Introduction
Breast cancer is the second most frequent
cancer-related death worldwide (1).
In China, it is predicted that breast cancer incidence will
increase to 85/100,000 women by 2021 (2). Estrogen is a key hormone in breast
cancer development. Estrogen exerts its biological activity,
including enhancing cell proliferation related to breast cancer
development, through actions mediated by two estrogen receptor
subtypes, ERα and ERβ (reviewed in ref. 3). Although both estrogen receptors (ERs)
share homology in their DNA and ligand-binding domains, they are
the products of independent genes (reviewed in refs. 4,5).
Moreover, unlike ERα whose expression is limited to breast
epithelial cells, ERβ expression is widely distributed, not only in
epithelial cell nuclei of normal and malignant glands but also in
stromal and endothelial cell nuclei and in the myoepithelium of
normal glands (6). These
observation are consistent with the concept that ERβ may have a
biological role distinct from ERα.
Generally, ERα is the key ER contributing to the
onset and progression of breast cancer and is a good prognostic
marker predicting the likelihood of the patient response to
adjuvant endocrine therapy (reviewed in refs. 7,8). ERβ
is a negative modulator of ERα, suppressing ERα transcription and
inhibiting the sensitivity to estrogen, thus being regarded as a
protective factor in the development of endocrine tumors including
breast cancer (reviewed in refs. 9,10).
However, there are conflicting studies regarding these ERs.
ERα-positive breast cancers seem to show less invasiveness and more
favorable prognosis (7–9,11,12),
and a significant loss of ERβ expression is observed in a
proportion of invasive carcinomas (6). Clinical-based studies have shown that
~30% of ER α-positive breast cancer cells do not respond to
first-line endocrine therapies, and the majority of relapsing
tumors still expresses ERα (reviewed in ref. 13). Thus, the role of ER subtypes in the
progression and endocrine resistance of breast cancer has not been
clearly elucidated.
Cytokines are proteins or peptides mainly produced
by immune cells, which can promote the expression of a variety of
genes involved in the survival or death of different target cells
(14). In the tumor
microenvironment, IL-4 or IFN-γ has been shown to exert both
positive and negative effects on the development and progression of
cancer (15,16). There is evidence suggesting that the
IL-4 receptor (IL-4R) and the IFN-γ receptor (IFN-γR) are detected
on most immune and epithelial cells and a variety of human tumors
of epithelial origin (17–19). Various studies illustrated that
IL-4R expression level in tumor cells is much higher than in normal
cells (20–24). Tumor cells are often surrounded by
infiltrating inflammatory cells, particularly lymphocytes and
macrophages, which are the main sources of cytokines. Notably,
immunohistochemistry and cell-based studies in vitro show
tumor tissues and tumor cell lines, including pancreatic, prostate,
breast and bladder cancer, may be another source of cytokines in
the tumor microenvironment (14,25,26).
Recently, the novel immunoediting theory emphasizes the dynamic
interaction between the host immune system and the developing
tumor. The theory points out that substances produced by tumor
tissues contribute to suppressing antitumor immunity, which may
provide an obstacle for exploring antitumor cytokine approaches
(15). In thyroid cancer, the
presence of IL-4 and IL-10 in the tumor microenvironment
contributes to thyroid cancer cell survival and proliferation
(27). Furthermore, several
investigators have provided evidence that there is a correlation
between IL-4 expression and local intracrine activity of estradiol
(E2) as well as the expression of estrogen receptor in breast
cancer. In the different breast cancer cell lines ZR-75-1 and
T-47D, IL-4 shows increasing or decreasing effect on sex steroid
production, respectively (28). In
addition, intratumoral IL-4 levels tend to be negatively correlated
with the level of hormone receptor, which seems to indicate
crosstalk between cytokine receptors and ER may result in
ligand-independent activation of ER signaling pathways (29).
Based on these studies demonstrating interactions
between ERs and cytokine pathways, we evaluated interaction between
the two ERs and the cytokines IL-4 and IFN-γ, and whether this
interaction modulates malignant behavior.
Materials and methods
Cell lines and cell culture
Human breast cancer MCF-7 and MDA-MB-231 cell lines
were obtained from the American Type Culture Collection. Cells were
cultured in RPMI-1640 containing 10% fetal bovine serum (FBS) (both
from Life Technologies, Inc., Gaithersburg, MD, USA).
Cell transfection with pGenesil-ERα small
interfering RNA (siRNA), pGenesil-ERβ siRNA and pGenesil-scrambled
siRNA vectors and generation of stable cell lines
Using the pGenesil-1 vector encoding enhanced green
fluorescent protein and kanamycin/neomycin-resistance genes,
pGenesil-ERα siRNA, pGenesil-ERβ siRNA and pGenesil-scrambled siRNA
plasmids were constructed by Wuhan Genesil Biotechnology Co., Ltd.
The sense sequences of the ERα siRNA pair (underlined sequence)
were as follows: 5′-GATCCG(CTCATCCTCTCCCACATCA)TTCAAGACG(TGATGTGGGAGAGGATGAG)TTTTTTGTCGACA-3′,
and the antisense sequence of the ERα siRNA was,
5′-AGCTTGTCGTACAAAAAA(CTCATCCTCTCCCACATCA)CGTCTTGAA(TGATGTGGGAGAGGATGAG)CG-3′.
The sense sequences of the ERβ siRNA pair (underlined sequence)
were as follows: 5′-GATCC(GCCCTGCTGTGATGAATTA)TTCAAGACG(TAATTCATCACAGCAGGGC)TTTTTTGTCGACA-3′,
and the antisense sequences of the ERβ siRNA were:
5′-AGCTTGTCGACAAAAAA(GCCCTGCTGTGATGAATTA)CGTCTTGAA(TAATTCATCACAGCAGGGC)G-3′.
The sense sequences of the scrambled siRNA pair (underlined
sequence) were as follows: 5′-GATCC(GACTTCATAAGGCGCATGC)TTCAAGACG(GCATGCGCCTTATGAAGTC)TTTTTTGTCGACA-3′,
and the antisense sequence of the scrambled siRNA was:
5′-AGCTTGTCGACAAAAAA(GACTTCATAAGGCGCATGC)CGTCTTGAA(GCATGCGCCTTATGAAGTC)G-3′.
To generate the stable ERα siRNA or ERβ siRNA clones in MCF-7
cells, the pGenesil-ERα siRNA or pGenesil-ERβ siRNA construct was
transfected into MCF-7 cells expressing endogenous ERα and ERβ by
Lipofectamine™ 2000 (Invitrogen, San Diego, CA, USA), according to
the manufacturer's protocol. MCF-7 cells (4×105) were
plated into 6-well plates until 90–95% confluence before
transfection and were then transfected with 4 µg of
pGenesil-ERα siRNA or pGenesil-ERβ siRNA. Selection for the
neomycin gene was initiated 48 h after transfection by adding 400
µg/ml of G418 (Invitrogen) to the supplemented culture
medium. This selection medium was changed every 2 days for 4 weeks,
until all non-transfected cells died. Resistant cell clones were
isolated and expanded for further characterization.
pGenesil-scrambled siRNA was also transfected into MCF-7 cells as
non-specific siRNA control.
Treatment of human breast cancer MCF-7
cells in the absence or presence of rhIFN-γ or rhIL-4
MCF-7 cells were cultured in 60-mm culture dishes
(1×106 cells/dish) overnight, and then switched to
medium containing 1% FBS for 24 h. The cells were treated in 1% FBS
with rhIFN-γ (0.1, 1, 10, 50, 100 or 250 ng/ml) or rhIL-4 (0.1, 1,
5, 10, 25 ng/ml) (Sigma, St. Louis, MO, USA) in 0.1%
BSA-phosphate-buffered saline (PBS) for 1, 2, 3 or 4 days. Control
cells were treated with 0.1% bovine serum albumin (BSA) (Sigma)-PBS
vehicle as previously described.
Tamoxifen (TAM) treatment of human breast
cancer MCF-7 cells and their stable transfected clones with ERα
siRNA, ERβ siRNA or scrambled siRNA
MCF-7 cells and their stable transfected clones with
ERα siRNA, ERβ siRNA or scrambled siRNA were plated into 96-well
(4×103 cells/well) plates overnight, and then switched
to medium containing 1% FBS for 24 h. The cells were treated in 1%
FBS with TAM (0.1, 0.5, 1, 5 and 10 µM) (Sigma) in
dimethylsulfoxide (DMSO) for 24 h, or pretreated in 1% FBS with
rhIFN-γ (0.5 or 100 ng/ml) for 72 h and then treated with TAM (10
µM) for 24 h. The final DMSO concentration was 0.1%. Control
cells were treated with DMSO vehicle or 0.1% BSA-PBS vehicle as
previously described.
Semi-quantitative RT-PCR
Total RNA was isolated from cells with TRIzol
(Invitrogen) according to the manufacturer's instructions. Primer
sequences were designed by Vector NTI 8 software and synthesized by
Takara Biotechnology Co., Ltd. (Dalian, China). The primer
sequences were as follows: ERα, 5′-AACAAAGGCATGGAGCATCTGT-3′
(forward) and 5′-GTGATGTAATACTTTTGCAAGG-3′ (reverse); for ERβ,
5′-GCGCTGTCTGCAGCGATTACGC-3′ (forward) and
5′-CACCATTCCCACTTCGTAACAC-3′ (reverse); for Bcl-2,
5′-TGCACCTGACGCCCTTCAC-3′ (forward) and
5′-AGACAGCCAGGAGAAATCAAACAG-3′ (reverse); for Bcl-xL,
5′-ATGTCTCAGAGCAACCGGGAGC-3′ (forward) and
5′-GCGATCCGACTCACCAATACCT-3′ (reverse); for XIAP,
5′-ATGATACCATCTTCCAAAATCC-3′ (forward) and
5′-TTTCTGTAATGAAGTCTGACTT-3′ (reverse); for β-actin,
5′-TGGAATCCTGTGGCATCCATGAAAC-3′ (forward) and
5′-TAAAACGCAGCTCAGTAACAGTCC-3′ (reverse). One-Step RNA PCR kit
(AMV) (Takara Biotechnology) was used for RT-PCR. PCR products were
fractionated on 1.5% agarose gel and analyzed with Quantity One
4.5.6 software (Bio-Rad, Hercules, CA, USA). The results were
normalized against β-actin and presented as target mRNA:β-actin
ratio.
Western blot analysis
Cells were lysed in ice-cold RIPA buffer (150 mM
NaCl, 1% Nonidet P-40, 0.5% deoxycho-late, 0.1% SDS, 50 mM Tris-HCl
pH 8.0, 1 mM PMSF, 10 µg/ml leupeptin and 100 µg/ml
aprotinin) for 45 min on ice. The lysates were centrifuged to
remove cellular debris. Supernatants were analyzed for protein
concentration using the bicinchoninic acid assay kit (Pierce
Biochemicals, Rockford, IL, USA). Total cell lysates (40 µg)
were subjected to 8–10% SDS-PAGE gels and analyzed by blotting with
rabbit polyclonal anti-ERα and anti-ERβ antibody (Santa Cruz
Biotechnology, Santa Cruz, CA. USA), respectively. Membranes were
stripped by incubating with stripping buffer at 50°C for 30 min and
then blotted with mouse monoclonal anti-β-actin antibody (Sigma).
Immunodetection was performed using the corresponding secondary
HRP-conjugated antibody, and HRP activity was detected using
chemiluminescent substrate kit (SuperSignal® West Pico
Trial kit; Pierce Biochemicals).
MTT assay
Cell proliferation was measured with MTT assay.
Briefly, human breast cancer cells were cultured into 96-well
plates at 4×103 cells/well and treated with rhIFN-γ,
rhIL-4 or TAM as previously described. After treatment, the cells
were incubated with 100 µl of MTT solution (0.5 mg/ml;
Sigma) for 4 h at 37°C. After centrifugation, 100 µl of 0.04
M HCl-isopropanol was added. The absorbance was measured at 492 nm
using ELISA microplate reader. Data represent the average
absorbance of five wells in one experiment. The experiment was
repeated twice with similar results.
Enzyme-linked immunosorbent assay
(ELISA)
To evaluate the in vitro production of IFN-γ
and IL-4 by breast cancer cells, 1×105 cells were seeded
into 24-well plates overnight, and the supernatants were collected
and clarified by centrifugation. The level of IFN-γ and IL-4 was
measured using ELISA kits (R&D Systems, Minneapolis, MN, USA)
according to the manufacturer's instructions.
Cell cycle distribution assay
Cells were cultured in serum-free medium for 6 h,
and then harvested and resuspended in PBS containing 0.1 M
propidium iodide solution, 0.1% Triton X-100 (Sigma) and 2% RNase
A. After incubation on ice, in the dark, for 2 h, the samples were
analyzed by flow cytometry. One hundred thousand events were
recorded and the proportion of cells in various phases of the cell
cycle was analyzed using the ModFit LT DNA analysis software
(Becton-Dickinson, San Jose, CA, USA).
Soft-agar colony formation assay
To assess the effect of anchorage-independent
growth, cells (1x104 cells/6-well plate) were grown in
soft agar for 14 days at 37°C using a two layer agar system and the
number of colonies quantitated as previously described (30).
In vitro cell adhesion assay
As previously described (30), 96-well plates were pre-coated with
Matrigel overnight at 37°C. After blocking with 2% BSA solution for
1 h at 37°C, 4×103 cells in 100 µl of serum-free
RPMI-1640 medium containing 0.1% BSA were placed into the wells
precoated with the reconstituted matrix for 1 h at 37°. The cells
were washed with PBS and the MTT assay was performed as previously
described. Data represent the average absorbance of five wells in
one experiment. The experiment was repeated twice with similar
results.
Statistical analysis
Multiple comparisons were performed using one-way
analysis of variance (ANOVA) with Fisher's protected least
significant difference method for post hoc analysis. All
statistical tests were two-sided. For all tests, the level of
significance was set at P<0.05. Statistical analysis was carried
out using SPSS version 11.0 software.
Results
Knockdown of ERβ causes increased cell
proliferation in MCF-7
In order to investigate the role of ER subtypes in
the cell growth of human breast cancer cells, we first analyzed the
expression of ERα and ERβ as well as the cell proliferation of two
breast cancer cell lines, MCF-7 and MDA-MB-231. As shown in
Fig. 1A, not only ERα but also ERβ
was expressed at the medium level in MCF-7, whereas in MDA-MB-231,
ERβ was expressed at a significantly higher level than that in
MCF-7. MTT assay demonstrated that cell proliferation ability in
MCF-7 was stronger than that in MDA-MB-231 after culturing for 4
days (Fig. 1B). These data are
consistent with the possibility that ERα is a positive factor in
promoting tumor growth; whereas, ERβ is a negative factor.
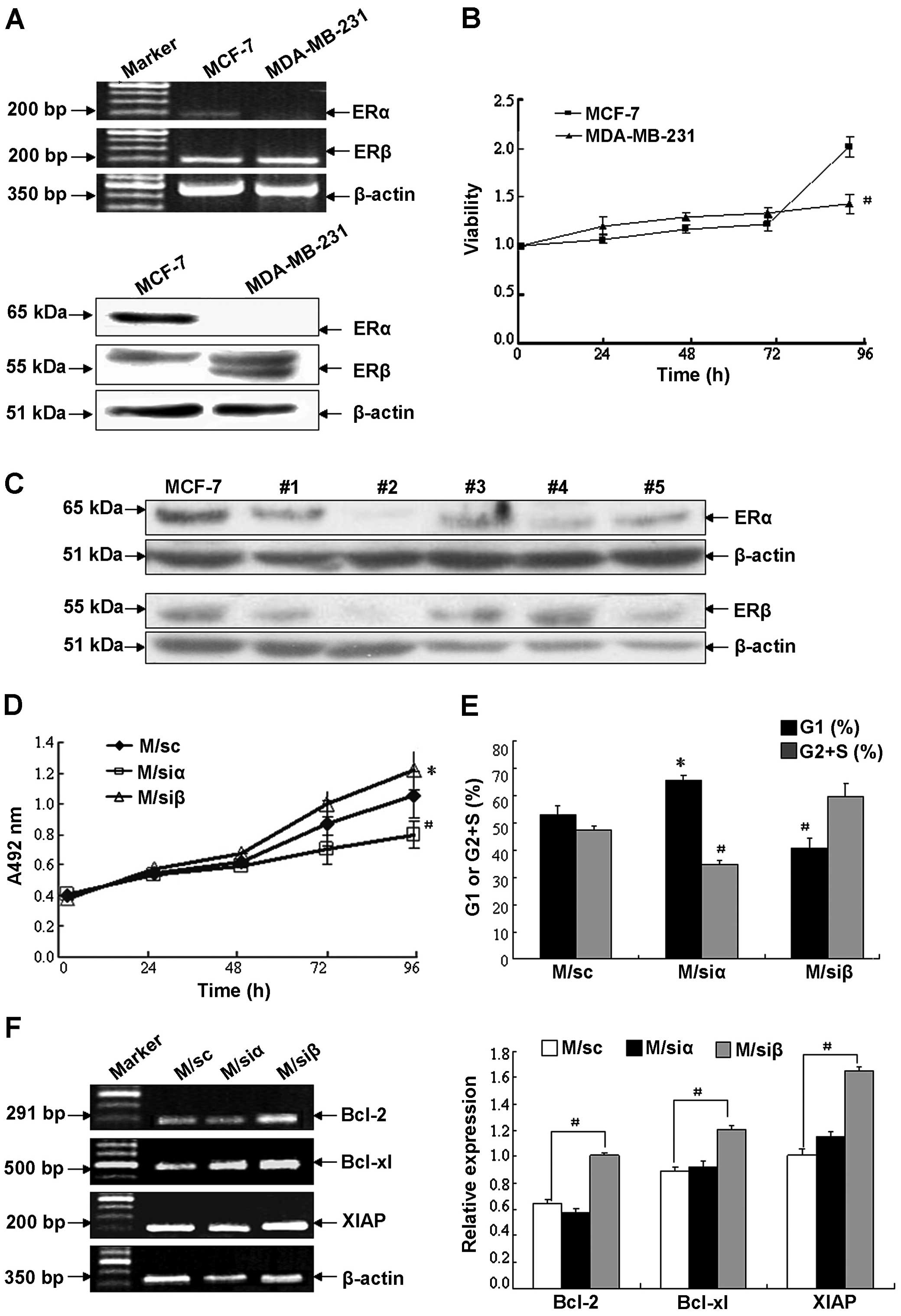 | Figure 1Regulation of cell growth in human
breast cancer cell lines by ER subtypes. (A) Detection of ERα and β
mRNA and proteins in human breast cancer cell lines MCF-7 and
MDA-MB-231 by semi-quantitative RT-PCR and western blot analysis.
The target gene expression was normalized using β-actin as control
of mRNA level. The target protein levels were normalized against
β-actin as control for variance in sample loading and transfer. The
experiment shown is representative of three separate experiments
with similar results. (B) Cell proliferation of MCF-7 and
MDA-MB-231 was assayed by MTT test. Data are shown as the mean ± SD
of three experiments with sextuple samples. #P<0.01
compared with MCF-7. (C) Confirmation of transfection efficacy as
well as expression level of ERα or β protein in MCF-7 cells and
their transfectants. MCF-7 cells were stably transfected with siRNA
targeting human ERα or β gene to silence corresponding ER subtype.
Stable MCF-7 transfected clones including #1–5 were obtained and
screened by western blotting with anti-ERα or anti-ERβ antibody.
Two stable transfected clones #2, with maximum knockdown of ERα or
ERβ protein ~77.7 and 68.3%, respectively (i.e., M/siα and M/siβ
cells), and scrambled siRNA-transfected MCF-7 cells (i.e., M/sc
cells) were chosen for subsequent studies. (D) Cell proliferation
of MCF-7 transfectants after culturing for 0, 24, 48, 72 or 96 h
was detected by MTT assay. Data are shown as the mean ± SD of two
separate experiments with sextuple samples. (E) Cell cycle
distribution in MCF-7 transfectants after culturing for 96 h were
assayed by flow cytometry. Black bar, G1 phase; gray bar, G2+S
phase. (F) mRNA levels of apoptosis suppressor genes in MCF-7
transfectants after culturing for 96 h were determined by
semi-quantitative RT-PCR method. White bar, M/sc; black bar, M/siα;
gray bar, M/siβ. The experiment shown is representative of three
independent experiments with similar results.
*P<0.05, #P<0.01 compared with the
control vector-tranfected MCF-7 cells. |
As previous studies reported, the expression of ER
subtypes may account for the confusion regarding the ERβ role in
cell proliferation. To determine whether expression of ER subtypes
correlates with cell proliferation, MCF-7 cells, which express both
ERα and β, were chosen to determine the impact of altering ERα and
β expression on cell growth. Stable clones with efficient knockdown
of ERα and β were made, two clones #2 with maximum knockdown of ERα
or ERβ ~77.7 or 68.3% (i.e., M/siα and M/siβ cells) and scrambled
siRNA-transfected MCF-7 cells (i.e., M/sc cells) were chosen for
subsequent studies (Fig. 1C). As
illustrated in Fig. 1D, compared
with the control cells (M/sc), cell proliferation decreased in
M/siα, which relatively overexpressed ERβ, whereas that in M/siβ,
which relatively overexpressed ERα, increased. The effect of ER
subtypes on cell proliferation was confirmed with FACS analysis of
synchronized MCF-7 cells (Fig. 1E).
Compared with the control cells, M/siα cells had a significant
increase in the G1 phase and decrease in the G2+S phase, whereas
M/siβ cells had a significant decrease in the G1 phase and increase
in the G2+S phase. These data suggest that knockdown of ERβ
promotes the proliferation of MCF-7 by altering cell cycle
distribution. We next analyzed the mRNA levels of apoptosis
suppressor genes (Fig. 1F).
Compared with the corresponding control cells, the expression of
Bcl-2, Bcl-xL and XIAP increased significantly in M/siβ cells
(P<0.01), whereas no difference was observed in M/siα cells.
Knockdown of ERβ in MCF-7 enhances
tumorigenicity and adhesion ability in vitro
To ascertain the biological effects of ER subtypes
on tumorigenicity of MCF-7, we assessed the anchorage-independent
growth of stable transfectants and the control cells in soft agar.
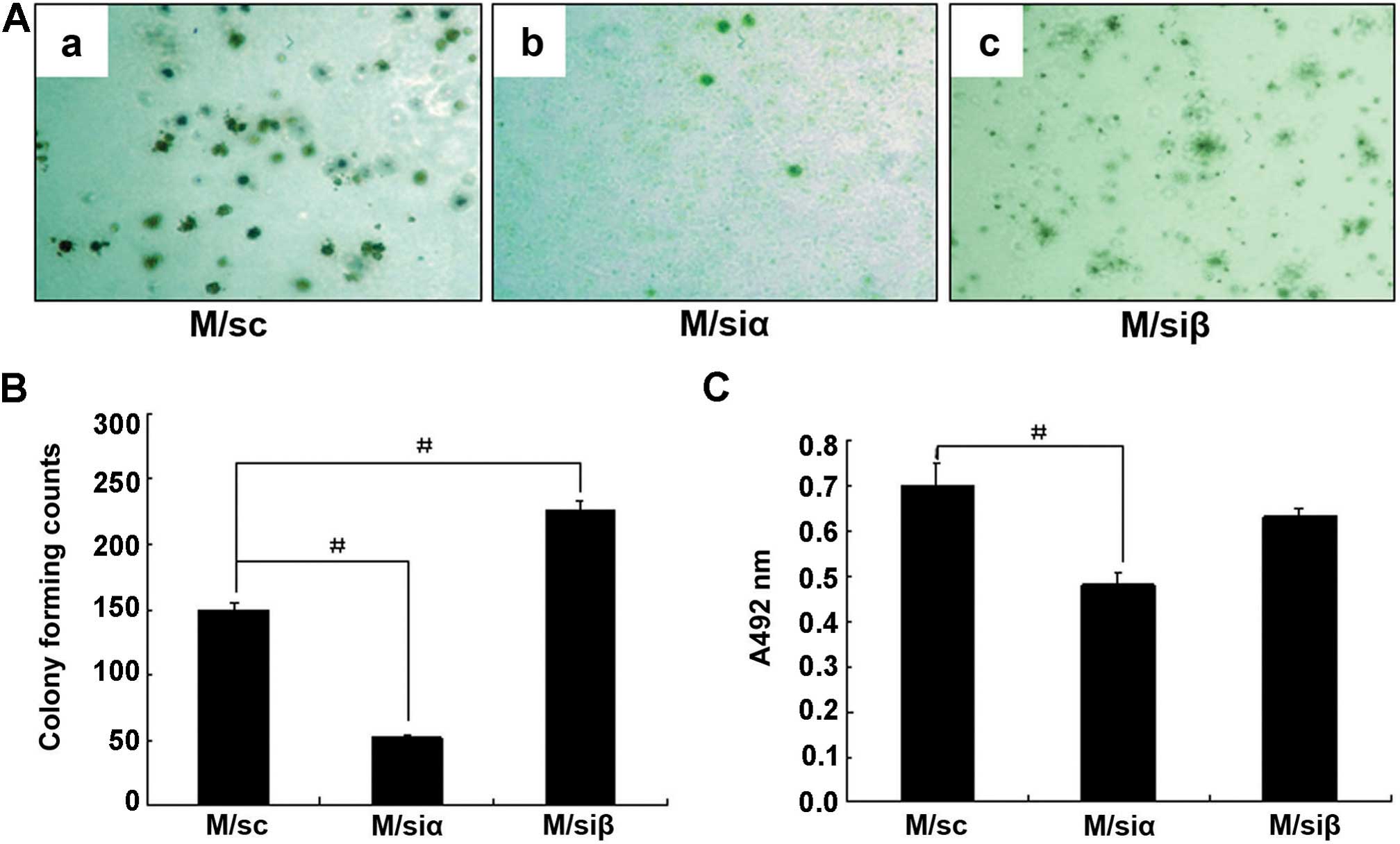
As shown in Fig. 2A and B, the
number of colonies formed by M/siα cells was significantly
decreased (~66.7% decrease) compared to those formed by the M/sc
cells. However, the number of colonies formed by M/siβ cells was
increased (~80.1% increase) compared to those formed by the M/sc
cells. These findings provide direct evidence that knockdown of ERβ
promotes tumorigenicity of MCF-7 cells.
Adhesion is a key function in the development of
metastasis. Accordingly, to examine whether ER subtype expression
correlates with adhesion ability of breast cancer cells, we
measured adhesion of MCF-7 stable transfectants and the control
cells. As shown in Fig. 2C,
compared with M/sc cells, ability of cell adhesion in M/si cells
decreased (P<0.01) markedly, while that in M/siβ cells had no
change. These results suggest that the increasement of relative
expression level of ERβ by knockdown of ERα may have an enhancing
effect on adhesion ability of MCF-7 cells, although direct
knockdown of ERβ plays only a small role.
Positive correlation exists between IFN-γ
at autocrine level and ERβ in MCF-7
Although some clinical-based studies indicate the
correlation between cytokine expression in tumor microenvironment
and ER levels in tumor tissues, the cell-based association study on
IFN-γ and IL-4 autocrine levels and ER subtypes in breast cancer
have not been previously reported. Accordingly, to examine whether
ER subtype expression correlates with autocrine IFN-γ and IL-4 in
human breast cancer cells, we first examined production of IFN-γ
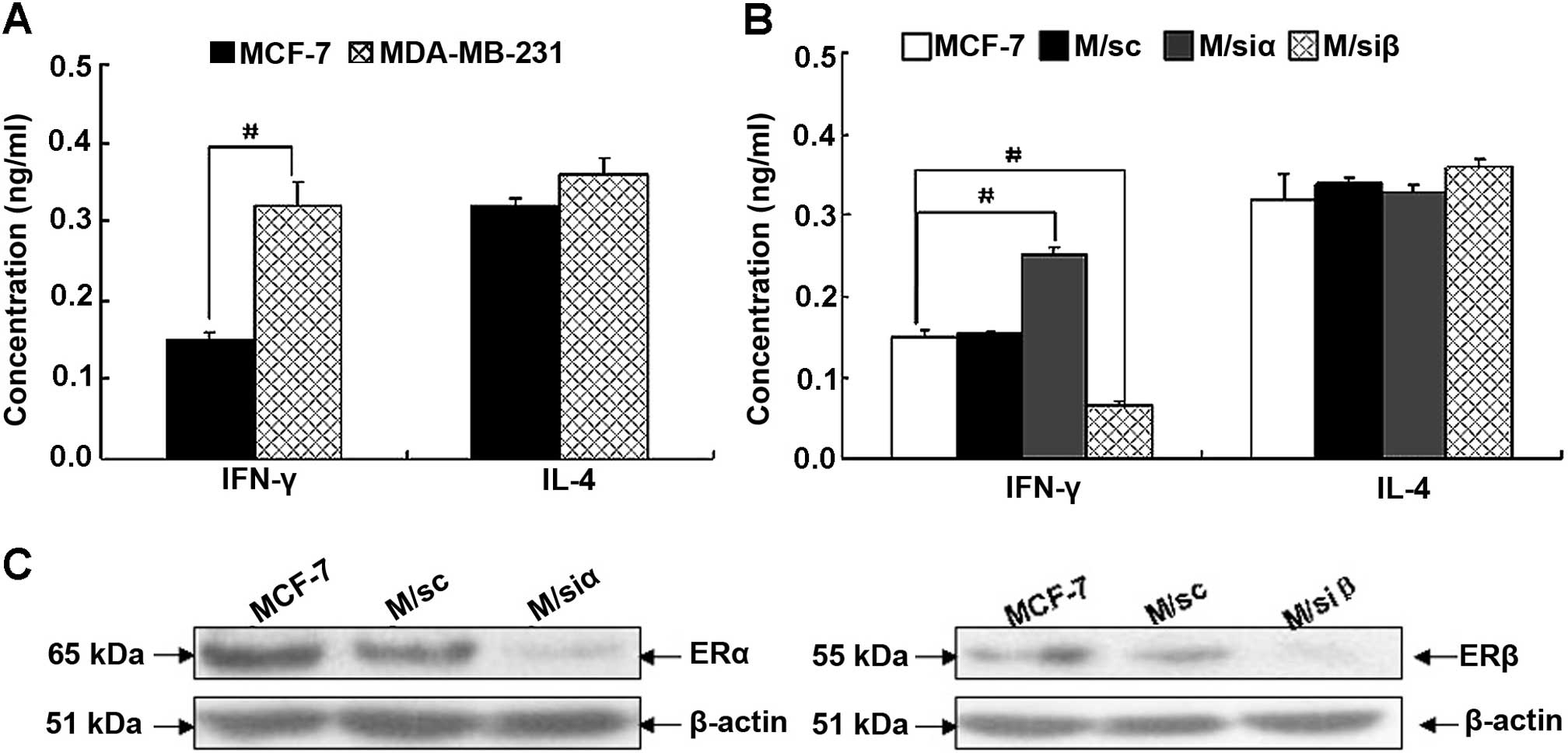
and IL-4 in MCF-7 and MDA-MB-231. As shown in Fig. 3A, IFN-γ production in MDA-MB-231 was
markedly higher than that in MCF-7. However, there was no
difference found in IL-4 secretion. The data illustrated in
Figs. 1A and 3A indicate that autocrine expression of
IFN-γ in breast cancer cells is inversely correlated with ERα;
whereas, positively correlated with ERβ. We then investigated the
secretion of IFN-γ and IL-4 in MCF-7 and its stable transfectants.
Consistent with the above results, compared with the corresponding
control cells, IFN-γ production in M/siα cells was significantly
increased (P<0.01); whereas, that in M/siβ cells was
significantly decreased (P<0.01) (shown in Fig. 3B and C). These results provide
further support that IFN-γ expression is differentially regulated
by ER subtype.
rhIFN-γ and rhIL-4 have effects on
biological behavior of MCF-7 cells
Tumor cells may produce a certain level of cytokines
to suppress antitumor immunity in tumor microenvironment. IFN-γ and
IL-4 have been shown to exert negative or positive effects on the
progression of cancer. Based on the above possibility, we first
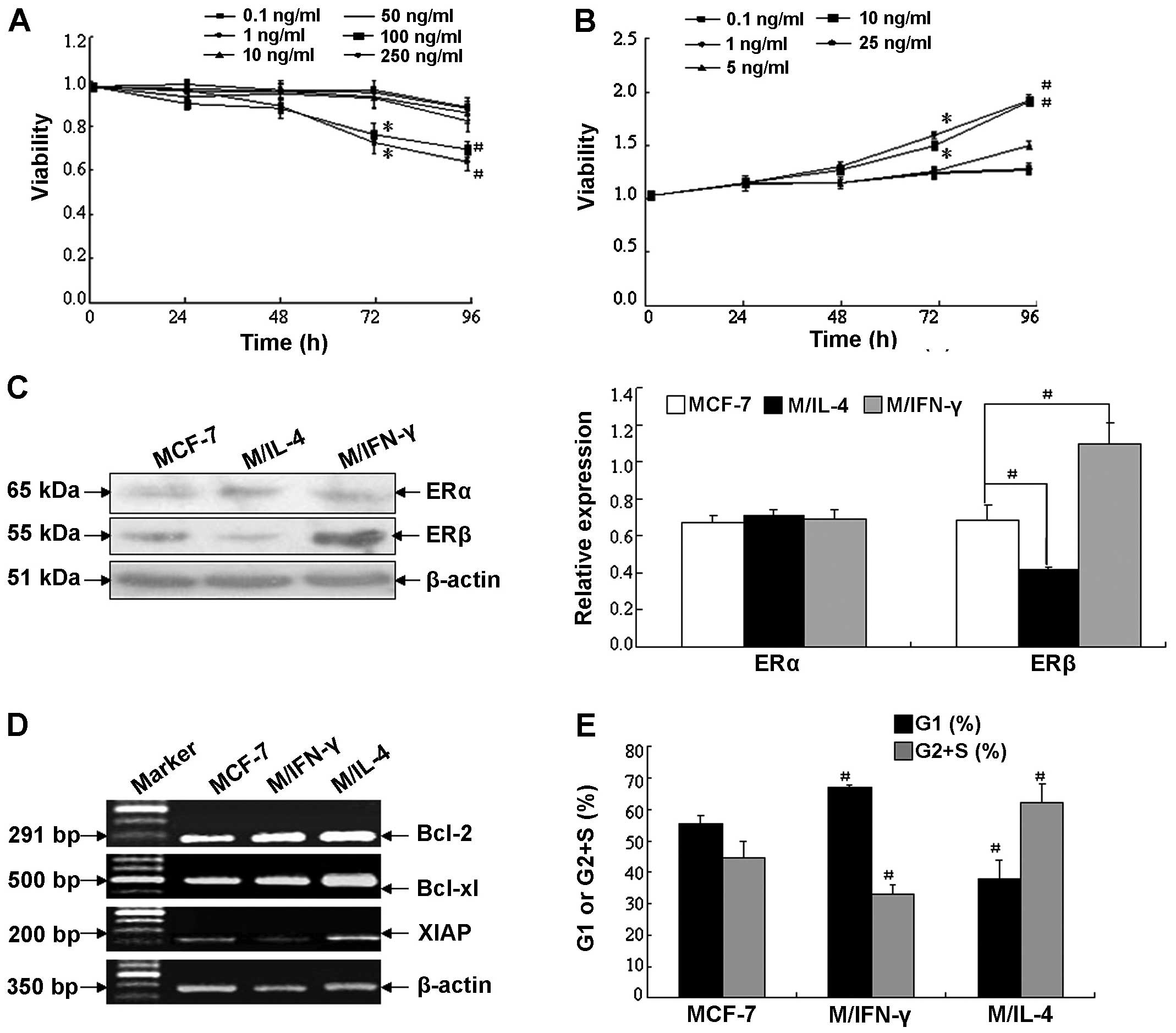
examined the impact of IFN-γ and IL-4 on MCF-7 cell growth
(Fig. 4A and B). As expected, IFN-γ
significantly inhibited proliferation at 100 and 250 ng/ml at 96-h
time point. In contrast, IL-4 significantly augmented cell growth
at 10 and 25 ng/ml at the same time point.
To explore whether IFN-γ and IL-4 were able to
modulate tumor cell growth by affecting the ER subtype expression
in breast cancer cells, we then investigated the protein levels of
ERα and ERβ in MCF-7, in the absence or presence of rhIFN-γ (100
ng/ml) or rhIL-4 (10 ng/ml) for 96 h, respectively. As shown in
Fig. 4C, rhIFN-γ markedly enhanced
expression of ERβ (~1.72-fold); whereas, rhIL-4 decreased
expression of ERβ (~0.57-fold), compared to the vehicle control,
whereas, neither IFN-γ nor IL-4 affected ERα protein level.
Furthermore, investigation illustrated that after treatment for 96
h, 100 ng/ml rhIFN-γ may induce cell apoptosis and inhibit
metastasis of human breast cancer cells; on the contrary, 10 ng/ml
rhIL-4 is able to promote tumor cell growth and metastasis. As
shown in Fig. 4D, rhIFN-γ decreased
mRNA level of the anti-apoptosis gene XIAP (~0.32-fold), but IL-4
increased mRNA level of the anti-apoptotic genes Bcl-2 and Bcl-xL
(~1.38- and 1.97-fold, respectively). Cell cycle distribution
analysis showed rhIFN-γ increased the percentage of G1 phase
(66.9%) and decreased the percentage of G2+S phase (33.1%) in MCF-7
compared with vehicle control (percentage of G1 and G2+S phase was
55.4 and 44.6%, respectively, P<0.01); whereas, IL-4 increased
the cell proportion in the G2+S phase (percentage of G1 and G2+S
phase was 37.8 and 62.2%, respectively, P<0.01) (Fig. 4E).
Furthermore, we ascertained whether IFN-γ or IL-4
had negative or positive effects on tumorigenicity and adhesion
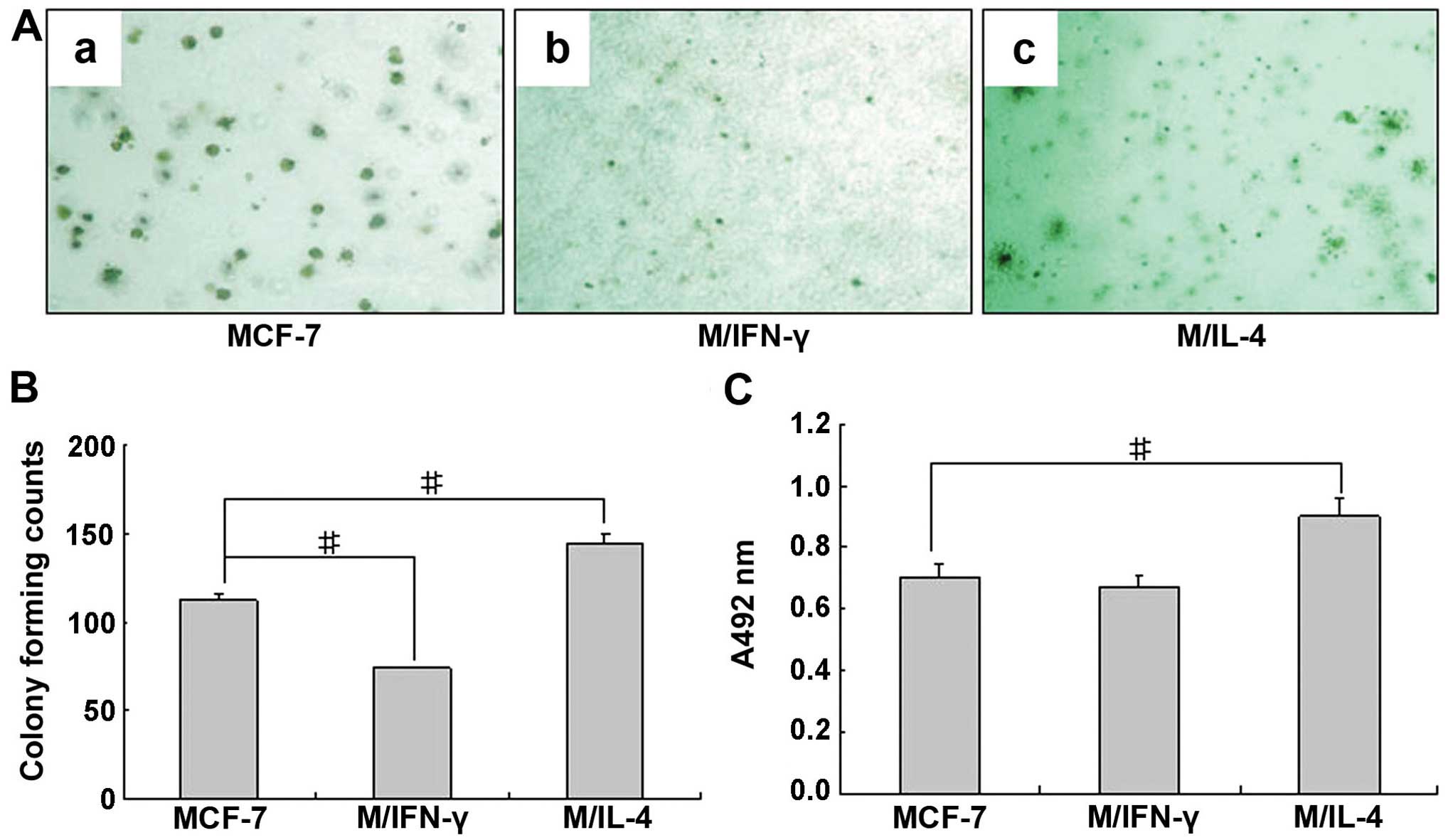
ability in MCF-7, respectively. As shown in Fig. 5A and B, compared with vehicle
control, the number of colonies formed in soft agar by MCF-7 in the
presence of IFN-γ significantly decreased (~33.9% decrease).
However, the number of colonies formed in the presence of rhIL-4
significantly increased (~28.6% increase). As illustrated in
Fig. 5C, compared with vehicle
control, adhesion ability of MCF-7 treated with rhIL-4 increased:
whereas, no difference was found regarding that of MCF-7 treated
with rhIFN-γ.
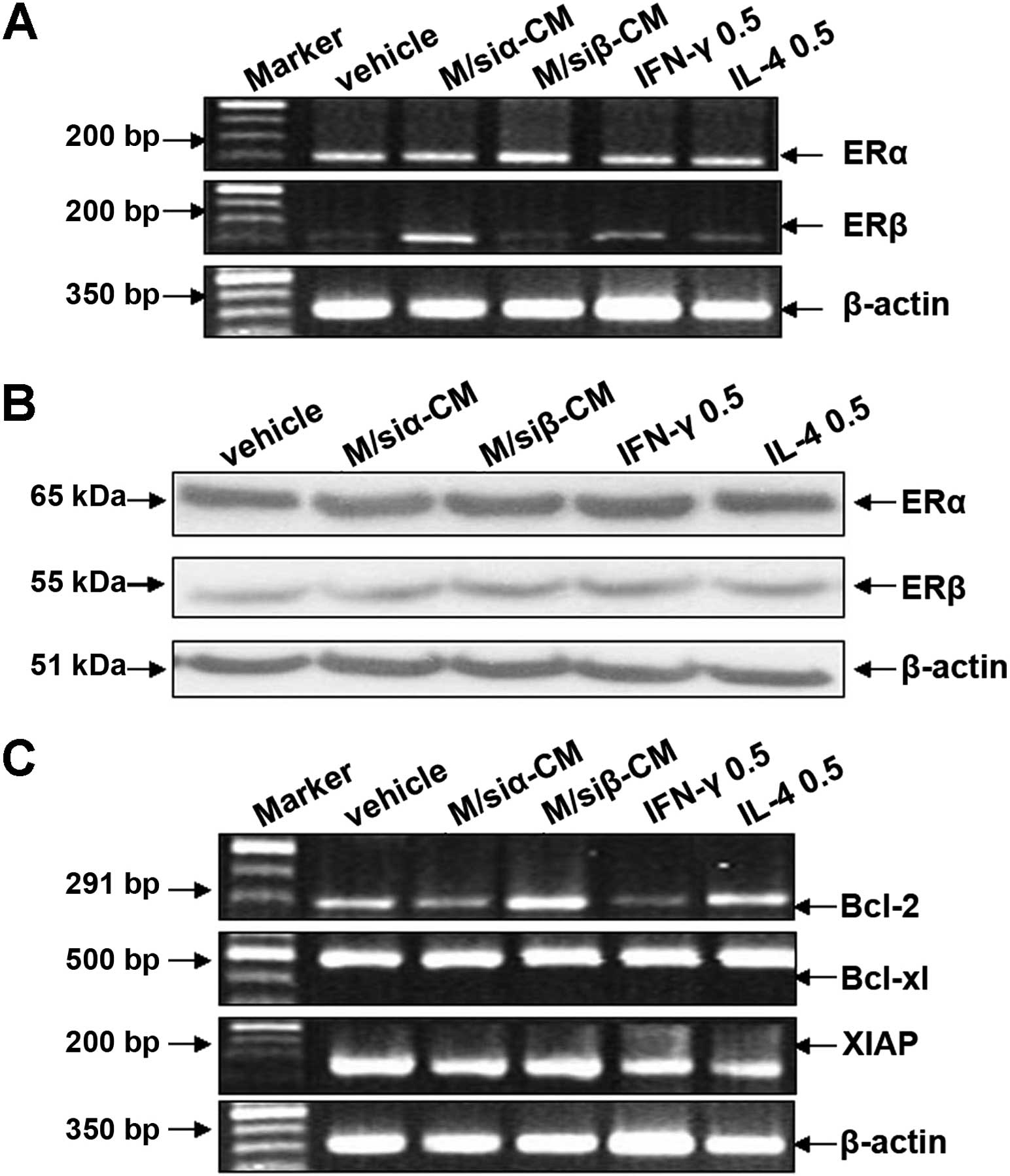
Autocrine level of IFN-γ affects mRNA
expression of ERβ and the apoptosis suppressor gene Bcl-2
Based on the above studies (Fig. 3A and B), we found the autocrine
production of IFN-γ and IL-4 by MCF-7 and MDA-MB-231 is below 1
ng/ml. We determined whether the level of IFN-γ and IL-4 affect the
expression of ER subtypes and the malignant behavior of breast
cancer cells. To address this problem, we used conditioned-medium
from M/siα or M/siβ, or 0.5 ng/ml rhIFN-γ or rhIL-4 to treat MCF-7
cells for 4 days. Then mRNA and protein expression of ER subtypes
and mRNA expression of the apoptosis suppressor genes were
detected. As illustrated in Fig.
6A, compared with control group, mRNA expression of ERβ in
MCF-7 treated with conditioned-medium from M/siα (in which the
level of IFN-γ was ~0.25 ng/ml), or 0.5 ng/ml rhIFN-γ was increased
(~1.86- and 1.28-fold, respectively); whereas, that in MCF-7
treated with conditioned medium of M/siβ (in which the level of
IL-4 was ~0.35 ng/ml) or 0.5 ng/ml rhIL-4 had no change. In
contrast, the treatment had no effect on mRNA expression of ERα. As
illustrated in Fig. 6B, there was
on difference in protein expression of ERα and β. In regards to
apoptosis suppressor genes, Fig. 6C
shows mRNA expression of Bcl-2 in MCF-7 cells treated with
conditioned-medium of M/siα or 0.5 ng/ml rhIFN-γ was decreased, in
contrast, that in MCF-7 treated with conditioned-medium of M/siβ or
0.5 ng/ml rhIL-4 was increased. These data suggest that autocrine
IFN-γ activity in breast cancer cells affects expression of ERβ and
Bcl-2.
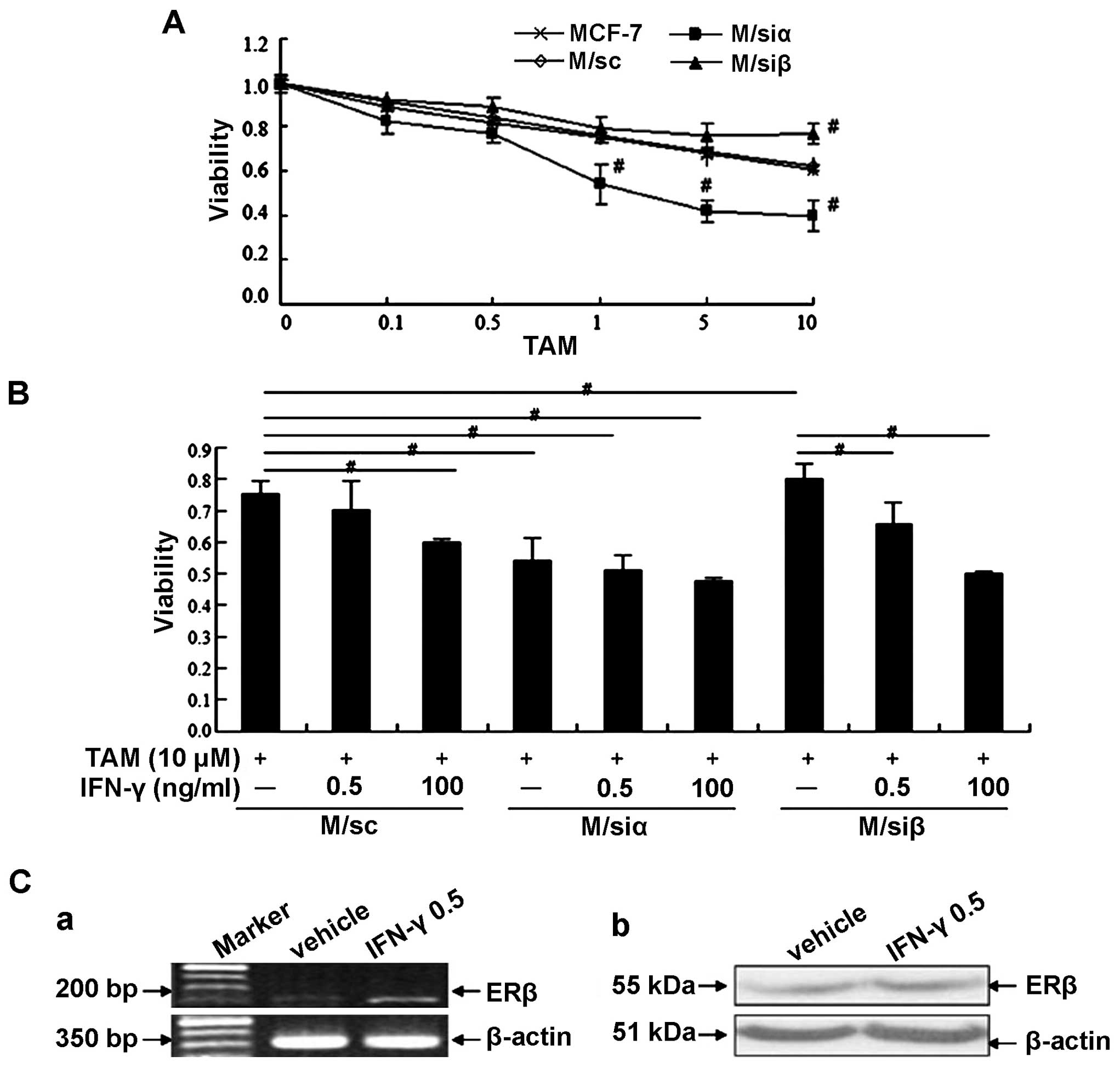
The pretreatment with IFN-γ increases the
sensitivity to TAM in MCF-7
ERα is regarded as a good biomarker for predicting
the efficiency of endocrine therapy of breast cancer. Previous data
show ERβ, in some cases such as ERα-negative breast cancer, may be
a better predictor to assess the effect of treatment. In the
present study, as shown in Fig. 7A,
we found knockdown of ERα in MCF-7 markedly enhanced the
sensitivity to TAM (1, 5 and 10 µM, compared with vehicle
control, P<0.01); whereas, knockdown of ERβ inversely inhibited
the sensitivity to TAM (10 µM, compared with vehicle
control, P<0.01). Further investigation showed that compared
with MCF-7 (both express ERα and β) and M/siα cells (express ERβ),
the sensitivity to TAM in M/siβ cells (express ERα) significantly
decreased. However, when we pretreated M/siβ cells with 0.5 ng/ml
rhIFN-γ (equivalent to autocrine level) or 100 ng/ml rhIFN-γ (level
based on MTT assay in Fig. 4A), the
sensitivity to TAM was significantly restored (Fig. 7B). As shown in Fig. 7C, 0.5 ng/ml rhIFN-γ promoted mRNA
expression of ERβ. These data suggest that autocrine IFN-γ may
upregulate the sensitivity to TAM of MCF-7 cells at least partially
via enhancing the expression of ERβ, although we only detected this
enhancement at mRNA level.
Discussion
A body of growing evidence shows that two distinct
estrogen receptors (ERs), ERα and ERβ, are critical in the
progression and prediction efficacy of antiestrogen therapy of
human breast cancer, and ER expression is a dynamic phenomenon and
is regulated by numerous factors, including cytokines in tumor
microenvironment. However, there are conflicting studies and it has
been unclear whether there is correlation between cytokines, ER
subtypes and malignant behavior of breast cancer cells. In the
present study, MCF-7, which expresses both ERα and ERβ, as a
representative breast cancer cell line, was used. Our results
indicated that ERβ inhibits the progression of breast cancer.
Within tumor microenvironment, IFN-γ and IL-4 may exert opposite
effects on malignant behavior. Moreover, we found that correlation
existed between expression of IFN-γ and ERβ. We also demonstrated
that autocrine IFN-γ activity in MCF-7 increases mRNA expression of
ERβ and enhances the sensitivity to tamoxifen (TAM).
Estrogens, particularly E2, have pleiotropic effects
on a wide variety of aspects through ERs, namely ERα and β. Since
its discovery in 1996, ERβ and its role in breast cancer is still
being explored. Most studies find ERα promotes progression of
breast cancer, but ERβ inhibits tumor formation due to its negative
modulation of ERα transcription. In contrast, Murphy et al
(32) suggested overexpression of
ERβ1 can increase proliferation, invasiveness and metastasis in a
ERα-negative breast cancer cell line (31–33).
The causes for the above differences include the presence of
multiple variant isoforms of ERβ, consideration on whether ERβ is
expressed alone or is co-expressed with ERα and methods of
determining ERβ expression (34–38).
In the present study, we used MCF-7 cells expressing both ERα and β
as the parental subject stably tranfected with ERα siRNA or ERβ
siRNA, and then analyzed the malignant behavior of MCF-7
transfectants. Antibodies used in western blotting recognized the
full length form of ERα and β (i.e. ERβ1) protein with 65 and 55
kD. Our studies suggested knockdown of ERβ in MCF-7 can promote
cell proliferation by increasing G1 phase distribution and
upregulation of mRNA levels of the apoptosis suppressor genes,
enhance the anchorage-independent growth ability and increase
adhesion. Autocrine production of IFN-γ by MCF-7 transfectants was
positively correlated with ERβ, but negatively with ERα. However,
there was no correlation found between autocrine IL-4 and ER
subtypes.
Previous studies have demonstrated cancer tissues
and cancer cells are another important resources of cytokines in
tumor microenvironment, besides tumor-infiltrating immune cells
(24–26,39).
IL-4 and IFN-γ, particularly produced by tumor or tumoral cells,
may re-educate cells in tumor niche to avoid host immunological
surveillance. It has been reported that IL-4 acts as an autocrine
growth factor in pancreatic cancer cells by promoting the
activation of AKT-1, Stat 3 and MAPK (25). In human primary prostate, breast and
bladder cancer cells, IL-4 induces upregulation of cFLIP and
Bcl-xL, which confer resistance to death receptor- and
chemotherapeutic drug-induced apoptosis (26). As for IFN-γ and other Th1 cytokines
(e.g. IL-12), it is reported that their production tends to be
suppressed, which often results in an inactivation of Th1 activity
(40). In the present study, we
illustrated a certain level of rhIFN-γ (i.e. 100 or 250 ng/ml) or
rhIL-4 (i.e. 10 or 25 ng/ml) may significantly inhibit or promote
breast cancer cell growth and metastasis in vitro,
respectively. Consistent with previous studies (29), we found correlation between IFN-γ
and IL-4 levels and ER expression. In the present study, we showed
that 100 ng/ml rhIFN-γ or 10 ng/ml rhIL-4 have direct positive or
negative effect on protein expression of ERβ, but not on ERα
expression.
In contrast, we noticed the autocrine production of
IFN-γ and IL-4, not only in breast cancer cell lines MCF-7 and
MDA-MB-231 but also in MCF-7 transfectants, was below 1 ng/ml
level. Accordingly, it was doubted whether autocrine IFN-γ or IL-4,
including conditioned medium from MCF-7 transfectants with
different ER subtypes expression and 0.5 ng/ml rhIFN-γ or rhIL-4
(equivalent to autocrine level), would affect ER expression in
MCF-7. We found autocrine IFN-γ markedly upregulated ERβ mRNA
expression, but did not affect ERβ protein expression, which may be
due to the insensitivity of antibodies recognizing ER in western
blotting. Our findings indicated that there may be a cycle in which
IFN-γ promotion by knockdown of ERα (i.e. relative overexpression
of ERβ) in MCF-7 would augment the level of ERβ further and vice
versa. Moreover, we also noted the apoptosis suppressor gene Bcl-2
was decreased by treatment with conditioned medium from M/siα or
0.5 ng/ml IFN-γ, which indicated the possible inhibition on cell
proliferation by autocrine IFN-γ.
As some previous studies show, ERα is regarded as a
good biomarker to assess the efficiency of endocrine therapy in
breast cancer for a long time. For this reason, patients with
ERα-positive tumors are treated with TAM, which blocks the action
of ERα or with aromatase inhibitors, which block the synthesis of
E2 (41,42). However, ERα seems not to act as a
predictor role invariably during the progression of breast cancer
(13,43,44).
Recently, data support ERβ may be a better predictor of TAM effect
than ERα in invasive breast cancers, which usually experience
reduced expression of ERα (45–48).
Consistent with the above studies, our results showed that
knockdown of ERα, i.e. relative overexpression of ERβ in MCF-7,
markedly enhanced the sensitivity to TAM, whereas knockdown of ERβ,
i.e. relative overexpression of ERα, inhibited TAM sensitivity.
These findings indicated that ERβ maybe a good predictor for
endocrine therapy in the cases of reduced expression of ERα during
the advanced stage of breast cancer. Compared with MCF-7 (both
express ERα and ERβ) and M/siα cells (express ERβ), the sensitivity
to TAM in M/siβ cells (express ERα) significantly decreased.
However, when we pretreated M/siβ cells with 0.5 ng/ml rhIFN-γ
(equivalent to autocrine level) or 100 ng/ml rhIFN-γ (chosen level
by MTT assay in Fig. 4A), the
sensitivity to TAM of these cells was significantly restored. These
data indicated that IFN-γ, including autocrine level, could restore
the sensitivity to TAM of M/siβ cells, at least, partially via
enhancing the expression of ERβ, although we only detected this
enhancement at mRNA level.
In conclusion, our findings provided evidence of
correlation between autocrine IFN-γ and ERβ in malignant
transformation and endocrine therapeutic prognosis. Furthermore, as
the IFN-γ exerted promotive activity on expression of ERβ and ERβ
is a protective factor to the progression of breast cancer, we
could deduce the combined treatment with IFN-γ and TAM may provide
a potentially benificial strategy in endocrine therapy of breast
cancer.
Acknowledgments
This study was supported by grants from the Tianjin
Municipal Science and Technology Commission (no.
15JCYBJC26000).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ziegler RG, Anderson WF and Gail MH:
Increasing breast cancer incidence in China: The numbers add up. J
Natl Cancer Inst. 100:1339–1341. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yue W, Yager JD, Wang JP, Jupe ER and
Santen RJ: Estrogen receptor-dependent and independent mechanisms
of breast cancer carcinogenesis. Steroids. 78:161–170. 2013.
View Article : Google Scholar
|
|
4
|
Mosselman S, Polman J and Dijkema R: ER
beta: Identification and characterization of a novel human estrogen
receptor. FEBS Lett. 392:49–53. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Williams C and Lin CY: Oestrogen receptors
in breast cancer: Basic mechanisms and clinical implications. E
Cancer Med Sci. 7:3702013.
|
|
6
|
Skliris GP, Munot K, Bell SM, Carder PJ,
Lane S, Horgan K, Lansdown MR, Parkes AT, Hanby AM, Markham AF, et
al: Reduced expression of oestrogen receptor beta in invasive
breast cancer and its re-expression using DNA methyl transferase
inhibitors in a cell line model. J Pathol. 201:213–220. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han SJ, Guo QQ, Wang T, Wang YX, Zhang YX,
Liu F, Luo YX, Zhang J, Wang YL, Yan YX, et al: Prognostic
significance of interactions between ER alpha and ER beta and lymph
node status in breast cancer cases. Asian Pac J Cancer Prev.
14:6081–6084. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Platet N, Cathiard AM, Gleizes M and
Garcia M: Estrogens and their receptors in breast cancer
progression: A dual role in cancer proliferation and invasion. Crit
Rev Oncol Hematol. 51:55–67. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pettersson K, Delaunay F and Gustafsson
JA: Estrogen receptor beta acts as a dominant regulator of estrogen
signaling. Oncogene. 19:4970–4978. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nilsson S, Mäkelä S, Treuter E, Tujague M,
Thomsen J, Andersson G, Enmark E, Pettersson K, Warner M and
Gustafsson JA: Mechanisms of estrogen action. Physiol Rev.
81:1535–1565. 2001.PubMed/NCBI
|
|
11
|
Fuqua SA, Schiff R, Parra I, Moore JT,
Mohsin SK, Osborne CK, Clark GM and Allred DC: Estrogen receptor
beta protein in human breast cancer: Correlation with clinical
tumor parameters. Cancer Res. 63:2434–2439. 2003.PubMed/NCBI
|
|
12
|
O'Neill PA, Davies MP, Shaaban AM, Innes
H, Torevell A, Sibson DR and Foster CS: Wild-type oestrogen
receptor beta (ERbeta1) mRNA and protein expression in
Tamoxifen-treated post-menopausal breast cancers. Br J Cancer.
91:1694–1702. 2004.PubMed/NCBI
|
|
13
|
Kerdivel G, Flouriot G and Pakdel F:
Modulation of estrogen receptor alpha activity and expression
during breast cancer progression. Vitam Horm. 93:135–160. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Stassi G, Todaro M, Zerilli M,
Ricci-Vitiani L, Di Liberto D, Patti M, Florena A, Di Gaudio F, Di
Gesù G and De Maria R: Thyroid cancer resistance to
chemotherapeutic drugs via autocrine production of interleukin-4
and interleukin-10. Cancer Res. 63:6784–6790. 2003.PubMed/NCBI
|
|
15
|
Mahmoud SM, Paish EC, Powe DG, Macmillan
RD, Grainge MJ, Lee AH, Ellis IO and Green AR: Tumor-infiltrating
CD8+ lymphocytes predict clinical outcome in breast
cancer. J Clin Oncol. 29:1949–1955. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Nagai S and Toi M: Interleukin-4 and
breast cancer. Breast Cancer. 7:181–186. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kmieciak M, Payne KK, Idowu MO, Grimes MM,
Graham L, Ascierto ML, Wang E, Wang XY, Bear HD and Manjili MH:
Tumor escape and progression of HER-2/neu negative breast cancer
under immune pressure. J Transl Med. 9:352011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen C, Guo L, Shi M, Hu M, Hu M, Yu M,
Wang T, Song L, Shen B, Qian L, et al: Modulation of IFN-γ receptor
1 expression by AP-2α influences IFN-γ sensitivity of cancer cells.
Am J Pathol. 180:661–671. 2012. View Article : Google Scholar
|
|
19
|
García-Tuñón I, Ricote M, Ruiz AA, Fraile
B, Paniagua R and Royuela M: Influence of IFN-gamma and its
receptors in human breast cancer. BMC Cancer. 7:1582007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Obiri NI, Siegel JP, Varricchio F and Puri
RK: Expression of high-affinity IL-4 receptors on human melanoma,
ovarian and breast carcinoma cells. Clin Exp Immunol. 95:148–155.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Obiri NI, Hillman GG, Haas GP, Sud S and
Puri RK: Expression of high affinity interleukin-4 receptors on
human renal cell carcinoma cells and inhibition of tumor cell
growth in vitro by interleukin-4. J Clin Invest. 91:88–93. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Varricchio F, Obiri NI, Haas GP and Puri
RK: Immunostaining of interleukin-4 receptor on human renal cell
carcinoma. Lymphokine Cytokine Res. 12:465–469. 1993.PubMed/NCBI
|
|
23
|
Toi M, Bicknell R and Harris AL:
Inhibition of colon and breast carcinoma cell growth by
interleukin-4. Cancer Res. 52:275–279. 1992.PubMed/NCBI
|
|
24
|
Todaro M, Lombardo Y, Francipane MG, Alea
MP, Cammareri P, Iovino F, Di Stefano AB, Di Bernardo C, Agrusa A,
Condorelli G, et al: Apoptosis resistance in epithelial tumors is
mediated by tumor-cell-derived interleukin-4. Cell Death Differ.
15:762–772. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Prokopchuk O, Liu Y, Henne-Bruns D and
Kornmann M: Interleukin-4 enhances proliferation of human
pancreatic cancer cells: Evidence for autocrine and paracrine
actions. Br J Cancer. 92:921–928. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Conticello C, Pedini F, Zeuner A, Patti M,
Zerilli M, Stassi G, Messina A, Peschle C and De Maria R: IL-4
protects tumor cells from anti-CD95 and chemotherapeutic agents via
up-regulation of antiapoptotic proteins. J Immunol. 172:5467–5477.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Todaro M, Zerilli M, Ricci-Vitiani L, Bini
M, Perez Alea M, Maria Florena A, Miceli L, Condorelli G, Bonventre
S, Di Gesù G, et al: Autocrine production of interleukin-4 and
interleukin-10 is required for survival and growth of thyroid
cancer cells. Cancer Res. 66:1491–1499. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Turgeon C, Gingras S, Carrière MC, Blais
Y, Labrie F and Simard J: Regulation of sex steroid formation by
interleukin-4 and interleukin-6 in breast cancer cells. J Steroid
Biochem Mol Biol. 65:151–162. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hong CC, Yao S, McCann SE, Dolnick RY,
Wallace PK, Gong Z, Quan L, Lee KP, Evans SS, Repasky EA, et al:
Pretreatment levels of circulating Th1 and Th2 cytokines, and their
ratios, are associated with ER-negative and triple negative breast
cancers. Breast Cancer Res Treat. 139:477–488. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li HZ, Wang Y, Gao Y, Shao J, Zhao XL,
Deng WM, Liu YX, Yang J and Yao Z: Effects of raf kinase inhibitor
protein expression on metastasis and progression of human
epithelial ovarian cancer. Mol Cancer Res. 6:917–928. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang B, Omoto Y, Iwase H, Yamashita H,
Toyama T, Coombes RC, Filipovic A, Warner M and Gustafsson JÅ:
Differential expression of estrogen receptor α, β1, and β2 in
lobular and ductal breast cancer. Proc Natl Acad Sci USA.
111:1933–1938. 2014. View Article : Google Scholar
|
|
32
|
Murphy LC and Leygue E: The role of
estrogen receptor-β in breast cancer. Semin Reprod Med. 30:5–13.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tonetti DA, Rubenstein R, DeLeon M, Zhao
H, Pappas SG, Bentrem DJ, Chen B, Constantinou A and Craig Jordan
V: Stable transfection of an estrogen receptor beta cDNA isoform
into MDA-MB-231 breast cancer cells. J Steroid Biochem Mol Biol.
87:47–55. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hou YF, Yuan ST, Li HC, Wu J, Lu JS, Liu
G, Lu LJ, Shen ZZ, Ding J and Shao ZM: ERbeta exerts multiple
stimulative effects on human breast carcinoma cells. Oncogene.
23:5799–5806. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fox EM, Davis RJ and Shupnik MA: ERbeta in
breast cancer -onlooker, passive player, or active protector?
Steroids. 73:1039–1051. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Skliris GP, Leygue E, Curtis-Snell L,
Watson PH and Murphy LC: Expression of oestrogen receptor-beta in
oestrogen receptor-alpha negative human breast tumours. Br J
Cancer. 95:616–626. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Monroe DG, Secreto FJ, Subramaniam M, Getz
BJ, Khosla S and Spelsberg TC: Estrogen receptor alpha and beta
heterodimers exert unique effects on estrogen- and
tamoxifen-dependent gene expression in human U2OS osteosarcoma
cells. Mol Endocrinol. 19:1555–1568. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Weitsman GE, Skliris G, Ung K, Peng B,
Younes M, Watson PH and Murphy LC: Assessment of multiple different
estrogen receptor-beta antibodies for their ability to
immunoprecipitate under chromatin immunoprecipitation conditions.
Breast Cancer Res Treat. 100:23–31. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jonsson P, Katchy A and Williams C:
Support of a bi-faceted role of estrogen receptor β (ERβ) in
ERα-positive breast cancer cells. Endocr Relat Cancer. 21:143–160.
2014. View Article : Google Scholar :
|
|
40
|
Chavey C, Bibeau F, Gourgou-Bourgade S,
Burlinchon S, Boissière F, Laune D, Roques S and Lazennec G:
Oestrogen receptor negative breast cancers exhibit high cytokine
content. Breast Cancer Res. 9:R152007. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Kidd P: Th1/Th2 balance: The hypothesis,
its limitations, and implications for health and disease. Altern
Med Rev. 8:223–246. 2003.PubMed/NCBI
|
|
42
|
Hartman J, Lindberg K, Morani A, Inzunza
J, Ström A and Gustafsson JA: Estrogen receptor beta inhibits
angiogenesis and growth of T47D breast cancer xenografts. Cancer
Res. 66:11207–11213. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Brueggemeier RW, Hackett JC and Diaz-Cruz
ES: Aromatase inhibitors in the treatment of breast cancer. Endocr
Rev. 26:331–345. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gutierrez MC, Detre S, Johnston S, Mohsin
SK, Shou J, Allred DC, Schiff R, Osborne CK and Dowsett M:
Molecular changes in tamoxifen-resistant breast cancer:
Relationship between estrogen receptor, HER-2, and p38
mitogen-activated protein kinase. J Clin Oncol. 23:2469–2476. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gallo D, De Stefano I, Grazia Prisco M,
Scambia G and Ferrandina G: Estrogen receptor beta in cancer: An
attractive target for therapy. Curr Pharm Des. 18:2734–2757. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Honma N, Horii R, Iwase T, Saji S, Younes
M, Takubo K, Matsuura M, Ito Y, Akiyama F and Sakamoto G: Clinical
importance of estrogen receptor-beta evaluation in breast cancer
patients treated with adjuvant tamoxifen therapy. J Clin Oncol.
26:3727–3734. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Pitta CA, Papageorgis P, Charalambous C
and Constantinou AI: Reversal of ER-β silencing by chromatin
modifying agents overrides acquired tamoxifen resistance. Cancer
Lett. 337:167–176. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Razandi M, Pedram A, Jordan VC, Fuqua S
and Levin ER: Tamoxifen regulates cell fate through mitochondrial
estrogen receptor beta in breast cancer. Oncogene. 32:3274–3285.
2013. View Article : Google Scholar
|