. One-way analysis
of variance (ANOVA) was performed, and significant differences were
indicated as: *P<0.05, **P<0.01. FXR,
farnesoid X receptor; Cdx2, caudal-related homeobox transcription
factor 2; MUC2, mucin 2; DCA, deoxycholic acid; GES-1, gastric
epithelial cells. |
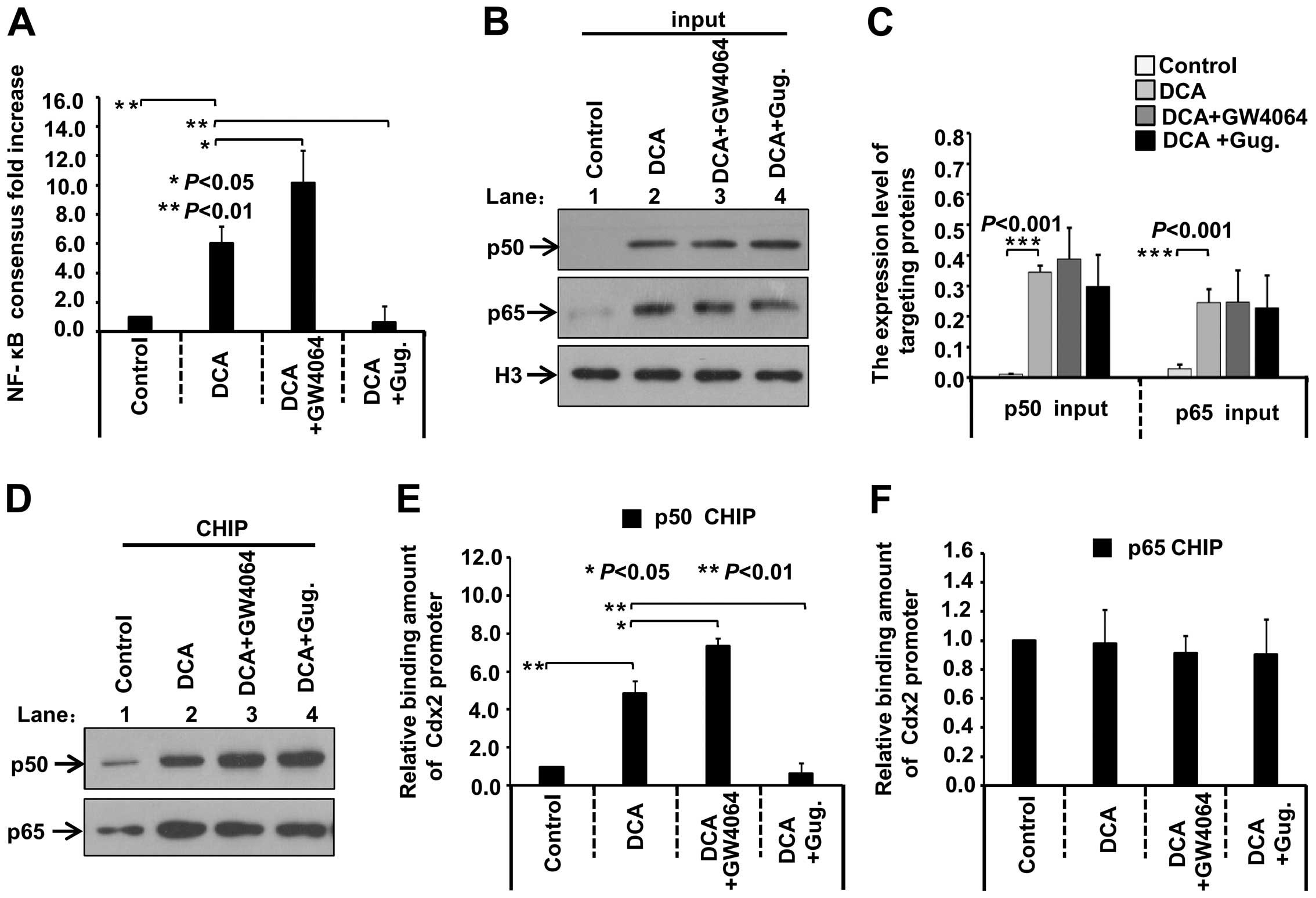
GW4064/Guggulsterone affect DCA-induced
NF-κB activity and binding of the Cdx2 promoter and p50
protein
Moreover, we analyzed the association between FXR
and NF-κB activity. GES-1 cells were co-transfected with pNF-κB-Luc
and pRL-TK plasmid. After 18 h, the cells were treated or not
treated with 400 µmol/l DCA, 400 µmol/l DCA plus 1.0
µmol/l GW4064 or 400 µmol/l DCA plus 50 µmol/l
Gug. for 6 h, respectively. The reporter activity was then
measured. As shown in Fig. 6A,
NF-κB activity in the GES-1 cells increased in response to DCA
stimulation, compared with the control group (P<0.01). GW4064
enhanced the activity (P<0.05), whereas Gug. significantly
reduced the activity (P<0.01), suggesting that DCA-induced FXR
activation is closely related with the NF-κB activity. We then
performed the CHIP assay to investigate the role of GW4064/Gug. in
the association between p50/p65 protein and Cdx2 promoter regions.
As shown in Fig. 6B–C, the input of
p50 and p65 increased in the DCA group, compared with the control
group (P<0.001), which may be due to the nuclear location or
upregulation of p50/p65 expression in response to the DCA
stimulation. However, the DCA-induced increase of nuclear p50/p65
protein was not affected by treatment with GW4064 or Gug. In
addition, the precipitated nuclear p50 and p65 increased in the DCA
group, compared with the control group (Fig. 6D). Thus, the amount of bound Cdx2
promoter fragment was normalized with the input fraction and the
band density value of the enriched p50 and p65 protein. The results
showed that DCA exposure enabled p50 protein to interact with the
Cdx2 promoter (Fig. 6E, P<0.01)
but not p65 (Fig 6F). GW4064
enhanced the binding activity of p50 protein and the Cdx2 promoter
region (Fig. 6E, P<0.05), By
contrast, Gug. reduced the Cdx2 target region enriched by the p50
protein (Fig. 6E, P<0.01). These
data demonstrated that DCA activated the FXR pathway and induced
the expression of Cdx2 by stimulating the NF-κB mediated
transcriptional activity of the Cdx2 promoter.
Discussion
Besides the intestinal system, kidneys, liver or
adrenal glands, FXR exhibits a low expression level in the stomach,
heart, lung and fat tissue (9,11,12,27–29).
In the present study, we observed FXR expression at a significantly
lower level in cultured human GES-1 gastric mucosa cells and
assessed the functional connection between bile acid-FXR pathway
and intestinal metaplasia of gastric mucosa cells.
As the major cause of gastric mucosal injury, bile
reflux is one of the risk factors for gastric cancer. However, the
exact molecular mechanism remains elusive. CDCA, DCA and bile acid
lithocholic acid are important ingredients that act as FXR ligands
to activate the FXR pathway (7,8). In
order to mimic the bile reflux, CDCA or DCA is often used to treat
gastric mucosal cells, and then further investigate the molecular
mechanism. It was reported that activation of the FXR pathway in
mice is able to resist the gastrointestinal mucosal damage of
non-steroidal anti-inflammatory drugs (NSAIDs) (30). FXR functions to protect gastric
epithelial cells against inflammation-mediated damage (11). Those findings suggest that proper
stimulation of FXR serves as a type of protective mechanism for
mucosa. However, in the present study, DCA was used to treat normal
human GES-1 cells to mimic the bile reflux. We found that
stimulation of the low-moderate concentration of DCA on human GES-1
cells for a short period of time increased cell viability and
activated the FXR pathway and the expression of Cdx2, the critical
modulator of intestinal metaplasia in gastric mucosa, which is in
concordance with the results obtained in a mouse model by Xu et
al (7). The results indicated
that activation of the FXR function differs in various ligands. In
the intestinal system, FXR expression contributes to the normal
cell function and inhibition of tumor promotion, whereas in
stomach, abnormal FXR predisposes individuals to intestinal
metaplasia in gastric mucosa and tumors occurrence.
Factors, such as NF-κB and MAPK pathways, contribute
to the enhancement of Cdx2 expression (15,18,31).
The transcription factor NF-κB activates the gene transcription of
many cell processes, including inflammation, cell proliferation,
differentiation and apoptosis (32,33). A
typical NF-κB complex contains the heterologous dimer p50-p65
(32,33). Under resting state, p50 and p65 are
associated with the IκB predominantly in the cytoplasm of the
majority of cells. When treated with toxins, lipopolysaccharides,
phospholipase A and other stimuli, NF-κB separates from IκB,
translocates into the nuclear compartment, and then binds to its
target gene promoter or enhancer to regulate gene transcription
(32,33). It was reported that Cdx2
promoter contains NF-κB binding sites, and can be combined with
different subunits of NF-κB. p65 and p50 exhibit different roles of
transcriptional activity of Cdx2 (31,34,35).
The translocation of NF-κB p50 and p65 subunits is associated with
the upregulation of Cdx2, and the binding of p50 subunit with the
Cdx2 promoter fragment leads to an increase in the promoter
activity of Cdx2 in human esophageal cells (34,36).
In the present study, we also observed activation of the NF-κB
pathway in the DCA-treated human GES-1 cells. Additionally, the
binding activity of NF-κB p50 protein and Cdx2 gene can be
affected by the FXR pathway inhibitors or activators. This effect
means that the NF-κB pathway is involved in the upregulation of
Cdx2 expression in the DCA-induced FXR pathway activation,
resulting in intestinal metaplasia of gastric mucosa.
Previous studies have shown that activated Cdx2
plays a role in the intestinal metaplasia of gastric mucosa and
gastric tumorigenesis through the association with the expression
of claudin-3 and -4, cyclooxygenase-2 (COX-2), Sonic hedgehog (Shh)
or mutations of p53, and APC genes (19,37–39).
For example, the homeobox protein Cdx2 reduces Cox-2 transcription
by inactivating the DNA-binding capacity of NF-κB (39). Cdx2 was reported to enhance
significantly the expression of MUC2 mRNA via binding to the
MUC2 gene promoter (26).
The present study has shown that the expression levels of Cdx2 and
MUC2 were enhanced by the DCA-induced FXR signal. The highest
expression level of MUC2 was at 12 h, later than FXR and Cdx2,
which is in concordance with the results of Xu et al
(7). It is possible that DCA
exposure first leads to the upregulation of FXR and Cdx2
expression, which then increases the expression of MUC2 in GES-1
cells. Similarly, the expression of other intestinal genes, such as
CDX1 and defensin-5, also delayed the Cdx2 protein in human gastric
mucosa with intestinal metaplasia (40). The findings suggest that DCA-induced
FXR-Cdx2 pathway is involved in the intestinal metaplasia of
gastric mucosa, the premalignant lesion of the intestinal type of
gastric cancer.
Chronic bile reflux, a common phenomenon after
subtotal gastrectomy, aggravates the damage of gastric and
esophageal mucosa, and even results in the occurence of gastric
ulcer, gastric cancer, Barrett's esophagus and esophageal cancer
(4,41,42).
Cdx2 protein is not expressed in normal gastric mucosa. However,
long-standing bile reflux alters the microenvironment of the
gastric mucosa through many factors, such as bile and gastric acid
and Hp, and thus form a type of microenvironment, similar to the
gut. Such a microenvironment contributes to the activation of FXR
or the NF-κB signaling pathway. Cdx2 protein regulates intestinal
gene expression in intestinal metaplasia of gastric mucosa.
However, the role of Cdx2 protein in the pathogenesis of cancer
remains to be elucidated and may be tissue-specific.
Upregulation of Cdx2 inhibits proliferation of the
MGC-803 gastric cancer cell line in vitro (43). Cdx2 was also reported to repress the
proliferation of pancreatic cancer cells through the modulation of
cyclin D1 transcriptional activity (44). These results suggest that
Cdx2 functions as the tumor-suppressor gene. It is
speculated that, in the early stage, ectopic expression of Cdx2
acts as the original cause of intestinal metaplasia, and maintains
the intestinal phenotype of gastric cells to repress the malignant
progression. Along with the enhancement of the extracellular
stimuli, such a role of Cdx2 was disturbed, and the precancerous
lesion of gastric mucosa was then transformed into gastric cancer.
With the increase of the malignant or differentiated extent of
tumor tissues, Cdx2 is downregulated or mutated (13,19–21,45).
Therefore, assessing and providing appropriate interventions on the
aberrant expression phase of FXR, CDX2 or MUC2, the related gene
mutation in gastric precancerous lesion and cancer contributes to
the early diagnosis of gastric cancer with the intestinal type, the
judgment of malignant degree, or even the preventiion or reversal
of gastric intestinal metaplasia in patients. In the present study,
the experiment based on the cell culture model was not effective
for the detection of FXR, CDX2 or MUC2 expression phase changes.
More experiments using clinical samples are to be performed in our
laboratory to verify the results obtained.
In summary, the stimulation of DCA is capable of
inducing the ectopic expression of Cdx2 and MUC2 in human gastric
epithelial cells via FXR-mediated estrogen receptor activation and
FXR-associated NF-κB activity. DCA, a key biological sensor or
metabolic regulator, acts as the activator of gastric mucosa FXR
and causes intestinal metaplasia. This plays an important role in
the intestinal metaplasia of gastric mucosa induced by bile reflux,
which is involved in the dysplasia or carcinogenesis of gastric
mucosa.
Acknowledgments
The present study was supported by a grant from the
National Natural Science Foundation of China (no. 81200261), and a
grant from the Science and Technology Fund Project of Tianjin
Health Bureau (07K222).
References
|
1
|
Park JY, von Karsa L and Herrero R:
Prevention strategies for gastric cancer: A global perspective.
Clin Endosc. 47:478–489. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Correa P, Piazuelo MB and Wilson KT:
Pathology of gastric intestinal metaplasia: Clinical implications.
Am J Gastroenterol. 105:493–498. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dinis-Ribeiro M, Areia M, de Vries AC,
Marcos-Pinto R, Monteiro-Soares M, O'Connor A, Pereira C,
Pimentel-Nunes P, Correia R, Ensari A, et al European Society of
Gastrointestinal Endoscopy; European Helicobacter Study Group;
European Society of Pathology; Sociedade Portuguesa de Endoscopia
Digestiva: Management of precancerous conditions and lesions in the
stomach (MAPS): Guideline from the European Society of
Gastrointestinal Endoscopy (ESGE), European Helicobacter Study
Group (EHSG), European Society of Pathology (ESP), and the
Sociedade Portuguesa de Endoscopia Digestiva (SPED). Endoscopy.
44:74–94. 2012. View Article : Google Scholar :
|
|
4
|
Dixon MF, Mapstone NP, Neville PM,
Moayyedi P and Axon AT: Bile reflux gastritis and intestinal
metaplasia at the cardia. Gut. 51:351–355. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Martinez-Augustin O and Sanchez de Medina
F: Intestinal bile acid physiology and pathophysiology. World J
Gastroenterol. 14:5630–5640. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Huo X, Juergens S, Zhang X, Rezaei D, Yu
C, Strauch ED, Wang JY, Cheng E, Meyer F, Wang DH, et al:
Deoxycholic acid causes DNA damage while inducing apoptotic
resistance through NF-κB activation in benign Barrett's epithelial
cells. Am J Physiol Gastrointest Liver Physiol. 301:G278–G286.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xu Y, Watanabe T, Tanigawa T, Machida H,
Okazaki H, Yamagami H, Watanabe K, Tominaga K, Fujiwara Y, Oshitani
N, et al: Bile acids induce cdx2 expression through the farnesoid ×
receptor in gastric epithelial cells. J Clin Biochem Nutr.
46:81–86. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mazuy C, Helleboid A, Staels B and
Lefebvre P: Nuclear bile acid signaling through the farnesoid X
receptor. Cell Mol Life Sci. 72:1631–1650. 2015. View Article : Google Scholar
|
|
9
|
Matsubara T, Li F and Gonzalez FJ: FXR
signaling in the enterohepatic system. Mol Cell Endocrinol.
368:17–29. 2013. View Article : Google Scholar
|
|
10
|
Li S, Ni A and Feng GS: Bridging cell
surface receptor with nuclear receptors in control of bile acid
homeostasis. Acta Pharmacol Sin. 36:113–118. 2015. View Article : Google Scholar
|
|
11
|
Lian F, Xing X, Yuan G, Schäfer C, Rauser
S, Walch A, Röcken C, Ebeling M, Wright MB, Schmid RM, et al:
Farnesoid X receptor protects human and murine gastric epithelial
cells against inflammation-induced damage. Biochem J. 438:315–323.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shaik FB, Prasad DV and Narala VR: Role of
farnesoid X receptor in inflammation and resolution. Inflamm Res.
64:9–20. 2015. View Article : Google Scholar
|
|
13
|
Yan LH, Wei WY, Xie YB and Xiao Q: New
insights into the functions and localization of the homeotic gene
CDX2 in gastric cancer. World J Gastroenterol. 20:3960–3966. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kerkhof M, Bax DA, Moons LM, van Vuuren
AJ, van Dekken H, Steyerberg EW, Kuipers EJ, Kusters JG and
Siersema PD; Cybar Study Group: Does CDX2 expression predict
Barrett's meta-plasia in oesophageal columnar epithelium without
goblet cells? Aliment Pharmacol Ther. 24:1613–1621. 2006.
View Article : Google Scholar
|
|
15
|
Coskun M, Troelsen JT and Nielsen OH: The
role of CDX2 in intestinal homeostasis and inflammation. Biochim
Biophys Acta. 1812:283–289. 2011. View Article : Google Scholar
|
|
16
|
Roessler K, Mönig SP, Schneider PM,
Hanisch FG, Landsberg S, Thiele J, Hölscher AH, Dienes HP and
Baldus SE: Co-expression of CDX2 and MUC2 in gastric carcinomas:
Correlations with clinico-pathological parameters and prognosis.
World J Gastroenterol. 11:3182–3188. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Boyd M, Hansen M, Jensen TG, Perearnau A,
Olsen AK, Bram LL, Bak M, Tommerup N, Olsen J and Troelsen JT:
Genome-wide analysis of CDX2 binding in intestinal epithelial cells
(Caco-2). J Biol Chem. 285:25115–25125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Houde M, Laprise P, Jean D, Blais M,
Asselin C and Rivard N: Intestinal epithelial cell differentiation
involves activation of p38 mitogen-activated protein kinase that
regulates the homeobox transcription factor CDX2. J Biol Chem.
276:21885–21894. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mutoh H, Sakurai S, Satoh K, Tamada K,
Kita H, Osawa H, Tomiyama T, Sato Y, Yamamoto H, Isoda N, et al:
Development of gastric carcinoma from intestinal metaplasia in
Cdx2-transgenic mice. Cancer Res. 64:7740–7747. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu Q, Teh M, Ito K, Shah N, Ito Y and
Yeoh KG: CDX2 expression is progressively decreased in human
gastric intestinal metaplasia, dysplasia and cancer. Mod Pathol.
20:1286–1297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Park Y, Srivastava A, Kim GH,
Mino-Kenudson M, Deshpande V, Zukerberg LR, Song GA and Lauwers GY:
CDX2 expression in the intestinal-type gastric epithelial
neoplasia: Frequency and significance. Mod Pathol. 23:54–61. 2010.
View Article : Google Scholar
|
|
22
|
Mutoh H, Satoh K, Kita H, Sakamoto H,
Hayakawa H, Yamamoto H, Isoda N, Tamada K, Ido K and Sugano K: Cdx2
specifies the differentiation of morphological as well as
functional absorptive enterocytes of the small intestine. Int J Dev
Biol. 49:867–871. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Silberg DG, Sullivan J, Kang E, Swain GP,
Moffett J, Sund NJ, Sackett SD and Kaestner KH: Cdx2 ectopic
expression induces gastric intestinal metaplasia in transgenic
mice. Gastroenterology. 122:689–696. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Johansson ME and Hansson GC: Mucus and the
goblet cell. Dig Dis. 31:305–309. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Johansson ME, Sjövall H and Hansson GC:
The gastrointestinal mucus system in health and disease. Nat Rev
Gastroenterol Hepatol. 10:352–361. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yamamoto H, Bai YQ and Yuasa Y:
Homeodomain protein CDX2 regulates goblet-specific MUC2 gene
expression. Biochem Biophys Res Commun. 300:813–818. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang XF, Zhao WY and Huang WD: FXR and
liver carcinogenesis. Acta Pharmacol Sin. 36:37–43. 2015.
View Article : Google Scholar
|
|
28
|
van de Winkel A, van Zoest KP, van Dekken
H, Moons LM, Kuipers EJ and van der Laan LJ: Differential
expression of the nuclear receptors farnesoid X receptor (FXR) and
pregnane X receptor (PXR) for grading dysplasia in patients with
Barrett's oesophagus. Histopathology. 58:246–253. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Khurana S, Raufman JP and Pallone TL: Bile
acids regulate cardiovascular function. Clin Transl Sci. 4:210–218.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fiorucci S, Mencarelli A, Cipriani S,
Renga B, Palladino G, Santucci L and Distrutti E: Activation of the
farnesoid-X receptor protects against gastrointestinal injury
caused by non-steroidal anti-inflammatory drugs in mice. Br J
Pharmacol. 164:1929–1938. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim S, Domon-Dell C, Wang Q, Chung DH, Di
Cristofano A, Pandolfi PP, Freund JN and Evers BM: PTEN and
TNF-alpha regulation of the intestinal-specific Cdx-2 homeobox gene
through a PI3K, PKB/Akt, and NF-kappaB-dependent pathway.
Gastroenterology. 123:1163–1178. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jing H and Lee S: NF-κB in cellular
senescence and cancer treatment. Mol Cells. 37:189–195. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chen LF and Greene WC: Shaping the nuclear
action of NF-kappaB. Nat Rev Mol Cell Biol. 5:392–401. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Debruyne PR, Witek M, Gong L, Birbe R,
Chervoneva I, Jin T, Domon-Cell C, Palazzo JP, Freund JN, Li P, et
al: Bile acids induce ectopic expression of intestinal guanylyl
cyclase C through nuclear factor-kappaB and Cdx2 in human
esophageal cells. Gastroenterology. 130:1191–1206. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kazumori H, Ishihara S, Rumi MA, Kadowaki
Y and Kinoshita Y: Bile acids directly augment caudal related
homeobox gene Cdx2 expression in oesophageal keratinocytes in
Barrett's epithelium. Gut. 55:16–25. 2006. View Article : Google Scholar
|
|
36
|
Huo X, Zhang HY, Zhang XI, Lynch JP,
Strauch ED, Wang JY, Melton SD, Genta RM, Wang DH, Spechler SJ, et
al: Acid and bile salt-induced CDX2 expression differs in
esophageal squamous cells from patients with and without Barrett's
esophagus. Gastroenterology. 139:194–203.e1. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Satake S, Semba S, Matsuda Y, Usami Y,
Chiba H, Sawada N, Kasuga M and Yokozaki H: Cdx2 transcription
factor regulates claudin-3 and claudin-4 expression during
intestinal differentiation of gastric carcinoma. Pathol Int.
58:156–163. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Mutoh H, Hayakawa H, Sashikawa M, Sakamoto
H and Sugano K: Direct repression of Sonic Hedgehog expression in
the stomach by Cdx2 leads to intestinal transformation. Biochem J.
427:423–434. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mutoh H, Hayakawa H, Sakamoto H and Sugano
K: Homeobox protein CDX2 reduces Cox-2 transcription by
inactivating the DNA-binding capacity of nuclear factor-kappaB. J
Gastroenterol. 42:719–729. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Eda A, Osawa H, Yanaka I, Satoh K, Mutoh
H, Kihira K and Sugano K: Expression of homeobox gene CDX2 precedes
that of CDX1 during the progression of intestinal metaplasia. J
Gastroenterol. 37:94–100. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zaninotto G, Portale G, Parenti A, Lanza
C, Costantini M, Molena D, Ruol A, Battaglia G, Costantino M,
Epifani M, et al: Role of acid and bile reflux in development of
specialised intestinal metaplasia in distal oesophagus. Dig Liver
Dis. 34:251–257. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Weijenborg PW and Bredenoord AJ: How
reflux causes symptoms: Reflux perception in gastroesophageal
reflux disease. Best Pract Res Clin Gastroenterol. 27:353–364.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xie Y, Li L, Wang X, Qin Y, Qian Q, Yuan X
and Xiao Q: Overexpression of Cdx2 inhibits progression of gastric
cancer in vitro. Int J Oncol. 36:509–516. 2010.PubMed/NCBI
|
|
44
|
Takahashi K, Hirano F, Matsumoto K, Aso K
and Haneda M: Homeobox gene CDX2 inhibits human pancreatic cancer
cell proliferation by downregulating cyclin D1 transcriptional
activity. Pancreas. 38:49–57. 2009. View Article : Google Scholar
|
|
45
|
Song JH, Kim CJ, Cho YG, Chae JS, Cao Z,
Nam SW, Lee JY and Park WS: Genetic alterations of the Cdx2 gene in
gastric cancer. APMIS. 116:74–80. 2008. View Article : Google Scholar : PubMed/NCBI
|