Introduction
Malignant gliomas are characterized by aggressive
tumor growth with a mean patient survival of 15–18 months and
frequently develop resistance to temozolomide (TMZ) which is the
first-line treatment for patients with high-grade gliomas (1,2).
Although TMZ has been well demonstrated to effectively prolong the
survival of patients with brain tumors in clinical application,
unfortunately, glioma cells showed resistance to TMZ in certain
cases (3–6). Therefore, current therapy is far from
satisfactory, indicating the need for new therapeutic agents and
approaches which can prolong the survival of glioma patients.
Recently, extensive study has been conducted to
evaluate the therapeutic efficacy and safety of natural compounds
for treating cancer. Rhein (4,5-dihydroxyanthraquinone-2-carboxylic
acid) is a natural product derived from the rhizome of rhubarb
which has been used medicinally in China for more than 1,000 years
(7). Rhein possesses diverse
biological properties such as antioxidant (8–10),
anti-inflammatory (11,12), antiviral (13), antibacterial (14), antifungal (15), anti-allergic (16) and anticancer (17–24).
The reported mechanism of the antitumor activity of rhein in cancer
cells is due to its ability to induce apoptosis and/or cell cycle
arrest in corresponding cancer cells (20,21).
Although rhein has many pharmacological effects, due to its
inability to dissolve in water, the use of rhein in the clinic is
limited.
Our previous studies showed that rhein lysinate
(RHL), the salt of rhein and lysine that is easily dissolved in
water, inhibited the proliferation of breast (25) and ovarian cancer (26), hepatocellular carcinoma (26), cervical cancer (27,28),
and lung carcinoma cells (29), and
human umbilical vein endothelial cells (HUVECs) (28). However, the IC50 was
found to be higher in normal cells, meaning that the drug can be
used to prevent the proliferation of cancer cells at a lower
concentration, without a strong effect on normal cells. Yet, its
effect on human glioma is still unknown.
In the present study, human glioma U87 cells and a
xenograft model in BALB/c nude mice were used to examine the
antitumor activity of RHL against human glioma, supporting the
potential use of rhein as an anti-glioma medicinal agent.
Materials and methods
Chemicals and reagents
Rhein (purity, 98%) was purchased from Nanjing
Qingze Medicine Ltd. (Nanjing, Jiangsu, China), while lysine was
purchased from Beijing Solarbio Science and Technology Co.
(Beijing, China). RHL was produced at the Department of Oncology of
the Institute of Medicinal Biotechnology, the Chinese Academy of
Medical Sciences (patent no. 200810089025.8).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and dimethylsulfoxide (DMSO) were obtained from Sigma-Aldrich
(Shanghai, China). Dichlorodihydrofluorescein-diacetate (DCFH-DA)
was obtained from Molecular Probes (Eugene, OR, USA). Antibodies
targeting Bcl-2, BAX, Bim, cyclin D and β-actin were purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). Secondary
antibodies against rabbit or mouse IgG were purchased from Cell
Signaling Technology (Danvers, MA, USA). The prestained protein
marker p7708V was purchased from New England Biolabs Ltd. (Beijing,
China). All other chemicals were of standard analytical grade.
Cell culture
The human glioma U87 cell line was cultured in
RPMI-1640 medium (Gibco-BRL, Grand Island, NY, USA) supplemented
with 10% heat-inactivated fetal bovine serum (Sigma Chemical Co.,
St. Louis, MO, USA), 100 U/ml penicillin and 100 μg/ml
streptomycin at 37°C in a humidified atmosphere containing 5%
CO2. The human glioma U87 cell line was obtained from
the Cell Center of the Institute of Basic Medical Sciences, Chinese
Academy of Medical Sciences and Peking Union Medical College
(Beijing, China).
In vivo therapeutic effects
Fourteen female BALB/c nude mice (20±2 g) obtained
from Vital River Laboratories (Beijing, China), at the age of 4–6
weeks were used for human glioma U87 xenografts. Mice were
maintained in a temperature-controlled room (22±2°C) with a 12-h
light/12-h dark cycle and a relative humidity of 40–60%. The mice
were given ad libitum access to food and water. All animal
experiments were approved by the Institutional Animal Care and Use
Committee of Beijing Normal University. U87 tumors for implantation
were initially grown by injection of U87 cells at a dose of
5×106 cells/mouse in two female BALB/c nude mice. A
tumor piece of 2–3 mm in diameter was implanted subcutaneously into
each experimental animal. After 9 days of tumor growth, the animals
were randomly divided into groups (n=6) in a manner that minimized
the difference in tumor size between the groups. Each animal
received 200 μl of either phosphate-buffered saline (PBS)
(vehicle control) or RHL by intraperitoneal injection every other
day for a consecutive 14 days. At the end of the experiment, the
body and tumor weight were measured.
Cell proliferation assay
Cell proliferation assays were performed using the
MTT method, according to the manufacturer's instructions. The cells
were seeded into 96-well plates (Costar, Cambridge, MA, USA) with
3,000 cells/well. Subsequent to overnight incubation, triplicate
wells were treated with various concentrations of RHL (0, 20, 40,
80 and 160 μmol/l) for 48 h. Next, 20 μl of MTT
solutions (5 mg/ml in PBS) were added to each well and incubated
for 4 h at 37°C. The MTT formazan was dissolved in 150 μl
DMSO and the absorbance was measured with a microplate reader
(Multiskan MK3; Thermo Labsystems, Waltham, MA USA) at a wavelength
of 570 nm.
Cell growth curve
Cells were plated into 96-well plates at a density
of 500 cells/well. Six plates were seeded with cells. Triplicate
wells/plate were treated with various concentrations of RHL (0, 20,
40, 80 and 160 μmol/l). One plate was assayed by MTT method
as described above each day until the sixth day. The cell growth
curve for various concentrations of RHL was constructed.
Detection of reactive oxygen species
(ROS) by DHCF-DA
Intracellular hydrogen peroxide levels were
monitored by fluorescence microscopy and flow cytometry after
staining with DCFH-DA. Cells were plated at 250,000 cells/flask.
Following a 24-h incubation period, the cells were treated with
various concentrations of RHL (0, 20, 40, 80 and 160
μmol/l). Forty-eight hours later, DCFH-DA (10 μmol/l)
was then added to each flask. After a 1-h incubation period, the
cells were monitored using an Olympus inverted fluorescence
microscope (Tokyo, Japan), and were subjected to flow cytometric
analysis with FACSCalibur and CellQuest software (Becton-Dickinson,
Franklin Lakes, NJ, USA). The percentage of cells displaying
increased dye uptake was used to reflect an increase in ROS
levels.
FITC-Annexin V/PI apoptosis assay
Cells were plated at 250,000 cells/flask. Following
a 24-h incubation period, the cells were treated with various
concentrations of RHL (0, 20, 40, 80 and 160 μmol/l).
Forty-eight hours later, the cells were collected and resuspended
in 200 μl binding buffer. Then, 10 μl FITC-labeled
enhanced Annexin V and 10 μl propidium iodide (PI) were
added. Upon incubation in the dark (15 min at room temperature or
30 min at 4°C), the samples were diluted with 300 μl binding
buffer. Cells were subjected to flow cytometric analysis with
FACSCalibur and CellQuest software.
Western blot analysis
Cells were harvested and washed with PBS. The whole
cellular extracts were prepared by incubating cells on ice in a
lysis buffer and a cocktail of phosphatase inhibitors (Roche,
Indianapolis, IN, USA). The cell lysates were cleared by
centrifugation at 12,000 × g for 20 min. Protein concentrations
were determined by Bradford assay. Equal amounts of lysate (40
μg) were resolved by SDS-PAGE and transferred to
polyvinylidene difluoride membranes (Millipore Corp., Bedford, MA,
USA). The membranes were blocked in TBST containing 5% non-fat skim
milk at room temperature for 2 h and probed with primary antibodies
overnight at 4°C. Then the membranes were blotted with an
appropriate horseradish peroxidase-linked secondary antibody (Santa
Cruz Biotechnology). Proteins were visualized using enhanced
chemiluminescence western blotting detection reagents (Amersham
Pharmacia Biotech, Inc., Piscataway, NJ, USA).
Statistical analysis
Results are expressed as the means ± SD. Treatment
effects were compared using one-way ANOVA and differences between
the means were considered to indicate a statistically significant
result when P<0.05.
Results
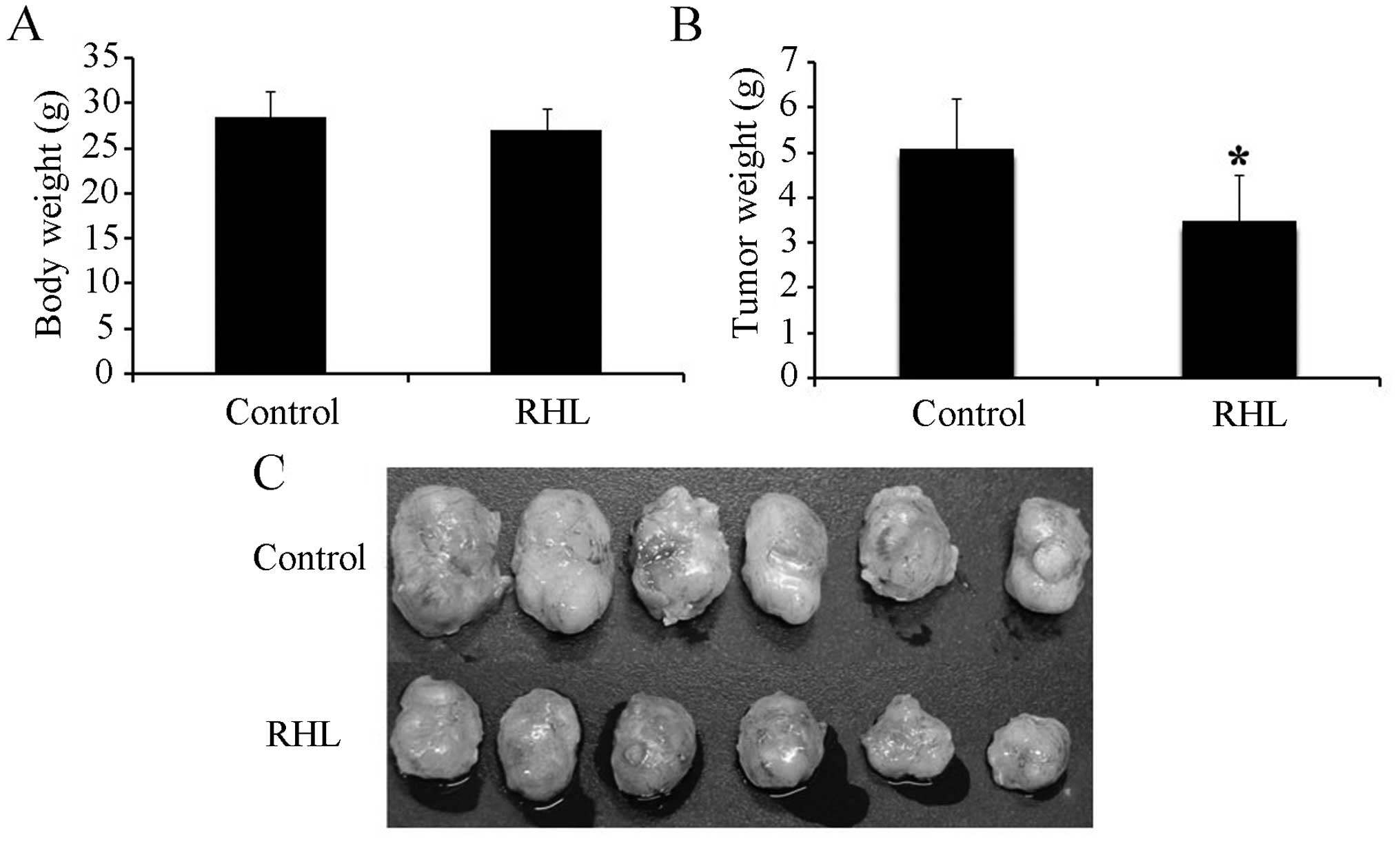
Inhibition of human glioma U87 xenograft
growth in BALB/c nude mice
Treatment was started on day 9 after tumor
transplantation. RHL was administered by intraperitoneal injection
at doses of 50 mg/kg every other day for consecutive 14 days.
Control mice were administered PBS vehicle only. The body weight of
the animals showed no significant differences between the control
and treated groups (Fig. 1A). The
growth of tumors in the RHL-treated BALB/c nude mice was
significantly suppressed compared with the controls (Fig. 1B and C). Treatment with RHL
inhibited the growth of human glioma U87 xenografts by 31.9%. These
findings suggest that RHL at a well-tolerated dose markedly
inhibited tumor growth.
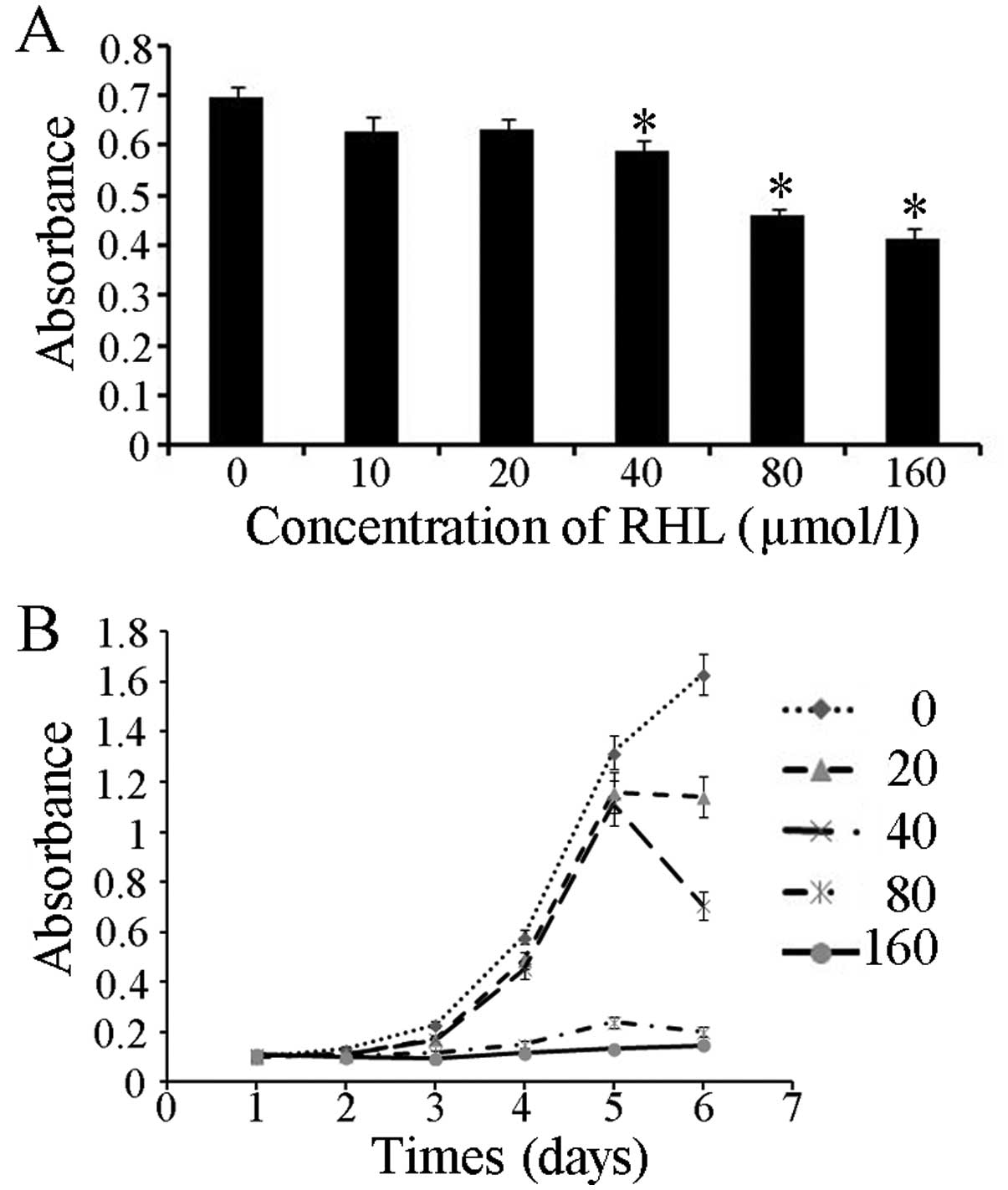
Growth inhibition of RHL in human glioma
U87 cells
The growth inhibitory effect of RHL on human glioma
U87 cells was examined with MTT assay. Cells were cultured for 48 h
(Fig. 2A) in the presence of
various concentrations of RHL. The U87 cells showed a decreased
cell proliferation in a dose-dependent manner after treatment with
RHL. In addition, RHL (160 μmol/l) inhibited the
proliferation of the U87 cells by 40% at 48 h. Compared with the
control group, RHL at 20 and 40 μmol/l inhibited U87 cell
proliferation on day 6. Nevertheless, RHL at 80 and 160
μmol/l inhibited U87 cell proliferation from initial
administration (Fig. 2B)
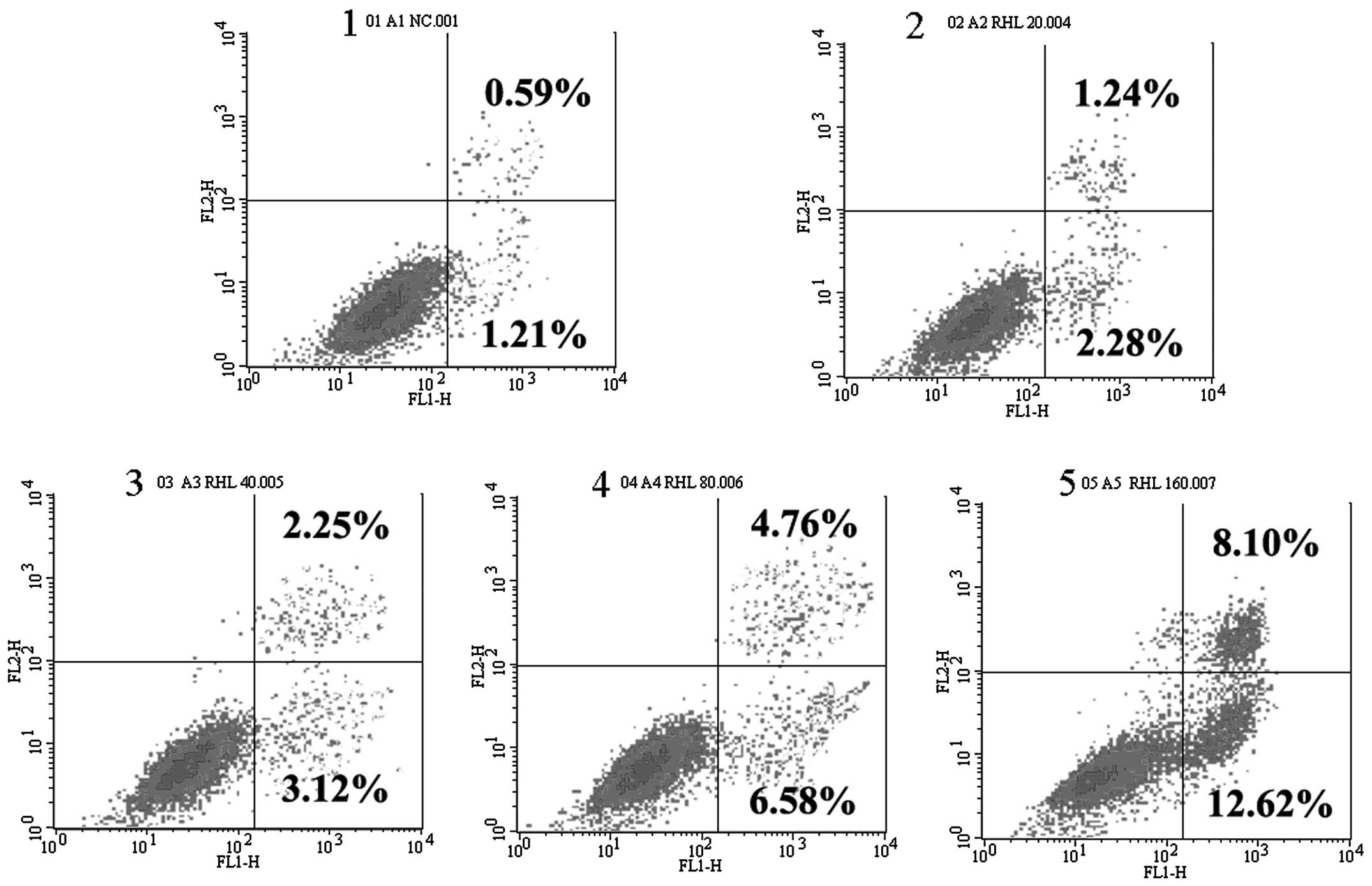
Induction of apoptosis by RHL in human
glioma U87 cells
Induction of apoptosis by RHL was confirmed by
FITC-Annexin V/PI staining. RHL at 80 μmol/l induced
apoptosis in the glioma U87 cells. The ratio of apoptosis was
significantly enhanced when cells were incubated with 160
μmol/l RHL for 48 h. This suggested that apoptosis was the
predominant mode of RHL-induced cell death (Fig. 3).
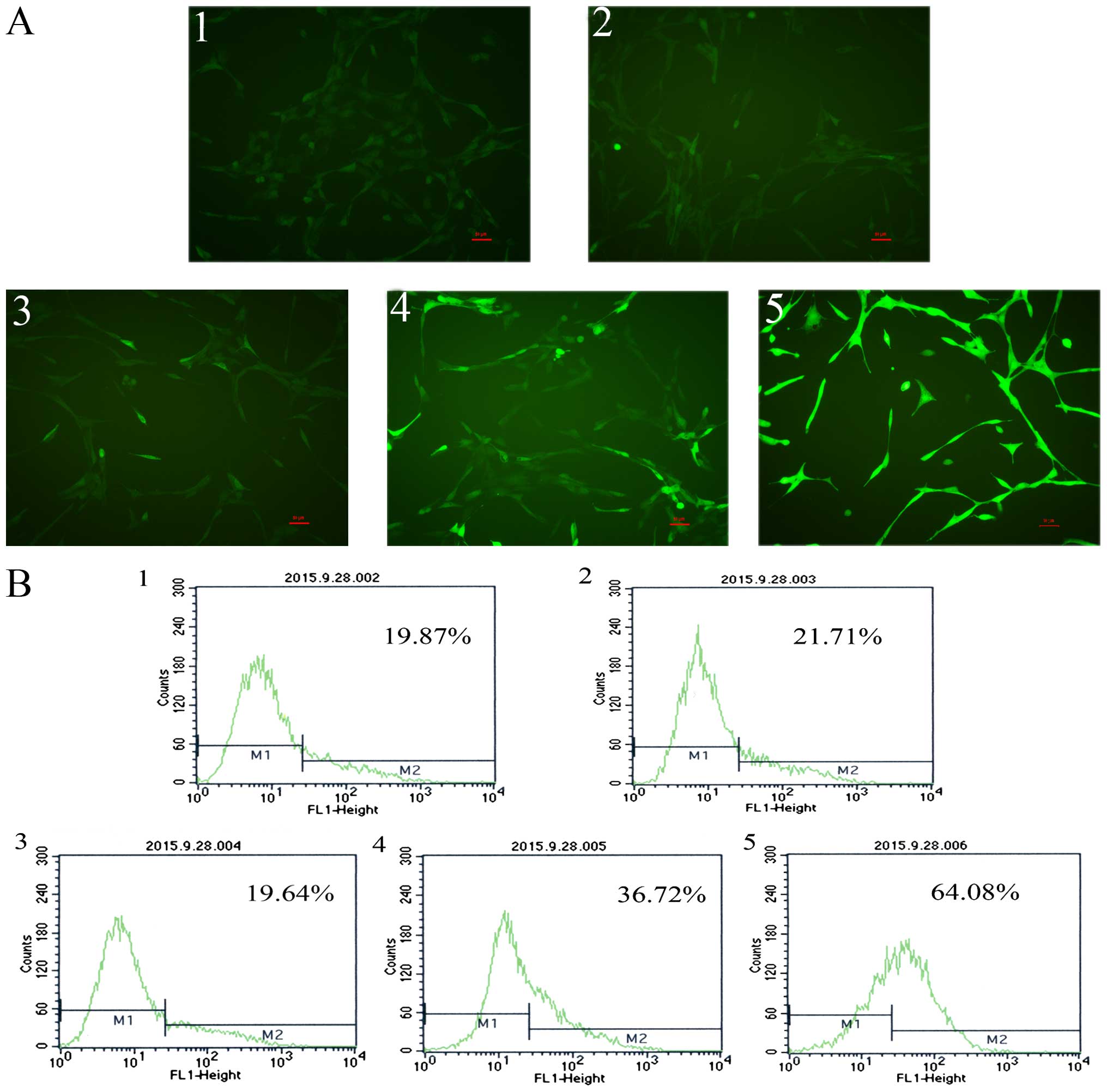
Induction of ROS production by RHL in
human glioma U87 cells
Induction of ROS production by RHL was confirmed by
DCFH-DA staining and detected by fluorescence microscopy and flow
cytometry. Fluorescence intensity/cell was significantly enhanced
when cells were treated with RHL for 48 h in dose-dependent manner.
This suggested that RHL induced ROS production in a dose-dependent
manner in glioma U87 cells (Fig.
4).
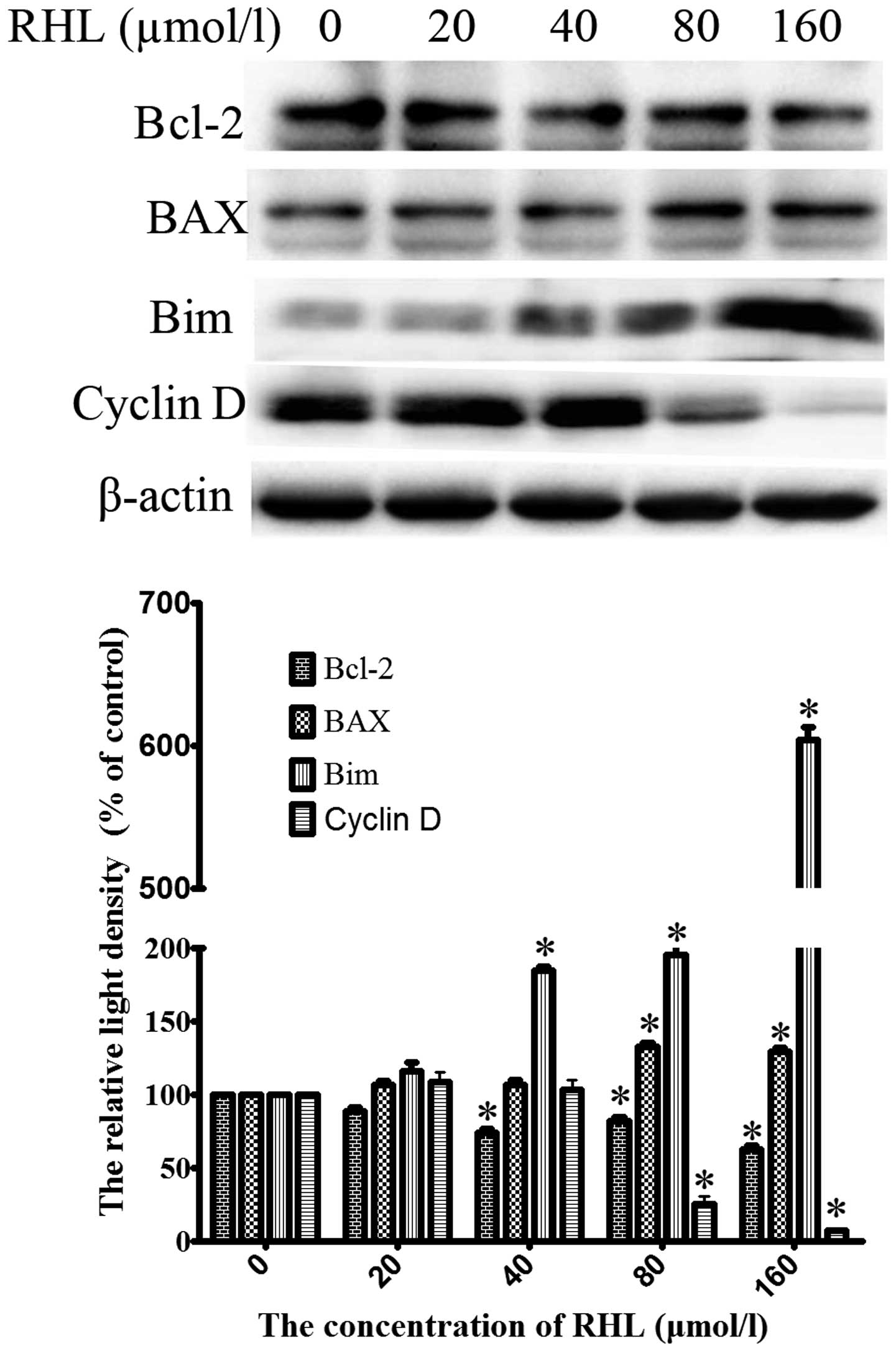
Downregulation of Bcl-2 and cyclin D and
upregulation of BAX and Bim in the RHL-induced apoptosis in glioma
U87 cells
In order to investigate the mechanism of RHL against
human glioma U87 cells, we examined whether RHL treatment modulates
the levels of apoptosis-associated and cell cycle-associated
proteins. As shown in Fig. 5,
expression of Bcl-2 was markedly downregulated by RHL treatment in
the U87 cells, whereas the levels of BAX and Bim were increased. In
addition, we also examined the expression of cyclin D. The level of
cyclin D was decreased after RHL treatment.
Discussion
Malignant glioma is one of the most deadly human
malignancies worldwide and its incidence has increased in recent
years (30). In spite of the
progress in the prognosis and treatment of glioma, the dismal
outcome has not improved substantially over the last three decades.
The survival of glioma patients is still quite short, particularly
when invasion and metastasis occur, even when aggressive surgical
resection, chemotherapy, and radiotherapy are carried out.
Temozolomide (TMZ) is a well-tolerated orally bioactive alkylating
agent used in glioma patients which has been adopted as the
first-line treatment in patients with high-grade gliomas. Although
TMZ has been well demonstrated to effectively prolong the survival
of brain tumor patients in clinical application (1,2),
unfortunately, glioma cells show resistance to TMZ in some cases
(3–6). Therefore, new therapeutic agents and
approaches are highly required for prolonging the survival of
glioma patients.
Our previous studies showed that rhein lysinate
(RHL), the salt of rhein and lysine that is easily dissolved in
water, inhibited the proliferation of breast (25), ovarian cancer (26), hepatocellular carcinoma (26), cervical cancer (27,28),
and lung carcinoma cells (29) and
human umbilical vein endothelial cells (28). However, the IC50 value is
higher in normal cells, meaning that the drug can be used to
prevent the proliferation of cancer cells at a lower concentration,
without a strong effect on normal cells. In addition, the in
vivo and in vitro therapeutic efficacy of RHL on human
glioma have not been evaluated. As known, human glioma U87
xenografts are useful models for testing the therapeutic effects of
anti-tumor agents in vivo (31–33).
Human glioma U87 cells and a xenograft model in BALB/c nude mice
were used to examine the antitumor activity of RHL against human
glioma in the present study. RHL exerted a significant inhibitory
effect on the growth of human glioma U87 xenografts in BALB/c nude
mice. There was no significant body weight loss in the treated
groups compared with the control group. All animals survived the
duration of the experiment. The results indicate that RHL shows a
high efficacy against human glioma U87 cells.
RHL showed a potent cytotoxic effect on U87 cells.
In the cell proliferation assays, RHL (160 μmol/l) inhibited
U87 cell proliferation by 40% at 48 h (Fig. 2A). In the cell growth curve assays,
RHL at concentrations of 80 and 160 μmol/l inhibited U87
cell proliferation from the initial administration (Fig. 2B). In the cell apoptosis assays, RHL
displayed a highly potent apoptosis-inducing effect on the U87
cells. It is evident that the induction of apoptosis appeared to be
the predominant mode of RHL-induced cell death particularly in the
high dose-RHL group (160 μmol/l).
Excessive intracellular ROS production induced by a
toxicant within the mitochondria or cytoplasm can damage many types
of biological macromolecules such as membrane lipids, DNA and
enzymes. Furthermore, ROS induce mitochondrial depolarization and
permeability transition (34,35).
In the present study, the DCF fluorescence assay showed that the
intracellular ROS level in the RHL-treated cells was higher than
that of the control cells. This observation suggests that RHL
exposure caused oxidative stress, mitochondrial permeability
transition, and apoptosis of U87 cells. Therefore, excessive ROS
induction by RHL exposure may be a key early factor in the cellular
damage and apoptosis of U87 cells.
As known, members of the Bcl-2 protein family act as
key regulators of cellular apoptosis and are important determinants
of cellular sensitivity or resistance to chemotherapy drugs
(36–38). Overexpression of Bcl-2, an
anti-apoptotic member of this family, is commonly observed in human
cancers, and Bcl-2 overexpression correlates with chemoresistance
in this disease. The anti-apoptotic protein, Bcl-2, has been
associated with inhibition of apoptosis and cell survival
mechanisms. The Bax and Bim proteins are pro-apoptotic members of
this family, and their increased expression is often associated
with increased apoptosis in target cells (39,40).
Next, we studied the molecular mechanism of apoptosis induced by
RHL in human glioma U87 cells. As shown in Fig. 5, when the U87 cells were treated
with RHL for 48 h, levels of Bax and Bim proteins were
significantly upregulated compared with the control group in a
dose-dependent manner. In contrast, compared with the control
group, the level of Bcl-2 protein in the cells treated with RHL was
significantly downregulated in a dose-dependent manner.
Furthermore, the level of cyclin D1 was decreased in the RHL group
in a dose-dependent manner. Cyclin D1 is a protein required for
progression through the G1 phase of the cell cycle. During the G1
phase, it is synthesized rapidly and accumulates in the nucleus,
and is degraded as the cell enters the S phase. Cyclin D1 is a
regulatory subunit of cyclin-dependent kinases CDK4 and CDK6. The
protein dimerizes with CDK4/6 to regulate G1/S phase transition and
entry into the S phase (41). It
can be concluded that RHL also inhibited cell proliferation by
blocking the G1/S phase of the cell cycle.
Our results showed that RHL is highly effective
against the growth of human glioma U87 xenografts in BALB/c nude
mice. In vitro, RHL induced apoptosis in human glioma U87
cells by decreasing Bcl-2 and increasing the expression of BAX and
Bim. The downregulation of the level of cyclin D was also involved
in the anticancer effects of RHL.
Acknowledgments
The present study was supported by grants from the
National Natural Science Foundation of China (81001439), and the
General Program of the Natural Science Foundation of Hebei Province
of China (H2012401030).
References
|
1
|
Gao J, Wang Z, Liu H, Wang L and Huang G:
Liposome encapsulated of temozolomide for the treatment of glioma
tumor: Preparation, characterization and evaluation. Drug Discov
Ther. 9:205–212. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kawaji H, Tokuyama T, Yamasaki T, Amano S,
Sakai N and Namba H: Interferon-β and temozolomide combination
therapy for temozolomide monotherapy-refractory malignant gliomas.
Mol Clin Oncol. 3:909–913. 2015.PubMed/NCBI
|
|
3
|
Yu Z, Xie G, Zhou G, Cheng Y, Zhang G, Yao
G, Chen Y, Li Y and Zhao G: NVP-BEZ235, a novel dual PI3K-mTOR
inhibitor displays anti-glioma activity and reduces chemoresistance
to temozolomide in human glioma cells. Cancer Lett. 367:58–68.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang X, Jia L, Jin X, Liu Q, Cao W, Gao X,
Yang M and Sun B: NF-κB inhibitor reverses temozolomide resistance
in human glioma TR/U251 cells. Oncol Lett. 9:2586–2590.
2015.PubMed/NCBI
|
|
5
|
Tian T, Li A, Lu H, Luo R, Zhang M and Li
Z: TAZ promotes temozolomide resistance by upregulating MCL-1 in
human glioma cells. Biochem Biophys Res Commun. 463:638–643. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou Q: BMS-536924, an ATP-competitive
IGF-1R/IR inhibitor, decreases viability and migration of
temozolomide-resistant glioma cells in vitro and suppresses tumor
growth in vivo. Onco Targets Ther. 8:689–697. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang Q, Lu G, Shen HM, Chung MC and Ong
CN: Anti-cancer properties of anthraquinones from rhubarb. Med Res
Rev. 27:609–630. 2007. View Article : Google Scholar
|
|
8
|
Tamura T, Yokoyama T and Ohmori K: Effects
of diacerein on indomethacin-induced gastric ulceration.
Pharmacology. 63:228–233. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhong XF, Huang GD, Luo T, Deng ZY and Hu
JN: Protective effect of rhein against oxidative stress-related
endothelial cell injury. Mol Med Rep. 5:1261–1266. 2012.PubMed/NCBI
|
|
10
|
Zhao YL, Zhou GD, Yang HB, Wang JB, Shan
LM, Li RS and Xiao XH: Rhein protects against acetaminophen-induced
hepatic and renal toxicity. Food Chem Toxicol. 49:1705–1710. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hu G, Liu J, Zhen YZ, Wei J, Qiao Y, Lin
YJ and Tu P: Rhein inhibits the expression of vascular cell
adhesion molecule 1 in human umbilical vein endothelial cells with
or without lipopolysaccharide stimulation. Am J Chin Med.
41:473–485. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cong XD, Ding MJ, Dai DZ, Wu Y, Zhang Y
and Dai Y: ER stress, p66shc, and p-Akt/Akt mediate
adjuvant-induced inflammation, which is blunted by argirein, a
supermolecule and rhein in rats. Inflammation. 35:1031–1040. 2012.
View Article : Google Scholar
|
|
13
|
Barnard DL, Huffman JH, Morris JL, Wood
SG, Hughes BG and Sidwell RW: Evaluation of the antiviral activity
of anthraquinones, anthrones and anthraquinone derivatives against
human cytomegalovirus. Antiviral Res. 17:63–77. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cyong J, Matsumoto T, Arakawa K, Kiyohara
H, Yamada H and Otsuka Y: Anti-bacteroides fragilis substance from
rhubarb. J Ethnopharmacol. 19:279–283. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Agarwal SK, Singh SS, Verma S and Kumar S:
Antifungal activity of anthraquinone derivatives from Rheum emodi.
J Ethnopharmacol. 72:43–46. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Singh B, Nadkarni JR, Vishwakarma RA,
Bharate SB, Nivsarkar M and Anandjiwala S: The hydroalcoholic
extract of Cassia alata (Linn.) leaves and its major compound rhein
exhibits antiallergic activity via mast cell stabilization and
lipoxygenase inhibition. J Ethnopharmacol. 141:469–473. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin ML, Chung JG, Lu YC, Yang CY and Chen
SS: Rhein inhibits invasion and migration of human nasopharyngeal
carcinoma cells in vitro by down-regulation of matrix
metalloproteinases-9 and vascular endothelial growth factor. Oral
Oncol. 45:531–537. 2009. View Article : Google Scholar
|
|
18
|
Fernand VE, Losso JN, Truax RE, Villar EE,
Bwambok DK, Fakayode SO, Lowry M and Warner IM: Rhein inhibits
angio-genesis and the viability of hormone-dependent and
-independent cancer cells under normoxic or hypoxic conditions in
vitro. Chem Biol Interact. 192:220–232. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen YY, Chiang SY, Lin JG, Ma YS, Liao
CL, Weng SW, Lai TY and Chung JG: Emodin, aloe-emodin and rhein
inhibit migration and invasion in human tongue cancer SCC-4 cells
through the inhibition of gene expression of matrix
metalloproteinase-9. Int J Oncol. 36:1113–1120. 2010.PubMed/NCBI
|
|
20
|
Shi P, Huang Z and Chen G: Rhein induces
apoptosis and cell cycle arrest in human hepatocellular carcinoma
BEL-7402 cells. Am J Chin Med. 36:805–813. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hsia TC, Yang JS, Chen GW, Chiu TH, Lu HF,
Yang MD, Yu FS, Liu KC, Lai KC, Lin CC, et al: The roles of
endoplasmic reticulum stress and Ca2+ on rhein-induced
apoptosis in A-549 human lung cancer cells. Anticancer Res.
29:309–318. 2009.PubMed/NCBI
|
|
22
|
Tsang SW and Bian ZX: Anti-fibrotic and
anti-tumorigenic effects of rhein, a natural anthraquinone
derivative, in mammalian stellate and carcinoma cells. Phytother
Res. 29:407–414. 2015. View
Article : Google Scholar
|
|
23
|
Huang HJ, Lin CC, Chou HC, Chen YW, Lin
ST, Lin YC, Lin DY, Lyu KW and Chan HL: Proteomic analysis of
rhein-induced cyt: ER stress mediates cell death in breast cancer
cells. Mol Biosyst. 10:3086–3100. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Du Q, Bian XL, Xu XL, Zhu B, Yu B and Zhai
Q: Role of mitochondrial permeability transition in human
hepatocellular carcinoma Hep-G2 cell death induced by rhein.
Fitoterapia. 91:68–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin YJ and Zhen YS: Rhein lysinate
suppresses the growth of breast cancer cells and potentiates the
inhibitory effect of Taxol in athymic mice. Anticancer Drugs.
20:65–72. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lin YJ, Zhen YZ, Shang BY and Zhen YS:
Rhein lysinate suppresses the growth of tumor cells and increases
the anti-tumor activity of Taxol in mice. Am J Chin Med.
37:923–931. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhen YZ, Lin YJ, Gao JL, Zhao YF and Xu
AJ: Rhein lysinate inhibits cell growth by modulating various
mitogen-activated protein kinases in cervical cancer cells. Oncol
Lett. 2:129–133. 2011.PubMed/NCBI
|
|
28
|
Lin YJ, Zhen YZ, Zhao YF, Wei J and Hu G:
Rhein lysinate induced S-phase arrest and increased the anti-tumor
activity of 5-FU in HeLa cells. Am J Chin Med. 39:817–825. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhen YZ, Hu G, Zhao YF, Yan F, Li R, Gao
JL and Lin YJ: Synergy of Taxol and rhein lysinate associated with
the down-regulation of ERK activation in lung carcinoma cells.
Oncol Lett. 6:525–528. 2013.PubMed/NCBI
|
|
30
|
Reardon DA, Galanis E, DeGroot JF,
Cloughesy TF, Wefel JS, Lamborn KR, Lassman AB, Gilbert MR, Sampson
JH, Wick W, et al: Clinical trial end points for high-grade glioma:
The evolving landscape. Neuro-oncol. 13:353–361. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chastagner P, Sudour H, Mriouah J,
Barberi-Heyob M, Bernier-Chastagner V and Pinel S: Preclinical
studies of pegylated- and non-pegylated liposomal forms of
doxorubicin as radiosensitizer on orthotopic high-grade glioma
xenografts. Pharm Res. 32:158–166. 2015. View Article : Google Scholar
|
|
32
|
Li XQ, Ouyang ZG, Zhang SH, Liu H, Shang
Y, Li Y and Zhen YS: Synergistic inhibition of angiogenesis and
glioma cell-induced angiogenesis by the combination of temozolomide
and enediyne antibiotic lidamycin. Cancer Biol Ther. 15:398–408.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang L and Zhao D: Liposomal
encapsulation enhances in vivo near infrared imaging of exposed
phosphatidylserine in a mouse glioma model. Molecules.
18:14613–14628. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Polunas M, Halladay A, Tjalkens RB,
Philbert MA, Lowndes H and Reuhl K: Role of oxidative stress and
the mitochondrial permeability transition in methylmercury
cytotoxicity. Neurotoxicology. 32:526–534. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li JP: Resveratrol caused apoptosis in
QGY-7701 cells. Eur Rev Med Pharmacol Sci. 19:3303–3308.
2015.PubMed/NCBI
|
|
36
|
Ma YX, Guo Z and Sun T: CGRP inhibits
norepinephrine induced apoptosis with restoration of Bcl-2/Bax in
cultured cardiomyocytes of rat. Neurosci Lett. 549:130–134. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zeng C, Ke Z, Song Y, Yao Y, Hu X, Zhang
M, Li H and Yin J: Annexin A3 is associated with a poor prognosis
in breast cancer and participates in the modulation of apoptosis in
vitro by affecting the Bcl-2/Bax balance. Exp Mol Pathol. 95:23–31.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Reed JC, Miyashita T, Takayama S, Wang HG,
Sato T, Krajewski S, Aimé-Sempé C, Bodrug S, Kitada S and Hanada M:
BCL-2 family proteins: Regulators of cell death involved in the
pathogenesis of cancer and resistance to therapy. J Cell Biochem.
60:23–32. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Labi V, Woess C, Tuzlak S, Erlacher M,
Bouillet P, Strasser A, Tzankov A and Villunger A: Deregulated cell
death and lymphocyte homeostasis cause premature lethality in mice
lacking the BH3-only proteins Bim and Bmf. Blood. 123:2652–2662.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View Article : Google Scholar
|
|
41
|
Baldin V, Lukas J, Marcote MJ, Pagano M
and Draetta G: Cyclin D1 is a nuclear protein required for cell
cycle progression in G1. Genes Dev. 7:812–821. 1993. View Article : Google Scholar : PubMed/NCBI
|